
Kocuria Strains from Unique Radon Spring Water from Jachymov Spa
by
 , , , and
, , , and
Elizaveta Timkina
1,* ,
,
Lucie Drábová
1,
Andrea Palyzová
2,
Tomáš Řezanka
2,
Olga Maťátková
1 and
Irena Kolouchová
1 1
Department of Biotechnology, University of Chemistry and Technology, 166 28 Prague, Czech Republic
2
Institute of Microbiology, The Czech Academy of Sciences, 142 20 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Fermentation 2022, 8(1), 35; https://doi.org/10.3390/fermentation8010035
Submission received: 10 December 2021
/
Revised: 7 January 2022
/
Accepted: 10 January 2022
/
Published: 16 January 2022
(This article belongs to the Special Issue Extremophiles—Source for Novel Biomolecules with Applied Potential)
Abstract
:Members of the genus Kocuria are often found in soils contaminated with toxic metals or exposed to high levels of ionizing radiation. The use of classical cultivation technics often leads to the isolation of Kocuria sp. from underground spring waters. These bacterial isolates have to adapt their metabolism to survive in such extreme environments. Four bacterial isolates of the genus Kocuria (Kocuria sp. 101, 208, 301, and 401) were obtained from radon spring water (Jachymov, Czech Republic). These isolates were tested for their ability to withstand stress and extreme conditions. Growth was observed at a temperature range of 10–45 °C with optimal growth temperature between 20 and 30 °C. The content of polyunsaturated fatty acids in all four isolates was proved to be temperature-dependent. The strain Kocuria sp. 301 showed high resistance to all studied extreme conditions (UV radiation, desiccation, and free radicals in medium). The results suggest that isolates from radioactive springs might have developed mechanisms that help them survive under several extreme conditions and could be used in biotechnological production.
1. Introduction
Groundwater mineral springs are environments in which unique microorganisms with specific development can be found [1]. Only a few microorganisms are able to exist under specific conditions, such as the complete absence of light; low availability of organic substrates; the presence of toxic substances, such as heavy metals and radionuclides. The specifics of the environment force these microorganisms to develop unique metabolic pathways that can be used, for example, in bioremediation technologies. Many chemical reactions used in biotechnological production could be improved by performing them at extremes of temperature, pressure, salinity, and pH for efficient and cost-effective outcomes. Therefore, it is essential to look for organisms that thrive in extreme environments found in nature and could thereby be a source of metabolites used in biotechnology. Bacterial communities from underground radioactive mineral spring water could be a potent source of microorganisms with extraordinary properties.
Mineral springs found in the small town of Jáchymov (Sankt Joachimsthal) in the Czech Republic are special and unique in the world due to the high concentration of dissolved radon. The spa town of Jáchymov is located in a protected valley on the southern slope of the Ore Mountains. It is famously known as the place where Marie and Pierre Curie isolated radium from the mineral uraninite for the first time in 1898. The discovery of radioactive thermal springs is closely connected with mining activities. Natural mineral water with a very high radon (222Rn) content originates in an old mine, where it is in contact with the uranium vein structure of the mountain granite massif. The water in the granite massif has a high temperature, which increases the capacity to dissolve mineral salts and gases. Currently, four springs (Agricola, Behounek, C1, and Curie) containing radioactive gas radon are being used for spa purposes, for example, for the treatment of nervous and rheumatic disorders by constantly produced radioactive gas radon (222Rn) dissolved in the water. The radioactivity may reach 24 kBq/L [2]. The reservoirs of such mineral waters provide a unique environment for the evolution and existence of extremophilic microorganisms.
One of these highly resistant genera of bacteria is bacteria of the genus Kocuria, which are often found in underground mineral waters [3,4]. In terms of biotechnological use, these bacteria have a diverse range of biotechnologically exploitable products and metabolic processes [5,6,7].
The genus Kocuria are Gram-positive, non-endospore-forming, catalase positive, coagulase-negative, non-hemolytic cocci. The Kocuria genus can be distinguished from other representatives of the order Actinomycetales based on the presence of galactosamine and glucosamine in the cell wall, the presence of polar lipids (diphosphatidylglycerol and phosphatidylglycerol), menaquinones MK-7 and MK-8, as well as their peptidoglycane type (L-Lys-Ala3/4). Their dominant fatty acids are saturated branched fatty acids such as anteiso-C15:0. Their genomic DNA consists of GC content between 60% and 70% [8]. In the case of individual members of the genus Kocuria, diversification could be performed based on physiology, sequence analysis of 16S rRNA genes, or DNA-DNA hybridization analysis. Individual strains of the genus Kocuria also differ in the types of phospholipids, different phenotypic, chemotypic, and molecular characterization, as well as biochemical tests, optimum growth conditions, and pigmentation [9,10].
In biotechnological applications are members of this genus rather underutilized, although they have significant potential for the production of enzymes, antibiotics, biosurfactants or pigments in food, cosmetics, and pharmaceutical industries. Bernal et al. (2006) identified and isolated a K. rosea LPB-3 strain which is able to degrade feathers as a source of carbon, energy, and nitrogen [5]. The keratinase present in this strain exhibited extremely high heat stability (70–90 °C) and resistance to alkaline pH (10–11). K. varians NCC 1482 was found to produce a lantibiotic variacin, inhibiting the growth of food pathogens such as Enterococcus, Staphylococcus aureus, Bacillus cereus, and Clostridium botulinum [11]. Sarafin et al. (2014) tested eight halophilic bacteria for biosurfactant activity. One of the tested bacterial strains was K. marina BS-15. This strain was proved to produce biosurfactants, which are able to emulsify coconut, olive, and crude oil [12].
Many studies, including metagenomics, are dedicated to studying extremophile prokaryotes for example from marine habitats, due to the presence of many extreme factors, such as high UV radiation levels [13,14,15]. These studies have identified such microorganisms as valuable sources of novel genes, which can be considered as a source of novel bioproducts, including enzymes and other bioactive metabolites [16]. Marine environment comprises a broad variation of habitats leading to the discovery of various new extremozymes such as proteases, lipases, glycoside hydrolases, etc., with promising technological potential in industrial processes [17,18]. Microorganisms from habitats exposed to high radiation doses may similarly harbor genes with biotechnological potential, although little is known about the metabolic pathways, resistance, or secondary metabolites of radiation-resistant bacteria. Production of primary and secondary metabolites with capacity to protect cells from ionizing radiation or oxidative stress is a potentially highly utilizable biotechnological property of radiation-resistant microorganisms. Most radiation-resistant bacteria produce pigments with a high antioxidant capacity [19]. Extremophilic radioresistant bacteria and algae can be used to develop radioprotective drugs to protect the skin from UV exposure or to suppress the development of cancer [20,21]. Extremophiles are, therefore, a sustainable source of valuable substances for the biotechnological industry that needs to be explored.
Bacteria of the genus Kocuria are unique due to their ability to survive in extreme conditions [22]. The defense mechanisms employed by these bacteria to survive high doses of radiation, desiccation, high or low temperatures may be useful for biotechnological (bioremediation), pharmaceutical, and therapeutic applications. It is known that they can produce biotechnologically interesting metabolites such as lipopeptide-based biotensides [12,23,24], auxins [6], siderophores [23,24], pigments (mainly carotenoids) [25,26], extracellular carbohydrates (e.g., kocuran isolated from the genus K. rosea) [27] and antibiotics (kocurin and kocumarin isolated from the genus K. marina) [28] under various, sometimes extreme, conditions.
This study aimed to examine and characterize four bacterial isolates, identified as bacteria of the genus Kocuria, all of which were isolated from radioactive radon (222Rn) spring water in Jachymov, Czech Republic. The isolation, identification process, and phylogenetic relationship was already described in our previous work [29]. In this study we examined their characteristics that contribute to resistance to extreme conditions, such as exposure to UV-C radiation, desiccation, and free radicals in the medium. Furthermore, we evaluated their metabolic adaptations which could be impacted by their origin, such as fatty acid, siderophores and antioxidant production or various carbon sources utilization.
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
The bacterial strains of Kocuria sp. 101, 208, 301, and 401 from four radioactive springs in Jachymov, Czech Republic (50°22.357 N, 12°54.663 E) were isolated. The isolation and identification process was already described previously. Partial sequences of 16S ribosomal RNA gene have been deposited at GenBank under the accession number MH538969 (isolate 101), MH539149 (isolate 208), MH539151 (isolate 301), MH539150 (isolate 401). Isolate 101 was obtained from water spring Agricola, isolate 208 from spring Behounek, isolate 301 from spring C1 and isolate from spring Curie [29]. Luria-Bertani (LB) medium (in g/L: tryptone 10, yeast extract 5, and NaCl 10) was used to prepare pre-inoculum cultures of the strain (100 mL cultures incubated for 72 h at 30 °C on an orbital shaker at 150 rpm).
For the preparation inoculum, a sterile LB medium (200 mL) was inoculated with the pre-inoculum to give a final OD600 of 0.2 ± 0.020. The inoculum was cultured for 24 h (30 °C, 150 rpm). Control strains were grown under aerobic conditions in B8 medium at temperatures 30 °C (Deinococcus radiodurans CCM 1700T) and in LB medium 37 °C (Escherichia coli CCM 4517).
2.2. Growth at Different Temperatures
Cultures (in triplicate) were inoculated with pelleted biomass from pre-cultures washed at a ratio of 10%. The cultures were incubated on an orbital shaker (150 rpm) at different temperatures (5, 10, 20, 30, 40, 45, 50 °C). The growth parameters (e.g., optical density, cell dry weight (cdw), number of viable cells (CFU)) were monitored spectrophotometrically at regular time intervals. The cultures in the stationary phase (72 h) of growth were harvested by centrifugation (10 min, 9000× g, 4 °C), the pellet was separated and washed twice. Biomass was frozen at −75 °C and lyophilized.
2.3. Isolation of Lipids and Fatty Acid Methyl Ester (FAME) Characterization
Cultures prepared at temperatures 10, 20, and 30 °C (see Section 2.1) were suspended in a chloroform–methanol mixture (2:1). The sample was centrifuged, and the lower phase was evaporated and washed three times with 1 M KCl. The resulting chloroform phase was evaporated to dryness.
The preparation and analysis of FAME were described previously [30,31]. After saponification of the total lipid extracts, fatty acids were esterified by methanolic BF3 and analyzed by GC-MS Finnigan 1020 B operated in the electron ionization mode (EI) on polar capillary column (Supelcowax 10; 60 m × 0.25 mm i.d., 0.25 mm film thickness; Supelco, Prague, Czech Republic). The GC conditions were as follows: splitless injection at 100 °C; oven temperature program: 100 °C (1 min); 20 °C.min−1 to 180 °C; 2 °C.min−1 to 280 °C (1 min). Carrier gas helium at a linear velocity of 60 cm⋅s−1. Spectra were scanned within the range of m/z 50–500, and the structures of FAME were confirmed by comparison of retention times and fragmentation patterns with those obtained by the analysis of the standard FAME (Supelco, Prague, Czech Republic) [29].
2.4. Carbon Source Utilization Characterization
The preparation of the bacterial inoculum was performed according to Section 2.1. The concentration of cells in physiological saline solution was adjusted to the desired value OD600 of 0.1 ± 0.01. For the characterization of the metabolic profile of the studied microorganisms, EcoPlates microtiter plates (Biolog, Hayward, USA) were used. EcoPlates contain three sets of 31 carbon sources and employ a tetrazolium redox dye as an indicator of microbial metabolism. The EcoPlate substrates were grouped into distinct biochemical categories: alcohols, amines, amino acids, carboxylic acids, esters, carbohydrates, and polymers. In total, 150 μL of the bacterial suspension were pipetted into each well of the EcoPlate. Incubation was static at 30 °C for 6 days and the substrate utilization was monitored by measuring absorbance at 590 nm each day with microplate reader Tecan Infinite 200 PRO. The average well color development (AWCD) of each plate was expressed according to Garland and Milles (1991) as:
where Ri was the optical density (OD590) of each substrate (where I = 1–31), respectively. The color development of these substrate groups was expressed as substrate average well color development (AWCD) [32].
2.5. Determination of Antioxidant Production
A total of 50 mL of liquid LB medium (see Section 2.1) were inoculated with one colony of isolated bacteria. DPPH (2,2-diphenyl-1-picrylhydrazyl radical) assay was used for the determination of antioxidant production. Cultivation was carried out for 7 days (30 °C, 150 rpm). The cultured colonies of isolated bacteria were then centrifuged, and 100 μL of the supernatant were mixed with 200 μL of DPPH-methanol solution (5.2 mg/L of DPPH) in a microtiter plate. After 15 min of incubation in the dark, the absorbance of the mixture was measured at 517 nm. Ascorbic acid (AA) was used as a standard for the calibration curve (2.5–15 mg/L), and the results were expressed as ascorbic acid equivalents (mgAA/L).
2.6. Determination of Siderophore Production
The universal Chrome Azurol S (CAS) assay was used to test the ability of strains to produce iron-binding compounds of siderophore-type in a solid medium [33]. LB agar plates supplemented with CAS (60 mg), hexadecyltrimethylammonium bromide (72.9 mg), piperazine-N,N’-bis(2-ethanesulfonic acid) (32.24 g), and Fe3+ (2.7 mg of FeCl3·6H2O dissolved in 10 mM HCl) were used to detect siderophore producing strains. The plates were incubated for 48 h at 30 °C. The CAS/HDTMA forms a tight complex with ferric iron to create a blue color in the medium. When a strong iron chelator such as siderophore removes iron from the dye complex, the colonies form orange/yellow zones, which is considered as a positive reaction.
2.7. Determination of UV-C Resistance
The concentration of cells in LB medium prepared according to Section 2.1 was adjusted to the desired value OD600 = 0.50 ± 0.05. Then, 2 mL of bacterial suspension were applied to the bottom of a sterile petri dish. Irradiation was performed using a UV lamp (Prolux GM 30 W), with the highest radiation intensity in the UV-C spectrum (100–280 nm), from a distance of 0.4 m. An unirradiated bacterial suspension was used as a control. Irradiation time was 1, 5, 15, and 60 min, the equivalent doses of UV-C radiation were as follows: 36, 180, 540, and 2160 mJ/cm2. Viable cell counts were determined by preparing serial dilutions of each sample and control, plating them on LB agar plates, and enumerating colony forming units (CFU) after 48 h of growth at 30 °C. Each measurement was performed in triplicate. D. radiodurans CCM 1700T and E. coli CCM 4517 were taken as positive and negative controls, respectively.
Inactivation curves were obtained by plotting the logarithm of the surviving fractions of bacteria log (N/N0) against UV-C doses. Chick–Watson law was used for determination of graph parameters: , where N0 stands for the number of viable cultivable non-irradiated bacteria before UV-C exposure, N is the number of viable cultivable bacteria after UV-C exposure, K is the Chick–Watson coefficient of specific lethality following UV irradiation, I is the UV-C intensity expressed in mW/cm2, n is the coefficient of dilution assumed to be 1, t is the exposure time (s). The greater the coefficient K is, the more liable microorganism is [34].
2.8. Determination of Tolerance to Hydrogen Peroxide (H2O2)
The inoculum prepared according to Section 2.1 was used for this test. In total, 125 μL bacterial suspension were pipetted into a sterile microtiter plate. Afterwards, 150 μL of sterile saline solution with H2O2 was added to the well to achieve a final concentration of 0–5% (0–1.46 M) of H2O2. The OD600 was measured at regular intervals. Each measurement was performed in 5 parallels. If the monitored strain was able to tolerate certain concentrations of H2O2 in the culture medium, cultivations were further performed in Erlenmayer flasks (20 mL media in 100 mL flasks). The preparation of the bacterial inoculum was again performed according to Section 2.1. Sterile LB medium containing the desired concentration of H2O2 was inoculated to an OD600 = 0.20 ± 0.02. The measurement was performed in triplicates. The growth of the bacterial isolates was monitored each day spectrophotometrically (OD600).
2.9. Determination of Desiccation Tolerance
First, 200 μL of the LB medium with the concentration of the cells adjusted to the value OD600 = 0.5 ± 0.05 was transferred into a microtiter plate in 5 parallels. The plate was placed in a desiccator at 30 °C. After 30 days in the desiccator, the cells were rehydrated with 200 μL phosphate-buffered saline (PBS). Determination of cell viability was performed using the fluorescent dyes propidium iodide (PI) and SYTO 9 n [35]. Then, 2 μL of SYTO 9 (0.1 mmol/L) and 2 μL of PI (1 g/L) were added, the suspension was vortexed and left in the dark for 5–10 min. Samples were analyzed by the flow cytometer (BD FACSAria III). Controls were obtained from a freshly prepared inoculum. An unstained bacterial sample was stained to set up the flow cytometer and confirm autofluorescence. The measurements were performed in duplicates.
2.10. Statistical Analysis
Dixon’s Q test was used for detection of outliers in data obtained by the determination of antioxidant capacity (the determination was performed in five parallels, the deviation of the five determinations was less than 5%). The determination of fatty acids composition, carbon source utilization was performed in three parallels. The standard deviation of the measurement was less than 5%.
3. Results and Discussion
This study characterizes four Kocuria sp. isolates (Kocuria sp. 101, 208, 301, 401) with a focus on their ability to withstand stress and extreme conditions represented by a wide range of temperatures, UV-C radiation, tolerance to hydrogen peroxide and desiccation and on their adaptation strategies, such as siderophore production, antioxidant production, and various carbon sources utilization.
One of the unique properties of these bacteria is the growth of strains over a wide range of temperatures. For example, the strain K. marina KMM 3905T isolated from marine sediment collected in the Western Siberian Sea was able to grow in temperatures ranging from 4 to 43 °C [36]. In our study, the growth parameters of OD600 and the specific growth rate at different temperatures (5–50 °C) were monitored for individual strains during cultivation in LB medium. As can be seen from Table 1, the highest specific growth rates, as well as the maximum OD600 values of the tested strains, were obtained for both 20 and 30 °C. In complex LB medium at 20 °C, the specific growth rate varied in the range of 0.068–0.083 h−1 and at 30 °C in the range of 0.066–0.085 h−1. Growth at 50 °C was not observed in any of the tested isolates. However, all studied strains of Kocuria sp. showed an ability to grow at 45 °C. A large number of representatives of the class Actinobacteria are known to be able to adapt to high temperatures. Thermotolerant representatives of the Actinobacteria class have developed various strategies for maintaining homeostasis at higher temperatures, such as high content of GC pairs in the genome, which are more thermally stable. In addition, a higher content of GC pairs leads to a higher proportion of polar amino acids (asparagine, glycine, arginine, and lysine) and a decrease in the proportion of acidic amino acids (aspartate and glutamate), thus increasing the thermostability of the proteins. For example, arginine is thought to form stronger ionic bonds due to resonant structures [37]. Furthermore, bacteria can adapt to high temperatures more easily due to specific components of their cell membranes. The predominant respiratory menaquinone of most thermotolerant bacteria is saturated menaquinone type 9. Members of the genus Kocuria were proved to have membrane isoprenoid menaquinone types 7, 8, or 9, and the content of GC pairs in the genome is in the range of 60–75 mol% [8].
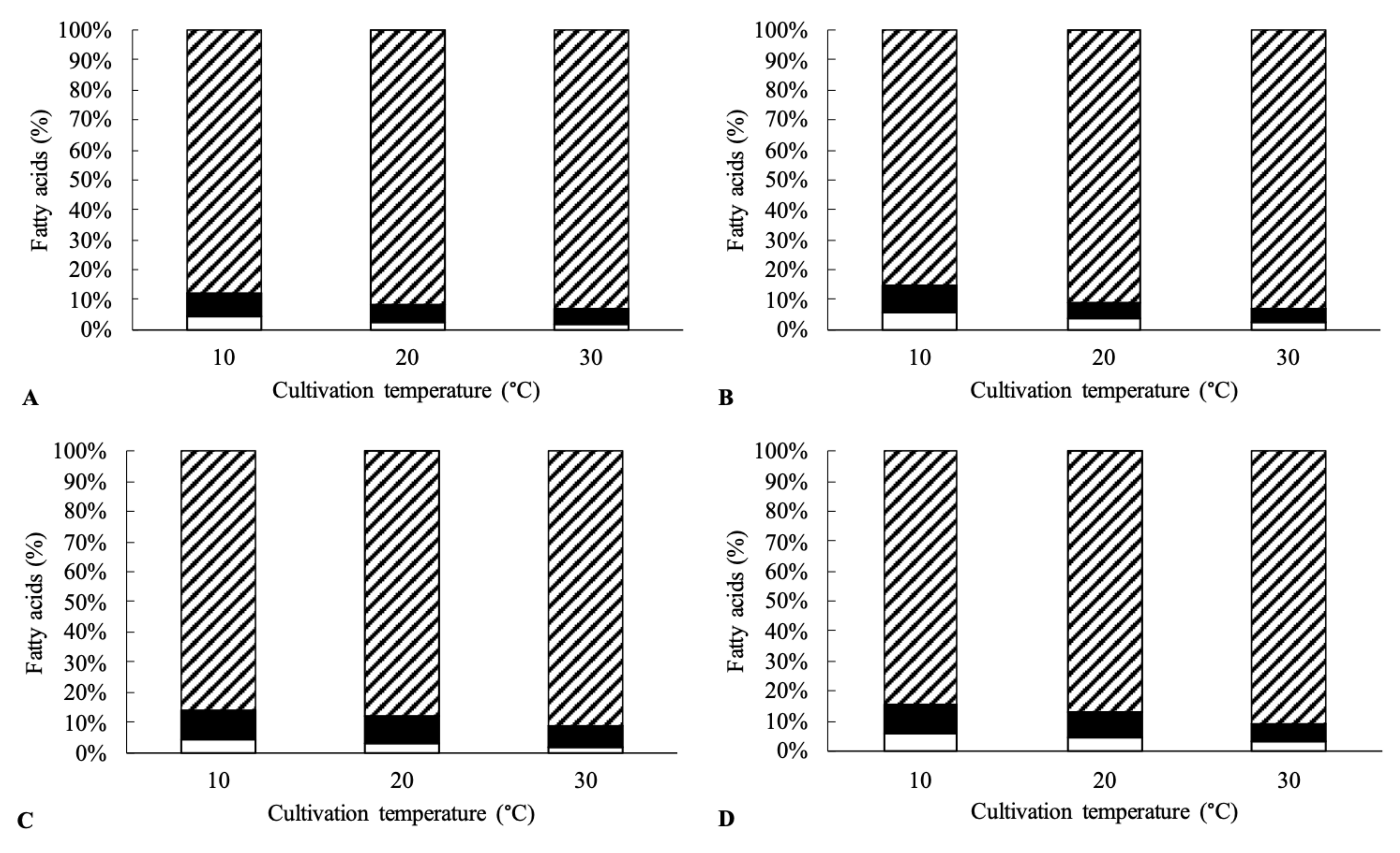
Out of the four tested strains, only two strains (Kocuria sp. 101 and Kocuria sp. 301) were able to grow at 5 °C, even though the specific growth rate of these two bacterial strains was almost three times lower than at 20 and 30 °C. The ability of the tested strains to grow at low temperatures may be attributed to the production of polyunsaturated fatty acids, which helps maintain the fluidity of cell membranes at low culture temperatures. Polyunsaturated fatty acids have a low melting point and thus contribute to increased membrane fluidity [38]. At 10 °C, the total unsaturated fatty acids in the cells ranged from 12.5% (Kocuria sp. 101) to 15.5% (Kocuria sp. 401) of the total fatty acids (see Figure 1). With increasing temperature, the content of unsaturated fatty acids in the cells decreased and, at 30 °C reached values ranging from 6.8% (Kocuria sp. 101) to 9.4% (Kocuria sp. 401) of the total amount of fatty acids. For all strains, the content of polyunsaturated fatty acids, specifically arachidonic (C20:4 ω-6) and eicosapentaenoic acid (C20:5 ω-3), ranged from 1.8 to 3.6% at 30 °C and 4.9–6.5% at 10 °C. The main monounsaturated fatty acids included palmitoleic acid (C16:1 ω-7) and 14-methylpalmitic acid (anteiso-C17:1 ω-9).
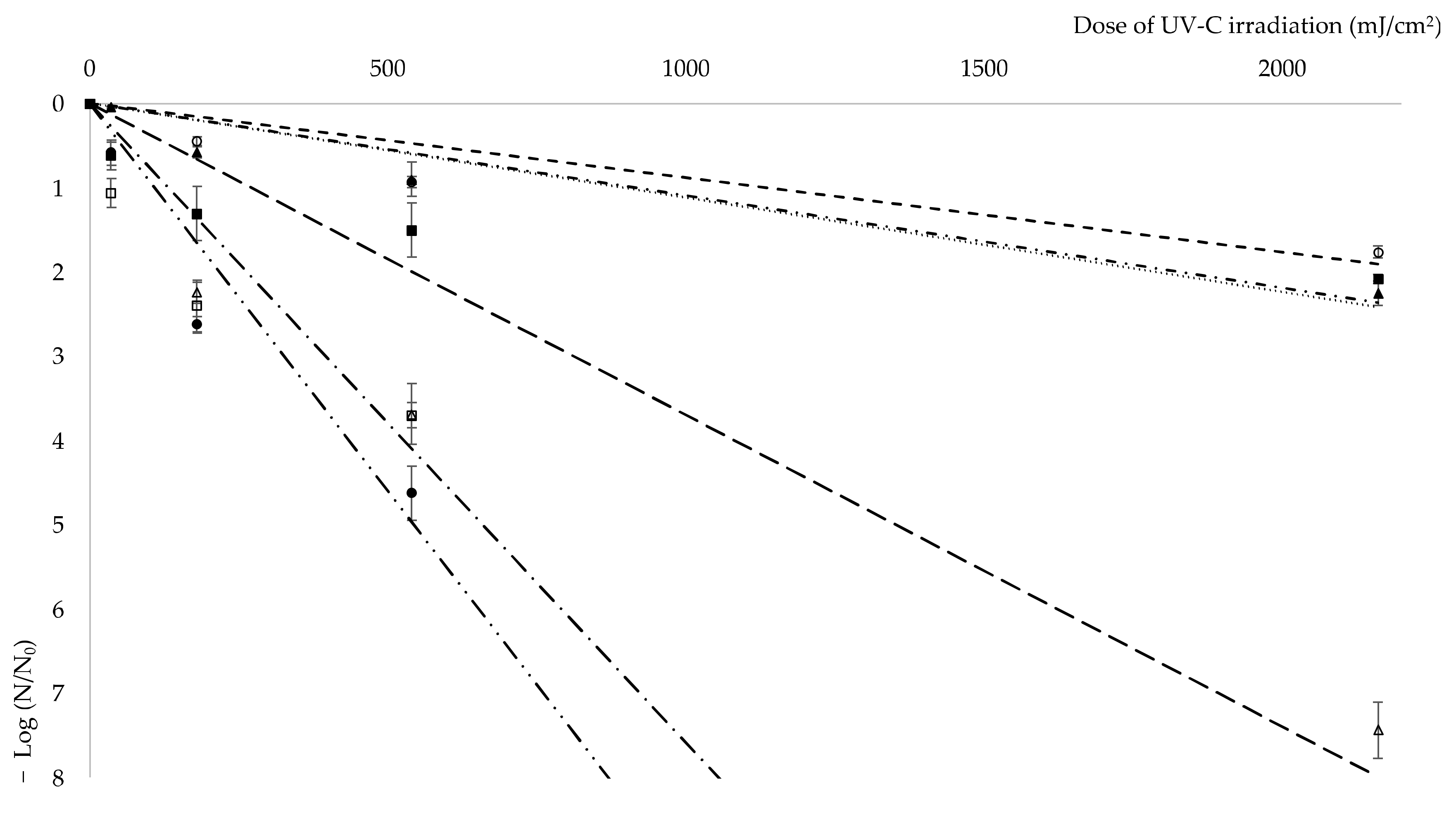
The resistance of bacterial cells to ionizing radiation is another unique property of some bacteria species. Radiation causes direct damage (through direct biomolecules damage) and indirect damage (through the formation of reactive oxygen species) to the living cells [39,40]. The molecules of water inside the cells are subjected to radiolysis, which involves homolytic cleavage of bonds to form the hydroxyl radicals HO•. The hydroxyl radical has a great oxidizing capacity but a short lifetime, and therefore only damages molecules which are close by. The hydroxyl radical has no reaction specificity and damages all organic molecules (DNA, RNA, lipids, and proteins). These radicals damage DNA molecules in ionizing radiation resistant and liable cells in the same way. However, sensitive cells are easily subjected to oxidative damage of proteins by ionizing radiation. Based on these findings, it can be assumed that the ability of resistant strains to minimize protein damage in a short time helps them to maintain functional and effective enzymatic equipment for repairing DNA damage [41,42]. UV radiation is a common type of radiation with great impact on human health [43]. Compounds able to protect bacterial cells against UV radiation and other oxidative damage are of great value. Due to the rising level of UV exposure, the search for novel sunscreen compounds is important [44]. UV-C resistance of the four studied Kocuria sp. strains was investigated. The survival diagram (Figure 2) shows a sharp decline in living Kocuria sp. 208 and 401 cells after irradiation. In case of isolates 208 and 401, coefficient of specific lethality (K = 0.0076 and K = 0.0092, respectively) is even greater than of E. coli CCM 4517 (K = 0.0037). No colony forming units of Kocuria sp. 401 were observed after as little as 5 min of irradiation (dose 180 mJ/cm2) and after 15 min (dose 540 mJ/cm2) of irradiation in the case of Kocuria sp. 208, as well as negative control E. coli CCM 4517. After 15 min of irradiation, cell viability of Kocuria sp. 101 decreased to 13% and of Kocuria sp. 301 to 3%, positive control D. radiodurans CCM 1700T showed a similar survival rate after 30 min (dose 1080 mJ/cm2) and 45 min (dose 1620 mJ/cm2) of irradiation, respectively. After 60 min of irradiation, under 1% of viable Kocuria sp. 101 and 301 cells were detected. Coefficient of specific lethality for isolates 101 and 301 was like that of D. radiodurans CCM 1700T: 0.0011, 0.0011, and 0.0009, respectively. In comparison with isolates from this work, Pseudomonas aeruginosa, a common opportunistic pathogen of humans, shows decrease in viability by 6-fold after a dose of 150–200 mJ/cm2, whereas isolates 101 and 301 went down by 2-fold after a dose of 2160 mJ/cm2 [45].
A secondary source of hydroxyl radicals in UV-C exposed cells is the Fenton reaction (Fe2+ + H2O2 → Fe3+ + HO• + OH−). Cells with a high Mn:Fe ratio have been shown to display significantly higher resistance to ionizing radiation than cells with a high iron content. Manganese ions can replace ferrous ions in the active sites of enzymes and thus maintain their activity, while reducing the intensity of the Fenton reaction (Daly, 2009) [46]. In order to obtain iron from the environment bacteria produce siderophores. Siderophores bind Fe3+ and facilitate its transfer into the cell, where Fe3+ gets reduced to Fe2+ [47]. The production of siderophores leads to an increase in the concentration of Fe2+ ions in the cell and thus may lead to a decrease in resistance to UV radiation. The production of siderophores was demonstrated in all four Kocuria sp. isolates, but only with low activity. In the case of siderophore production, the iron is bounded from the agar and the color changes from green to orange. Larger halo zones could be observed in strains of Kocuria sp. 208 and Kocuria sp. 401. Kocuria sp. 101 and Kocuria sp. 301 produced visibly smaller halo zones on CAS agars and both strains were also more resistant to UV-C radiation.
Another mechanism to reduce oxidative stress in cells before it damages vital molecules is the production of compounds with antioxidant activity [19]. As shown in Table 2, the highest antioxidant activity in cultivation media determined by DPPH method was observed in the strain Kocuria sp. 301 (8.9 mgAA/L). For the other studied strains, Kocuria sp. 101, Kocuria sp. 208, and Kocuria sp. 401, the antioxidant activity was similar and ranged from 4.1 to 5.4 mgAA/L.
The production of antioxidants may be closely related to tolerance to H2O2 in cultivation media. The screening tests in microtiter plates showed that even small amounts of H2O2 inhibited the growth of Kocuria sp. 208 and Kocuria sp. 401. Kocuria sp. 101 and Kocuria sp. 301 were able to tolerate 1% of H2O2 in cultivation media. These results were further confirmed on a larger scale in Erlenmeyer flasks. In the case of Kocuria sp. 101, growth in Erlenmeyer flasks was observed only at 0.5% H2O2. The strain Kocuria sp. 301, which was proved to produce the largest amount of antioxidants (see Table 2), was able to grow in media with 1% of H2O2. A known mechanism that helps microorganisms tolerate free radicals is the production of eicosapentaenoic acid [48]. As shown in Table 2, out of all four tested strains, Kocuria sp. 301 produced the highest amount of eicosapentaenoic acid out of all the tested strains (2.1–4.5% of total fatty acids).
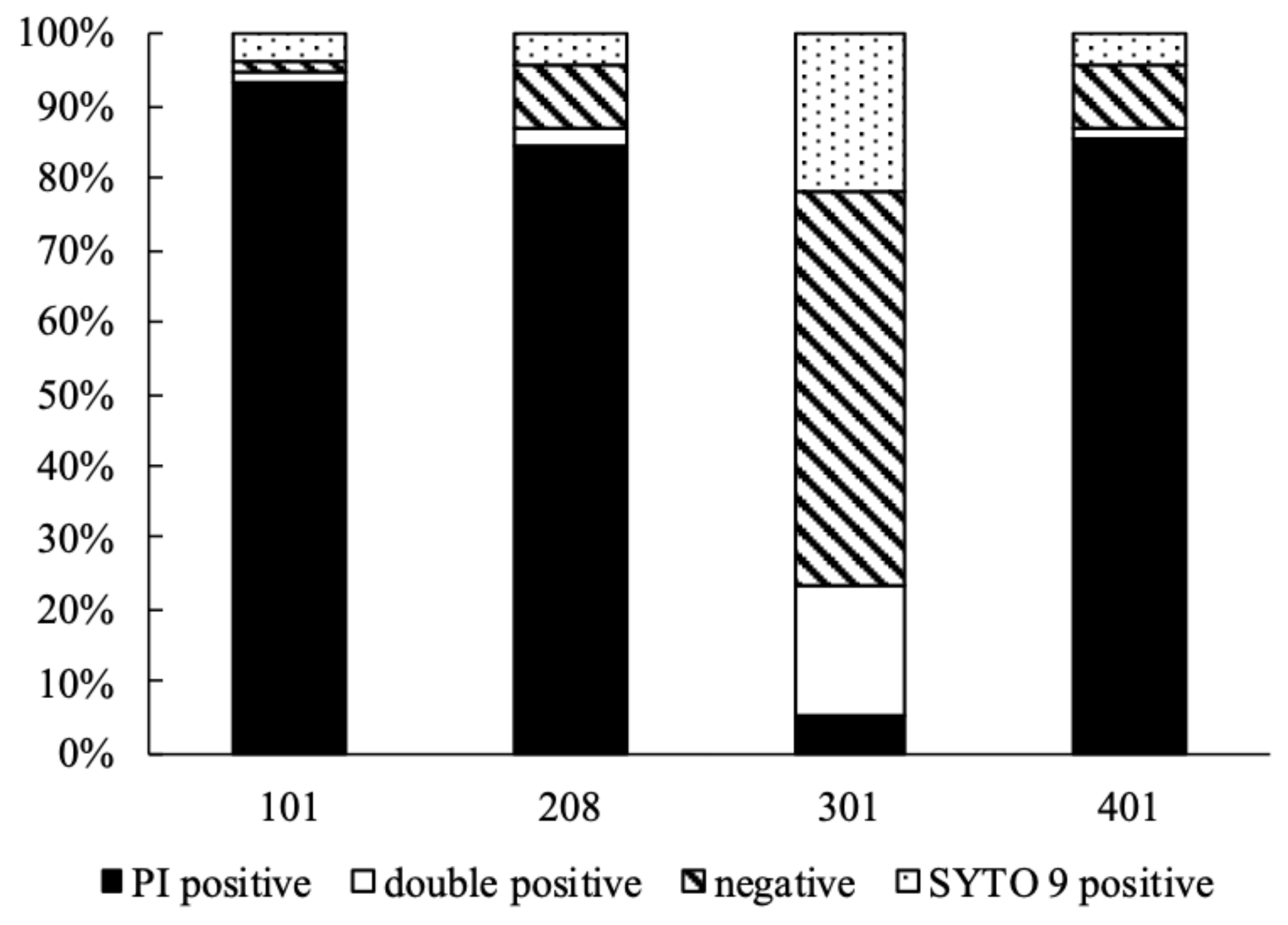
Desiccation has a similar effect on cells as ionizing radiation, which means that DNA molecules are damaged. After any DNA damage, the activity of repair mechanisms determines the ability of cells to survive. Studies show that in cells damaged by desiccation or ionizing radiation the activity of similar proteins increases [49]. During co-staining with PI and SYTO 9, SYTO 9 enters all cells regardless of their membrane integrity, binds to DNA and RNA and emits green fluorescence, while PI dyes cells with damaged cell membranes, binds to DNA and RNA and emits a red fluorescent signal [50,51]. Several Kocuria sp. isolates were proved to survive desiccation [4,22]. According to the obtained results by flow cytometry, the strain Kocuria sp. 301 had the largest share of the population stained with SYTO 9. Cell viability of this strain after 30 days of desiccation decreased to 20% while for other strains this value was around 4%. All tested strains had a large proportion of the population either stained with both dyes (PI and SYTO 9) or not stained at all. In each case, the proportion of unstained or double stained cells was similar in populations after desiccation and control population. Cells stained with both dyes can be considered dead [51], while unstained cells might indicate that they are alive because of the presence of active secretion mechanisms in living cells [35]. Taking this into account, the cell viability of Kocuria sp. 301 would be as high as 75% and only 5–13% for Kocuria sp. 101, 208, and 401 (see Figure 3).
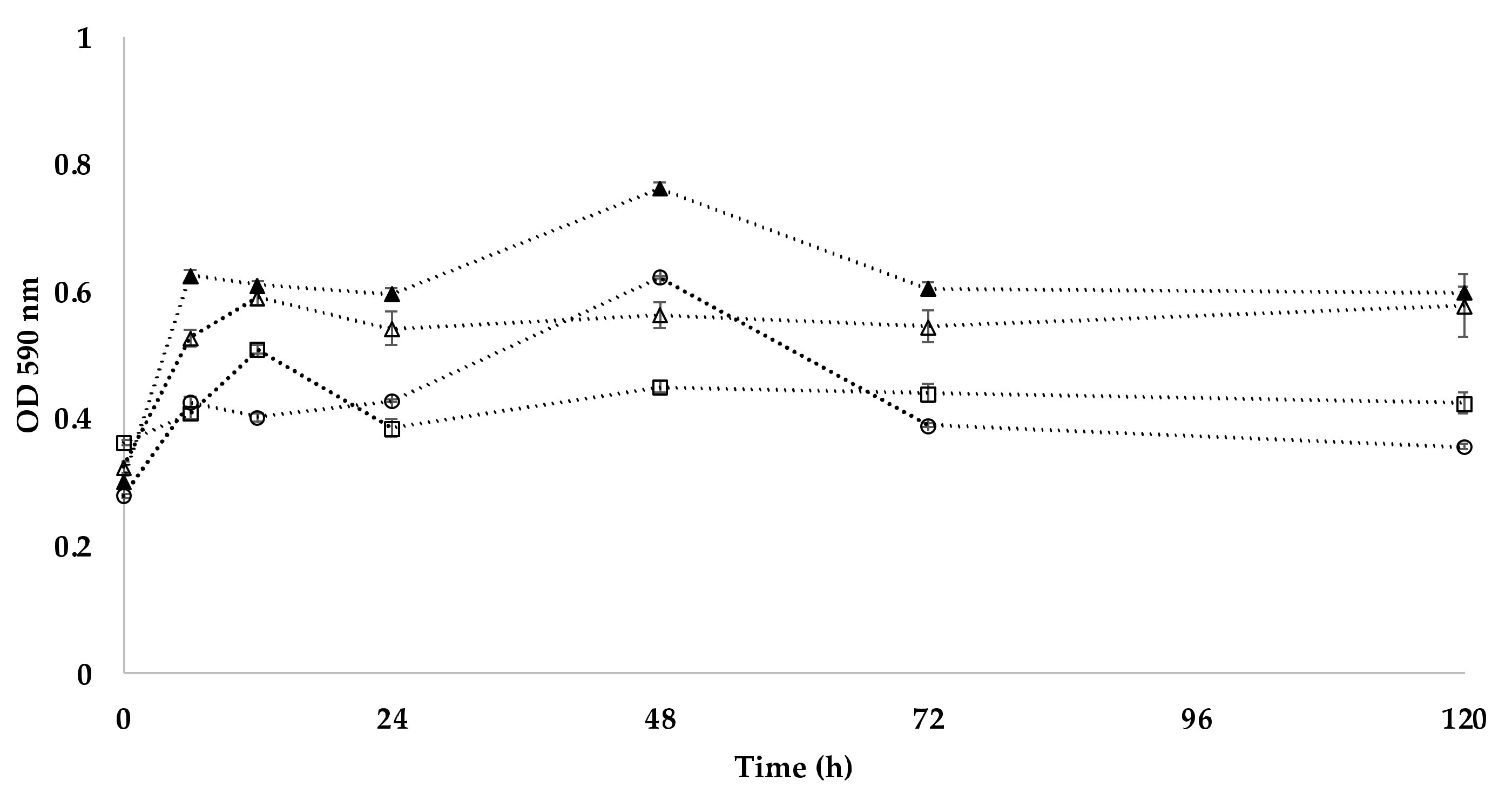
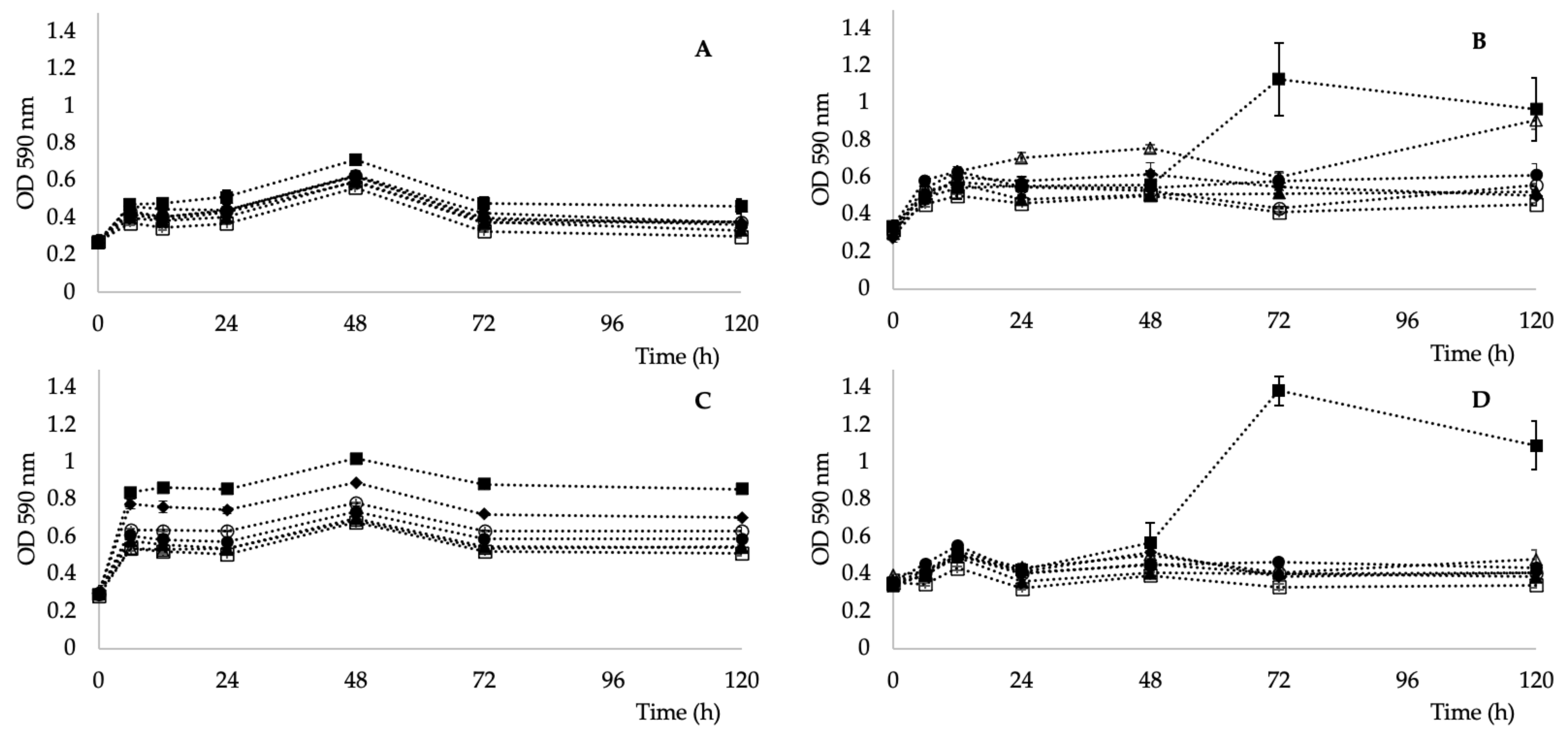
The last parameter tested was the metabolic activity, as bacterial isolates were obtained from an unusual habitat with low nutrient availability. As shown in Figure 4, the highest ability to utilize individual carbon sources (determined through AWCD) for all the tested carbon sources (alcohols, amines, amino acids, carboxylic acids, esters, hydrocarbons, and polymers) was observed in the strain Kocuria sp. 301 and the lowest in Kocuria sp. 101. All four strains were able to metabolize pyruvic acid methyl esters (see Figure 5). This ability has also been described in a K. rhizophila strain [52] and a K. arsenatis strain [53]. Esterase activity was also demonstrated in a K. pelophila strain isolated from the mangrove rhizosphere [54].
Biotechnological and pharmaceutical application of microorganisms resistant against high doses of radiation or intensive oxidative damage is conditioned by unique secondary metabolites production. Numerous biologically active compounds were proved to have protective properties against ionizing radiation like UV, α, β and γ rays. These compounds may be used for therapeutics production, such as antioxidants, anticancer drugs, or cell cycle inhibitors, etc. [55]. For example, scytonemin, a hydrophobic dimeric molecule composed of an indolic and a phenolic subunit with high absorption in UVB and UVA spectrum. Scytonemine has been characterized as a biochemical inhibitor for PLK1 and recognized as a non-specific ATP competitor. Stevenson et al. (2002) demonstrated that scytonemine can inhibit PLK1 in screening assays which reveals its ability to treat hyperproliferative skin disorders [56]. Many radiation resistant microorganisms produce carotenoid pigments [57,58]. Recent studies show that deinoxanthin, a carotenoid produced by D. radiodurans, can be used for the production of functionalized gold nanoparticles (DX-AuNPs). Those nanoparticles were shown to induce ROS formation in cancer cells by upregulation of several genes’ expression, which leads to cell apoptosis [59]. Among other bacteria, Actinobacteria are still the biggest source of antibiotics. Arid environment inhabiting Actinobacteria from Atacama Desert soils were shown to produce new ansamycin and 22-membered macrolactones with antibacterial and antitumor activity [60,61]. Another study reported novel aminobenzoquinones from Actinobacteria strain isolated from the Chilean highland soil. Molecules showed inhibitory activity against bacteria and dermatophytic fungi [62]. The pharmaceutical potential of Actinobacteria, including Kocuria sp., is huge. The more resistant strains need to be studied in more detail for their further possible use in the biotechnology and pharmaceutical industries.
4. Conclusions
In this work, the four bacterial isolates of the genus Kocuria (designated 101, 208, 301, and 401) were investigated for their ability to withstand stress and extreme conditions. The isolates were obtained from mineral radon water in Jachymov, Czech Republic, from the springs Agricola, Běhounek, C1, and Curie. Strains Kocuria sp. 101 and 301 were able to grow at temperatures ranging from 10 to 45 °C. Out of the four tested strains, Kocuria sp. 301 seems to be the most promising. Its high resistance against UV radiation, similar to the extremely radiotolerant D. radiotolerans, free radicals, and desiccation could be attributed to production of antioxidants, the presence of the polyunsaturated fatty acids, especially eicosapentaenoic acid, and also to low production of siderophores, which reduces the intracellular iron content. The results obtained are so unique that further detailed studies based on whole genome analysis will be performed to reveal unique metabolic pathways and protein or secondary metabolite production that would allow the properties of these extremely resistant microorganisms to be exploited in biotechnology.
Author Contributions
Conceptualization, I.K.; Formal analysis, E.T. and L.D.; Funding acquisition, I.K.; Investigation, E.T. and T.Ř.; Project administration, O.M.; Resources, O.M.; Supervision, O.M.; Visualization, L.D.; Writing—original draft, I.K.; Writing—review & editing, E.T., A.P. and T.Ř. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Czech Science Foundation (GACR) project 18-00036S. This research was funded with institutional support RVO 61388971.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors thanking Pihera and Ing. Loukota from Svornost mine for enabling sample collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Griebler, C.; Lueders, T. Microbial biodiversity in groundwater ecosystems. Freshw. Biol. 2009, 54, 649–677. [Google Scholar] [CrossRef]
- Krejbichova, Z. Radioactivity of mineral waters in Bohemia. Czech. J. Phys. 1999, 49, 127–132. [Google Scholar] [CrossRef]
- Asgarani, E.; Soudi, M.R.; Borzooee, F.; Dabbagh, R. Radio-resistance in psychrotrophic Kocuria sp ASB 107 isolated from Ab-e-Siah radioactive spring. J. Environ. Radioact. 2012, 113, 171–176. [Google Scholar] [CrossRef]
- Gholami, M.; Etemadifar, Z.; Bouzari, M. Isolation a new strain of Kocuria rosea capable of tolerating extreme conditions. J. Environ. Radioact. 2015, 144, 113–119. [Google Scholar] [CrossRef]
- Bernal, C.; Cairo, J.; Coello, N. Purification and characterization of a novel exocellular keratinase from Kocuria rosea. Enzym. Microb. Technol. 2006, 38, 49–54. [Google Scholar] [CrossRef]
- Goswami, D.; Pithwa, S.; Dhandhukia, P.; Thakker, J.N. Delineating Kocuria turfanensis 2M4 as a credible PGPR: A novel IAA-producing bacteria isolated from saline desert. J. Plant Interact. 2014, 9, 566–576. [Google Scholar] [CrossRef]
- Nesheli, M.A.; Asgarani, E.; Dabbagh, R. Biosorption potential of Cr(VI) by Kocuria sp ASB107, a radio-resistant bacterium isolated from Ramsar, Iran. Chem. Ecol. 2018, 34, 163–176. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Koch, C.; Gvozdiak, O.; Schumann, P. Taxonomic Dissection Of The Genus Micrococcus-Kocuria Gen-Nov, Nesterenkonia Gen-Nov, Kytococcus Gen-Nov, Dermacoccus Gen-Nov, And Micrococcus Cohn 1872 Gen Emend. Int. J. Syst. Bacteriol. 1995, 45, 682–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dastager, S.G.; Tang, S.K.; Srinivasan, K.; Lee, J.C.; Li, W.J. Kocuria indica sp nov., isolated from a sediment sample. Int. J. Syst. Evol. Microbiol. 2014, 64, 869–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, M.S.; Wang, E.J.; Zimmermann, S.; Boutin, S.; Wagner, H.; Wink, M. Kocuria tytonicola, new bacteria from the preen glands of American barn owls (Tyto furcata). Syst. Appl. Microbiol. 2019, 42, 198–204. [Google Scholar] [CrossRef]
- Pridmore, D.; Rekhif, N.; Pittet, A.C.; Suri, B.; Mollet, B. Variacin, a new lanthionine-containing bacteriocin produced by Micrococcus varians: Comparison to lacticin 481 of Lactococcus lactis. Appl. Environ. Microbiol. 1996, 62, 1799–1802. [Google Scholar] [CrossRef] [Green Version]
- Sarafin, Y.; Birdilla, M.; Donio, S.; Velmurugan, S.; Michaelbabu, M.; Citarasu, T. Kocuria marina BS-15 a biosurfactant producing halophilic bacteria isolated from solar salt works in India. Saudi J. Biol. Sci. 2014, 21, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Budel, B.; Karsten, U.; GarciaPichel, F. Ultraviolet-absorbing scytonemin and mycosporine-like amino acid derivatives in exposed, rock-inhabiting cyanobacterial lichens. Oecologia 1997, 112, 165–172. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C. Mycosporine-like amino acids and related gadusols: Biosynthesis, accumulation, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 2002, 64, 223–262. [Google Scholar] [CrossRef] [Green Version]
- Dunlap, W.C.; Shick, J.M. Ultraviolet radiation-absorbing mycosporine-like amino acids in coral reef organisms: A biochemical and environmental perspective. J. Phycol. 1998, 34, 418–430. [Google Scholar] [CrossRef]
- Trincone, A. Potential biocatalysts originating from sea environments. J. Mol. Catal. B-Enzym. 2010, 66, 241–256. [Google Scholar] [CrossRef]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulzele, R.; DeSa, E.; Yadav, A.; Shouche, Y.; Bhadekar, R. Characterization of Novel Extracellular Protease Produced By Marine Bacterial Isolate from the Indian Ocean. Braz. J. Microbiol. 2011, 42, 1364–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, B.; Xu, Z.J.; Sun, Z.T.; Lin, J.; Hua, Y.J. Evaluation of the antioxidant effects of carotenoids from Deinococcus radiodurans through targeted mutagenesis, chemiluminescence, and DNA damage analyses. Biochim. Biophys. Acta-Gen. Subj. 2007, 1770, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Tristan, S.; Parra-Saldivar, R.; Iqbal, H.M.N.; Carrillo-Nieves, D. Bioinspired biomolecules: Mycosporine-like amino acids and scytonemin from Lyngbya sp. with UV-protection potentialities. J. Photochem. Photobiol. B-Biol. 2019, 201, 11. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, C.S.; Capper, E.A.; Roshak, A.K.; Marquez, B.; Eichman, C.; Jackson, J.R.; Mattern, M.; Gerwick, W.H.; Jacobs, R.S.; Marshall, L.A. The identification and characterization of the marine natural product scytonemin as a novel antiproliferative pharmacophore. J. Pharmacol. Exp. Ther. 2002, 303, 858–866. [Google Scholar] [CrossRef]
- Shukla, M.; Chaturvedi, R.; Tamhane, D.; Vyas, P.; Archana, G.; Apte, S.; Bandekar, J.; Desai, A. Multiple-stress tolerance of ionizing radiation-resistant bacterial isolates obtained from various habitats: Correlation between stresses. Curr. Microbiol. 2007, 54, 142–148. [Google Scholar] [CrossRef]
- Haidar, B.; Ferdous, M.; Fatema, B.; Ferdous, A.S.; Islam, M.R.; Khan, H. Population diversity of bacterial endophytes from jute (Corchorus olitorius) and evaluation of their potential role as bioinoculants. Microbiol. Res. 2018, 208, 43–53. [Google Scholar] [CrossRef]
- Retamal-Morales, G.; Mehnert, M.; Schwabe, R.; Tischler, D.; Zapata, C.; Chavez, R.; Schlomann, M.; Levican, G. Detection of arsenic-binding siderophores in arsenic-tolerating Actinobacteria by a modified CAS assay. Ecotoxicol. Environ. Saf. 2018, 157, 176–181. [Google Scholar] [CrossRef]
- Samanta, A.K.; Chaudhuri, S.; Dutta, D. Antioxidant efficacy of carotenoid extract from bacterial strain Kocuria marina DAGII. Mater. Today-Proc. 2016, 3, 3427–3433. [Google Scholar] [CrossRef]
- Mal, S.A.; Ibrahim, G.S.; Al Khalaf, M.I.; Al-Hejin, A.M.; Bataweel, N.M.; Abu-Zaid, M. Production and Partial Characterization of Yellow Pigment Produced by Kocuria flava Isolate and Testing its Antioxidant and Antimicrobial activity. Int. J. Life Sci. Pharma Res. 2020, 10, L58–L66. [Google Scholar] [CrossRef]
- Kumar, C.G.; Sujitha, P. Kocuran, an exopolysaccharide isolated from Kocuria rosea strain BS-1 and evaluation of its in vitro immunosuppression activities. Enzym. Microb. Technol. 2014, 55, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Palomo, S.; Gonzalez, I.; de la Cruz, M.; Martin, J.; Tormo, J.R.; Anderson, M.; Hill, R.T.; Vicente, F.; Reyes, F.; Genilloud, O. Sponge-Derived Kocuria and Micrococcus spp. as Sources of the New Thiazolyl Peptide Antibiotic Kocurin. Mar. Drugs 2013, 11, 1071–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezanka, T.; Gharwalova, L.; Novakova, G.; Kolouchova, I.; Uhlik, O.; Sigler, K. Kocuria Bacterial Isolates from Radioactive Springs of Jachymov spa (Joachimsthal) as Sources of Polyunsaturated Fatty Acids. Lipids 2019, 54, 177–187. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rezanka, T.; Rozentsvet, O.A. Lipid-Composition of 3 Macrophytes from the Caspian Sea. Phytochemistry 1993, 33, 1015–1019. [Google Scholar] [CrossRef]
- Vancura, A.; Rezanka, T.; Marslalek, J.; Melzoch, K.; Basarova, G.; Kristan, V. Metabolism of L-Threonine and Fatty-Acids and Tylosin Biosynthesis in Streptomyces-Fradiae. FEMS Microbiol. Lett. 1988, 49, 411–415. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and Characterization of Heterotrophic Microbial Communities on the Basis of Patterns of Community-Level Sole-Carbon-Source Utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal Chemical-Assay for the Detection and Determination of Siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Chick, H. An investigation of the laws of disinfection. J. Hyg. 1908, 8, 92–158. [Google Scholar] [CrossRef] [Green Version]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.B.; Nedashkovskaya, O.I.; Mikhailov, V.V.; Han, S.K.; Kim, K.O.; Rhee, M.S.; Bae, K.S. Kocuria marina sp. nov., a novel actinobacterium isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 2004, 54, 1617–1620. [Google Scholar] [CrossRef]
- Shivlata, L.; Satyanarayana, T. Thermophilic and alkaliphilic Actinobacteria: Biology and potential applications. Front. Microbiol. 2015, 6, 1015. [Google Scholar] [CrossRef] [PubMed]
- Russell, N.J.; Nichols, D.S. Polyunsaturated fatty acids in marine bacteria—A dogma rewritten. Microbiology 1999, 145, 767–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Close, D.; Nelson, W.; Bernhard, W. DNA Damage by the Direct Effect of Ionizing Radiation: Products Produced by Two Sequential One-Electron Oxidations. J. Phys. Chem. A 2013, 117, 12608–12615. [Google Scholar] [CrossRef]
- Pavlopoulou, A.; Savva, G.D.; Louka, M.; Bagos, P.G.; Vorgias, C.E.; Michalopoulos, I.; Georgakilas, A.G. Unraveling the mechanisms of extreme radioresistance in prokaryotes: Lessons from nature. Mutat. Res.-Rev. Mutat. Res. 2016, 767, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Leapman, R.D.; Lai, B.; Ravel, B.; Li, S.M.W.; Kemner, K.M.; et al. Protein oxidation implicated as the primary determinant of bacterial radioresistance. PLoS. Biol. 2007, 5, 769–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rich, T.; Allen, R.L.; Wyllie, A.H. Defying death after DNA damage. Nature 2000, 407, 777–783. [Google Scholar] [CrossRef]
- Rivas, M.; Rojas, E.; Calaf, G.M.; Barberan, M.; Liberman, C.; Correa, M.D. Association between non-melanoma and melanoma skin cancer rates, vitamin D and latitude. Oncol. Lett. 2017, 13, 3787–3792. [Google Scholar] [CrossRef] [PubMed]
- Gabani, P.; Singh, O.V. Radiation-resistant extremophiles and their potential in biotechnology and therapeutics. Appl. Microbiol. Biotechnol. 2013, 97, 993–1004. [Google Scholar] [CrossRef]
- Ben Ghorbal, S.K.; Chatti, A.; Sethom, M.M.; Maalej, L.; Mihoub, M.; Kefacha, S.; Feki, M.; Landoulsi, A.; Hassen, A. Changes in Membrane Fatty Acid Composition of Pseudomonas aeruginosa in Response to UV-C Radiations. Curr. Microbiol. 2013, 67, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Venkateswaran, A.; Hess, M.; Omelchenko, M.V.; Kostandarithes, H.M.; Makarova, K.S.; et al. Accumulation of Mn(II) in, Deinococcus radiodurans facilitates gamma-radiation resistance. Science 2004, 306, 1025–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cain, T.J.; Smith, A.T. Ferric iron reductases and their contribution to unicellular ferrous iron uptake. J. Inorg. Biochem. 2021, 218, 9. [Google Scholar] [CrossRef]
- Yoshida, K.; Hashimoto, M.; Hori, R.; Adachi, T.; Okuyama, H.; Orikasa, Y.; Nagamine, T.; Shimizu, S.; Ueno, A.; Morita, N. Bacterial Long-Chain Polyunsaturated Fatty Acids: Their Biosynthetic Genes, Functions, and Practical Use. Mar. Drugs 2016, 14, 23. [Google Scholar] [CrossRef]
- Fredrickson, J.K.; Li, S.-M.W.; Gaidamakova, E.K.; Matrosova, V.Y.; Zhai, M.; Sulloway, H.M.; Scholten, J.C.; Brown, M.G.; Balkwill, D.L.; Daly, M.J. Protein oxidation: Key to bacterial desiccation resistance? ISME J. 2008, 2, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Wang, L.L.; Chen, Y.J.; Long, Y. Optimization of staining with SYTO 9/propidium iodide: Interplay, kinetics and impact on Brevibacillus brevis. Biotechniques 2020, 69, 89. [Google Scholar] [CrossRef]
- Stocks, S.M. Mechanism and use of the commercially available viability stain, BacLight. Cytom. Part A 2004, 61A, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.; Burghardt, J.; Pradella, S.; Schumann, P.; Stackebrandt, E.; Marialigeti, K. Kocuria palustris sp. nov. and Kocuria rhizophila sp. nov., isolated from the rhizoplane of the narrow-leaved cattail (Typha angustifolia). Int. J. Syst. Bacteriol. 1999, 49, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Roman-Ponce, B.; Wang, D.; Vasquez-Murrieta, M.S.; Chen, W.F.; Estrada-de los Santos, P.; Sui, X.H.; Wang, E.T. Kocuria arsenatis sp nov., an arsenic-resistant endophytic actinobacterium associated with Prosopis laegivata grown on high-arsenic-polluted mine tailing. Int. J. Syst. Evol. Microbiol. 2016, 66, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.; Shibata, C.; Tamura, T.; Nurkanto, A.; Ratnakomala, S.; Lisdiyanti, P.; Suzuki, K. Kocuria pelophila sp nov., an actinobacterium isolated from the rhizosphere of a mangrove. Int. J. Syst. Evol. Microbiol. 2016, 66, 3276–3280. [Google Scholar] [CrossRef]
- Singh, O.V.; Gabani, P. Extremophiles: Radiation resistance microbial reserves and therapeutic implications. J. Appl. Microbiol. 2011, 110, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, C.S.; Capper, E.A.; Roshak, A.K.; Marquez, B.; Grace, K.; Gerwick, W.H.; Jacobs, R.S.; Marshall, L.A. Scytonemin—A marine natural product inhibitor of kinases key in hyperproliferative inflammatory diseases. Inflamm. Res. 2002, 51, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Lemee, L.; Peuchant, E.; Clerc, M.; Brunner, M.; Pfander, H. Deinoxanthin: A new carotenoid isolated from Deinococcus radiodurans. Tetrahedron 1997, 53, 919–926. [Google Scholar] [CrossRef]
- Shahmohammadi, H.R.; Asgarani, E.; Terato, H.; Saito, T.; Ohyama, Y.; Gekko, K.; Yamamoto, O.; Ide, H. Protective roles of bacterioruberin and intracellular KCl in the resistance of Halobacterium salinarium against DNA-damaging agents. J. Radiat. Res. 1998, 39, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Li, J.L.; Pang, R.J.; Dai, S.; Li, T.; Weng, Y.L.; Jin, Y.; Hua, Y.J. Gold Nanoparticles Biosynthesized and Functionalized Using a Hydroxylated Tetraterpenoid Trigger Gene Expression Changes and Apoptosis in Cancer Cells. ACS Appl. Mater. Interfaces 2018, 10, 37353–37363. [Google Scholar] [CrossRef]
- Santhanam, R.; Rong, X.Y.; Huang, Y.; Andrews, B.A.; Asenjo, J.A.; Goodfellow, M. Streptomyces bullii sp nov., isolated from a hyper-arid Atacama Desert soil. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2013, 103, 367–373. [Google Scholar] [CrossRef]
- Rateb, M.E.; Houssen, W.E.; Harrison, W.T.A.; Deng, H.; Okoro, C.K.; Asenjo, J.A.; Andrews, B.A.; Bull, A.T.; Goodfellow, M.; Ebel, R.; et al. Diverse Metabolic Profiles of a Streptomyces Strain Isolated from a Hyper-arid Environment. J. Nat. Prod. 2011, 74, 1965–1971. [Google Scholar] [CrossRef] [PubMed]
- Schulz, D.; Beese, P.; Ohlendorf, B.; Erhard, A.; Zinecker, H.; Dorador, C.; Imhoff, J.F. Abenquines A-D: Aminoquinone derivatives produced by Streptomyces sp strain DB634. J. Antibiot. 2011, 64, 763–768. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Proportion of (▨) saturated, (■) monounsaturated, and (□) polyunsaturated fatty acids in Kocuria sp. 101 (A), 208 (B), 301 (C) and 401 (D) at different cultivation temperatures.
Figure 1.
Proportion of (▨) saturated, (■) monounsaturated, and (□) polyunsaturated fatty acids in Kocuria sp. 101 (A), 208 (B), 301 (C) and 401 (D) at different cultivation temperatures.

Figure 2.
Kinetics of Kocuria sp. isolates reduction according to the Chick–Watson model. N0 stands for the number of viable cultivable non-irradiated bacteria before UV-C exposure, N is the number of viable cultivable bacteria after a certain UV-C dose exposure. Symbols used in the graph correlate with: ![Fermentation 08 00035 i001]() E. coli CCM 4517,
E. coli CCM 4517, ![Fermentation 08 00035 i002]() D. radiodurans CCM 1700T,
D. radiodurans CCM 1700T, ![Fermentation 08 00035 i003]() isolate 101,
isolate 101, ![Fermentation 08 00035 i004]() isolate 208,
isolate 208, ![Fermentation 08 00035 i005]() isolate 301,
isolate 301, ![Fermentation 08 00035 i006]() isolate 401.
isolate 401.
 E. coli CCM 4517,
E. coli CCM 4517,  D. radiodurans CCM 1700T,
D. radiodurans CCM 1700T,  isolate 101,
isolate 101,  isolate 208,
isolate 208,  isolate 301,
isolate 301,  isolate 401.
isolate 401.
Figure 2.
Kinetics of Kocuria sp. isolates reduction according to the Chick–Watson model. N0 stands for the number of viable cultivable non-irradiated bacteria before UV-C exposure, N is the number of viable cultivable bacteria after a certain UV-C dose exposure. Symbols used in the graph correlate with: ![Fermentation 08 00035 i001]() E. coli CCM 4517,
E. coli CCM 4517, ![Fermentation 08 00035 i002]() D. radiodurans CCM 1700T,
D. radiodurans CCM 1700T, ![Fermentation 08 00035 i003]() isolate 101,
isolate 101, ![Fermentation 08 00035 i004]() isolate 208,
isolate 208, ![Fermentation 08 00035 i005]() isolate 301,
isolate 301, ![Fermentation 08 00035 i006]() isolate 401.
isolate 401.
E. coli CCM 4517, D. radiodurans CCM 1700T, isolate 101, isolate 208, isolate 301, isolate 401.
Figure 3.
Survival diagram of Kocuria sp. 101, 208, 301, and 401 after 30-day desiccation.

Figure 4.
Substrate average well color development (AWCD) of Kocuria sp. isolate 101 (○), isolate 208 (△), isolate 301 (▲), and isolate 401 (□).
Figure 4.
Substrate average well color development (AWCD) of Kocuria sp. isolate 101 (○), isolate 208 (△), isolate 301 (▲), and isolate 401 (□).

Figure 5.
Well color development of individual groups of substrates for Kocuria sp. 101 (A), 208 (B), 301 (C), and 401 (D) of alcohols (○), amines (△), amino acids (▲), carboxylic acids (□), esters (■), hydrocarbons (●), polymers (♦).
Figure 5.
Well color development of individual groups of substrates for Kocuria sp. 101 (A), 208 (B), 301 (C), and 401 (D) of alcohols (○), amines (△), amino acids (▲), carboxylic acids (□), esters (■), hydrocarbons (●), polymers (♦).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of different temperatures on specific growth rate and maximum OD600 of isolated Kocuria sp. strains.
Table 1.
Effect of different temperatures on specific growth rate and maximum OD600 of isolated Kocuria sp. strains.
| Temperature (°C) | Kocuria sp. 101 | Kocuria sp. 208 | Kocuria sp. 301 | Kocuria sp. 401 | |
|---|---|---|---|---|---|
| 5 | Specific growth rate (h−1) OD600 max | 0.013 ± 0.005 | - | 0.013 ± 0.002 | - |
| 10 | Specific growth rate (h−1) | 0.021 ± 0.004 | - | 0.020 ± 0.005 | - |
| OD600 max | 13.5 | 10.8 | |||
| 20 | Specific growth rate (h−1) | 0.070 ± 0.010 | 0.068 ± 0.009 | 0.074 ± 0.002 | 0.083 ± 0.006 |
| OD600 max | 18.9 | 15.3 | 15.5 | 12.6 | |
| 30 | Specific growth rate (h−1) | 0.066 ± 0.008 | 0.080 ± 0.012 | 0.072 ± 0.009 | 0.085 ± 0.013 |
| OD600 max | 19.1 | 14.9 | 15.7 | 12.6 | |
| 40 | Specific growth rate (h−1) | 0.042 ± 0.007 | 0.064 ± 0.011 | 0.053 ± 0.002 | 0.061 ± 0.005 |
| OD600 max | 6.8 | 7.5 | 6.0 | 4.9 | |
| 45 | Specific growth rate (h−1) | 0.021 ± 0.006 | 0.031 ± 0.007 | 0.025 ± 0.002 | 0.020 ± 0.008 |
| OD600 max | 6.3 | 6.2 | 5.7 | 4.0 |
Table 2.
Polyunsaturated fatty acid content at 20 °C, siderophore and antioxidant production of four Kocuria sp. isolates from Jachymov radon springs.
Table 2.
Polyunsaturated fatty acid content at 20 °C, siderophore and antioxidant production of four Kocuria sp. isolates from Jachymov radon springs.
| Siderophore Production | Antioxidant Activity (mgAA/L) | C20:4 ω-6 (% of Total Fatty Acid) | C20:5 ω-3 (% of Total Fatty Acid) | |
|---|---|---|---|---|
| Kocuria sp. 101 | + | 4.1 ± 1.3 | 0.31 ± 0.09 | 2.21 ± 0.12 |
| Kocuria sp. 208 | + | 4.2 ± 0.3 | 1.01 ± 0.13 | 2.11 ± 0.17 |
| Kocuria sp. 301 | + | 8.9 ± 0.6 | 1.21 ± 0.17 | 3.71 ± 0.13 |
| Kocuria sp. 401 | + | 5.4 ± 0.4 | 1.11 ± 0.08 | 3.31 ± 0.22 |
Mean ± S.D. from three measurements, i.e., relative %.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Timkina, E.; Drábová, L.; Palyzová, A.; Řezanka, T.; Maťátková, O.; Kolouchová, I. Kocuria Strains from Unique Radon Spring Water from Jachymov Spa. Fermentation 2022, 8, 35. https://doi.org/10.3390/fermentation8010035
AMA Style
Timkina E, Drábová L, Palyzová A, Řezanka T, Maťátková O, Kolouchová I. Kocuria Strains from Unique Radon Spring Water from Jachymov Spa. Fermentation. 2022; 8(1):35. https://doi.org/10.3390/fermentation8010035
Chicago/Turabian StyleTimkina, Elizaveta, Lucie Drábová, Andrea Palyzová, Tomáš Řezanka, Olga Maťátková, and Irena Kolouchová. 2022. "Kocuria Strains from Unique Radon Spring Water from Jachymov Spa" Fermentation 8, no. 1: 35. https://doi.org/10.3390/fermentation8010035
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.