
Characteristics of Soil Structure and Greenhouse Gas Fluxes on Ten-Year Old Skid Trails with and without Black Alders (Alnus glutinosa (L.) Gaertn.)


Abstract
:1. Introduction
2. Materials and Methods
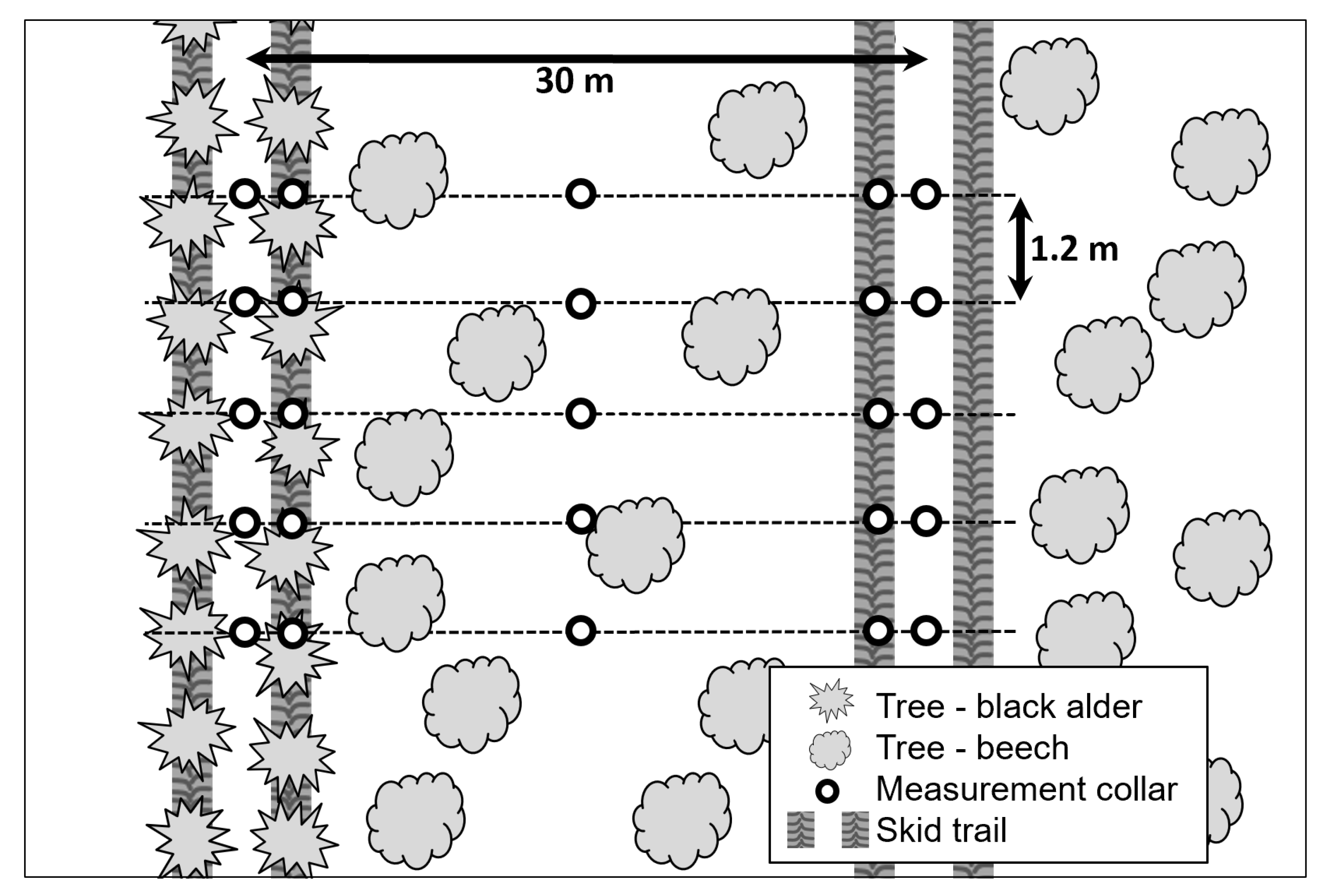
2.1. Site Description
2.2. Filed Measurements
2.3. Soil Sampling and Laboratory Anayses
2.4. Statistical Anayses
3. Results
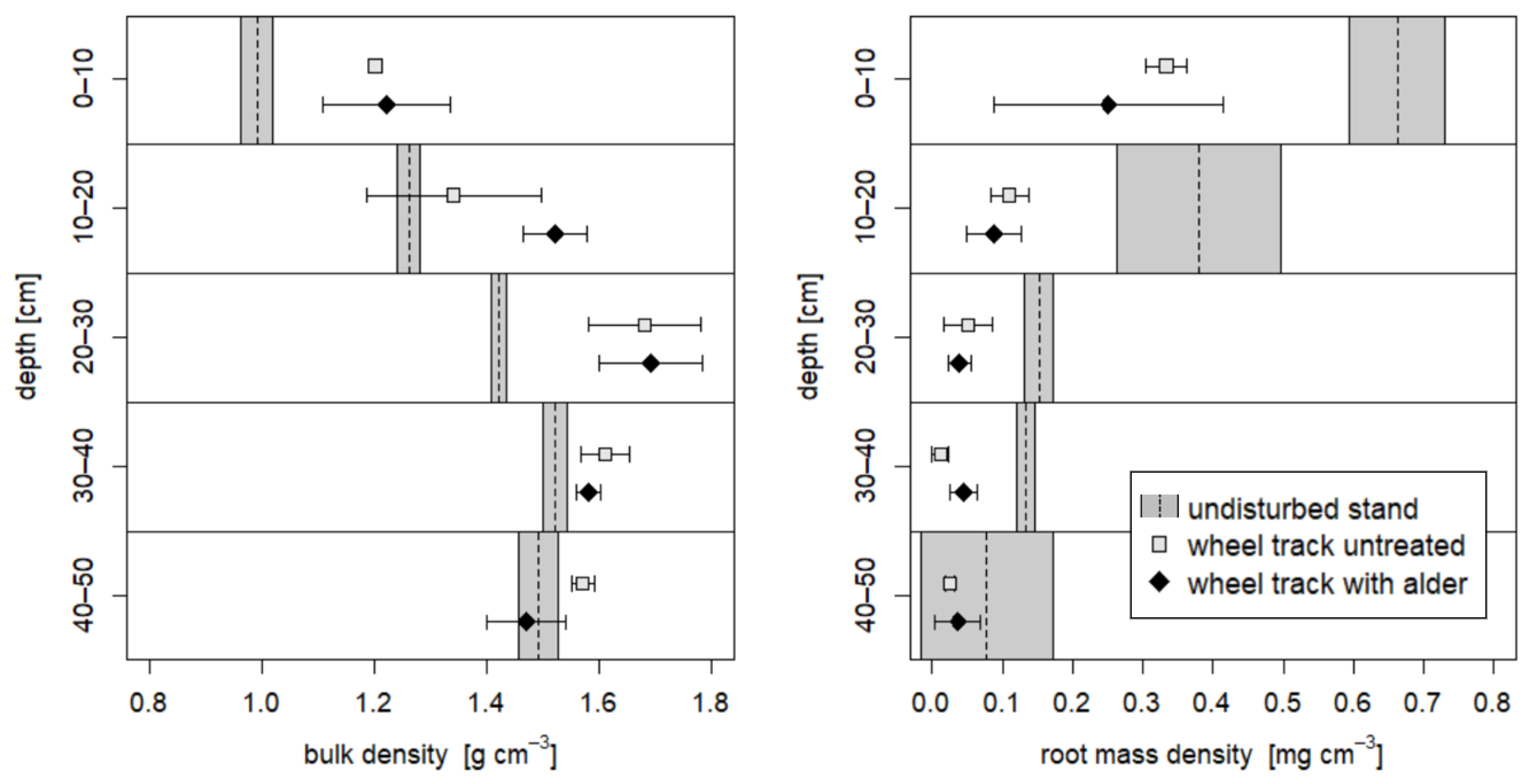
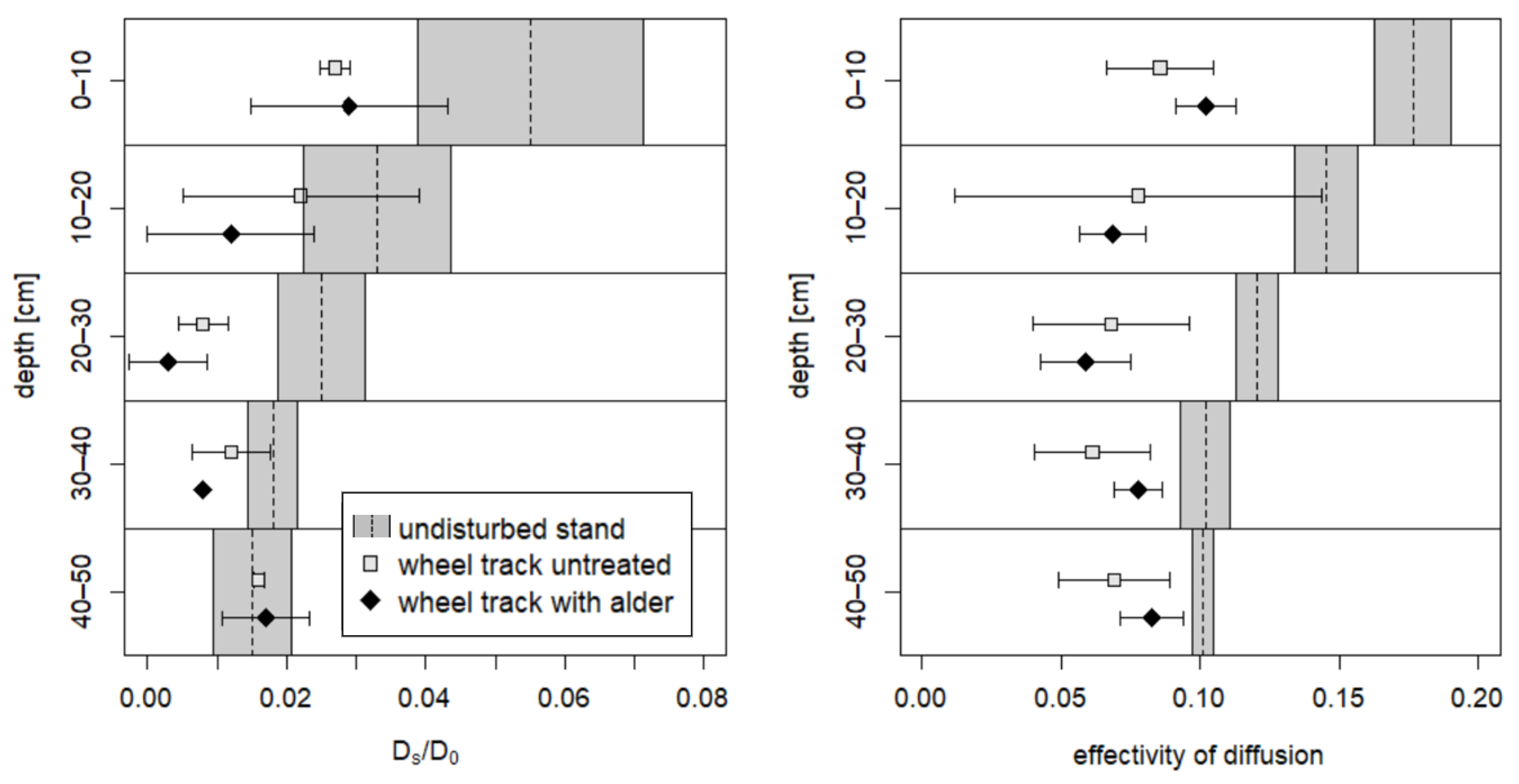
3.1. Soil Physical Parameters
3.2. Comparison of Soil Physical Parameters in 2008 and 2018
3.3. Soil Carbon and Nitrogen Contents
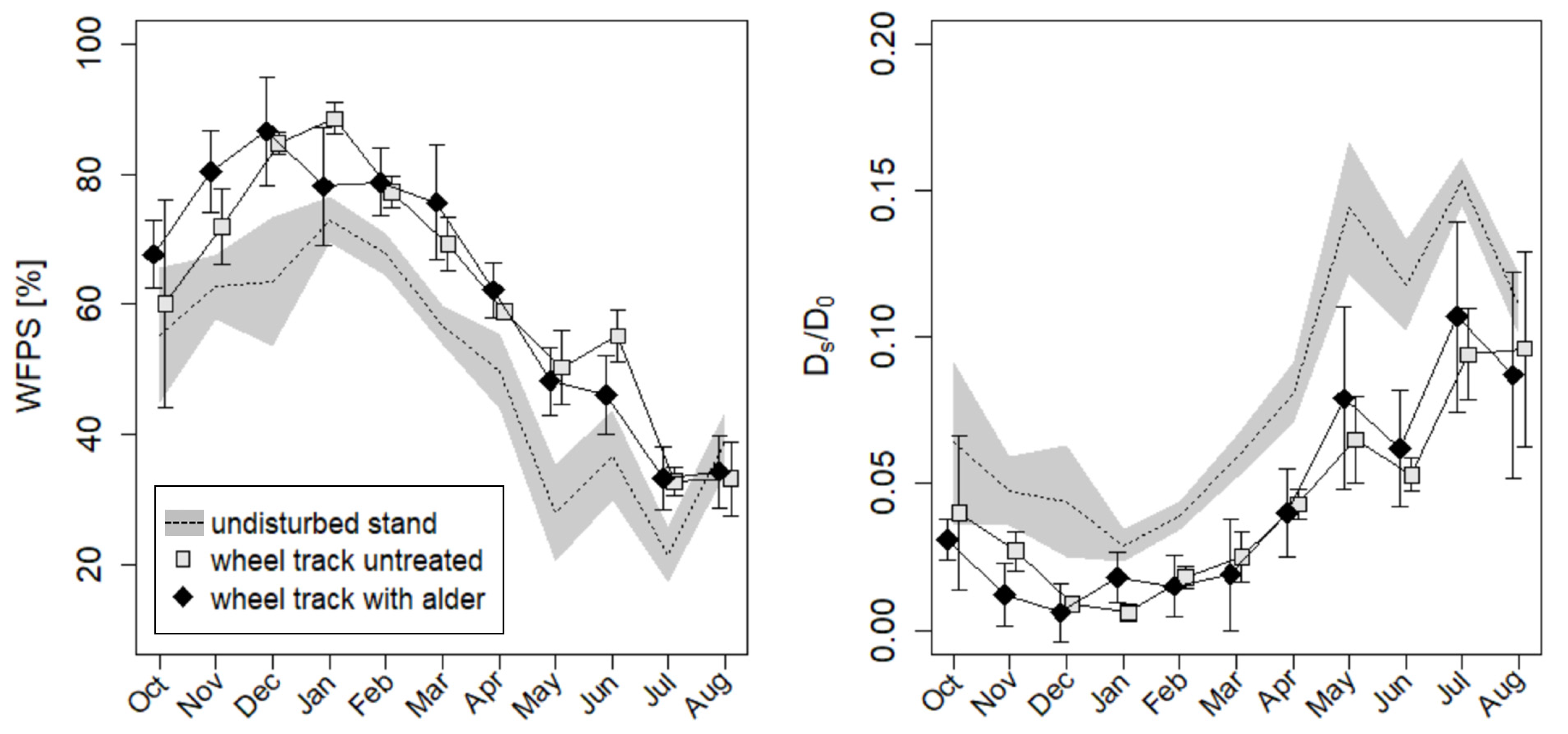
3.4. Environmental Conditions during Greenhous Gas Measurements
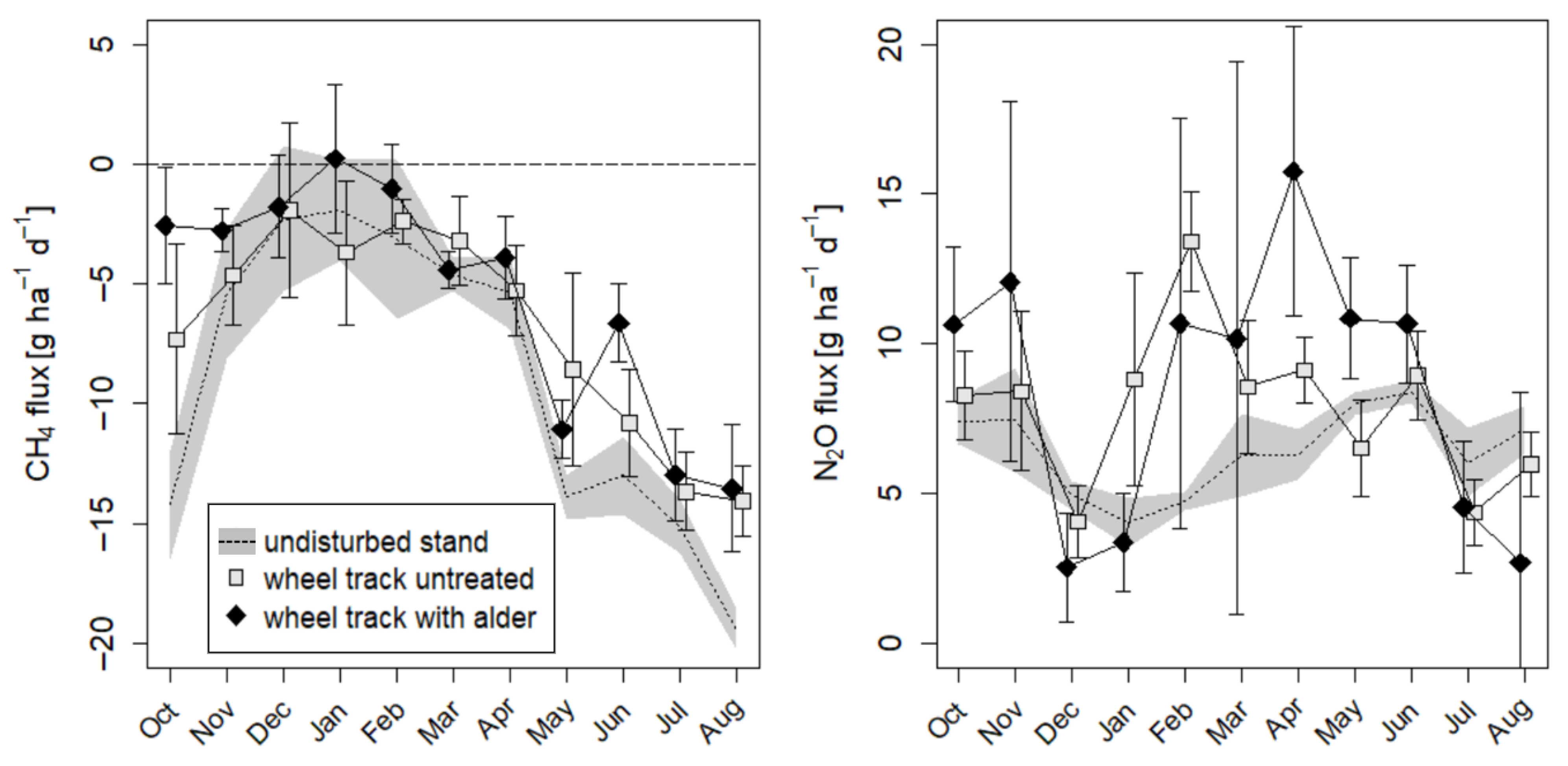
3.5. Greenhouse Gas Fluxes
4. Discussion
4.1. Soil Recovery on the Skid Trail without Treatment
4.2. Soil Recovery on the Skid Trail Planted with Black Alders
4.3. Greenhouse Gas Fluxes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riggert, R.; Fleige, F.; Kietz, B.; Gaertig, T.; Horn, R. Stress distribution under forestry machinery and consequences for soil stability. Soil Sci. Soc. Am. J. 2016, 80, 38–47. [Google Scholar] [CrossRef]
- Horn, R.; Vossbrink, J.; Peth, S.; Becker, S. Impact of modern forest vehicles on soil physical properties. For. Ecol. Manag. 2007, 248, 56–63. [Google Scholar] [CrossRef]
- Gebauer, R.; Martinková, M. Effects of pressure on the root systems of Norway spruce plants (Picea abies [L.] Karst.). J. For. Sci. 2005, 51, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Ampoorter, E.; Goris, R.; Cornelis, W.M.; Verheyen, K. Impact of mechanized logging on compaction status of sandy forest soils. For. Ecol. Manag. 2007, 241, 162–174. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. Production, Oxidation, Emission and Consumption of Methane by Soils: A Review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Dobbie, K.E.; Smith, K.A. Nitrous Oxide Emission Factors for Agricultural Soils in Great Britain: The Impact of Soil Water-filled Pore Space and Other Controlling Variables. Glob. Change Biol. 2003, 9, 204–218. [Google Scholar] [CrossRef]
- Ampoorter, E.; De Schrijver, A.; De Frenne, P.; Hermy, M.; Verheyen, K. Experimental assessment of ecological restoration options for compacted forest soils. Ecol. Eng. 2011, 37, 1734–1746. [Google Scholar] [CrossRef]
- von Wilpert, K. Forest Soil—What’ s Their Peculiarity? Soil Syst. 2022, 6, 5. [Google Scholar] [CrossRef]
- von Wilpert, K.; Schäffer, J. Ecological effects of soil compaction and initial recovery dynamics: A preliminary study. Eur. J. For. Res. 2006, 125, 129–138. [Google Scholar] [CrossRef]
- Garland, J.J. Designated Skid Trails Minimize Soil Compaction. In The Woodland Workbook; Oregon State University Extension Service: Corvalis, OR, USA, 1983; pp. 1–7. [Google Scholar]
- Angers, D.A.; Caron, J. Plant-induced changes in soil structure: Processes and feedbacks. Biogeochemistry 1998, 42, 55–72. [Google Scholar] [CrossRef]
- Carminati, A.; Vetterlein, D.; Weller, U.; Vogel, H.; Oswald, S.E. When roots lose contact. Vadose Zone J. 2009, 8, 805–809. [Google Scholar] [CrossRef]
- Meyer, C.; Lüscher, P.; Schulin, R. Recovery of forest soil from compaction in skid tracks planted with black alder (Alnus glutinosa (L.) Gaertn.). Soil Tillage Res. 2014, 143, 7–16. [Google Scholar] [CrossRef]
- Meyer, C.; Lüscher, P.; Schulin, R. Enhancing the regeneration of compacted forest soils by planting black alder in skid lane tracks. Eur. J. For. Res. 2014, 133, 453–465. [Google Scholar] [CrossRef]
- Flores Fernández, J.L.; Hartmann, P.; Schäffer, J.; Puhlmann, H.; von Wilpert, K. Initial recovery of compacted soil—planting and technical treatments decrease CO2concentrations in soil and promote root growth. Ann. For. Sci. 2017, 74, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Vergani, C.; Graf, F. Soil permeability, aggregate stability and root growth: A pot experiment from a soil bioengineering perspective. Ecohydrology 2016, 9, 830–842. [Google Scholar] [CrossRef]
- Ebeling, C.; Lang, F.; Gaertig, T. Structural recovery in three selected forest soils after compaction by forest machines in Lower Saxony, Germany. For. Ecol. Manag. 2016, 359, 74–82. [Google Scholar] [CrossRef]
- Warlo, H.; von Wilpert, K.; Lang, F.; Schack-Kirchner, H. Black Alder (Alnus glutinosa (L.) Gaertn.) on Compacted Skid Trails: A Trade-off between Greenhouse Gas Fluxes and Soil Structure Recovery? Forests 2019, 10, 726. [Google Scholar] [CrossRef] [Green Version]
- Kätzel, R. Zum physiologischen Anpassungspotenzial der Schwarz-Erle (Alnus glutinosa [L.] GAERTN.). Eberswalder Forstl Schr. 2003, 17, 39–46. [Google Scholar]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A review of the characteristics of black alder (Alnus glutinosa (L.) Gaertn.) and their implications for silvicultural practices. Forestry 2010, 83, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Klingensmith, K.M.; Van Cleve, K. Denitrification and nitrogen fixation in floodplain successional soils along the Tanana River, interior Alaska. Can. J. For. Res. 1993, 23, 956–963. [Google Scholar] [CrossRef]
- Mogge, B.; Kaiser, E.A.; Munch, J.C. Nitrous oxide emissions and denitrification N-losses from forest soils in the bornhoved lake region (Northern Germany). Soil Biol. Biochem. 1998, 30, 703–710. [Google Scholar] [CrossRef]
- Frey, B.; Niklaus, P.A.; Kremer, J.; Lüscher, P.; Zimmermann, S. Heavy-machinery traffic impacts methane emissions as well as methanogen abundance and community structure in oxic forest soils. Appl. Environ. Microbiol. 2011, 77, 6060–6068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Rome World Soil Resour; Rep. No 106; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2015; p. 203.
- Lüscher, P.; Sciacca, S.; Thees, O. Bestrebungen zur Verbesserung des Bodenschutzes in der Schweiz. LWF Aktuell 2008, 67, 19–21. [Google Scholar]
- Schäffer, J.; von Wilpert, K.; Kublin, E. Analysis of fine rooting below skid trails using linear and generalized additive models. Can. J. For. Res. 2009, 39, 2047–2058. [Google Scholar] [CrossRef]
- Teepe, R.; Brumme, R.; Beese, F.; Ludwig, B. Nitrous Oxide Emission and Methane Consumption Following Compaction of Forest Soils. Soil Sci. Soc. Am. J. 2004, 68, 605. [Google Scholar] [CrossRef]
- Maier, M.; Longdoz, B.; Laemmel, T.; Schack-Kirchner, H.; Lang, F. 2D profiles of CO2, CH4, N2O and gas diffusivity in a well aerated soil: Measurement and Finite Element Modeling. Agric. For. Meteorol. 2017, 247, 21–33. [Google Scholar] [CrossRef]
- Muukkonen, P.; Häkkinen, M.; Mäkipää, R. Spatial variation in soil carbon in the organic layer of managed boreal forest soil-implications for sampling design. Environ. Monit. Assess. 2009, 158, 67–76. [Google Scholar] [CrossRef]
- Hartmann, M.; Niklaus, P.A.; Zimmermann, S.; Schmutz, S.; Kremer, J.; Abarenkov, K.; Lüscher, P.; Widmer, F.; Frey, B. Resistance and resilience of the forest soil microbiome to logging-associated compaction. ISME J. 2014, 8, 226–244. [Google Scholar] [CrossRef] [PubMed]
- Loftfield, N.; Flessa, H.; Augustin, J.; Beese, F. Automated Gas Chromatographic System for Rapid Analysis of the Atmospheric Trace Gases Methane, Carbon Dioxide, and Nitrous Oxide. J. Environ. Qual. 1997, 26, 560–564. [Google Scholar] [CrossRef]
- Hutchinson, G.L.; Livingston, G.P. Soil-Atmosphere Gas Exchange. In Methods of Soil Analysis: Part 4 Physical Methods; Soil Science Society of America Book Ser. 5: Madison, WI, USA, 2002; pp. 1159–1182. [Google Scholar]
- Maechler, M.; Rousseeuw, P.; Croux, C.; Todorov, V.; Ruckstuhl, A.; Salibian-Barrera, M.; Verbeke, T.; Koller, M.; Conceicao, E.L.; di Palma, M.A. robustbase: Basic Robust Statistics; R Package Version 0.93-7; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Frey, B.; Kremer, J.; Rüdt, A.; Sciacca, S.; Matthies, D.; Lüscher, P. Compaction of forest soils with heavy logging machinery affects soil bacterial community structure. Eur. J. Soil Biol. 2009, 45, 312–320. [Google Scholar] [CrossRef]
- Hartge, K.H.; Horn, R. Die Physikalische Untersuchung von Böden, 3rd ed.; Ferdinand Enke Verlag: Stuttgart, Germany, 2011. [Google Scholar]
- Kühne, A.; Schack-Kirchner, H.; Hildebrand, E.E. Gas diffusivity in soils compared to ideal isotropic porous media. J. Plant Nutr. Soil Sci. 2012, 175, 34–45. [Google Scholar] [CrossRef]
- Schack-Kirchner, H.; Gaertig, T.; von Wilpert, K.; Hildebrand, E.E. A modified McIntyre and Phillip approach to measure top-soil gas diffusivity in-situ. J. Plant Nutr. Soil Sci. 2001, 164, 253–258. [Google Scholar] [CrossRef]
- Perdok, U.D.; Kroesbergen, B.; Hoogmoed, W.B. Possibilities for modelling the effect of compression on mechanical and physical properties of various Dutch soil types. Soil Tillage Res. 2002, 65, 61–75. [Google Scholar] [CrossRef]
- Dinno, A. dunn.test: Dunn’s Test of Multiple Comparisons Using Rank Sums; R Package Version 1.3.5; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- McGill, R.; Tukey, J.W.; Larsen, W.A. Variation of box plots. Am. Stat. 1978, 32, 12–16. [Google Scholar]
- Ampoorter, E.; Van Nevel, L.; De Vos, B.; Hermy, M.; Verheyen, K. Assessing the effects of initial soil characteristics, machine mass and traffic intensity on forest soil compaction. For. Ecol. Manag. 2010, 260, 1664–1676. [Google Scholar] [CrossRef] [Green Version]
- Schäffer, J.; Buberl, H.; von Wilpert, K. Deformation damages in forest topsoils—An assessment based on Level-I soil monitoring data from Baden-Württemberg (SW Germany). J. Plant Nutr. Soil Sci. 2012, 175, 24–33. [Google Scholar] [CrossRef]
- Qi, B.; Marshall, J.D.; Mattson, K.G. High soil carbon dioxide concentrations inhibit root respiration of Douglas fir. New Phytol. 1994, 128, 435–442. [Google Scholar] [CrossRef]
- Hildebrand, E.E. Der Einfluß der Bodenverdichtung auf die Bodenfunktionen im forstlichen Standort. Forstwiss. Cent. 1983, 102, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Hodgkinson, L.; Jin, K.; Watts, C.W.; Ashton, R.W.; Shen, J.; Ren, T.; Dodd, I.C.; Binley, A.; Phillips, A.L.; et al. Deep roots and soil structure. Plant Cell Environ. 2016, 39, 1662–1668. [Google Scholar] [CrossRef] [Green Version]
- Dexter, A.R. Compression of soil around roots. Plant Soil 1987, 97, 401–406. [Google Scholar] [CrossRef]
- Sommer, M.; Ehrmann, O.; Friedel, J.K.; Martin, K.; Vollmer, T.; Turian, G. Böden als Lebensraum für Organismen-Regenwürmer, Gehäuseschnecken und Bodenmikroorganismen in Wäldern Baden-Württembergs. Hohenh. Bodenkd. Hefte 2002, 63, 98–100. [Google Scholar]
- Capowiez, Y.; Cadoux, S.; Bouchand, P.; Roger-Estrade, J.; Richard, G.; Boizard, H. Experimental evidence for the role of earthworms in compacted soil regeneration based on field observations and results from a semi-field experiment. Soil Biol. Biochem. 2009, 41, 711–717. [Google Scholar] [CrossRef]
- Ponder, F.; Li, F.; Jordan, D.; Berry, E.C. Assessing the impact of Diplocardia ornata on physical and chemical properties of compacted forest soil in microcosms. Biol. Fertil. Soils. 2000, 32, 166–172. [Google Scholar] [CrossRef]
- Larink, O.; Werner, D.; Langmaack, M.; Schrader, S. Regeneration of compacted soil aggregates by earthworm activity. Biol. Fertil. Soils. 2001, 33, 395–401. [Google Scholar]
- Greacen, E.L.; Sands, R. Compaction of Forest Soils. A Review. Aust. J. Soil Res. 1980, 18, 163–189. [Google Scholar] [CrossRef]
- Lavelle, P.; Barot, S.; Blouin, M.; Decaens, T.; Jimenez, J.J.; Jouquet, P. Earthworms as key factor in self-organized soil systems. In Ecosystem Engeneers: Plants to Protists; Cuddington, K., Byers, J.E., Wilson, W.G., Hastings, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 77–106. [Google Scholar]
- Fisher, R.F.; Binkley, D. Ecology and Management of Forest Soils, 3rd ed.; Wiley: New York, NY, USA, 2000. [Google Scholar]
- Othman, M.A.; Benson, C.H. Effect of freeze–thaw on the hydraulic conductivity and morphology of compacted clay. Can. Geotech J. 1993, 30, 236–246. [Google Scholar] [CrossRef]
- Uri, V.; Aosaar, J.; Varik, M.; Becker, H.; Ligi, K.; Padari, A.; Kanal, A.; Löhmus, K. The dynamics of biomass production, carbon and nitrogen accumulation in grey alder (Alnus incana (L.) Moench) chronosequence stands in Estonia. For. Ecol. Manag. 2014, 327, 106–117. [Google Scholar] [CrossRef]
- Haynes, R.J.; Naidu, R. Influence of lime, fertilizer and manure applications on soil organic matter content and soil physical conditions: A review. Nutr. Cycl. Agroecosyst. 1998, 51, 123–137. [Google Scholar] [CrossRef]
- Epron, D.; Plain, C.; Ndiaye, F.K.; Bonnaud, P.; Pasquier, C.; Ranger, J. Effects of compaction by heavy machine traffic on soil fluxes of methane and carbon dioxide in a temperate broadleaved forest. For. Ecol. Manag. 2016, 382, 1–9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Depth * | Beech Stand | Skid Trail (Untreated) | Skid Trail (Alder) | |||
|---|---|---|---|---|---|---|
| 2008 | 2018 | 2008 | 2018 | 2018 | ||
| Bulk density [g m−3] | 1 | 1.23 ± 0.04 a | 0.99 ± 0.03 b | 1.52 ± 0.04 a | 1.23 ± 0.01 b | 1.22 ± 0.11 b |
| 2 | 1.49 ± 0.03 a | 1.42 ± 0.01 b | 1.56 ± 0.04 a | 1.68 ± 0.10 a | 1.69 ± 0.10 a | |
| Total porosity [%] | 1 | 50.1 ± 0.7 a | 65.2 ± 1.8 b | 43.3 ± 2.0 a | 57.1 ± 3.0 b | 57.9 ± 8.7 b |
| 2 | 42.4 ± 1.0 a | 47.7 ± 0.1 b | 39.8 ± 0.3 a | 40.9 ± 4.2 a | 37.7 ± 2.0 a | |
| Macro porosity [%] | 1 | 13.3 ± 2.4 a | 15.8 ± 1.6 a | 3.3 ± 0.6 a | 19.8 ± 3.1 b | 18.3 ± 8.7 b |
| 2 | 9.8 ± 1.6 a | 8.5 ± 0.5 a | 4.2 ± 0.5 a | 11.1 ± 3.3 b | 3.6 ± 7.7 b | |
| Beech Stand | Skid Trail Untreated | Skid Trail Alder | |
|---|---|---|---|
| CH4 (kg ha−1 yr−1) | −2.53 ± 0.79 a | −2.14 ± 0.62 ab | −1.67 ± 0.56 b |
| N2O (kg ha−1 yr−1) | 2.51 ± 0.23 a | 2.49 ± 0.35 a | 3.23 ± 0.70 b |
| CO2 (Mg ha−1 yr−1) | 26.1 ± 5.5 a | 26.7 ± 5.3 a | 28.0 ± 6.5 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warlo, H.; Zimmermann, S.; Lang, F.; Schack-Kirchner, H. Characteristics of Soil Structure and Greenhouse Gas Fluxes on Ten-Year Old Skid Trails with and without Black Alders (Alnus glutinosa (L.) Gaertn.). Soil Syst. 2022, 6, 43. https://doi.org/10.3390/soilsystems6020043
Warlo H, Zimmermann S, Lang F, Schack-Kirchner H. Characteristics of Soil Structure and Greenhouse Gas Fluxes on Ten-Year Old Skid Trails with and without Black Alders (Alnus glutinosa (L.) Gaertn.). Soil Systems. 2022; 6(2):43. https://doi.org/10.3390/soilsystems6020043
Chicago/Turabian StyleWarlo, Hannes, Stephan Zimmermann, Friederike Lang, and Helmer Schack-Kirchner. 2022. "Characteristics of Soil Structure and Greenhouse Gas Fluxes on Ten-Year Old Skid Trails with and without Black Alders (Alnus glutinosa (L.) Gaertn.)" Soil Systems 6, no. 2: 43. https://doi.org/10.3390/soilsystems6020043