
The “True Colours” of Golden Loaches (Teleostei: Cobitidae)
by
 , , , , , and
, , , , , and
Jakub Fedorčák
1,* ,
,
Radek Šanda
2,
Tihomir Stefanov
3,
Jan Mendel
4,
Michal Nowak
5,
Peter Križek
6,
Anabel Perdices
7,
Jasna Vukić
8 and
Ján Koščo
1 1
Department of Ecology, Faculty of Humanities and Natural Sciences, University of Presov, Ul. 17. Novembra 1, 080 16 Prešov, Slovakia
2
Department of Zoology, National Museum, Václavské Náměstí 68, 115 79 Prague, Czech Republic
3
Vertebrates Department, National Museum of Natural History, Bulgarian Academy of Science, 1 Tsar Osvoboditel Blvd, 1000 Sofia, Bulgaria
4
Institute of Vertebrate Biology, Czech Academy of Sciences, Květná 8, 603 65 Brno, Czech Republic
5
Mazanów Fish Farm, Mazanów 8A, 24-340 Jozefow nad Wislą, Poland
6
Slovak Angling Association—The Board, Andreja Kmet’a 20, 010 55 Žilina, Slovakia
7
Department of Biodiversity and Evolutionary Biology, Museo Nacional de Ciencias Naturales, José Gutiérrez Abascal 2, 28006 Madrid, Spain
8
Department of Ecology, Faculty of Science, Charles University, Viničná 7, 128 44 Prague 2, Czech Republic
*
Author to whom correspondence should be addressed.
Fishes 2023, 8(2), 119; https://doi.org/10.3390/fishes8020119
Submission received: 30 December 2022
/
Revised: 16 February 2023
/
Accepted: 17 February 2023
/
Published: 20 February 2023
(This article belongs to the Section Biology and Ecology)
Abstract
:Loaches of the genus Sabanejewia are model organisms for many ichthyologic studies focusing on morphological and colour variation. The bright lateral spots—a part of the mimicry of this fish—cause a taxonomic puzzle. Common practice is that morphometric and molecular studies lack habitat assessments of species originating from different environments. In this study, we analysed body shape and colour variation within two species of Sabanejewia in order to reveal whether they are more affected by phylogeny or environmental conditions. The environmental parameters were obtained using satellite mapping tools, body shape using geometric morphometry, and the molecular identification of specimens using an analysis of mitochondrial DNA (sequence of the cytochrome b gene). Generalized linear mixed models were used for the analysis of lateral spot numbers, and principal components analysis to describe the changes in landmark position defined by the permutation tests. Results show a significant impact of the locality elevation and specimen’s sex on the change of the body shape. We identified significant differences in the number of lateral spots, which are considered mimicry variation across an elevation gradient. Morphological differences of Sabanejewia spp. vary with ecological factors along the river continuum.
Keywords:
ecology; cryptic colouration; habitat; sequencing; museum collections; ecomorphotype; body shape; fish complex; Danube; Balkan1. Introduction
Species delimitation is a key task of systematic biology since species is the basic unit for evolutionary or conservation studies. Most freshwater fish species are restricted to a few river basins, but some, e.g., species of the genus Sabanejewia, depart from this general rule. The genus Sabanejewia was described based on the sexual dimorphism of males [1] and several studies have analysed their external characters [2,3,4,5,6]. The genus Sabanejewia includes ten species, distributed from the Aral Sea to Italy and from Asia Minor to the Baltic Sea [7,8,9,10,11], while the validity of several other nominal Sabanejewia species is still unclear [11], the existence of unknown species is possible [12]. From the Danube River basin and Balkan watersheds, the “Danube–Balkan (DB) complex” [13] of sympatric Sabanejewia lineages have been described. Within sympatric species, a morphological cline (number of spots, body height, length of caudal peduncle, etc.) reflecting the character of the habitat was observed [3]. Molecular studies on the DB complex have established six mitochondrial lineages [13]. Morphological studies [14,15] of some individuals of the DB complex showed, however, that their morphological identification does not reflect their genetic identity. Although the morphological cline in the Danube lineages was related to their habitat [3], in most recent taxonomic studies, the influence of habitat has not been considered. Recently, Križek et al. [15] have confirmed the impact of the microhabitat parameters on morphological characters unrelated to the genetic identity of analysed specimens.
In this study, we evaluate the influence of environmental parameters and sex on the body shape exhibited by two Sabanejewia mitochondrial lineages from the DB complex. For the expansion of knowledge within the target genus, we analysed the most widespread lineages of the DB complex: the Balkan golden loach (S. balcanica Karaman, 1922) and the Bulgarian golden loach (S. bulgarica Drensky, 1928), including samples from type localities of both species and specimens from the adjacent river basins.
2. Materials and Methods
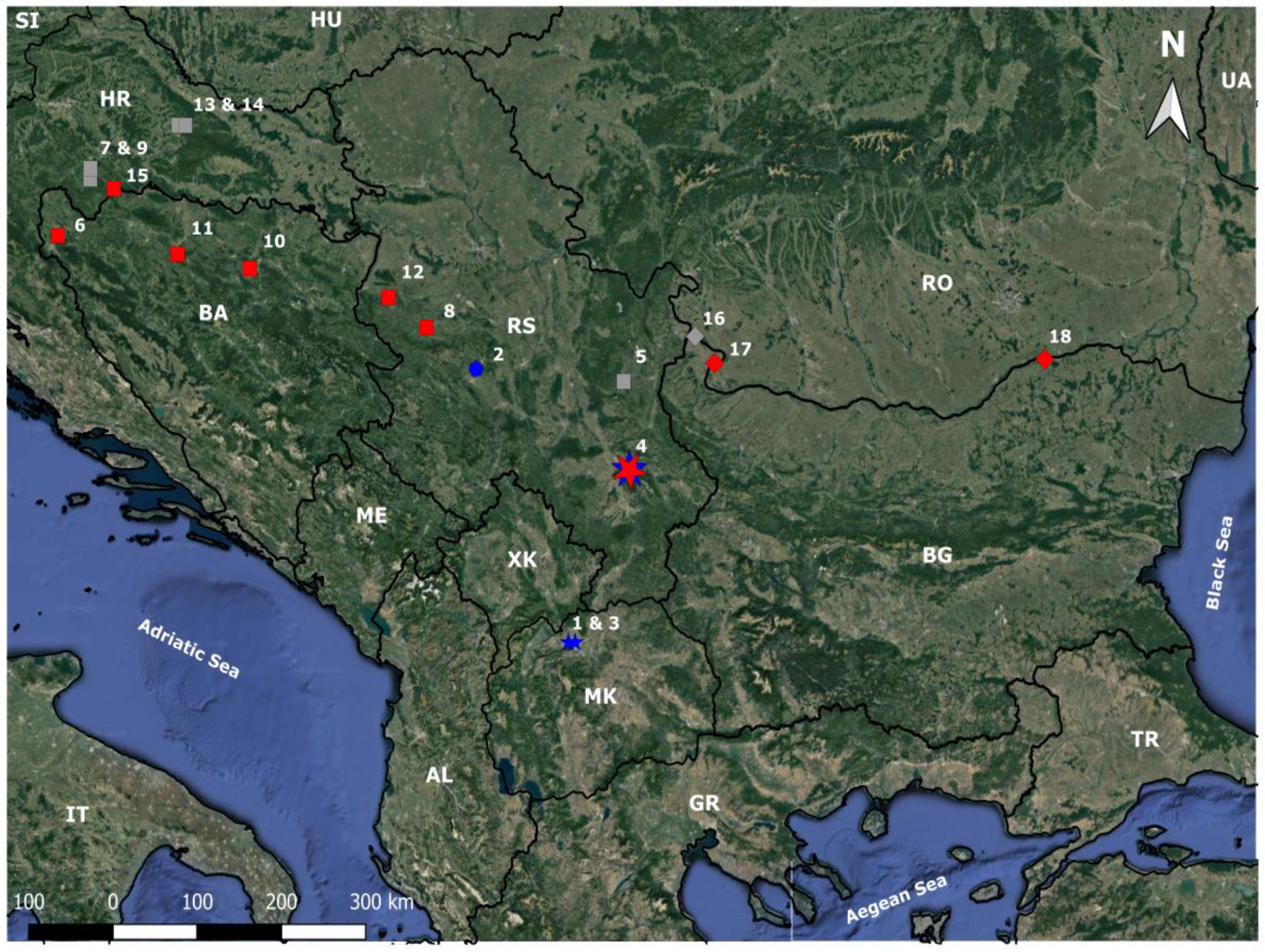
Samples: Individuals of Sabanejewia balcania and S. bulgarica analysed in this study are preserved in the ichthyological collections of the National Museum in Prague (CZ), the National Museum of Natural History in Sofia (BG) and the Department of Ecology of the University of Prešov (SK) (collection numbers listed in Table S1). All examined individuals were preserved by the same procedure (fixed in 4% formaldehyde solution and later transferred to 70% ethanol for permanent storage). The data set (133 specimens) was sex and length (SL) balanced (the length of both sexes did not differ statistically; χ2 = 0.03, d.f. = 1, p > 0.85) and length balanced between the species (χ2 = 0.13, d.f. = 1, p > 0.72). Out of 133 specimens, we included 111 specimens (24 S. balcanica and 87 S. bulgarica) in the morphometric analysis and 54 on the cyt b analysis (selected specimens were analysed morpologically as well as genetically, Table S1). Analysed specimens originated from the Danube drainage (Sava R., Danube R., Crni Timok R., Great Morava R.—Black Sea drainage) and from the Vardar drainage (Aegean Sea drainage) (Figure 1).
The localities represent different habitats at different elevations (Table S1). In addition, we analysed ten specimens of the S. bulgarica from the type locality (Danube River in Vidin) and twelve specimens of S. balcanica from the Vardar River catchment (Lepenac R., Treska R.) in Skopje, which are considered part of the type locality [16]. We also analysed specimens of NM Prague (Table S1) representing samples genetically characterised by Marešová et al. [17].
Morphometry: We analysed 11 homologous landmarks defined according to Mousavi-Sabet et al. [18] (Figure 2—excluding the center of the eye) on 111 specimens (Table S1—Geomorph).
Photographs of all specimens were taken with a fixed camera on a tripod, setting the height and standard lighting to minimise shooting errors. Standard length (SL; mm) was measured in TPS with graph paper as the calibration scale. The fish samples were pinned on a hard cover to avoid small body deformities. For landmark analysis, we used TPS landmarks based on Rohlf’s [19] procedure. Procrustes superimposition of landmarks, which eliminates the effect of different size, position and rotation of compared objects, was performed with the geomorph package [20] in the R statistical environment [21], according to Park et al. [22]. We checked outliers of transformed landmarks using the plot outliers function. Lateral spots were counted on the left side of the body.
Statistical analysis: All analyses were performed in the R statistical environment with a threshold for statistical significance α = 0.05. For data evaluation, we used several statistical approaches: (1) We assessed the permutational multivariate analysis of variance (PERMANOVA, Euclid., 9999 perm.) using the function adonis, according to Anderson [20], to define differences in landmark position depending on the explanatory variable (elevation, slope, sex); (2) Principal components analysis (PCA) [20] was performed to plot results of the PERMANOVA and deformation grids; (3) Comparison of the morphological disparity (within/between localities) was done by the function morphol.disparity of the geomorph package [23] under 999 iterations of the permutation test. The probabilities of the posterior population membership of individuals obtained by all grouping methods were converted to between-individual Euclidean distances. We also performed pairwise comparisons of these distance matrices using the Mantel test—ade4 package [24]. (4) For the evaluation of the lateral spot numbers of 111 specimens, we fitted generalised linear mixed models (GLMMs) with the response variable (number of lateral spots) and explanatory variables (elevation, slope, SL, sex, double interactions). Mixed models were fitted with the random factor of locality for correction of the spatial pseudo-replication between specimens from the same locality [25]. The GLMM models were submitted to ANOVA type II (testing the main effect after the other main effects) and III (in the case of significant interaction) [26]. The minimum Akaike information criterion (AIC) was applied for multiple model comparisons and the best model fit (quality/size).
Environmental parameters: We obtained environmental values (river bottom slope and the elevation) of the sampling localities using Google Earth Pro (ver.7.3.2.) according to the GPS locations of the voucher material. For a more accurate estimate of the slope of locality, we calculated the average river slope from 300 m of river transects up to the GPS coordinates.
Molecular analysis: A total of 54 sequences/49 haplotypes (Table S1) of the complete mitochondrial cytochrome b (cyt b; 1140 bp) of Sabanejewia spp. of the lineages S. bulgarica from the Danube River basin ([15,17], this work Table S1) and S. balcanica from the Vardar River and Danube River basin [17], together with sequences of the other four lineages of the DB complex ([13], this work Table S1 (OK032158-63)) were used for the construction of a consensus tree. We constructed the consensus tree in PopArt software [27]. The sequence of S. romanica (GenBank accession no. AF499196) was used as the outgroup for phylogenetic reconstruction. To avoid potential misidentification based on morphology, the new Sabanejewia specimens were not identified to the species level.
For the newly added sequences (Table S1), the total genomic DNA was extracted from a piece of fin tissue using a commercial kit (GT300, Geneaid). The sequences were deposited in the GenBank database (under Accession Nos. OK032158-63). We amplified the cyt b gene by PCR with the primer pair GluDG.L [28] and H16460 [29]. We performed 25-µL PCR reactions with the PPP master kit mix (Top-Bio) in a Mastercycler Pro (Eppendorf). The PCR conditions were: one initial denaturation cycle at 94 °C for 1 min, followed by 30 cycles of denaturation at 94 °C for 15 s, annealing at 60 °C for 15 s, extension at 72 °C for 1 min and a final extension at 72 °C for 7 min. The amplicons were visualised in 1.7% agarose gel by electrophoresis. The PCR products were purified with the DNA Clean & Concentrator-5 Kit (Zymo Research) and the complete cyt b (1140 base pairs) gene was sequenced at Macrogen Europe with the same PCR primers. All PCR amplicons were sequenced in both directions to ensure high quality reads. The DNA sequences were edited and aligned using the SeqMan module within Lasergene 15.0 (DNASTAR Inc., Madison, WI, USA) and also checked manually. Furthermore, as a final quality control, the cyt b sequences were translated to verify that they were free of stop codons, frame-shifts and gaps. A detailed list of all studied taxa, their haplotype classification relative to sampling sites, haplotype frequencies and GenBank accession numbers are shown in Table S1.
For the phylogenetic reconstructions, the genetic dataset was analysed by Bayesian inference (BI) using MrBayes 3.1.2 [30], the maximum likelihood (ML) method using PhyML [31] and the neighbour-joining algorithm (NJ) using PAUP* 4.0B.10 [32]. The best-fit model of molecular evolution was determined for the mitochondrial dataset using the Akaike Criterion (AIC) in jModeltest 2.1.4 [33]. MrBayes was run with six substitution types (nst = 6) and considered gamma-distributed rate variation and the proportion of invariable positions (GTR + G + I). For BI, we ran four simultaneous Monte Carlo Markov chains for 2 million generations, with a sampling frequency every 100 generations. The first 5000 trees were excluded as the burn-in. The remaining trees were used to compute a 50% majority-rule consensus tree. For ML analysis, we conducted heuristic searches under the TN93 + G + I model of mutation. The neighbour-joining algorithm (NJ) was used to calculate the genetic distances according to the TN93 model. The robustness of the inferred trees was assessed by bootstrapping (1000 replicates) in ML or NJ analyses and posterior probability values in BI analysis. Branch support values were evaluated in accordance with Yang et al. [34], where good support was defined as bootstrap values of 75–88% and posterior probabilities of 0.85–0.94, strong support as bootstrap values of 89–100% and posterior probabilities of 0.95–1. A number of polymorphic sites (S), nucleotide diversity (π), haplotype diversity (Hd) and neutrality tests were calculated using DnaSP 6 [35].
3. Results
The analysed individuals of Sabanejewia spp. originate from 18 localities with an elevational range from 15 to 269 m above sea level and a bottom slope varying from 0.4 to 6.8% (Table S1). The locality parameters represented different environmental conditions from lowland rivers (metapotamon) to sub-montane river habitats (metarithron).
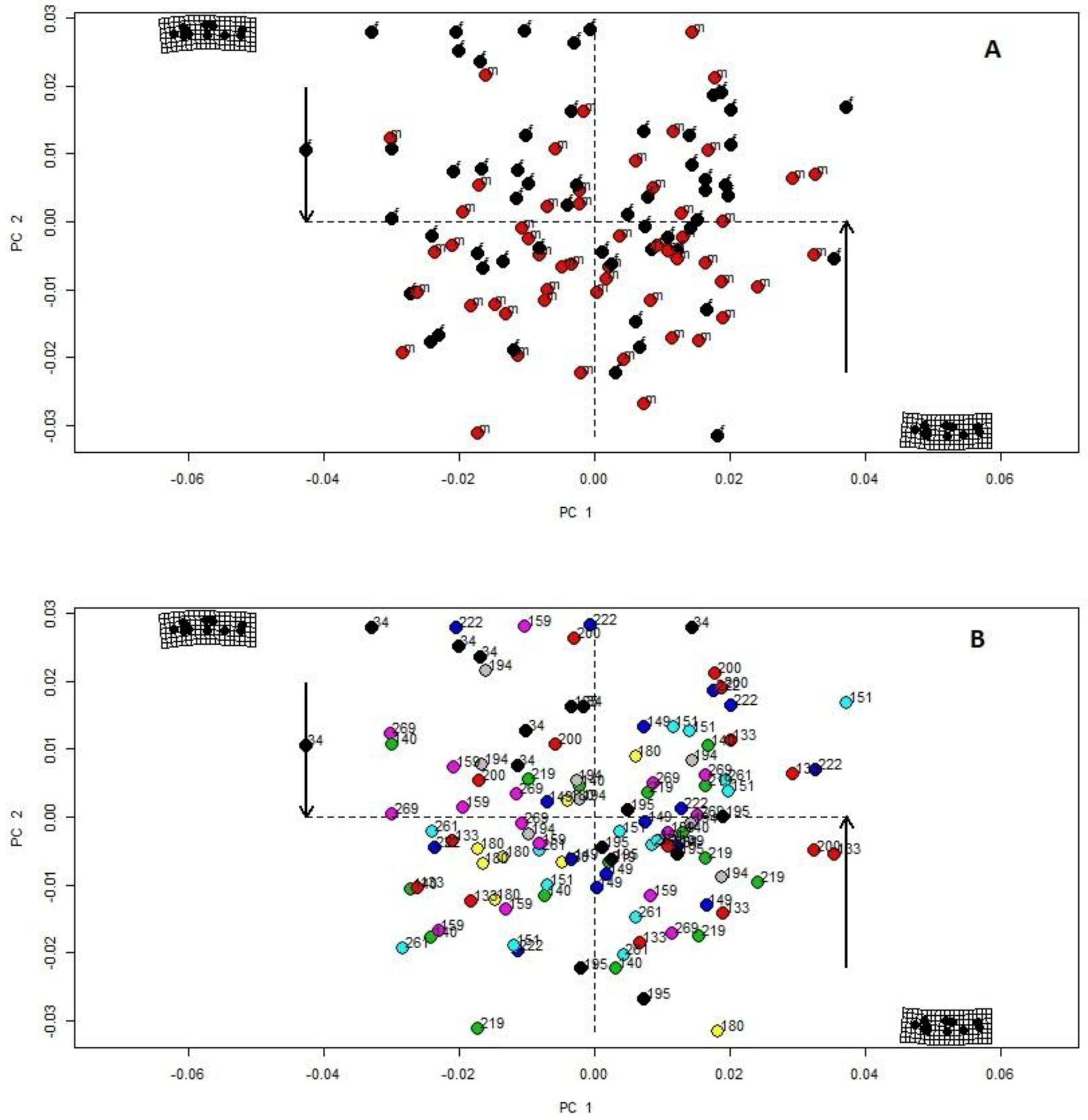
Body shape at the minima and maxima of the first two PCA axes explained 38.5% of the variability (PC1–23.4%, PC2–15.1%), with visible deformation in the pre-dorsal part represented by landmarks 2 and 6 (Figure 2). According to the permutation test results (Table 1), the significant impact on a specimen’s landmark variance was best explained by sex and the locality elevation.
Subsequently, we set PCA plot based on sex categories and elevation levels (Table S1). We retained two principal components according to the PCA variance graph, two categories for sex and fourteen categories for locality elevation (Figure 3A,B), responsible for approximately 35.5% of the morphological variation. The deformation grids in PCA plot (Figure 3A,B) showed the importance of the association with the change of landmark position representing the dorsum, the caudal peduncle and the base of the anal fin.
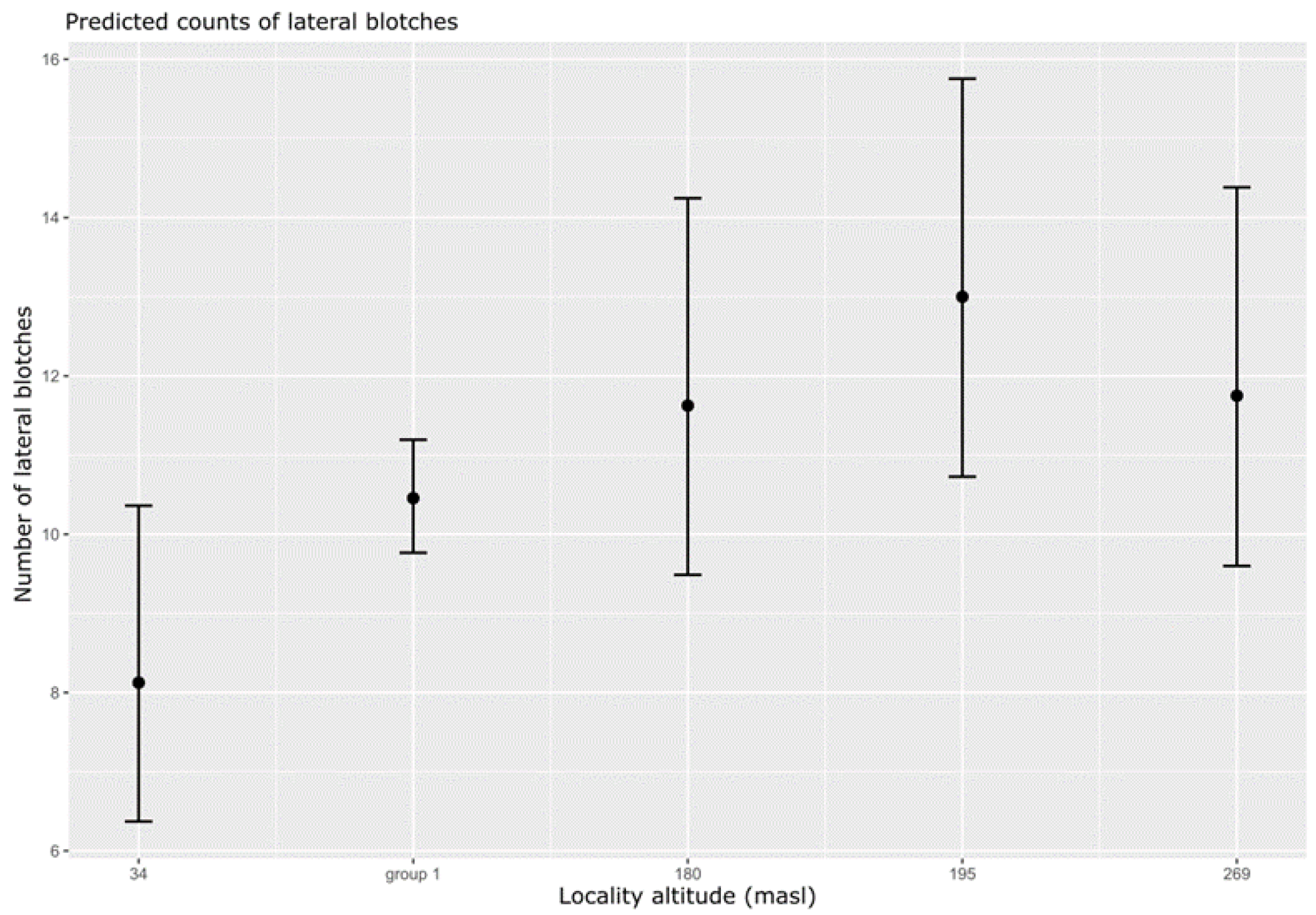
The body shape variability varied with the elevation (Figure 3B, Table 2) and the number of lateral spots decreased with elevation (Figure 4, Table 3).
Specimens from the type locality of S. balcanica in the Vardar River basin (Lepenac R., Treska R.: Table 1 No. 1, 3, elevation 269 m) had a significantly higher number of lateral spots (Table 3—Model fit 3, Figure 4). In contrast, individuals from the S. bulgarica type locality in the Danube River (Table S1. No. 17, elevation 34 m) had a lower number of spots (Table S1, Table 3—Model fit 3, Figure 4) and a different body shape (Table 1, Figure 3A). Based on the GLMM results (Table 3), we found a significant correlation between the locality elevation (grouped according to significant levels of the GLMM summary) and the number of lateral spots for the populations from the Danube River (34 m a.s.l.), group 1 of the localities (133–261 m a.s.l.), the Petrinjčica River (180 and 195 m a.s.l.) and the population from the Treska River (269 m a.s.l.). Based on our results (Figure 4) and the field observation, we suggest three ecological groups within the river continuum: (1) ≤132 m a.s.l. (lowland rivers–metapotamon lowland forms/monotypic fine sediments prevalent, lowland ecomorph dominant); (2) 133–260 m a.s.l. (middle river–epipotamon, variability of substrate grain size, high variability of ecomorphs); and (3) >180 m a.s.l. (foothills–hyporithron, fine sediments very rare or absent, increased number of spots to support surface mimicry).
Analysis of the mitochondrial sequences from 54 individuals identified 49 cyt b haplotypes based on 85 variable nucleotides and 49 parsimony informative sites. Overall, haplotype diversity was high (0.995 ± 0.003) with relatively low nucleotide diversity (0.0109 ± 0.0007). The values of Tajima’s D and Fu & Li’s D neutrality tests were negative (−1.3290 and −1.8483, respectively), but not significant, indicating an excess of low-frequency polymorphisms relative to expectation. The phylogenetic trees clustered analysed Sabanejewia haplotypes into six lineages (Figure 5).
All analysed samples of S. balcanica and S. bulgarica belonged to their expected lineages, S. balcanica (lineage II) and S. bulgarica (lineage III), respectively. The lineages were defined by the presence of sequences of specimens from the type localities of both species. The specimens analysed in this study belonged to subclades II (S. doiranica–S. balcanica) and III (S. montana–S. bulgarica–S. balcanica) within the DB complex according to Perdices et al. [13]. The mean genetic TN93 distance among lineages included in our study was 0.018 (range 0.009–0.027), while intraspecific/intragroup distances ranged from 0.001 to 0.003 (Table 4). Estimated molecular divergence between the sequences from the two type localities was 0.017.
4. Discussion
Molecular studies that have used genetic data to infer the phylogenetic relationships of Sabanejewia have shown that there is a complex of evolutionary lineages inhabiting the Danube–Balkanian region [11,13,14]; however, morphology does not parallel their taxonomy. Our study evaluates the importance of habitat type (environment) on external morphological characters (body shape and colouration).
The distribution of Sabanejewia spp. within a geographic region has been very often characterised by the optimal substrate composition [36,37] and habitats [15,38]. However, the ecological adaptability of the genus representatives is great and suggests genus distribution from river delta [39,40] to streams at an altitude of 750 m a.s.l. in Europe [1], and almost to 2000 m a.s.l. in Asia [41]. Our results showed a geographical gradient of phenotypes similar to those described by Bănărescu et al. [3]. The difference of phenotypes increases at the boundaries (min/max. elevation) within the analysed range of locality elevations (Figure 3B and Figure 4). Furthermore, the local habitat conditions give rise to different phenotypes characterised by body shape (Figure 3B) and the number of lateral spots (Figure 4). The intergradation of the phenotypes described by Bănărescu et al. [3] is evident in streams traversing larger elevation differences (Fedorčák, pers. obs.) characterised by diverse microhabitats, reflecting the transition between foothills to lowlands [15]. The cline of the external characters (body shape, lateral spots pattern) reflects the environmental conditions (hydrogeology), which include a range of ecological factors (water velocity, substrate composition, depth, etc.). Based on our data (Figure 4), we identified three ecological groups characterised by a different number of lateral spots and corresponding to the specific habitat conditions: (1) ≤132 m a.s.l. (lowland rivers–metapotamon, lowland forms, mean 8 spots); (2) 133–260 m a.s.l. (epipotamon, a high variability of forms, mean 10.5 spots); and (3) >180 m a.s.l. (hyporithron, sub-montane forms, mean 12 spots). Similar results have been described for populations of the Timiș River [3] (p. 30). Spot groups may overlap (Figure 4) or show a slightly different cline according to the specific environmental gradient in the context of local specifics of the river basin (length of the river basin, hydrology, geology, etc.) [42].
Our results also show that the specimens from the type localities of S. balcanica and S. bulgarica, analysed by us, represent different phenotypes from different ecological conditions (habitat) characterised by different elevations (Table S1—loc. 1, 3, 17). Moreover, the phylogenetic difference between S. balcanica (Figure 5. blue clade, lineage II) and S. bulgarica (Figure 5. red clade, lineage III) specimens from their type localities (Figure 5 and Figure 6A,B) is well distinguishable (0.009–0.027).
The haplotypic similarity found in S. balcanica among specimens from the type locality (Lepenac and Treska Rivers in the Vardar River basin) and Lake Doirani showed that S. balcanica haplotypes are present at both localities (Figure 5). However, in the Danube River basin, the S. balcanica haplotypes were observed only in two rivers (Dičina, Kutinska) belonging to the Great Morava River basin in Serbia [17], which are closest in geographic proximity to the Vardar basin. Consequently, we consider the presence S. balcanica lineage (II) in the Danube River basin of Bosnia and Herzegovina [44] to be questionable. In contrast, the presence of S. bulgarica lineage (III) has never been confirmed south of the Great Morava River basin (Serbia) and the Bulgarian Danube River basin. Furthermore, Križek et al. [15] suppose that S. bulgarica (lineage III) is widely distributed in the Danube drainage basin. We also propose that the distribution of lineage III is likely to be broader as it included specimens from the Croatian Danube tributaries recorded by Buj et al. [14]. The specimens from the type locality of S. bulgarica (lineage III, Vidin) have a different body shape (Table 1; Figure 3A elevation 34, Figure 6B). The evident morphological differences (bigger body, the shape of the mental lobe, longer barbels, etc.) of S. bulgarica specimens from Vidin have been previously documented also by Drensky [45] and Križek et al. [15]. Our samples differ mainly at the pre/post dorsal part (Figure 3A,B and Figure 6B–D). These differences are explained by the elevation of the locality and specimen’s sex (Table 2; Figure 3A,B). Križek et al. [15] also showed that the humpback shape (aD) increases with the presence of fine sediments (the most common sediments of lowland habitats). We hypothesise that Sabanejewia spp. from slow-flowing waters (the Danube River in Vidin) generally have laterally compressed and deeper bodies (Figure 2 and Figure 3C), which make them more flexible for maneuvering between river obstacles [46]. Taeniform (ribbon) shape fish are laterally compressed, which is generally advantageous for hiding in bottom shelters (macrophyte roots, river woods, stony particles, etc.). The “hump-backed” appearance pointed out by Iftime [5] is associated with downstream (lowland) forms and hormonal activity during the spawning period. Our results (Table 1; Figure 3B) similarly confirm shape differences affected by sex. We propose that these differences may be caused by an enlargement of the gonads during the spawning season with consequent differences during the season. Changes in the shape of the body of males known as the so-called lateral swelling [1] is linked to spawning behaviour [47].
Finally, S. bulgarica lineage (III) in their type locality (Danube River; Vidin) is externally well distinguishable due to their adaptation for specific habitat parameters given by low locality elevation (Table S1, No. 17). Križek et al. [15] reported the distribution of the S. bulgarica in the large area within the Danube basin, but the type locality (Danube River in Vidin) of this species is located at the lower edge of the elevational distribution (near the Danube River delta). Quite the opposite, S. balcanica (lineage II) from the type locality (Vardar River Basin; Figure 3B elevation 269, Figure 6A) does not differ morphologically from other specimens analysed (Figure 3B, elevations 149, 151, 159, 194, 200 m). This means that S. balcanica (lineage II) has the same body shape as S. bulgarica (lineage III) specimens occurring in a similar environment (environmental conditions at higher elevation). This result is supported by the observation that the body shape of S. balcanica (lineage II) from the lower Vardar River (Axios River, Greece) could be much more similar to the body shape of S. bulgarica from the type locality (Danube River, Bulgaria). This hypothesis is supported by findings of Economidis and Nalbant [48], who identified lowland morphotypes (seven spot forms) of S. balcanica at the streams around the Vardar (Axios) River in the lowlands of Greece. We can confirm a statistically significant difference in the number of lateral spots of S. balcanica (lineage II, type locality Vardar River) and S. bulgarica (lineage III, type locality Danube River) from different elevations (Table 3, Figure 3). This cline of Sabanejewia colour forms has been observed several times [3,4,49]. In addition, Križek et al. [14] confirmed the increase of the number of the lateral spots with the presence of a coarse substrate in foothill river zones. Here, we hypothesise that: (1) the deep habitat and higher turbidity of lowland rivers (metapotamon) favour the mechanoreceptor–barbels and affect diurnal activity of spined loaches [50,51], and (2) shallow (sub-mountain) habitats and the coarse-grained bottom of foothill rivers (metarithron) favour the photoreceptor (eye) and body mimicry (surface–spots). The high mimicry adaptability of the genus Sabanejewia is evident and appears at the boundaries within the elevational distribution and in slow-flowing and stagnant habitats. Among well-known examples is the striped form of S. caspia from the wetlands of Iran (Figure 6D) or high colour variation of S. baltica (Figure 6E,F) which can be even greater based on actualized distribution [11]. Further, small spotted S. larvata [6,52] were recorded in palustrine of the Po River [53] and karst lakes of Croatia [54]. Another visual difference can be caused by the presence of intergeneric crossbreds [55,56] and specific habitat conditions affecting water velocity and the process of particle sedimentation (e.g., distance from a confluence, presence of river obstacles, and flood dynamic change of bottom deposits). Interesting is also the colour peculiarity of lowland Sabanejewia characterized by the violet-metallic sheen of the body surface (Figure 6C) observed also by Sivkov [5] in the Arges River (Romania).
5. Conclusions
The results of our work confirm the phylogenetic differentiation between S. balcanica (Figure 5. blue clade, lineage II) and S. bulgarica (Figure 5. red clade, lineage III). Both species can have various phenotypes depending on the environmental conditions on their habitat. The lowland phenotype is independent of the evolutionary lineage and is found in different molecular clades. Different phenotypes are well distinguished by the lateral colour pattern which reflects the different ecological conditions, mainly elevation and type of the bottom substrate. The colour pattern of cobitid fishes is a character that is more adaptable than body shape and vary across the wide spectrum of the populated environment [57]. The influence of local habitat parameters on the external characteristics of fishes [58] is a key in phenotype adaptability to local habitat conditions [59]. The body shape of individuals of both species also varies depending on specimen’s sex. In further morphological studies of Sabanejewia DB complex, we recommend to take into consideration the type of habitat and the level of gonadal maturity.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fishes8020119/s1, Table S1: List of examined samples.
Author Contributions
J.F. contributed to conception, study design, statistics, morphometry, graphics and environmental data assessment; R.Š., T.S., M.N. and J.V. provide the samples and contributed to text of manuscript; J.M. processed molecular data, P.K. contributed to construction of the distribution map and consensus tree; A.P., R.Š, J.V. and J.K. supervised concept and reviewed text of manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The authors (JF: JK) gratefully acknowledge support from the Slovak Research and Development Agency under the project SK-AT-20-0009, the Scientific Grant Agency of the Slovak Republic VEGA 1/0553/23 and JF to the Slovak Academic Information Agency (no. 39062). RŠ acknowledges the Ministry of Culture of the Czech Republic (DKRVO 2019-2023/6.V.e National Museum, 00023272). AP was supported by the Synthesys projects CZ-TAF-8225, CA-TAF-8389. The authors would like to also thank Maroš Kubala and Yuliya Kutsokon for their consulting, technical help or provision of material.
Institutional Review Board Statement
Analysis was carried out on museum material. Sampling of museum material was in accordance with the national law for the collection of fish samples by all participating authors. Details are given in the material and methods section of the manuscript.
Data Availability Statement
Data are be available on request from the corresponding author. Will be applied after acceptance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vladykov, V. Sur un nouveau genre de Cobitides: Sabanejewia. Bull. Mus. Natl. D’hist. Nat. (Sér. 2) 1929, 2, 85–90. [Google Scholar]
- Banarescu, V.P. Intraspecific variation, subspeciation and speciation in Roumanian fresh-water fishes. J. Zool. Syst. Evol. Res. 1966, 4, 378–396. [Google Scholar] [CrossRef]
- Bănărescu, P.M.; Nalbant, T.T.; Chelmu, S. Revision and geographical variation of Sabanejewia aurata in Romania and the origin of S. bulgarica and S. romanica (Pisces, Cobitidae). Annot. Zool. Bot. Bratisl. 1972, 75, 49. [Google Scholar]
- Sivkov, Y. Morphological characteristic of the Danubian loach Sabanejewia bulgarica (Drensky, 1928) (Pisces, Cobitidae). Acta Zool. Bulg. 1991, 42, 34–43. [Google Scholar]
- Iftime, A. Considerations over the taxonomic status of the balkan golden loach (Sabanejewia balcanica) (Pisces: Ostariophysi: Cobitidae) in Romania and Republic of Moldova. Trav. Mus. Natl. D’hist. Nat. Grigore Antipa 2002, 44, 335–355. [Google Scholar]
- Freyhof, J.; Kottelat, M. Handbook of European Freshwater Fishes; Publications Kottelat: Cornol, Switzerland; Berlin, Germany, 2007; pp. 321–327. [Google Scholar]
- Bănărescu, P. Zoogeography of Fresh Waters. Distribution and Dispersal of Freshwater Animals in North America and Eurasia; AULA-Verlag: Wiesbaden, Germany, 1992; Volume 2. [Google Scholar]
- Witkowski, A. Morphological characteristics of Sabanejewia aurata (De Filippi, 1865) from the Odra River Basin, with description of a new subspecies (Teleostei: Cypriniformes: Cobitidae). Zool. Abh. Staatl. Mus. Tierkd. Dresd. 1994, 48, 23–51. [Google Scholar]
- Kottelat, M. Conspectus Cobitidum: An inventory of the loaches of the world (Teleostei: Cypriniformes: Cobitoidei). Raffles Bull. Zool. 2012, 26, 1–199. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References; California Academy of Sciences: San Francisco, CA, USA, 2022; Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 17 December 2022).
- Vasil’eva, E.D.; Solovyeva, E.N.; Vasil’ev, V.P. Molecular phylogeny of the Spined Loach Genus Sabanejewia (Osteichthyes: Cobitidae) Revised. J. Ichthyol. 2022, 62, 812–827. [Google Scholar] [CrossRef]
- Marić, S.P.; Bănăduc, D.; Gajić, Đ.D.; Šanda, R.; Veličković, T.Z. Sabanejewia romanica (Băcescu, 1943) (Actinopterygii: Cobitidae), a new species for the ichthyofauna of Serbia. Acta Zool. Bulg. 2022, 74, 369–377. [Google Scholar]
- Perdices, A.; Doadrio, I.; Economidis, P.S.; Bohlen, J.; Bǎnǎrescu, V.P. Pleistocene effects on the European freshwater fish fauna: Double origin of the cobitid genus Sabanejewia in the Danube basin (Osteichthyes: Cobitidae). Mol. Phylogenet. Evol. 2003, 26, 289–299. [Google Scholar] [CrossRef]
- Buj, I.; Podnar, M.; Mrakovčić, M.; Ćaleta, M.; Mustafić, P.; Zanella, D.; Marčić, Z. Morphological and genetic diversity of Sabanejewia balcanica in Croatia. Folia Zool. 2008, 57, 100–110. [Google Scholar]
- Križek, P.; Mendel, J.; Fedorčák, J.; Koščo, J. In the foothill zone—Sabanejewia balcanica (Karaman 1922), in the lowland zone—Sabanejewia bulgarica (Drensky, 1928): Myth or reality? Ecol. Evol. 2021, 10, 7929–7947. [Google Scholar] [CrossRef] [PubMed]
- Kottelat, M. European freshwater fishes. Biologia 1997, 52, 1–271. [Google Scholar] [CrossRef]
- Marešová, E.; Delić, A.; Kostov, V.; Marić, S.; Mendel, J.; Šanda, R. Genetic diversity of Sabanejewia balcanica (Actinopterygii: Cobitidae) in the western Balkans and comparison with other regions. Folia Zool. 2011, 60, 335–342. [Google Scholar] [CrossRef]
- Mousavi-Sabet, H.; Anvarifar, H. Landmark-based morphometric variation between Cobitis keyvani and Cobitis faridpaki (Pisces: Cobitidae), with new habitat for C. faridpaki in the southern Caspian Sea basin. Folia Zool. 2013, 62, 167–175. [Google Scholar] [CrossRef]
- Rohlf, F.J. TpsDig; Version 1.4; Stony Brook University: Stony Brook, NY, USA, 2004; Available online: https://life.bio.sunysb.edu/morph/ (accessed on 1 January 2020).
- Baken, E.; Collyer, M.; Kaliontzopoulou, A.; Adams, D. geomorph v4.0 and gmShiny: Enhanced analytics and a new graphical interface for a comprehensive morphometric experience. Methods Ecol. Evol. 2021, 12, 2355–2363. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 16 February 2023).
- Park, P.J.; Aguirre, W.E.; Spikes, D.A.; Miyazaki, J.M. Landmark-Based Geometric Morphometrics: What fish shapes can tell us about fish evolution. Proc. Assoc. Biol. Lab. Educ. 2013, 34, 361–371. [Google Scholar]
- Adams, D.; Collyer, M.; Kaliontzopoulou, A. Geomorph: Software for Geometric Morphometric Analyses. Version 3.2.1. 2020. Available online: https://cran.r-project.org/package=geomorph (accessed on 1 February 2023).
- Chessel, D.; Dufour, A.; Thioulouse, J. The ade4 package—I: One-table methods. R News 2004, 4, 5–10. Available online: https://cran.r-project.org/doc/Rnews/ (accessed on 1 February 2023).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 1 February 2023).
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Palumbi, S.R. Nucleic acids II: The polymerase chain reaction. Mol. Syst. 1996, 205–247. [Google Scholar]
- Perdices, A.; Doadrio, I. The Molecular systematics and biogeography of the European Cobitids Based on mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2001, 19, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Leford, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (* and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Posada, D. jModeltest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Yang, J.; He, S.; Freyhof, J.; Witte, K.; Liu, H. The phylogenetic relationships of the Gobioninae (Teleostei: Cyprinidae) inferred from mitochondrial cytochrome b gene sequences. Hydrobiologia 2006, 553, 255–266. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP v6: DNA sequence polymorphism analyses of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Marić, D.; Milošević, D. First record and morphological characteristics of the Balkan golden loach Sabanejewia balcanica (Cobitidae) in Montenegro. Period. Biol. 2010, 112, 149–152. [Google Scholar]
- Ratschan, C.; Gumpinger, C.; Schauer, M.; Wanzenböck, J.; Zauner, G. Arten-schutzprojekt Kleinfische und Neunaugen in Oberösterreich. Teil 2: Balkan-Goldsteinbeißer (Sabanejewia balcanica Karaman, 1922). Osterr. Fisch. 2011, 64, 174–188. [Google Scholar]
- Pekárik, L.; Koščo, J.; Košuthová, L.; Košuth, P. Coenological and habitat affinities of Cobitis elongatoides, Sabanejewia balcanica and Misgurnus fossilis in Slovakia. Folia Zool. 2008, 57, 172. [Google Scholar]
- Năstase, A.; Oţel, V.; Năvodaru, I. Ecological status of fish fauna in arms of the Danube Delta (Danube Delta Biosphere Reserve, Romania) at the beginning of the third Millennium. Acta Zool. Bulg. 2017, 69, 349–360. [Google Scholar]
- Sayyadzadeh, G.; Abbasi, K.; Esmaeili, H.R. Review and re-description of Sabanejewia species in Iran (Teleostei: Cobitidae). Iran. J. Ichthyol. 2019, 5, 277–292. [Google Scholar]
- Kaya, C.; Bayçelebi, E.; Turan, D. Taxonomic assessment and distribution of fishes in upper Kura and Aras river drainages. Zoosyst. Evol. 2020, 96, 325–344. [Google Scholar] [CrossRef]
- Erős, T.; Botta-Dukat, Z.; Grossman, G.D. Assemblage structure and habitat use of fishes in a Central European submontane stream: A patch-based approach. Ecol. Freshw. Fish 2003, 12, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Kutsokon, Y. Northern golden loach (Sabanejewia baltica Witkowski, 1994) in Ubort River. Stud. Biol. 2014, 8, 255–258. [Google Scholar] [CrossRef]
- Bajrić, A.; Buj, I.; Adrović, A.; Hajdarević, E. Morphological and genetic diversity of Sabanejewia balcanica (Cobitidae, Actinopterygii) in Bosnia and Herzegovina. J. Appl. Ichthyol. 2020, 37, 89–98. [Google Scholar] [CrossRef]
- Drensky, P. Die Fische der Familie Cobitidae in Bulgarien. Izv. Tsarskite Prir. Inst. Sofia 1928, 8, 156–181. [Google Scholar]
- Shuai, F.; Yu, S.; Lek, S.; Li, X. Habitat effects on intra-species variation in functional morphology: Evidence from freshwater fish. Ecol. Evol. 2018, 8, 10902–10913. [Google Scholar] [CrossRef]
- Bohlen, J. First report of the spawning behaviour of a golden spined loach, Sabanejewia vallachica (Teleostei: Cobitidae). Folia Zool. 2008, 57, 139–146. [Google Scholar]
- Economidis, P.S.; Nalbant, T.T. A study of the loaches of the genera Cobitis and Sabanejewia (Pisces, Cobitidae) of Greece, with description of six new taxa. Trav. Mus. Natl. D’hist. Nat. Grigore Antipa 1996, 36, 297–347. [Google Scholar]
- Fedorčák, J.; Šanda, R.; Stefanov, T.; Mendel, J.; Koščo, J. Influence of habitat on the external morphology of Sabanejewia (Cypriniformes: Cobitidae) specimens. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Nowak, M.; Klaczak, A.; Szczerbik, P.; Popek, W. Diel differences in the exploitation of shallow inshore habitats by three small bottom-dwelling fishes with implications for their monitoring in a large lowland river. Arch. Pol. Fish. 2018, 26, 159–164. [Google Scholar] [CrossRef]
- Nowak, M.; Klaczak, A.; Koščo, J.; Szczerbik, P.; Fedorčák, J.; Hajdú, J.; Popek, W. Diel changeover of fish assemblages in shallow sandy habitats of lowland rivers of different sizes. Knowl. Manag. Aquat. Ecosyst. 2019, 420, 41. [Google Scholar] [CrossRef] [Green Version]
- Ittiofauna. Available online: https://tinyurl.com/45jyrwhc (accessed on 7 February 2023).
- Siti. Biosphere Reserve Nomination Form Collina Po. Ente di Gestione Aree Protette Po e Collina Torinese. 2015. Available online: www.parchipocollina.to.it/PDF/mab_collinapo.pdf (accessed on 7 February 2023).
- Buj, I.; Mustafić, P.; Ćaleta, M.; Marčić, Z.; Ivić, L.; Žalac, S.; Raguž, L. Peculiar occurrence of Cobitis bilineata Canestrini, 1865 and Sabanejewia larvata (De Filippi, 1859) (Cobitidae, Actinopteri) in the Danube River basin in Croatia. Fundam. Appl. Limnol. 2021, 194, 201–213. [Google Scholar] [CrossRef]
- Mezhzherin, S.V.; Pukhtayevych, P.P.; Tsyba, A.A. Absorbing hybridization of Cobitis taenia and Sabanejewia aurata (Cypriniformes, Cobitidae) in water reservoirs of northern Ukraine connected with diploid-polyploid complex formation. Vestn. Zool. 2014, 48, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Vasil’eva, E.D.; Vasil’ev, V.P. Natural hybridization in spined loaches of the genera Cobitis and Sabanejewia (Cobitidae). J. Ichthyol. 2019, 59, 776–785. [Google Scholar] [CrossRef]
- Saitoh, K.; Aizawa, H. Local differentiation within the striated spined loach (the striata type of Cobitis taenia complex). Jpn. J. Ichthyol. 1987, 34, 334–345. [Google Scholar] [CrossRef]
- Poisot, T.; Verneau, O.; Desdevises, Y. Morphological and molecular evolution are not linked in Lamellodiscus (Plathyhelminthes, Monogenea). PLoS ONE 2011, 6, e26252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuigan, K.; Franklin, C.E.; Moritz, C.; Blows, M.W. Adaptation of rainbow fish to lake and stream habitats. Evolution 2003, 57, 104–118. [Google Scholar] [CrossRef]
Figure 1.
Map showing sampling localities; the numbers (1–18) correspond to the numbers of the localities in Table S1. Colours depict haplotype groups: red—S. bulgarica, blue—S. balcanica, grey—localities are not included in cyt b analysis. Shapes: red square—S. bulgarica in Danube River basin; blue circle—S. balcanica in Danube River basin; blue/red star—co-occurrence of S. balcanica and S. bulgarica in Danube River basin (Black Sea drainage); blue stars—S. balcanica in Vardar River catchment (Aegean Sea drainage).
Figure 1.
Map showing sampling localities; the numbers (1–18) correspond to the numbers of the localities in Table S1. Colours depict haplotype groups: red—S. bulgarica, blue—S. balcanica, grey—localities are not included in cyt b analysis. Shapes: red square—S. bulgarica in Danube River basin; blue circle—S. balcanica in Danube River basin; blue/red star—co-occurrence of S. balcanica and S. bulgarica in Danube River basin (Black Sea drainage); blue stars—S. balcanica in Vardar River catchment (Aegean Sea drainage).

Figure 2.
Sabanejewia body with 11 landmarks (middle)—1. Tip of snout, 2. Forehead (end of frontal bone), 3. End of operculum (ventral part), 4. End of the operculum (lateral), 5. Dorsal origin of pectoral fin, 6. Origin of dorsal fin, 7. Origin of ventral fin, 8. Termination of dorsal fin, 9. Origin of anal fin, 10. Base of caudal fin (dorsal), 11. Base of caudal fin (ventral).
Figure 2.
Sabanejewia body with 11 landmarks (middle)—1. Tip of snout, 2. Forehead (end of frontal bone), 3. End of operculum (ventral part), 4. End of the operculum (lateral), 5. Dorsal origin of pectoral fin, 6. Origin of dorsal fin, 7. Origin of ventral fin, 8. Termination of dorsal fin, 9. Origin of anal fin, 10. Base of caudal fin (dorsal), 11. Base of caudal fin (ventral).

Figure 3.
Principal components (PCA) of geometric morphometry based on 11 landmarks (Figure 2) of 111 individuals of Sabanejewia spp. based on specimens’ sex ((A) black dots–females, red dots–males) from 15 localities differing in elevation ((B)numbers and colours represent locality elevation). Significant explanatory variables are identified by PERMANOVA (Table 1).
Figure 3.
Principal components (PCA) of geometric morphometry based on 11 landmarks (Figure 2) of 111 individuals of Sabanejewia spp. based on specimens’ sex ((A) black dots–females, red dots–males) from 15 localities differing in elevation ((B)numbers and colours represent locality elevation). Significant explanatory variables are identified by PERMANOVA (Table 1).

Figure 4.
Generalised linear mixed model (GLMM, results of Table 3, Model fit 3) plot showing a significant correlation between the number of lateral spots and elevation (numbers represent locality altitude in meters) (results of Table 3., model 3.) for 111 individuals of Sabanejewia spp. Elevation: 34 m–River Danube (type locality Vidin, Vrav, BG); 180 m and 195 m–River Petrinjčica (loc. Miočinovići, Dodoši, HR); 269 m–River Treska (type locality Skopje, NMK), group1–other localities (133–261 m elevation) (Table S1).
Figure 4.
Generalised linear mixed model (GLMM, results of Table 3, Model fit 3) plot showing a significant correlation between the number of lateral spots and elevation (numbers represent locality altitude in meters) (results of Table 3., model 3.) for 111 individuals of Sabanejewia spp. Elevation: 34 m–River Danube (type locality Vidin, Vrav, BG); 180 m and 195 m–River Petrinjčica (loc. Miočinovići, Dodoši, HR); 269 m–River Treska (type locality Skopje, NMK), group1–other localities (133–261 m elevation) (Table S1).

Figure 5.
Bayesian consensus tree of Sabanejewia DB complex haplotypes resulting from the analysis of cyt b sequence data. Bayesian posterior probabilities and ML/NJ bootstrap values are listed above the branches (only value 1 for BI and ≥75% for ML and NJ are shown). The letters in brackets indicate the international country codes. The tip labels are river names and haplotypes or the GenBank accession number (letter H–from [17]; letters AF, AY–from [13], letters V from [15] letters B, T–our samples; see Table S1) * S. bulgarica type locality, ** S. balcanica type locality.
Figure 5.
Bayesian consensus tree of Sabanejewia DB complex haplotypes resulting from the analysis of cyt b sequence data. Bayesian posterior probabilities and ML/NJ bootstrap values are listed above the branches (only value 1 for BI and ≥75% for ML and NJ are shown). The letters in brackets indicate the international country codes. The tip labels are river names and haplotypes or the GenBank accession number (letter H–from [17]; letters AF, AY–from [13], letters V from [15] letters B, T–our samples; see Table S1) * S. bulgarica type locality, ** S. balcanica type locality.

Figure 6.
Colouration patterns of the Sabanejewia spp. originating from different types of habitat and elevation. (A) S. balcanica, type locality River Treska in Skopje (269 m a.s.l.); (B) S. bulgarica, type locality River Danube in Vidin (34 m a.s.l.); (C) surface metallic sheen of S. bulgarica originating in the Danube delta of Ukraine (Figure A. M. Naseka in [6]); (D) S. caspia–Anzali wetland (28 m a.s.l.) (in [40]); (E) S. baltica–River Ubort near Maidan-Kopishchenskii (154 m a.s.l., river near wetlands) (Figure in [43]); and (F) S. baltica–River Stradomka near Chrostowa (215 m a.s.l.) (Figure Michal Nowak).
Figure 6.
Colouration patterns of the Sabanejewia spp. originating from different types of habitat and elevation. (A) S. balcanica, type locality River Treska in Skopje (269 m a.s.l.); (B) S. bulgarica, type locality River Danube in Vidin (34 m a.s.l.); (C) surface metallic sheen of S. bulgarica originating in the Danube delta of Ukraine (Figure A. M. Naseka in [6]); (D) S. caspia–Anzali wetland (28 m a.s.l.) (in [40]); (E) S. baltica–River Ubort near Maidan-Kopishchenskii (154 m a.s.l., river near wetlands) (Figure in [43]); and (F) S. baltica–River Stradomka near Chrostowa (215 m a.s.l.) (Figure Michal Nowak).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
PERMANOVA results (Adonis [20]; 9999 permutations) for geometric morphometric data and explanatory variables (sex, elevation, slope) and their double interaction based on Euclidian distances. (Abbreviations: SS–sum of squares, MS–mean sum of squares, d.f–degrees of freedom).
Table 1.
PERMANOVA results (Adonis [20]; 9999 permutations) for geometric morphometric data and explanatory variables (sex, elevation, slope) and their double interaction based on Euclidian distances. (Abbreviations: SS–sum of squares, MS–mean sum of squares, d.f–degrees of freedom).
| Main Effects | d.f. | SS | MS | Pseudo F | R2 |
|---|---|---|---|---|---|
| Elevation | 13 | 0.032 | 0.003 | 2.468 | <0.001 |
| Sex | 1 | 0.005 | 0.005 | 4.828 | <0.001 |
| Slope | 1 | 0.0008 | 0.0008 | 0.821 | >0.076 |
| Residuals | 95 | 0.096 | 0.001 | ||
| Total | 110 | 0.134 | |||
| Interaction terms | d.f. | SS | MS | pseudo F | R2 |
| Elevation | 13 | 0.032 | 0.003 | 2.543 | <0.05 |
| Sex | 1 | 0.004 | 0.005 | 4.975 | <0.001 |
| Slope | 1 | 0.0008 | 0.0008 | 0.847 | 0.033 |
| Elevation:sex | 13 | 0.016 | 0.001 | 1.239 | >0.06 |
| Sex:slope | 1 | 0.0008 | 0.0008 | 0.783 | >0.33 |
| Residuals | 81 | 0.079 | 0.001 | ||
| Total | 110 | 0.134 |
Table 2.
Morphometric variance of 111 individuals of Sabanejewia spp. within the groups and pairwise comparisons of group variances (two ways) based on elevation of the localities listed in Table S1. Elevation in m a.s.l.
Table 2.
Morphometric variance of 111 individuals of Sabanejewia spp. within the groups and pairwise comparisons of group variances (two ways) based on elevation of the localities listed in Table S1. Elevation in m a.s.l.
| Observed Variances by Group | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.00053 | 0.00086 | 0.00068 | 0.00056 | 0.00072 | 0.00076 | 0.00077 | 0.00053 | 0.00044 | 0.00087 | 0.00086 | 0.00085 | 0.00066 | 0.00101 | ||
| Elevation | 34 | 133 | 140 | 149 | 151 | 159 | 180 | 194 | 195 | 200 | 219 | 222 | 261 | 269 | |
| Pairwise P-values between variances | 34 | 1 | |||||||||||||
| 133 | 0.1069 | 1 | |||||||||||||
| 140 | 0.4537 | 0.3722 | 1 | ||||||||||||
| 149 | 0.8656 | 0.1542 | 0.5719 | 1 | |||||||||||
| 151 | 0.333 | 0.5124 | 0.8185 | 0.4335 | 1 | ||||||||||
| 159 | 0.2417 | 0.647 | 0.6647 | 0.3221 | 0.8451 | 1 | |||||||||
| 180 | 0.2402 | 0.6701 | 0.6458 | 0.3146 | 0.8215 | 0.9811 | 1 | ||||||||
| 194 | 1 | 0.1108 | 0.4723 | 0.8704 | 0.3411 | 0.2494 | 0.2355 | 1 | |||||||
| 195 | 0.6642 | 0.0401 | 0.2455 | 0.5447 | 0.1653 | 0.1127 | 0.1062 | 0.6664 | 1 | ||||||
| 200 | 0.1059 | 0.9546 | 0.3666 | 0.1473 | 0.4917 | 0.6234 | 0.6309 | 0.1118 | 0.0436 | 1 | |||||
| 219 | 0.11 | 0.9903 | 0.374 | 0.1518 | 0.5194 | 0.6557 | 0.6691 | 0.1147 | 0.0456 | 0.9473 | 1 | ||||
| 222 | 0.1197 | 0.9542 | 0.4058 | 0.1678 | 0.5491 | 0.6929 | 0.7053 | 0.1229 | 0.0491 | 0.914 | 0.9611 | 1 | |||
| 261 | 0.4992 | 0.3404 | 0.9529 | 0.6062 | 0.7781 | 0.6153 | 0.6109 | 0.5072 | 0.264 | 0.3318 | 0.3453 | 0.3737 | 1 | ||
| 269 | 0.0211 | 0.4488 | 0.0964 | 0.031 | 0.1597 | 0.2322 | 0.2305 | 0.0215 | 0.0055 | 0.4998 | 0.4412 | 0.4147 | 0.0897 | 1 | |
Table 3.
Summary of GLMM (with mixed model effect of the localities) for lateral blotches (spots) of the 111 Sabanejewia spp. and explanatory variables.
Table 3.
Summary of GLMM (with mixed model effect of the localities) for lateral blotches (spots) of the 111 Sabanejewia spp. and explanatory variables.
| GLMM Family | Response | Model Fit | Explanatory | df | Chi2 | p | ANOVA | AIC | Df |
|---|---|---|---|---|---|---|---|---|---|
| Poisson | Lateral blotches | Intercept (full model) | 1 | 3.9 | <0.05 | Type 3 | 500 | 6 | |
| 3 (ANOVA summary) | Elevation2 (groups) | 4 | 10.9 | <0.05 | |||||
| Model summary | Est | SE | Pr (>|z|) | ||||||
| Intercept (Elevation 34) | 2.09 | 0.12 | <0.01 | ||||||
| (Elevation group1) | 0.25 | 0.13 | <0.05 | ||||||
| (Elevation 180) | 0.36 | 0.16 | <0.05 | ||||||
| (Elevation 195) | 0.47 | 0.16 | <0.01 | ||||||
| (Elevation 269) | 0.37 | 0.16 | <0.05 | ||||||
| 2 | Intercept | 1 | 3.9 | <0.05 | Type 3 | 517 | 18 | ||
| Elevation | 13 | 13.97 | >0.37 | ||||||
| Sex | 1 | 0.79 | >0.77 | ||||||
| Slope | 1 | 0.58 | >0.44 | ||||||
| SL | 1 | 0.68 | >0.40 | ||||||
| 1 | Elevation | 13 | 13.97 | >0.37 | Type 2 | 569 | 46 | ||
| Sex | 1 | 0.03 | >0.86 | ||||||
| Slope | 1 | 0.16 | >0.68 | ||||||
| SL | 1 | 0.72 | >0.39 | ||||||
| Double int. (all exp.) | no sig. effect of interaction | ||||||||
Table 4.
Genetic divergences (by the TN93 model) on cyt b sequences between the molecularly studied Sabanejewia species. (The number of base differences per site from averaging over all sequence pairs between groups are shown; standard error estimate(s) are shown above the diagonal. Distances within groups are shown diagonally in bold).
Table 4.
Genetic divergences (by the TN93 model) on cyt b sequences between the molecularly studied Sabanejewia species. (The number of base differences per site from averaging over all sequence pairs between groups are shown; standard error estimate(s) are shown above the diagonal. Distances within groups are shown diagonally in bold).
| S. bulgarica | S. balcanica | “S. radnensis” | “S. thrakica” | Lin. VI. | S. vallachica | |
| S. bulgarica | 0.003 | 0.003 | 0.003 | 0.004 | 0.003 | 0.004 |
| S. balcanica | 0.018 | 0.003 | 0.002 | 0.003 | 0.003 | 0.004 |
| “S. radnensis” | 0.014 | 0.013 | 0.003 | 0.003 | 0.002 | 0.004 |
| “S. thrakica” | 0.019 | 0.018 | 0.013 | 0.002 | 0.003 | 0.005 |
| Lineage VI. | 0.015 | 0.015 | 0.009 | 0.014 | 0.001 | 0.004 |
| S. vallachica | 0.027 | 0.026 | 0.019 | 0.026 | 0.02 | 0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fedorčák, J.; Šanda, R.; Stefanov, T.; Mendel, J.; Nowak, M.; Križek, P.; Perdices, A.; Vukić, J.; Koščo, J. The “True Colours” of Golden Loaches (Teleostei: Cobitidae). Fishes 2023, 8, 119. https://doi.org/10.3390/fishes8020119
AMA Style
Fedorčák J, Šanda R, Stefanov T, Mendel J, Nowak M, Križek P, Perdices A, Vukić J, Koščo J. The “True Colours” of Golden Loaches (Teleostei: Cobitidae). Fishes. 2023; 8(2):119. https://doi.org/10.3390/fishes8020119
Chicago/Turabian StyleFedorčák, Jakub, Radek Šanda, Tihomir Stefanov, Jan Mendel, Michal Nowak, Peter Križek, Anabel Perdices, Jasna Vukić, and Ján Koščo. 2023. "The “True Colours” of Golden Loaches (Teleostei: Cobitidae)" Fishes 8, no. 2: 119. https://doi.org/10.3390/fishes8020119