3.1. LFQ Analysis of Rat Spleen Lymphocytes after 7-Day Treatment with Morphine, LYS739, and LYS744: The Dose-Effect of 3 mg/kg
The study had some limitations. The first one is that we had to work with small amounts of peptides
LYS739 and
LYS744. For that reason, each testing group included only three of four animals. Second, this study was not designed to search for differences between male and female rats and only included male rats as in our previous proteomic analyses [
13,
14,
15,
18]. The third one is connected with mass spectrometry-based data. Some measurements may result in technical failures. Therefore, three technical replicates of each sample enabled us to perform statistical analysis. Label-free quantification (LFQ) analysis revealed 27 altered proteins in rat spleen lymphocytes after 7-day morphine treatment (3 mg/kg). The minimum of 2-fold expression differences were calculated for at least 2 measured values from 3 technical replicates,
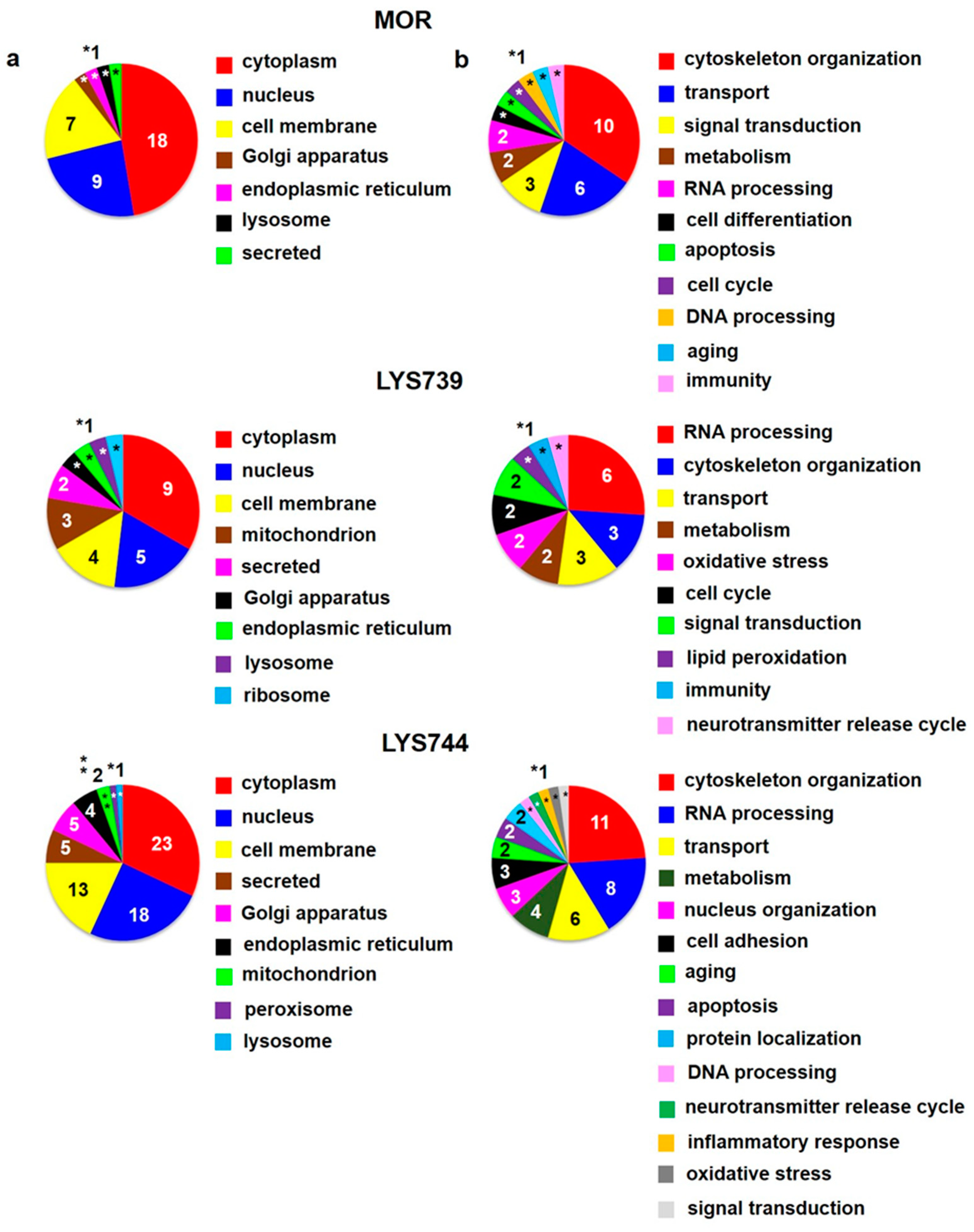
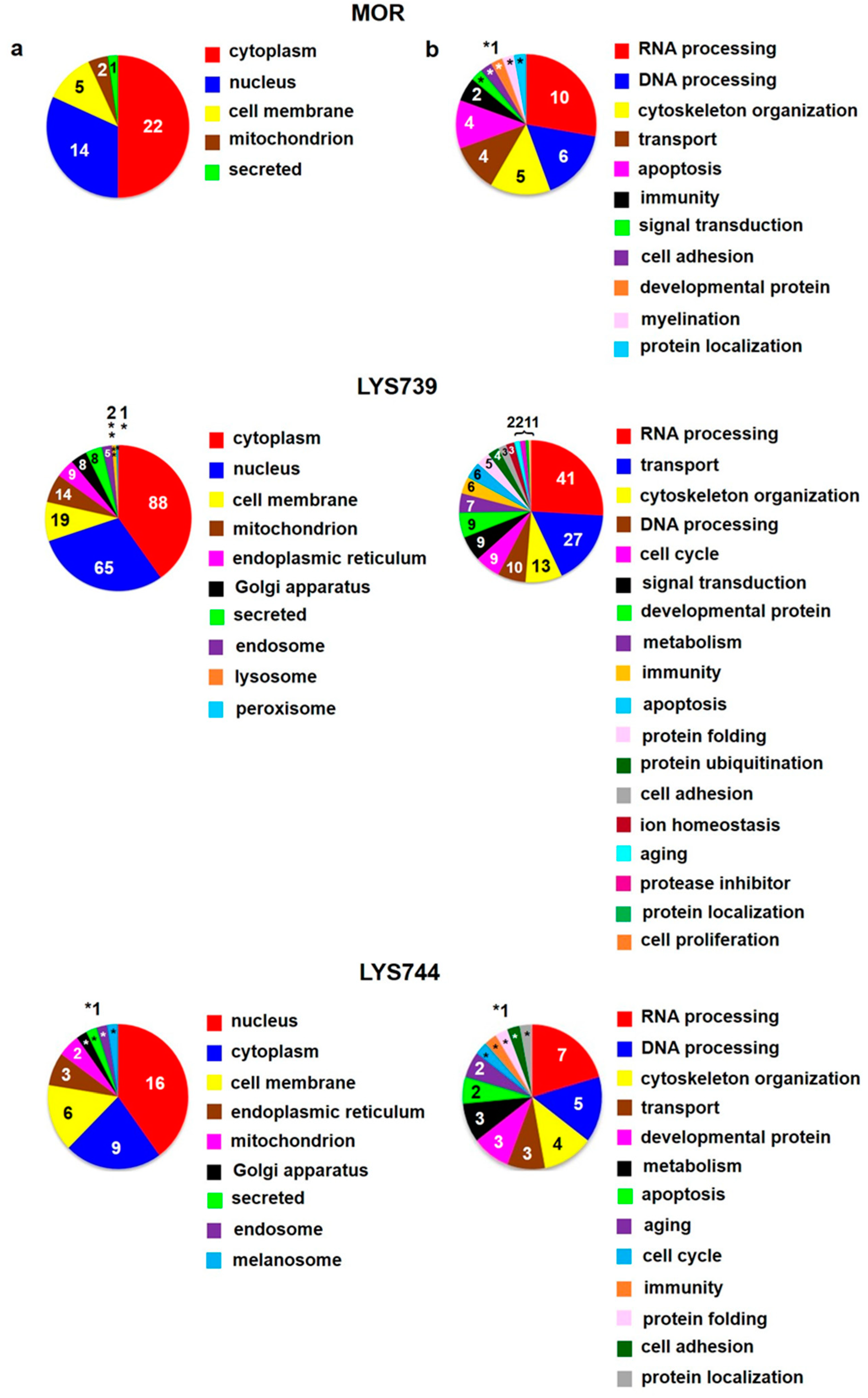
Table S1a. The 12 proteins were upregulated, and 15 proteins were downregulated. Their subcellular localization and functional significance unveiled most proteins of cytoplasmic origin related to the cytoskeletal changes,
Figure 1a,b (upper panels). These proteins were associated with actin filament bundle assembly: beta-adducin (
Add2, ↑4.0-fold), spectrin beta chain (
Sptbn2, ↑3.9-fold); microtubule polymerization: tubulin polymerization-promoting protein family member 3 (Tppp3, ↑2.6-fold), tubulin beta-3 chain (
Tubb3, ↓5.1-fold), or intermediate filament organization: vimentin (
Vim, ↓2.8-fold),
Table S1a.
Proteomic analysis indicated that 7-day treatment with 3 mg/kg of
LYS739 altered the expression level of 20 proteins more than twice, 5 were upregulated and 15 downregulated,
Table S1a. Among these, 6 proteins were involved in RNA processing: H/ACA ribonucleoprotein complex subunit 4 (
Dkc1, ↑2.2-fold), hepatoma-derived growth factor-related protein 2 (
Hdgfrp2, ↑2.0-fold), treacle ribosome biogenesis factor 1 (
Tcof1, ↓3.0-fold), 60S acidic ribosomal protein P2 (
Rplp2, ↓2.6-fold), serine/arginine repetitive matrix 2 (
Srrm2, ↓2.6-fold), and CREB-regulated transcription coactivator 2 (
Crtc2, ↓2.0-fold). Modulation of transcription factors (TF), such as cyclic AMP response element binding protein (CREB) may represent a potential mechanism for persisted opioid-induced plasticity in the brain [
21]. According to Algmandi and Alshehri [
22], morphine administration was accompanied by a reduced mRNA expression level of CREB in the nucleus accumbens. Other functional categories associated with the identified proteins were organization of the cytoskeleton (3), transport (3), metabolism (2), oxidative stress (2), cell cycle (2), signal transduction (2), lipid peroxidation (1), immunity (1), and neurotransmitter release cycle (1),
Figure 1a,b (middle panels).
LYS744 had the most significant effect on the change of protein expression levels in rat spleen lymphocytes. LFQ analysis detected 45 altered proteins: 29 proteins upregulated more than 2-fold and 16 proteins downregulated,
Table S1a. More than half of these were related to actin cytoskeleton organization (11), RNA processing (8), and transport (6),
Figure 1a,b (lower panels). DNA helicase (
Chd4, ↑3.8-fold) was found to be the most significantly upregulated, while globin c2 (
Hba-a2, ↓11.6-fold) the most decreased. DNA helicases play key roles in DNA replication, transcription, DNA repair, and many other processes to maintain genomic integrity and cellular homeostasis [
23]. Helicase variants are frequently upregulated in cancerous tissues [
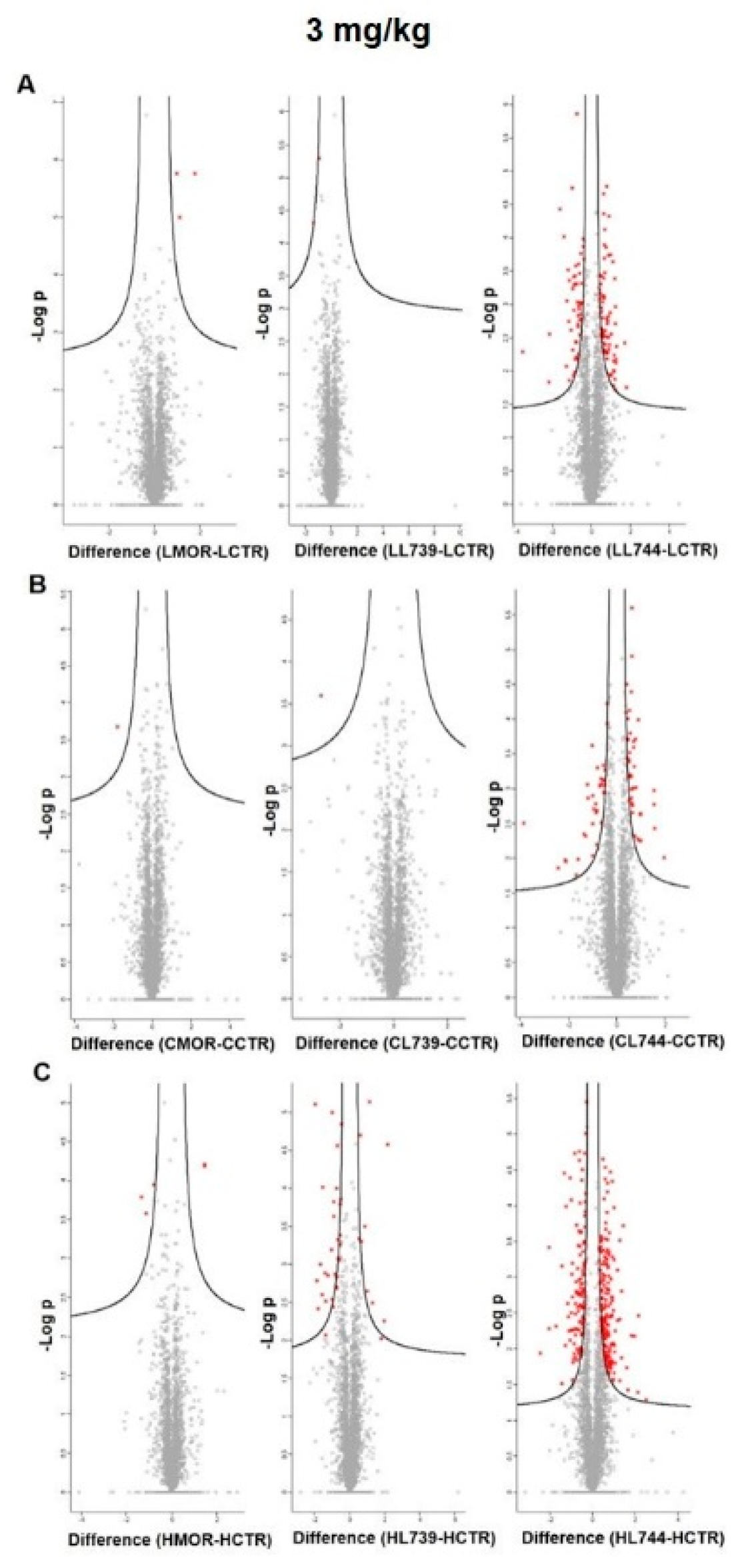
24]. Volcano plots representing significantly altered proteins identified in rat spleen lymphocytes after treatment with 3 mg/kg of morphine,
LYS739, and
LYS744 are depicted in
Figure 2A. Hierarchical heatmap clustering of all identified protein expression profiles in rat spleen lymphocytes is presented in
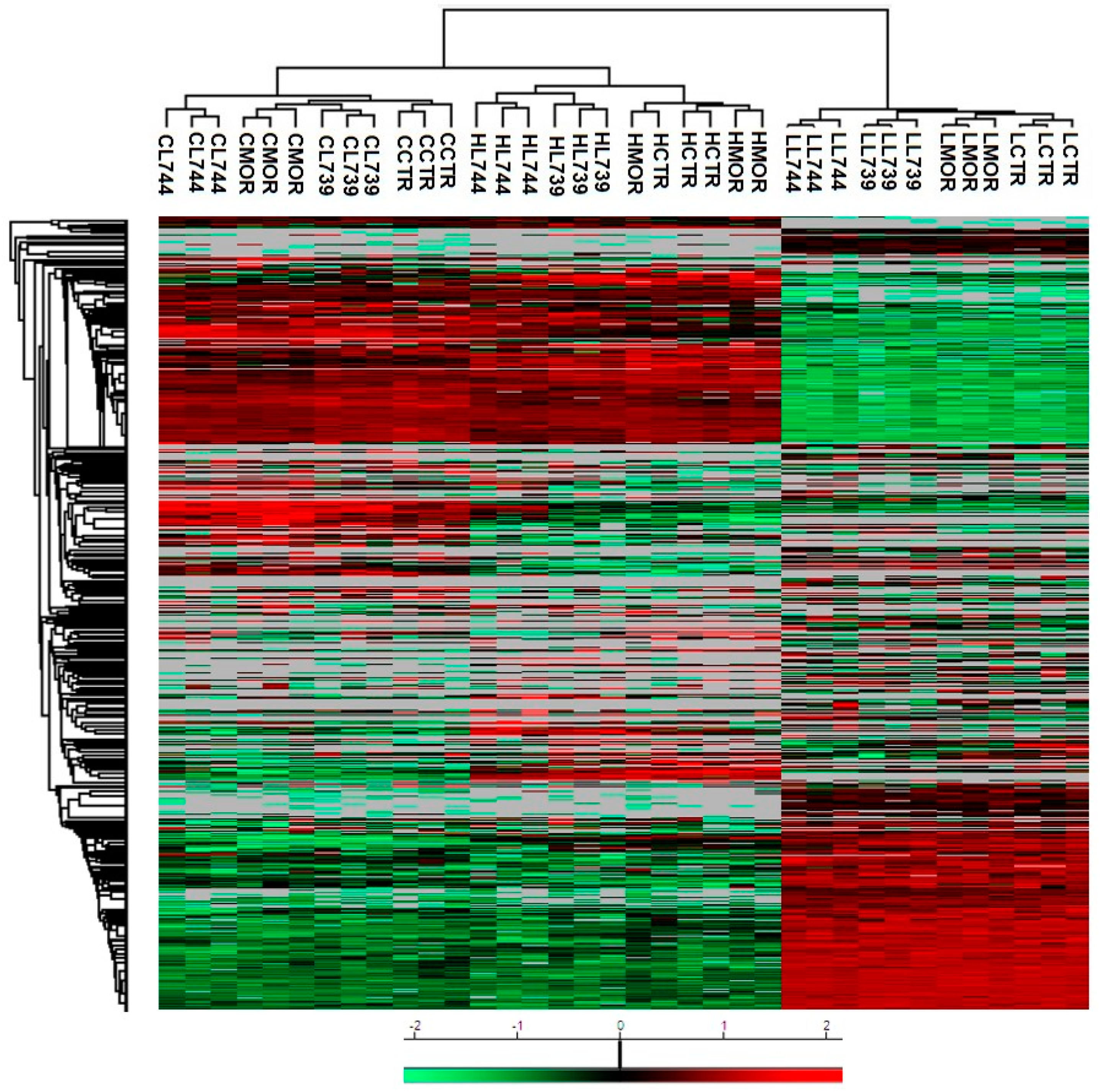
Figure 3.
3.2. LFQ Analysis of Rat Brain Cortex after 7-Day Treatment with Morphine, LYS739, and LYS744: The Dose-Effect of 3 mg/kg
The effect of 7-day morphine treatment (3 mg/kg) on protein alterations in the rat brain cortex was slightly less than that detected in the proteome profile of spleen lymphocytes. In cortical samples, LFQ analysis detected 5 proteins significantly upregulated and 10 proteins were downregulated,
Table S1b. From the functional point of view, 3 proteins were involved in RNA processing: U1 small nuclear ribonucleoprotein 70 kDa (
Snrnp70, ↓3.5-fold), DAZ-associated protein 1 (
Dazap1, ↓2.3-fold), nucleolar RNA helicase 2 (
Ddx21, ↓2.2-fold), 3 proteins participated in oxygen and protein transport: globin c2 (
Hba-a2, ↓13.6-fold), exportin 5 (
Xpo5, ↓2.4-fold), alpha globin (
Hba-a3, ↓2.2-fold), and 3 proteins were related to apoptosis: ribosomal protein S6 kinase (
Rps6ka3, ↑2.7-fold), protein S100-A9 (
S100a9, ↓3.7-fold), programmed cell death protein 4 (
Pdcd4, ↓2.3-fold).
S100a9 is a member of the S100 family proteins representing essential factors in inflammation and apoptosis-inducing activities [
25]. Decreased levels of
S100a9 and
Pdcd4 may suggest the activation of compensatory defense mechanisms, as chronic morphine treatment was related to a strong upregulation of the pro-apoptotic Fas receptor and a moderate downregulation of the anti-apoptotic Bcl-2 [
26]. The other functional categories were associated with immunity (3), metabolism (2), signal transduction (2), cell adhesion (1), cytoskeleton organization (1), and protein ubiquitination (1),
Figure 4a,b (upper panels).
LYS739 treatment resulted in 20 altered proteins (7 upregulated, 13 downregulated), mainly involved in RNA processing: polypyrimidine tract-binding protein 1 (
Ptbp1, ↓8.4-fold), U1 small nuclear ribonucleoprotein 70 kDa (
Snrnp70, ↓6.5-fold), heterogeneous nuclear ribonucleoprotein U-like 2 (
Hnrnpul2, ↓5.4-fold), probable ATP-dependent RNA helicase DDX46 (
Ddx46, ↓4.2-fold), nucleolar RNA helicase 2 (
Ddx21, ↓3.2-fold), splicing factor 3b, subunit 1 (
Sf3b1, ↓2.2-fold), and small nuclear ribonucleoprotein Sm D2 (
Snrpd2, ↓2.0-fold). Six cytoskeletal proteins were found to differ significantly: filamin A (
Flna, ↑3.5-fold), prefoldin 1 (
Pfdn1, ↑2.4-fold), formin-like 1 (
Fmnl1, ↑2.3-fold), protein S100-A9 (
S100a9, ↓10.5-fold), IQ motif-containing GTPase-activating protein 1 (
Iqgap1, ↓5.2-fold), and CD2-associated protein (
Cd2ap, ↓4.7-fold),
Table S1b,
Figure 4a,b (middle panels).
As in the case of spleen lymphocytes, treatment with
LYS744 identified the highest number of alterations in the brain cortex. Among 31 proteins, 11 were upregulated, and 20 proteins had the decreased expression levels,
Table S1b. LFQ analysis identified proteins related to the regulation of RNA processes (7), regulation of cell cycle (4), immunity (4), apoptotic processes (3), reorganization of the cytoskeleton (2), protein transport (2), signal transduction (2), brain development, (1) or protein folding (1),
Figure 4a,b (lower panels).
Flna (↑3.9-fold),
Pfdn1 (↑2.6-fold),
Ptbp1 (↓14.4-fold),
S100a9 (↓5.4-fold),
Snrnp70 (↓4.4-fold),
Cd2ap (↓3.2-fold),
Hba-a2 (↓2.6-fold),
Dazap1 (↓2.4-fold), and
Ddx21 (↓2.0-fold) were found dysregulated after morphine administration and
LYS739 treatment as well, see above. Volcano plots representing significantly altered proteins identified in rat brain cortex after treatment with 3 mg/kg of morphine,
LYS739, and
LYS744 are depicted in
Figure 2B. Hierarchical heatmap clustering of all identified protein expression profiles in the rat brain cortex is presented in
Figure 3.
3.3. LFQ Analysis of Rat Hippocampus after 7-Day Treatment with Morphine, LYS739, and LYS744: The Dose-Effect of 3 mg/kg
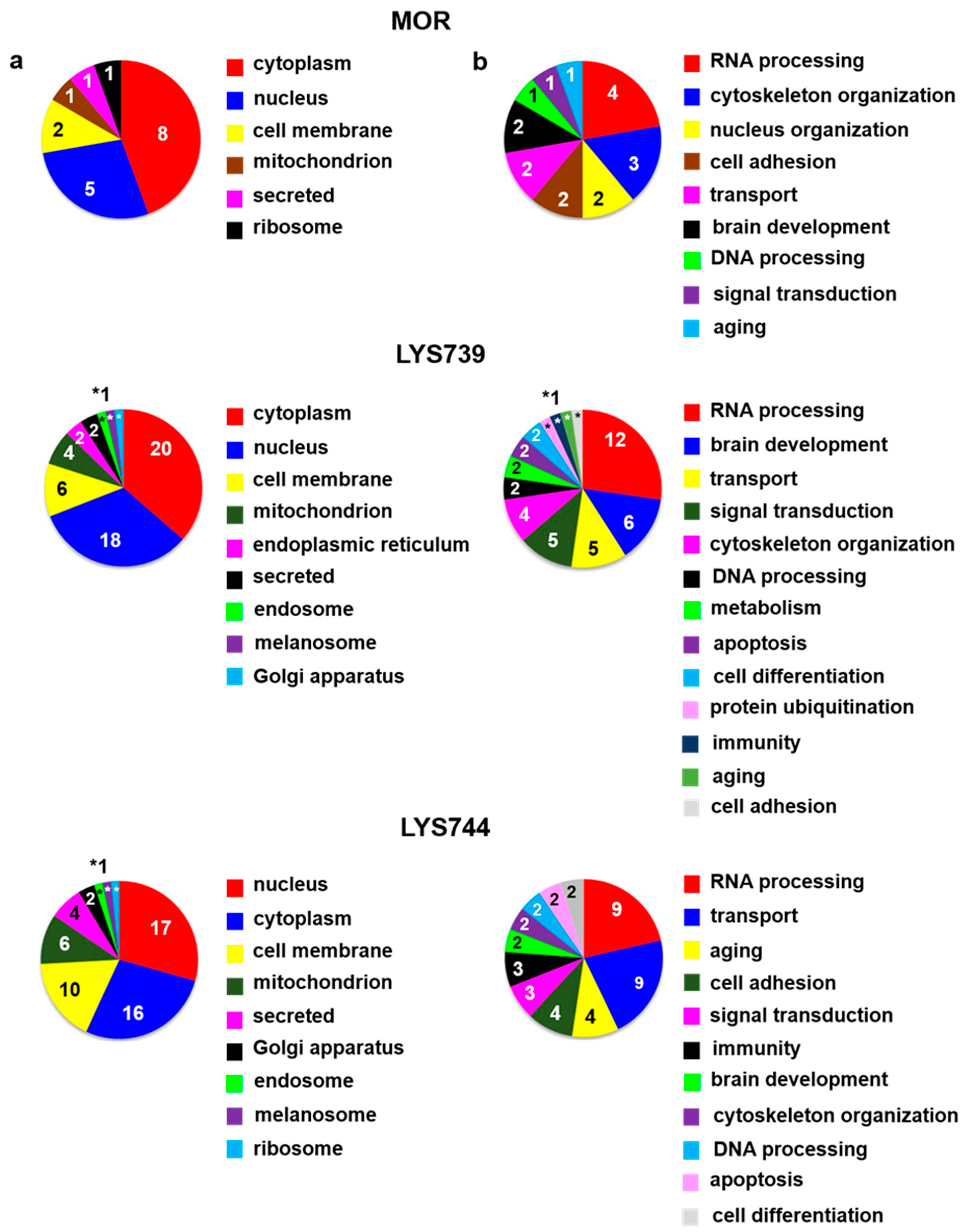
LFQ analysis indicated 14 proteins with changed expression levels at least 2-fold after 7-day treatment with morphine. This amount is similar to the number of alterations in the rat brain cortex (15) but lower twice in comparison with spleen lymphocytes (27). Eight proteins were upregulated, 6 had decreased levels,
Table S1c. According to the current annotations in the Uniprot database, these proteins were mainly associated with the regulation of transcription and translation: methyl-CpG-binding protein 2 (
Mecp2, ↑2.6-fold), general transcription factor II-I (
Gtf2i, ↑2.2-fold), CXXC-type zinc finger protein 1 (
Cxxc1, ↑2.2-fold), 60S ribosomal protein L29 (
Rpl29, ↑2.0-fold), and cytoskeletal changes: vasodilator-stimulated phosphoprotein (
Vasp, ↑4.1-fold), transgelin (
Tagln, ↑2.7-fold), alpha-internexin (
Ina, ↓2.2-fold),
Figure 5a,b (upper panels). Interestingly, in our previous study, the level of
Ina was detected as hypophosphorylated (↓2.3-fold) in rat hippocampal samples after 3-month morphine withdrawal [
15].
The effect of
LYS739 on rat hippocampal protein profile was higher than that of morphine treatment; proteomic analysis detected 36 altered proteins (8 upregulated, 28 downregulated), as shown in
Table S1c. Among different biological processes, most proteins were involved in RNA processing (12), brain development (6), transport (5), and signal transduction (5),
Figure 5a,b (middle panels). Notably, all proteins involved in brain development were decreased: cell cycle exit and neuronal differentiation protein 1 (
Cend1, ↓3.9-fold), neurofilament heavy polypeptide (
Nefh, ↓3.7-fold), metallothionein-3 (
Mt3, ↓3.5-fold),
Ina (↓2.0-fold), plasma membrane calcium-transporting ATPase 4 (
Atp2b4, ↓2.0-fold), myristoylated alanine-rich C-kinase substrate (
Marcks, ↓2.0-fold). Opioid drugs were reported to change hippocampal plasticity and inhibit adult neurogenesis [
27]. Contrarily, opioid withdrawal was connected with increased neuronal plasticity [
28,
29].
Treatment with
LYS744 revealed 42 alterations in protein expression levels, which is more than in the brain cortex (31) and similar to spleen lymphocytes (45). Twenty-four proteins were upregulated and 18 proteins were downregulated,
Table S1c. RNA processing (9) and transport (9) were the main functional categories,
Figure 5a,b (lower panels).
Mecp2 (↑2.6-fold) and
Rpl29 (↑2.0-fold) were found significantly upregulated after morphine treatment as well, see above. Among upregulated proteins, four of these were associated with the change in energy metabolism: electron transfer flavoprotein-ubiquinone oxidoreductase, mitochondrial (
Etfdh, ↑2.6-fold), ATPase inhibitor, mitochondrial (
Atpif,↑2.4-fold), complex I-B17 (
Ndufb6, ↑2.4-fold), and cytochrome c oxidase subunit 2 (
Mtco2, ↑2.3-fold). We previously reported that the majority of altered hippocampal proteins were related to energy metabolism after both chronic morphine treatment (10–50 mg/kg, 10 days) and subsequent drug withdrawal (3 weeks, 3 months, 6 months) [
14,
15,
16]. Volcano plots representing significantly altered proteins identified in rat hippocampus after treatment with 3 mg/kg of morphine,
LYS739, and
LYS744 are depicted in
Figure 2C. Hierarchical heatmap clustering of all identified protein expression profiles in rat hippocampus is presented in
Figure 3.
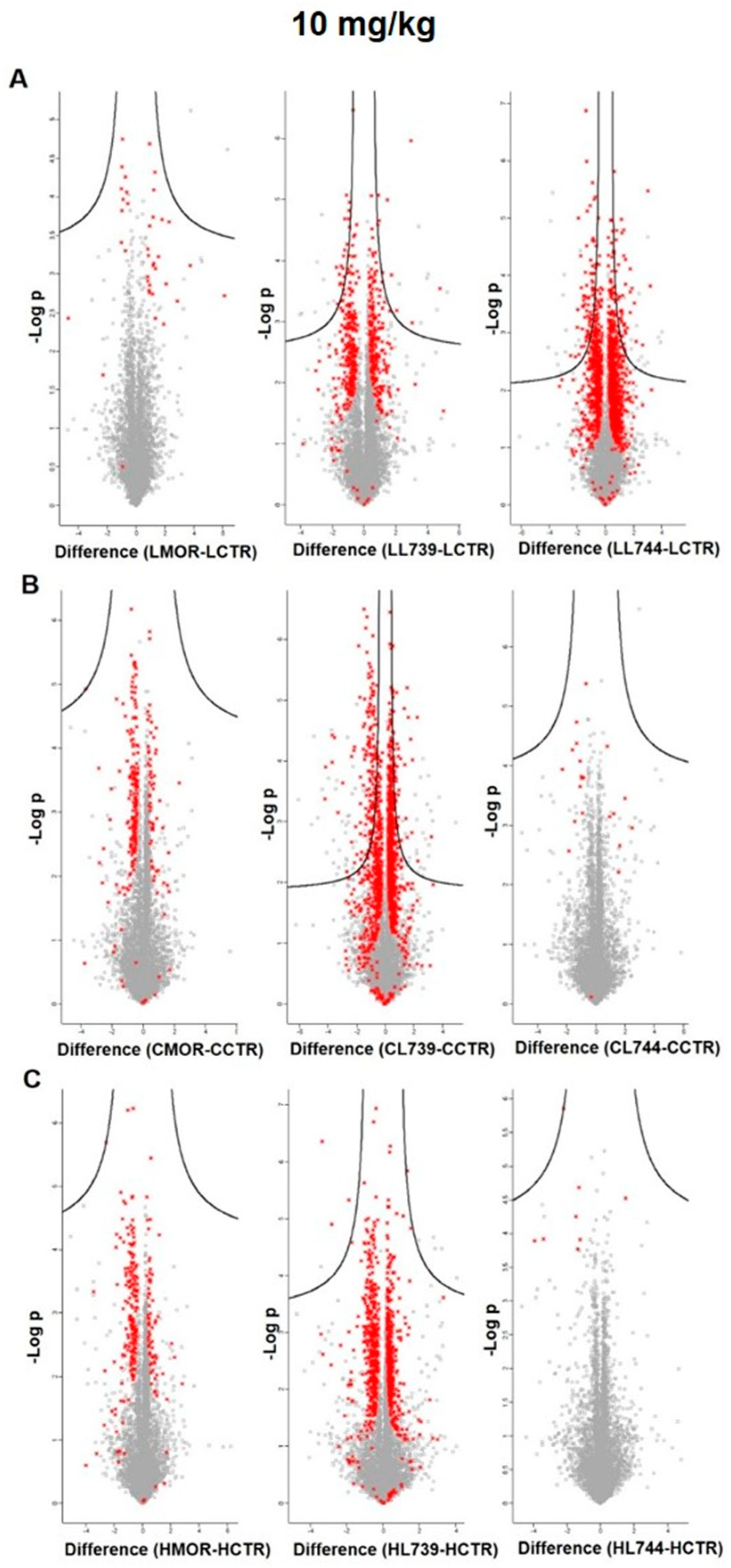
3.4. LFQ Analysis of Rat Spleen Lymphocytes after 7-Day Treatment with Morphine, LYS739, and LYS744: The Dose-Effect of 10 mg/kg
In the second round of our experiments, we increased the dose of morphine and enkephalin analogs from 3 mg/kg to 10 mg/kg. We applied this amount of OR drugs to rats for 7 consecutive days to test the effect of increased doses on protein changes in spleen lymphocytes, cortex, and hippocampus. This scheme was modified according to our previously established protocols [
4,
5,
13,
14,
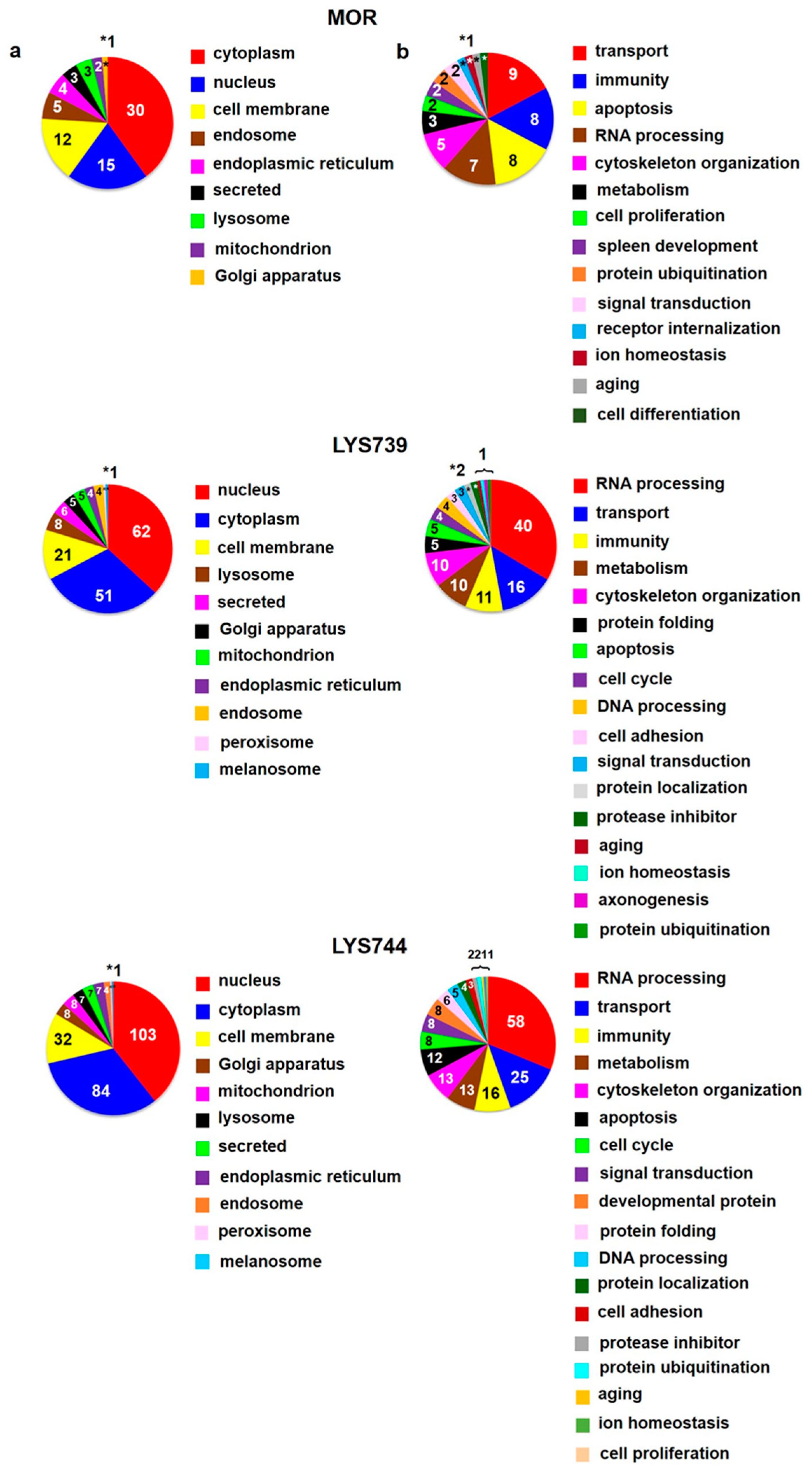
18]. LFQ analysis revealed 45 altered proteins (at least 2-fold) in spleen lymphocytes after morphine treatment. Upregulation was found for 29 proteins; 16 proteins were downregulated,
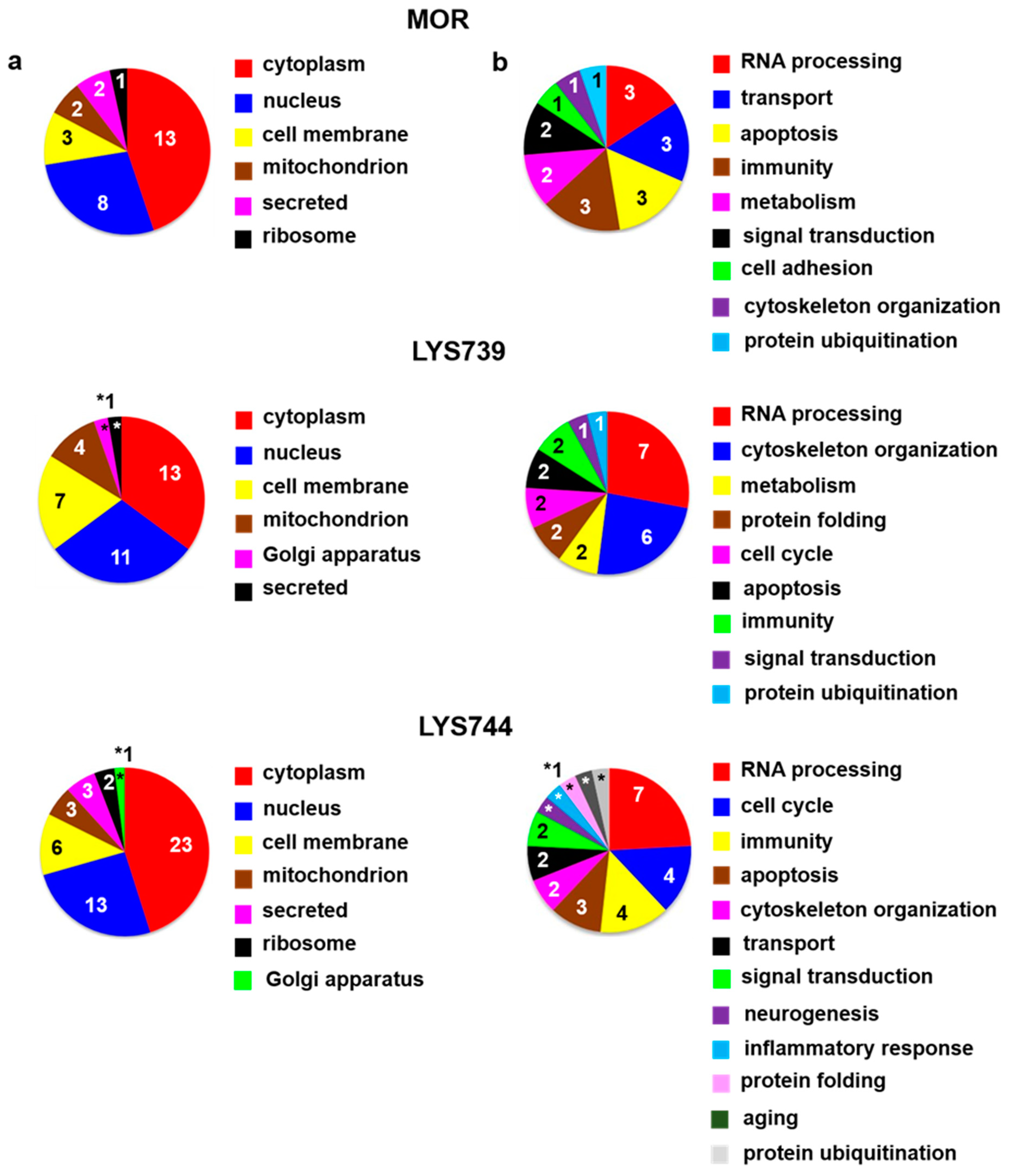
Table S2a. According to the current annotations in the Uniprot database, their molecular functions and biological processes were mostly involved in transport (9), immunity (8), apoptosis (8), and RNA processing (7),
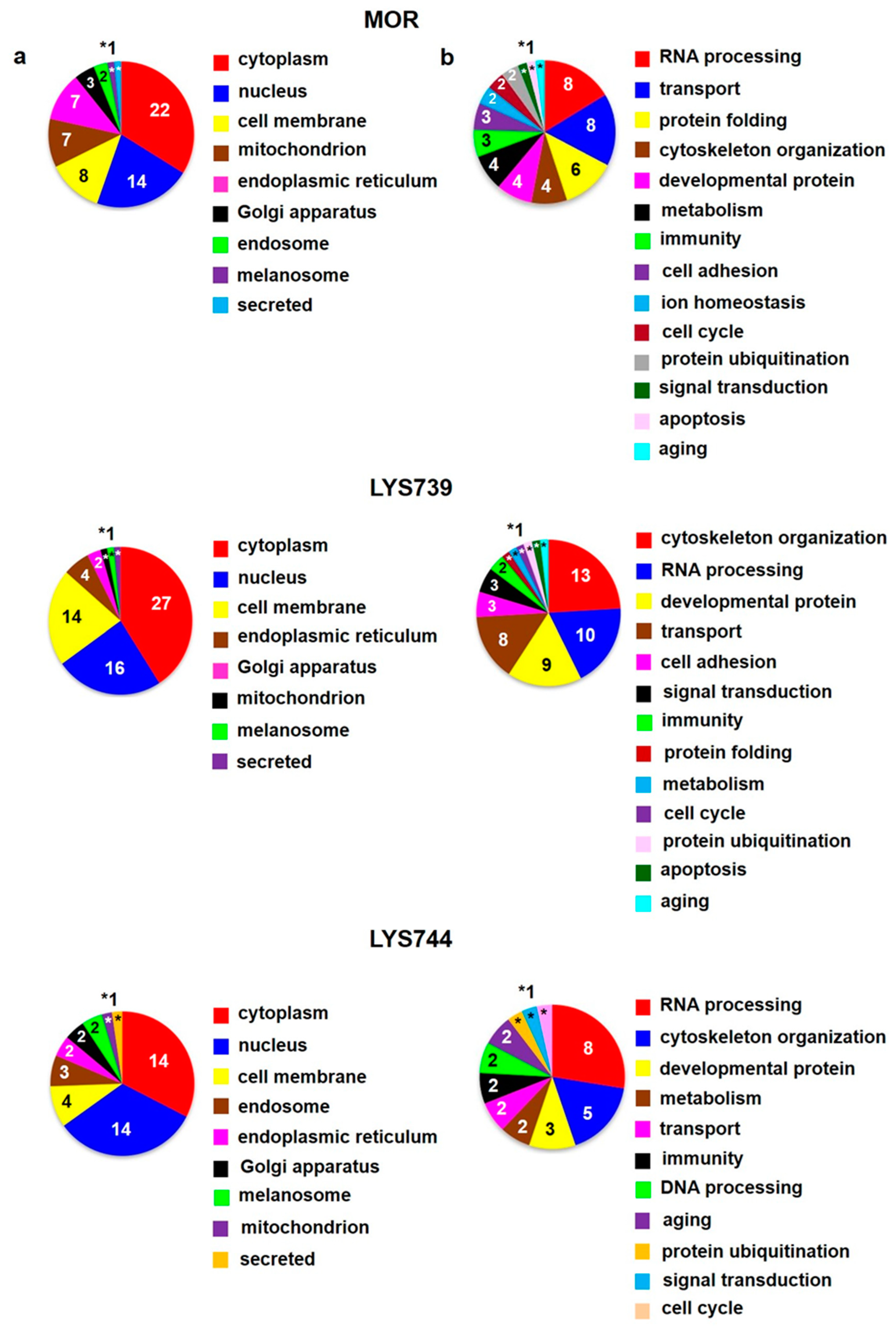
Figure 6a,b (upper panels). When compared with the effect of a lower dose, the number of proteins associated with immune responses and apoptotic processes was increased, suggesting the state of cell dysfunction and neuroinflammation after chronic morphine administration. Among upregulated apoptotic proteins were interferon-induced protein with tetratricopeptide repeats 3 (
Ifit3, ↑4.9-fold), sialic acid-binding Ig-like lectin 1 (
Siglec1, ↑3.8-fold), interferon-induced protein with tetratricopeptide repeats 2 (
Ifit2, ↑3.4-fold), MAP kinase-activating death domain protein (
Madd, ↑2.8-fold), galectin-5 (
Lgals5, ↑2.3-fold), and interferon activated gene 204 (
Mnda, ↑2.2-fold).
LYS739 treatment resulted in the change of 119 proteins (51 proteins were upregulated, 68 downregulated). The number of altered proteins was six times higher than after the dose of 3 mg/kg,
Table S2a. The majority of identified proteins were related to RNA processing (40), transport (16), immunity (11), metabolism (10), and cytoskeletal changes (10),
Figure 6a,b (middle panels). Seven proteins that participated in immune responses were upregulated: Ig gamma-2C chain C region (
N/A, ↑27.5-fold), proto-oncogene vav (
Vav1, ↑9.5-fold), tyrosine-protein kinase (
Btk, ↑3.4-fold),
Mnda (↑2.8-fold), macrophage-expressed gene 1 protein (
Mpeg1, ↑2.4-fold), tap-binding protein (
Tapbp, ↑2.3-fold), TNF receptor-associated factor 6 (
Traf6, ↑2.2-fold).
The treatment with
LYS744 caused the most significant differences in protein expression levels of spleen lymphocytes. Four times more proteins were altered (182 proteins, 88 upregulated, 94 downregulated) in comparison with the lower dose of
LYS744,
Table S2a. Identified proteins were primarily associated with RNA processing (58), transport (25), immune responses (16), metabolic changes (13), cytoskeletal changes (13), and apoptotic processes (12),
Figure 6 a,b (lower panels).
Vav1 (↑3.7-fold),
Mnda (↑3.4-fold),
Mpeg1 (↑2.6-fold), and
Btk (↑2.5-fold) were identified in samples treated with LYS739 as well, see above. Both immunosuppressive and immuno-stimulating effects have been described after opioid administration [
30,
31,
32]. Recent studies have shown that
Vav proteins are essential for the homeostasis of the central nervous, cardiovascular, and immune systems. They are considered potential therapeutic targets for several pathological conditions [
33,
34]. Volcano plots representing significantly altered proteins identified in rat spleen lymphocytes after treatment with 10 mg/kg of morphine,
LYS739, and
LYS744 are depicted in
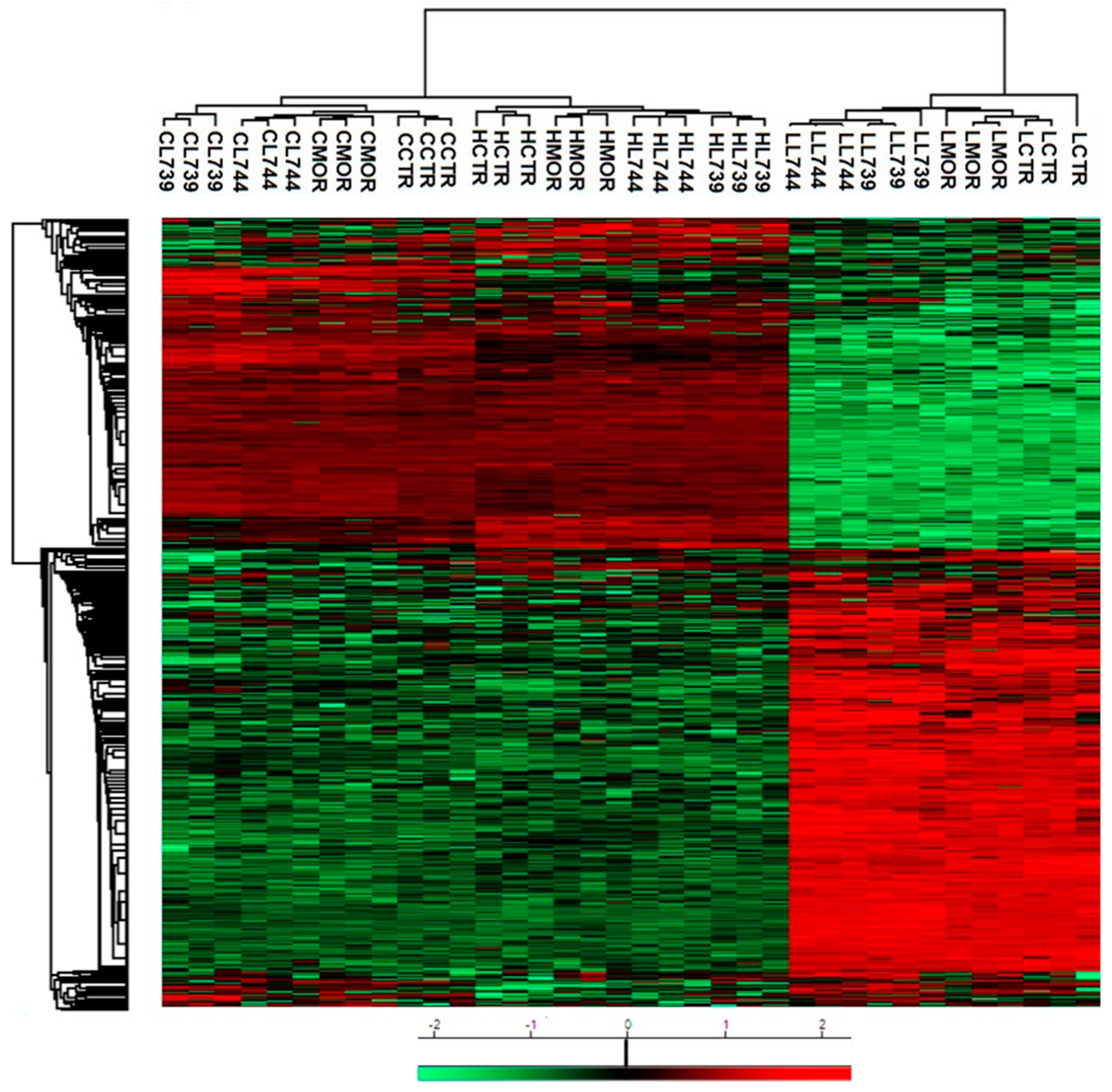
Figure 7A. Hierarchical heatmap clustering of all identified protein expression profiles in rat spleen lymphocytes is presented in
Figure 8.
3.5. LFQ Analysis of Rat Brain Cortex after 7-Day Treatment with Morphine, LYS739, and LYS744: The Dose-Effect of 10 mg/kg
The effect of a higher morphine dose on protein composition in the brain cortex was increased from 15 to 31 proteins (9 were upregulated, 22 downregulated) when compared with a lower dose,
Table S2b. The functional significance of these proteins was mainly related to RNA processing (10), DNA processing (6), cytoskeletal changes (5), transport (4), and apoptosis (4),
Figure 9a,b (upper panels). Among the ten morphine-regulated proteins involved in RNA processes were: Mtr4 exosome RNA helicase (
Mtrex, ↑3.2-fold), thyroid hormone receptor-associated protein 3 (
Thrap3, ↑2.5-fold), cyclin-dependent kinase 17 (
Cdk17, ↑2.2-fold), lamina-associated polypeptide 2, isoform beta (
Tmpo, ↓7.3-fold), A-kinase anchor protein 8 (
Akap8, ↓7.2-fold),
Snrnp70 (↓7.1-fold), non-histone chromosomal protein HMG-17 (
Hmgn2, ↓3.2-fold),
Ddx46 (↓2.8-fold),
Mecp2 (↓2.2-fold), and UPF3B, regulator of nonsense mediated mRNA decay (
Upf3b, ↓2.1-fold). The results of Wu et al. [
35] revealed that dependence-associated genes and transcription regulators have essential roles in opioid-induced analgesia and tolerance. Morphine regulates the expression of transcription factors, such as CREB, ΔFosB, and nuclear factor-kappa B (NF-κB). Of particular interest in the study of addiction is ΔFosB, which is expressed in the brain after acute drug treatment but can persist for weeks or even months after the drug is withdrawn. It represents a mechanism by which drugs of abuse produce lasting changes in gene expression patterns long after the cessation of drug administration [
21,
36].
LYS739 treatment unveiled 152 altered cortical proteins, which is 4.9 times more than morphine and 7.6 times more than
LYS739 lower dose. Interestingly, only 30 proteins exhibited significant upregulation, whereas 122 proteins were downregulated,
Table S2b. These proteins were described to participate in RNA processes (41), transport-related processes (27), organization of the cytoskeleton (13), DNA processes (10), cell cycle (9), signal transduction (9), brain development (9), metabolic changes (7), immunity (6), apoptosis-related pathways (5), protein folding (5), protein ubiquitination (4), cell adhesion (3), ion homeostasis (3), aging (2), protein localization (1), and cell proliferation (1),
Figure 9a,b (middle panels). The high number of proteins exhibiting the decreased protein expression levels suggests a state of disturbed cell homeostasis. Notably, decreased level of ubiquitin carboxyl-terminal hydrolase 30 (
Usp30, ↓2.6-fold), small ubiquitin-related modifier 2 (
Sumo2, ↓2.3-fold), Ube 2l3 protein (
Ube2l3, ↓2.1-fold), and UBX domain-containing protein 1 (
Ubxn1, ↓2.1-fold) may indicate attenuation of protein degradation pathways in the etiology of opioid dependence and addiction [
37].
LFQ analysis revealed 26 altered proteins in the brain cortex after
LYS744 treatment. This is a similar number to morphine-induced alterations (31) and, noticeably, less than after the administration of the lower dose of
LYS744 (31). Six proteins were upregulated and 20 were downregulated,
Table S2b. According to the current annotations in the Uniprot database, these proteins were associated with RNA processing (7), DNA processing (5), cytoskeletal changes (4), transport (3), brain development (3), metabolism (3), apoptosis (2), aging (2), cell cycle (1), immunity (1), protein folding (1), cell adhesion (1), and protein localization (1),
Figure 9a,b (lower panels).
S100a9 was the most downregulated protein (↓5.4-fold), which was found to decrease in cortical samples after morphine administration (↓13.2-fold) and treatment with
LYS739 (↓13.4-fold) as well. Apart from its role in inflammation and apoptosis [
25], Toleikis et al. [
38] described its involvement in neurodegenerative disorders, such as Alzheimer’s and Parkinson’s diseases. Their data have shown that even small concentrations of
S100a9 resulted in the aggregation and formation of α-synuclein (
Snca) fibrils.
Snca is the main protein found in amyloid plaques in the brains of patients suffering from Parkinson’s disease. Volcano plots representing significantly altered proteins identified in rat brain cortex after treatment with 10 mg/kg of morphine,
LYS739, and
LYS744 are depicted in
Figure 7B. Hierarchical heatmap clustering of all identified protein expression profiles in rat brain cortex is presented in
Figure 8.
3.6. LFQ Analysis of Rat Hippocampus after 7-Day Treatment with Morphine, LYS739, and LYS744: The Dose-Effect of 10 mg/kg
Proteomic analysis unveiled 42 altered proteins in rat hippocampus after the treatment with the higher morphine dose; 8 proteins were upregulated, 34 were downregulated,
Table S2c. This is three times more than the effect of a lower morphine dose (14). The functional significance of hippocampal proteins was mainly related to RNA processes (8), transport (8), protein folding (6), cytoskeletal changes (4), brain development (4), metabolic changes (4), immunity (3), and cell adhesion (3),
Figure 10a,b (upper panels).
Interestingly, 6 decreased proteins were identified as protein chaperones: tubulin-specific chaperone A (
Tbca, ↓3.7-fold), BAG cochaperone 3 (
Bag3, ↓2.4-fold), H2-K region express gene 2, rat orthologue (
Pfdn6, ↓2.2-fold), HscB mitochondrial iron-sulfur cluster co-chaperone (
Hscb, ↓2.1-fold), RING-type E3 ubiquitin transferase (
Stub1, ↓2.0-fold), and mitochondrial import inner membrane translocase subunit Tim13 (
Timm13, ↓2.0-fold). Molecular chaperones prevent protein misfolding and aggregation-induced neurotoxicity. A growing body of evidence suggests that they play a significant role in neuron degeneration and may participate in Parkinson’s disease, Alzheimer’s disease, and Huntington’s disease [
39]. Okuyama et al. [
40] described that the accumulation of misfolded proteins in the endoplasmic reticulum (ER) induced the unfolded protein response (UPS) contributing to diverse pathological conditions, such as opioid tolerance development. They investigated that pharmacological chaperones 4-phenylbutyric acid (PBA) and tauroursodeoxycholic acid (TUDCA) suppressed the development of morphine tolerance and restored analgesia. Of note is an increased level of proteins involved in cell adhesion and maintenance of tissue integrity: desmoplakin (
Dsp, ↑7.0-fold), junction plakoglobin (
Jup, ↑2.3-fold), and integrin beta (
Itgb8, ↑2.1-fold). Civciristov et al. [
41] reported that the stimulation of µ-OR caused an enrichment of proteins associated with the formation of desmosomes.
LYS739 treatment resulted in the change of 45 proteins, upregulation was found for 20 proteins, 25 proteins were downregulated,
Table S2c. When compared with the brain cortex (152), the number of altered proteins in the hippocampus was 3.4 times lower. These proteins were predominantly involved in the cytoskeletal dynamics (13), RNA processing (10), brain development (9), transport (8), and cell adhesion (3),
Figure 10a,b (middle panels). Among identified cytoskeletal proteins were: neurofilament light polypeptide (
Nefl, ↑2.9-fold), tropomyosin alpha-4 chain (
Tpm4, ↑2.8-fold), Ras/Rap GTPase-activating protein SynGAP (
Syngap1, ↑2.6-fold), tropomyosin alpha-3 chain (
Tpm3, ↑2.6-fold), neurofilament medium polypeptide (
Nefm, ↑2.2-fold), hematopoietic cell-specific LYN substrate 1 (
Hcls1, ↑2.1-fold), nuclear mitotic apparatus protein 1 (
Numa1, ↓11.7-fold), myosin-11 (
Myh11, ↓10.1-fold), vimentin (
Vim, ↓7.2-fold), transgelin (
Tagln, ↓7.1-fold), filamin A (
Flna, ↓2.7-fold), non-muscle caldesmon (
Cald1, ↓2.1-fold), and microtubule-associated protein (
Map2, ↓2.0-fold). Neuronal plasticity is required to respond to external stimuli and is achieved by cytoskeletal reorganization controlled by phosphorylation of cytoskeletal proteins [
42]. Rothenfluh and Covan [
43] discussed that cytoskeleton plays an essential role in neuronal, dendritic, and behavioral plasticity observed in addicted animals. Drastichova et al. [
44] identified altered phosphorylated proteins associated with microtubule dynamics and the actin-spectrin network in rat brains isolated from animals even after six months of morphine withdrawal.
The effect of a higher
LYS744 dose on hippocampal proteome was surprisingly decreased from 42 to 26 proteins (6 were upregulated, 20 downregulated) when compared with the lower dose,
Table S2c. The functional significance of these proteins was associated with RNA processing (8), cytoskeletal changes (5), brain development (3), metabolism (2), transport (2), immunity (2), DNA processing (2), aging (2), protein ubiquitination (1), signal transduction (1), and cell cycle (1), as shown in
Figure 10a,b (lower panels). Cytoskeletal proteins:
Myh11 (↓15.0-fold),
Vim (↓10.4-fold),
Tagln (↓4.6-fold),
Flna (↓2.6-fold), and
Cald1 (↓2.3-fold) were found dysregulated after
LYS739 treatment as well, see above. Volcano plots representing significantly altered proteins identified in rat hippocampus after treatment with 10 mg/kg of morphine,
LYS739, and
LYS744 are depicted in
Figure 7C. Hierarchical heatmap clustering of all identified protein expression profiles in the rat hippocampus is presented in
Figure 8.
3.7. Summary of Results
3.7.1. Effects of Low Doses of Morphine, LYS739, and LYS744 (3 mg/kg)
LFQ analysis revealed 27 altered proteins in rat spleen lymphocytes after 7-day treatment of male rats with 3 mg/kg of morphine. The 12 proteins were upregulated, downregulation was detected for 15 proteins. Their subcellular localization and functional significance unveiled most proteins to be of cytoplasmic origin and functionally related to the organization of cytoskeleton. Fewer proteins were altered by LYS739, 5 proteins were upregulated, 15 downregulated. LYS744 induced more alterations than morphine, 29 proteins were upregulated, and 16 proteins were downregulated. More than half of these proteins were functionally related to actin cytoskeleton organization (11), RNA processing (8), and transport (6).
In the brain cortex, morphine administration resulted in the change of 15 proteins; 5 proteins were upregulated, 10 proteins were downregulated. Contrary to the spleen lymphocytes, LYS739 treatment resulted in a greater number of protein alterations than morphine, with 7 proteins upregulated and 13 downregulated. As in the case of spleen lymphocytes, treatment with LYS744 exhibited the highest number of alterations in cortical proteome—11 proteins were upregulated, 20 proteins were downregulated.
In the hippocampus, LFQ analysis revealed 14 proteins altered by morphine, 8 proteins were upregulated, 6 were downregulated. This change was similar to that found in the brain cortex (15) but lower twice in comparison with spleen lymphocytes (27). The effect of LYS739 on rat hippocampal protein profile was stronger than that of morphine with 8 proteins upregulated and 28 downregulated. Treatment with LYS744 unveiled 42 alterations in protein expression levels, which is more than in the brain cortex (31) and similar to that found in spleen lymphocytes (45); 24 proteins were upregulated and 18 proteins were downregulated.
3.7.2. Effects of Higher Doses of Morphine, LYS739, and LYS744 (10 mg/kg)
Proteomic analysis revealed 45 altered proteins in spleen lymphocytes after morphine treatment. Upregulation was found for 29 proteins; 16 proteins were downregulated. Their molecular functions and biological processes were mostly involved in transport (9), immunity (8), apoptosis (8), and RNA processing (7). When compared with the effect of a lower dose, the number of proteins associated with immune responses and apoptotic processes was increased, suggesting the state of cell dysfunction and neuroinflammation after chronic morphine administration. LYS739 treatment resulted in the change of 119 proteins, 51 proteins were upregulated, 68 downregulated. Thus, the number of altered proteins was six times higher than that obtained after the administration of a lower dose. The majority of altered proteins were related to RNA processing (40), transport (16), immunity (11), metabolism (10), and cytoskeletal changes. Higher dose of LYS744 caused the most significant change in protein expression levels of spleen lymphocytes. Four times more proteins were altered (182 proteins, 88 were upregulated, 94 downregulated) in comparison with the lower dose of LYS744. The identified proteins were primarily associated with RNA processing (58), transport (25), immune responses (16), metabolic changes (13), cytoskeletal changes (13), and apoptotic processes (12).
In the brain cortex, the effect of a higher morphine dose on protein composition was increased from 15 to 31 proteins (9 proteins were upregulated, 22 downregulated) when compared with a lower dose. The functional significance of these proteins was related to RNA processing (10), DNA processing (6), cytoskeletal changes (5), transport (4), and apoptosis (4). LYS739 treatment unveiled 152 altered cortical proteins, which is 4.9 times more than morphine and 7.6 times more than LYS739 lower dose. Interestingly, only 30 proteins exhibited significant upregulation, 122 proteins were downregulated. These proteins were described to participate in RNA processes (41), transport-related processes (27), organization of the cytoskeleton (13), DNA processes (10), cell cycle (9), signal transduction (9), brain development (9), metabolic changes (7), immunity (6), apoptosis-related pathways (5), protein folding (5), protein ubiquitination (4), cell adhesion (3), ion homeostasis (3), aging (2), protein localization (1), and cell proliferation (1). The high number of proteins exhibiting the decreased expression levels suggests a state of disturbed cell homeostasis. LYS744 treatment resulted in the change of 26 proteins, with 6 proteins upregulated and 20 downregulated.
In the hippocampus, proteomic analysis unveiled 42 altered proteins after the treatment with the higher morphine dose, 8 proteins were upregulated, 34 were downregulated. This is three times more than the effect of a lower morphine dose (14). The functional significance of hippocampal proteins was mainly related to RNA processes (8), transport (8), protein folding (6), cytoskeletal changes (4), brain development (4), metabolic changes (4), immunity (3), and cell adhesion (3). LYS739 treatment resulted in the change of 45 proteins, upregulation was found for 20 proteins, 25 proteins were downregulated. When compared with the brain cortex (152), the number of altered proteins in the hippocampus was 3.4 times lower. The altered proteins were predominantly involved in the cytoskeletal dynamics (13), RNA processing (10), brain development (9), transport (8), and cell adhesion (3). The effect of a higher dose of LYS744 on hippocampal proteome was surprisingly decreased from 42 to 26 proteins when compared with the lower dose (6 proteins were upregulated, 20 downregulated). The functional significance of these proteins was associated with RNA processing (8), cytoskeletal changes (5), brain development (3), metabolism (2), transport (2), immunity (2), DNA processing (2), aging (2), protein ubiquitination (1), signal transduction (1), and cell cycle (1).


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









