
Pitfalls of NMDA Receptor Modulation by Neuroactive Steroids. The Effect of Positive and Negative Modulation of NMDA Receptors in an Animal Model of Schizophrenia

 ,
, 
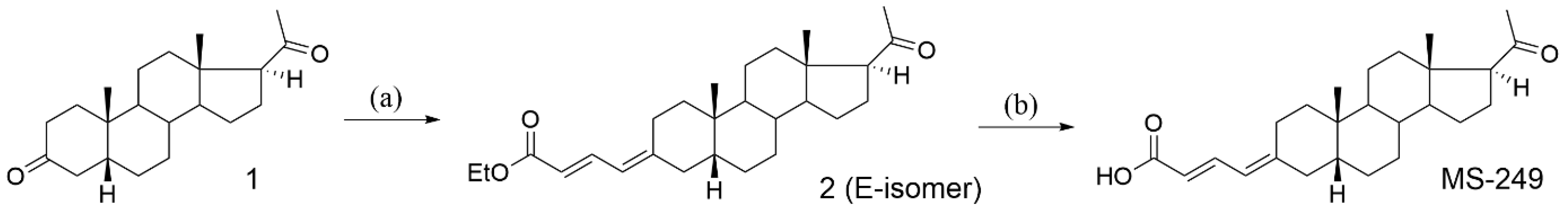{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Neuroactive Steroids
2.3. Electrophysiology
2.4. [35S]-Tert-Butylbicyclophosphorothionate ([35S]TBPS) Displacement
2.5. Behavioral Tests
2.5.1. Open Field (OF)
2.5.2. Elevated Plus Maze (EPM)
2.5.3. Pre-Pulse Inhibition of Acoustic Startle Response (PPI)
2.5.4. Step-Through Passive Avoidance (PA)
2.5.5. Carousel Maze (CM)
2.5.6. Statistical Analysis
3. Results
3.1. Electrophysiology
3.2. [35S]-Tert-Butylbicyclophosphorothionate ([35S]TBPS) Displacement
3.3. Behavioral Tests
3.3.1. Open Field (OF)
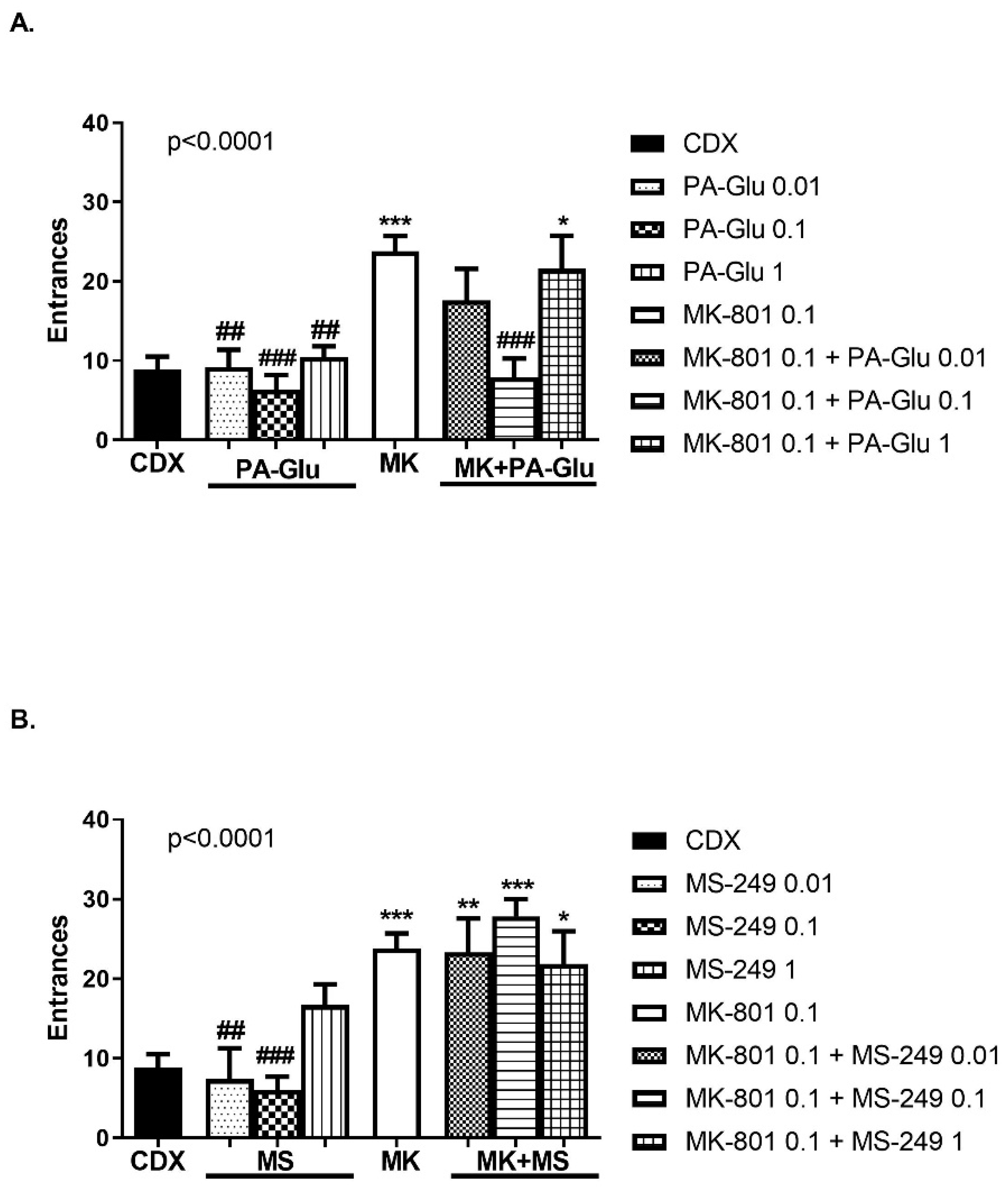
3.3.2. Elevated Plus Maze (EPM)
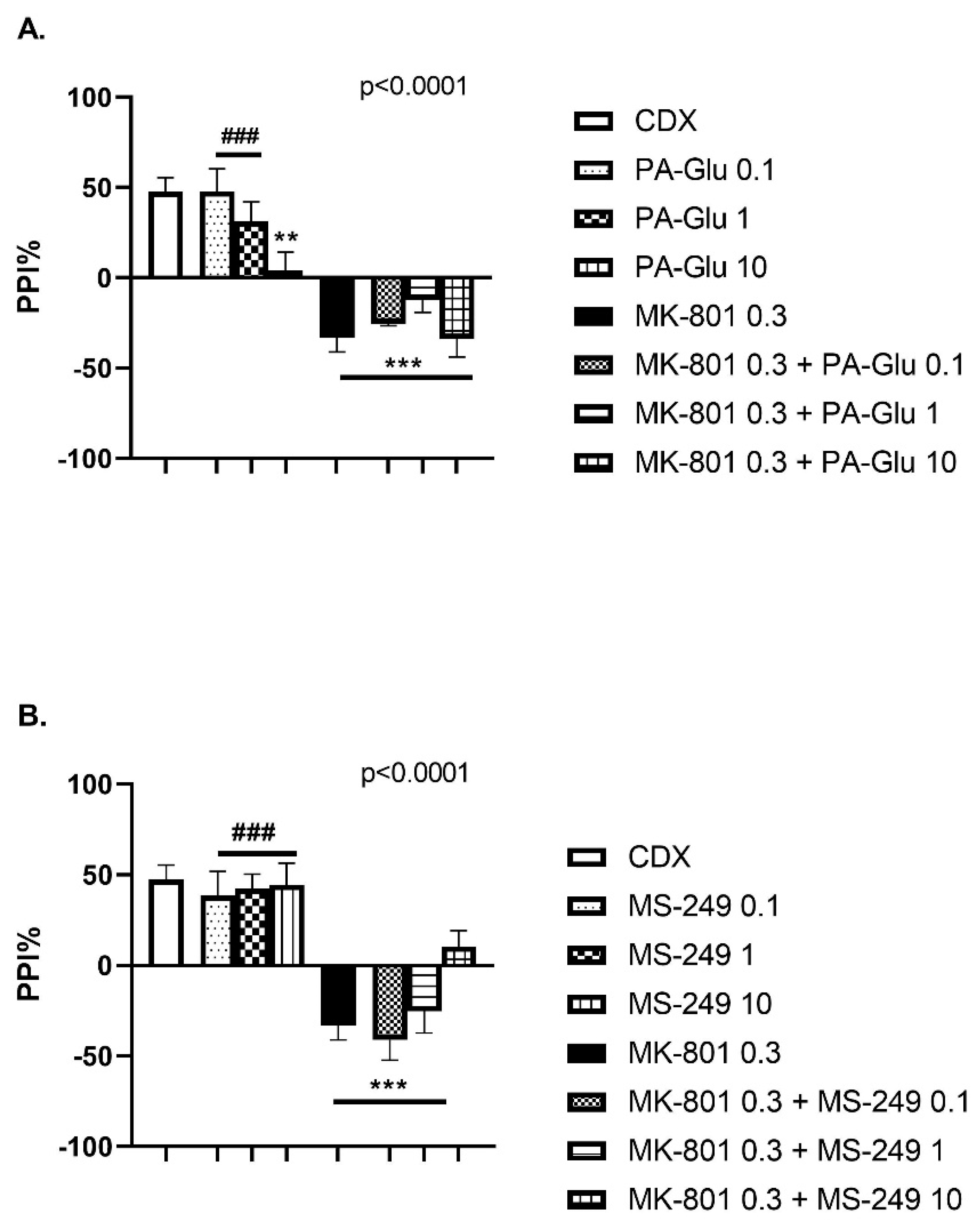
3.3.3. Pre-Pulse Inhibition of Startle Response (PPI)
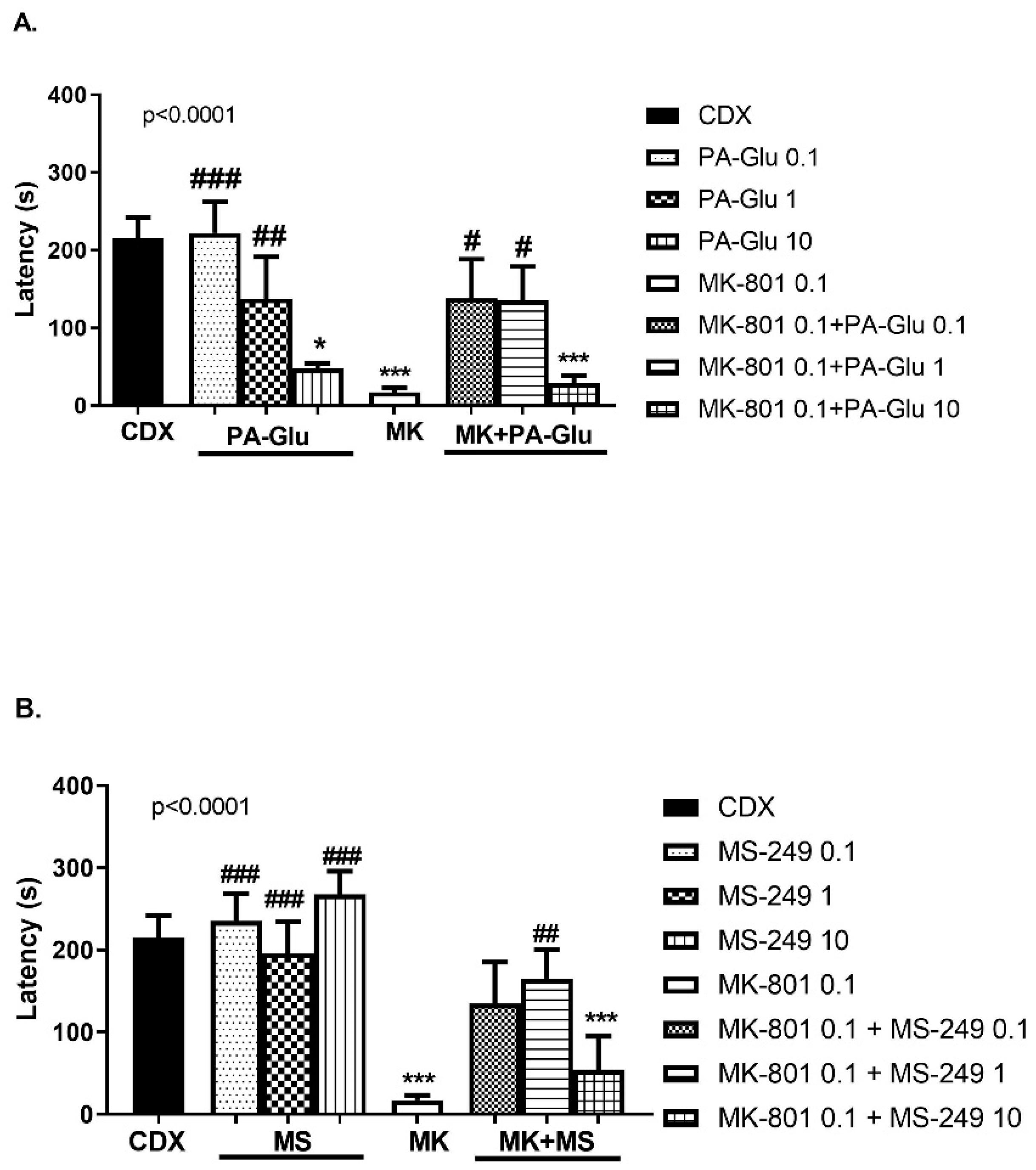
3.3.4. Step-Through Passive Avoidance (PA)
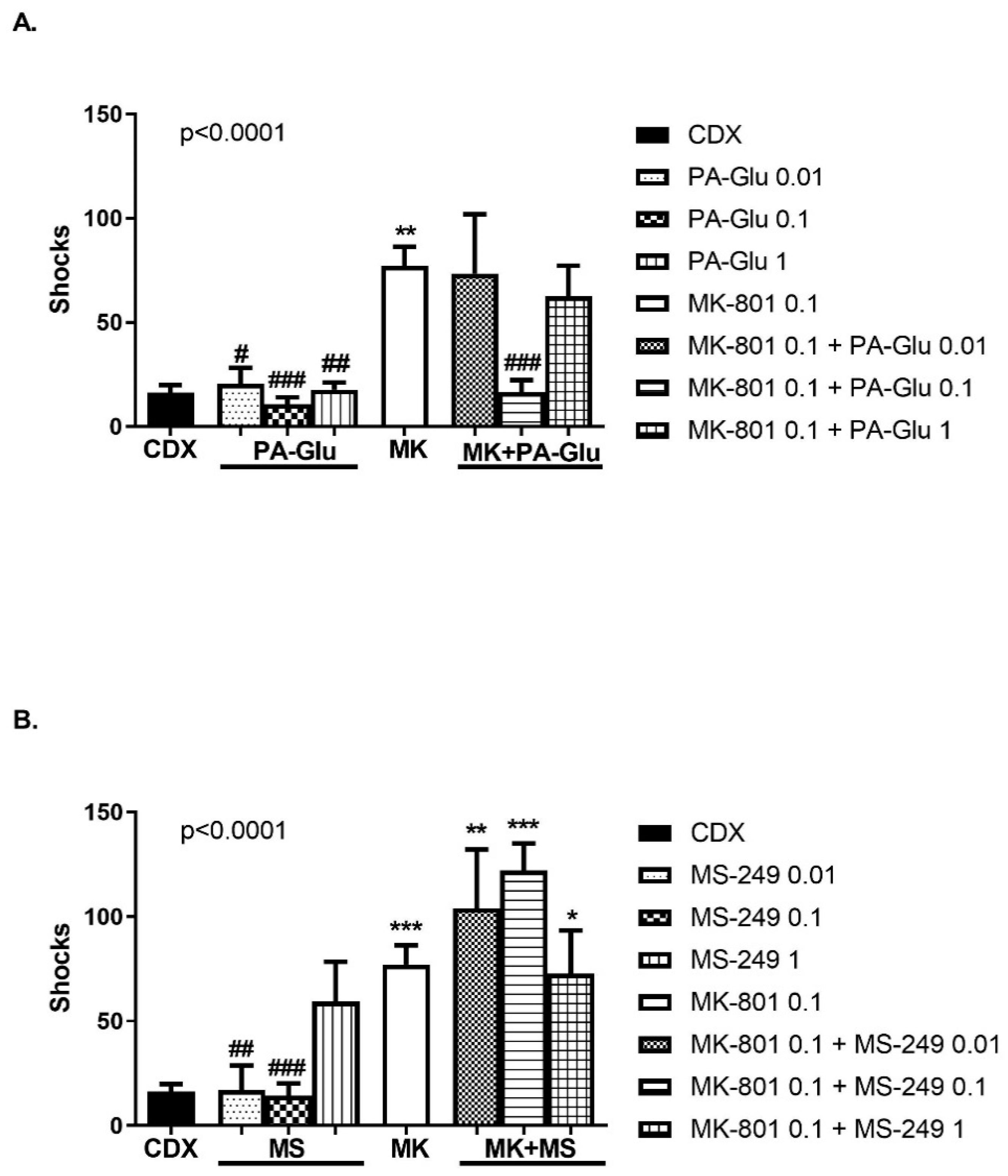
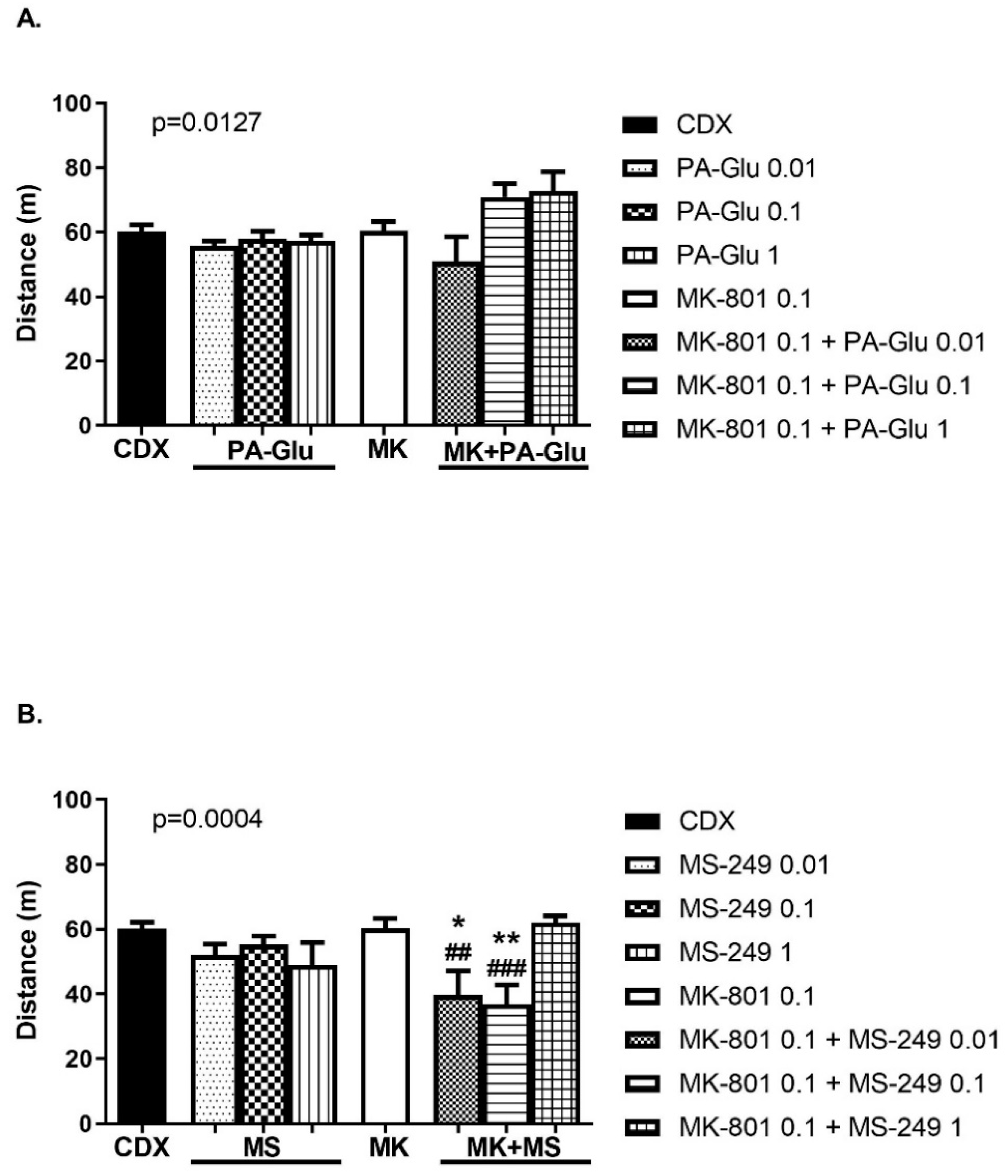
3.3.5. Carousel Maze (CM)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gibb, J.; Hayley, S.; Gandhi, R.; Poulter, M.O.; Anisman, H. Synergistic and additive actions of a psychosocial stressor and endotoxin challenge: Circulating and brain cytokines, plasma corticosterone and behavioral changes in mice. Brain Behav. Immun. 2008, 22, 573–589. [Google Scholar] [CrossRef]
- Snyder, M.A.; Gao, W.J. NMDA hypofunction as a convergence point for progression and symptoms of schizophrenia. Front. Cell. Neurosci. 2013, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kehrer, C. Altered excitatory-inhibitory balance in the NMDA-hypofunction model of schizophrenia. Front. Mol. Neurosci. 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyklicky, V.; Korinek, M.; Smejkalova, T.; Balik, A.; Krausova, B.; Kaniakova, M.; Lichnerova, K.; Cerny, J.; Krusek, J.; Dittert, I.; et al. Structure, function, and pharmacology of NMDA receptor channels. Physiol. Res. 2014, 63, S191–S203. [Google Scholar] [CrossRef] [PubMed]
- Augustine, G.J.; Santamaria, F.; Tanaka, K. Local calcium signaling in neurons. Neuron 2003, 40, 331–346. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.C.; Blackwell, K.T. Calcium: Amplitude, duration, or location? Biol. Bull. 2015. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef]
- Le Meur, K.; Galante, M.; Angulo, M.C.; Audinat, E. Tonic activation of NMDA receptors by ambient glutamate of non-synaptic origin in the rat hippocampus. J. Physiol. 2007. [Google Scholar] [CrossRef]
- Yao, L.; Grand, T.; Hanson, J.; Paoletti, P.; Zhou, Q. Higher ambient synaptic glutamate at inhibitory versus excitatory neurons differentially impacts NMDA receptor activity. Nat. Commun. 2018. [Google Scholar] [CrossRef]
- Lester, R.A.J.; Clements, J.D.; Westbrook, G.L.; Jahr, C.E. Channel kinetics determine the time course of NMDA receptor-mediated synaptic currents. Nature 1990. [Google Scholar] [CrossRef]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [Green Version]
- Marx, C.E.; Keefe, R.S.E.; Buchanan, R.W.; Hamer, R.M.; Kilts, J.D.; Bradford, D.W.; Strauss, J.L.; Naylor, J.C.; Payne, V.M.; Lieberman, J.A.; et al. Proof-of-concept trial with the neurosteroid pregnenolone targeting cognitive and negative symptoms in schizophrenia. Neuropsychopharmacology 2009. [Google Scholar] [CrossRef]
- Marx, C.E.; Lee, J.; Subramaniam, M.; Rapisarda, A.; Bautista, D.C.T.; Chan, E.; Kilts, J.D.; Buchanan, R.W.; Wai, E.P.; Verma, S.; et al. Proof-of-concept randomized controlled trial of pregnenolone in schizophrenia. Psychopharmacology 2014. [Google Scholar] [CrossRef]
- Ritsner, M.S.; Bawakny, H.; Kreinin, A. Pregnenolone treatment reduces severity of negative symptoms in recent-onset schizophrenia: An 8-week, double-blind, randomized add-on two-center trial. Psychiatry Clin. Neurosci. 2014. [Google Scholar] [CrossRef] [PubMed]
- Kreinin, A.; Bawakny, N.; Ritsner, M.S. Adjunctive pregnenolone ameliorates the cognitive deficits in recent-onset schizophrenia: An 8-week, randomized, double-blind, placebo-controlled trial. Clin. Schizophr. Relat. Psychoses 2017. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.S.; Park, J.; Marx, C.E.; Hynan, L.S.; Gardner, C.; Davila, D.; Nakamura, A.; Sunderajan, P.; Lo, A.; Holmes, T. A randomized, double-blind, placebo-controlled trial of pregnenolone for bipolar depression. Neuropsychopharmacology 2014. [Google Scholar] [CrossRef] [PubMed]
- Sripada, R.K.; Marx, C.E.; King, A.P.; Rampton, J.C.; Ho, S.S.; Liberzon, I. Allopregnanolone elevations following pregnenolone administration are associated with enhanced activation of emotion regulation neurocircuits. Biol. Psychiatry 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, D.G. Progesterone in the treatment of acute traumatic brain injury: A clinical perspective and update. Neuroscience 2011, 191, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, L.; Soni, D.; Meltzer, H.Y. Neurosteroid pregnenolone sulfate, alone, and as augmentation of lurasidone or tandospirone, rescues phencyclidine-induced deficits in cognitive function and social interaction. Behav. Brain Res. 2018. [Google Scholar] [CrossRef]
- Korinek, M.; Kapras, V.; Vyklicky, V.; Adamusova, E.; Borovska, J.; Vales, K.; Stuchlik, A.; Horak, M.; Chodounska, H.; Vyklicky, L. Neurosteroid modulation of N-methyl-d-aspartate receptors: Molecular mechanism and behavioral effects. Steroids 2011, 76, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Horak, M.; Vlcek, K.; Petrovic, M.; Chodounska, H.; Vyklicky, L. Molecular mechanism of pregnenolone sulfate action at NR1/NR2B receptors. J. Neurosci. 2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horak, M.; Vlcek, K.; Chodounska, H.; Vyklicky, L. Subtype-dependence of N-methyl-D-aspartate receptor modulation by pregnenolone sulfate. Neuroscience 2006. [Google Scholar] [CrossRef] [PubMed]
- Krausova, B.H.; Kysilov, B.; Cerny, J.; Vyklicky, V.; Smejkalova, T.; Ladislav, M.; Balik, A.; Korinek, M.; Chodounska, H.; Kudova, E.; et al. Site of action of brain neurosteroid pregnenolone sulfate at the N-methyl-D-aspartate receptor. J. Neurosci. 2020. [Google Scholar] [CrossRef]
- Flood, J.F.; Morley, J.E.; Roberts, E. Memory-enhancing effects in male mice of pregnenolone and steroids metabolically derived from it. Proc. Natl. Acad. Sci. USA 1992. [Google Scholar] [CrossRef] [Green Version]
- Mathis, C.; Paul, S.M.; Crawley, J.N. The neurosteroid pregnenolone sulfate blocks NMDA antagonist-induced deficits in a passive avoidance memory task. Psychopharmacology 1994. [Google Scholar] [CrossRef]
- Lee, K.H.; Cho, J.H.; Choi, I.S.; Park, H.M.; Lee, M.G.; Choi, B.J.; Jang, I.S. Pregnenolone sulfate enhances spontaneous glutamate release by inducing presynaptic Ca2+-induced Ca2+ release. Neuroscience 2010, 171, 106–116. [Google Scholar] [CrossRef]
- Cheney, D.L.; Uzunov, D.; Guidotti, A. Pregnenolone sulfate antagonizes dizocilpine amnesia: Role for allopregnanolone. Neuroreport 1995. [Google Scholar] [CrossRef]
- Reddy, D.S.; Kulkarni, S.K. Possible role of nitric oxide in the nootropic and antiamnesic effects of neurosteroids on aging- and dizocilpine-induced learning impairment. Brain Res. 1998, 799, 215–229. [Google Scholar] [CrossRef]
- Marx, C.E.; Bradford, D.W.; Hamer, R.M.; Naylor, J.C.; Allen, T.B.; Lieberman, J.A.; Strauss, J.L.; Kilts, J.D. Pregnenolone as a novel therapeutic candidate in schizophrenia: Emerging preclinical and clinical evidence. Neuroscience 2011, 191, 78–90. [Google Scholar] [CrossRef]
- Vallée, M.; Mayo, W.; Darnaudéry, M.; Corpéchot, C.; Young, J.; Koehl, M.; Le Moal, M.; Baulieu, E.E.; Robel, P.; Simon, H. Neurosteroids: Deficient cognitive performance in aged rats depends on low pregnenolone sulfate levels in the hippocampus. Proc. Natl. Acad. Sci. USA 1997. [Google Scholar] [CrossRef] [Green Version]
- Vyklicky, V.; Krausova, B.; Cerny, J.; Balik, A.; Zapotocky, M.; Novotny, M.; Lichnerova, K.; Smejkalova, T.; Kaniakova, M.; Korinek, M.; et al. Block of NMDA receptor channels by endogenous neurosteroids: Implications for the agonist induced conformational states of the channel vestibule. Sci. Rep. 2015. [Google Scholar] [CrossRef] [Green Version]
- Kussius, C.L.; Kaur, N.; Popescu, G.K. Pregnanolone sulfate promotes desensitization of activated NMDA receptors. J. Neurosci. 2009. [Google Scholar] [CrossRef] [PubMed]
- Covey, D.F.; Evers, A.S.; Mennerick, S.; Zorumski, C.F.; Purdy, R.H. Recent developments in structure-activity relationships for steroid modulators of GABAA receptors. Brain Res. Rev. 2001, 37, 91–97. [Google Scholar] [CrossRef]
- Sedláček, M.; Kořínek, M.; Petrovic, M.; Cais, O.; Adamusová, E.; Chodounská, H.; Vyklický, L. Neurosteroid modulation of ionotropic glutamate receptors and excitatory synaptic transmission. Physiol. Res. 2008, 57, S49–S57. [Google Scholar] [CrossRef] [PubMed]
- Maksay, G.; Laube, B.; Betz, H. Subunit-specific modulation of glycine receptors by neurosteroids. Neuropharmacology 2001. [Google Scholar] [CrossRef]
- Kobayashi, T.; Washiyama, K.; Ikeda, K. Pregnenolone sulfate potentiates the inwardly rectifying K+ channel Kir2.3. PLoS ONE 2009, 4, e6311. [Google Scholar] [CrossRef] [PubMed]
- Horishita, T.; Ueno, S.; Yanagihara, N.; Sudo, Y.; Uezono, Y.; Okura, D.; Sata, T. Inhibition by pregnenolone sulphate, a metabolite of the neurosteroid pregnenolone, of voltage-gated sodium channels expressed in Xenopus oocytes. J. Pharmacol. Sci. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, M.; Liere, P.; Akwa, Y.; Rajkowski, K.; Griffiths, W.; Bodin, K.; Sjövall, J.; Baulieu, E.E. Pregnenolone sulfate in the brain: A controversial neurosteroid. Neurochem. Int. 2008. [Google Scholar] [CrossRef] [PubMed]
- Vyklicky, V.; Smejkalova, T.; Krausova, B.; Balik, A.; Korinek, M.; Borovska, J.; Horak, M.; Chvojkova, M.; Kleteckova, L.; Vales, K.; et al. Preferential inhibition of tonically over phasically activated nmda receptors by pregnane derivatives. J. Neurosci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Rambousek, L.; Bubenikova-Valesova, V.; Kacer, P.; Syslova, K.; Kenney, J.; Holubova, K.; Najmanova, V.; Zach, P.; Svoboda, J.; Stuchlik, A.; et al. Cellular and behavioural effects of a new steroidal inhibitor of the N-methyl-D-aspartate receptor 3α5β-pregnanolone glutamate. Neuropharmacology 2011, 61. [Google Scholar] [CrossRef]
- Vales, K.; Rambousek, L.; Holubova, K.; Svoboda, J.; Bubenikova-Valesova, V.; Chodounska, H.; Vyklicky, L.; Stuchlik, A. 3α5β-Pregnanolone glutamate, a use-dependent NMDA antagonist, reversed spatial learning deficit in an animal model of schizophrenia. Behav. Brain Res. 2012, 235. [Google Scholar] [CrossRef]
- Holubova, K.; Nekovarova, T.; Pistovcakova, J.; Sulcova, A.; Stuchlík, A.; Vales, K. Pregnanolone glutamate, a novel use-dependent NMDA receptor inhibitor, exerts antidepressant-like properties in animal models. Front. Behav. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borovska, J.; Vyklicky, V.; Stastna, E.; Kapras, V.; Slavikova, B.; Horak, M.; Chodounska, H.; Vyklicky, L. Access of inhibitory neurosteroids to the NMDA receptor. Br. J. Pharmacol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Stuchlík, A.; Petrásek, T.; Prokopová, I.; Holubová, K.; Hatalová, H.; Valeš, K.; Kubík, S.; Dockery, C.; Wesierska, M. Place avoidance tasks as tools in the behavioral neuroscience of learning and memory. Physiol. Res. 2013, 62, S1–S19. [Google Scholar] [CrossRef]
- Hawkinson, J.E.; Kimbrough, C.L.; Belelli, D.; Lambert, J.J.; Purdy, R.H.; Lan, N.C. Correlation of neuroactive steroid modulation of [35S]t- butylbicyclophosphorothionate and [3H]flunitrazepam binding and γ- aminobutyric acid(A) receptor function. Mol. Pharmacol. 1994, 46, 977–985. [Google Scholar]
- Majewska, M.D.; Harrison, N.L.; Schwartz, R.D.; Barker, J.L.; Paul, S.M. Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor. Science 1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Luo, C.; Mao, X.Y.; Li, X.; Yin, J.Y.; Zhang, W.; Zhou, H.H.; Liu, Z.Q. Metformin reverses the schizophrenia-like behaviors induced by MK-801 in rats. Brain Res. 2019, 1719, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Hackler, E.A.; Byun, N.E.; Jones, C.K.; Williams, J.M.; Baheza, R.; Sengupta, S.; Grier, M.D.; Avison, M.; Conn, P.J.; Gore, J.C. Selective potentiation of the metabotropic glutamate receptor subtype 2 blocks phencyclidine-induced hyperlocomotion and brain activation. Neuroscience 2010, 168, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghedim, F.V.; Fraga, D.D.; Deroza, P.F.; Oliveira, M.B.; Valvassori, S.S.; Steckert, A.V.; Budni, J.; Dal-Pizzol, F.; Quevedo, J.; Zugno, A.I. Evaluation of behavioral and neurochemical changes induced by ketamine in rats: Implications as an animal model of mania. J. Psychiatr. Res. 2012, 46, 1569–1575. [Google Scholar] [CrossRef] [PubMed]
- Inostroza, M.; Cid, E.; Brotons-Mas, J.; Gal, B.; Aivar, P.; Uzcateg, Y.G.; Sandi, C.; de la Prida, L.M. Hippocampal-Dependent spatial memory in the water maze is preserved in an experimental model of temporal lobe epilepsy in rats. PLoS ONE 2011, 6, e22372. [Google Scholar] [CrossRef]
- Shoji, H.; Miyakawa, T. Effects of test experience, closed-arm wall color, and illumination level on behavior and plasma corticosterone response in an elevated plus maze in male C57BL/6J mice: A challenge against conventional interpretation of the test. Mol. Brain 2021. [Google Scholar] [CrossRef] [PubMed]
- Shoji, H.; Miyakawa, T. Relationships between the acoustic startle response and prepulse inhibition in C57BL/6J mice: A large-scale meta-analytic study. Mol. Brain 2018, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Swerdlow, N.R.; Light, G.A.; Thomas, M.L.; Sprock, J.; Calkins, M.E.; Green, M.F.; Greenwood, T.A.; Gur, R.E.; Gur, R.C.; Lazzeroni, L.C.; et al. Deficient prepulse inhibition in schizophrenia in a multi-site cohort: Internal replication and extension. Schizophr. Res. 2018, 198, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawabe, K.; Miyamoto, E. Effects of early postnatal MK-801 treatment on behavioral properties in rats: Differences according to treatment schedule. Behav. Brain Res. 2019, 370. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Barba, R.; Morales-Muñoz, I.; del Manzano, B.Á.; Fernández-Guinea, S.; Caballero, M.; Martínez-Gras, I.; Rubio-Valladolid, G. Relationship between measures of inhibitory processes in patients with schizophrenia: Role of substance abuse disorders. Psychiatry Res. 2011, 190, 187–192. [Google Scholar] [CrossRef]
- Wang, X.; Ding, S.; Lu, Y.; Jiao, Z.; Zhang, L.; Zhang, Y.; Yang, Y.; Zhang, Y.; Li, W.; Lv, L. Effects of sodium nitroprusside in the acute dizocilpine (MK-801) animal model of schizophrenia. Brain Res. Bull. 2019. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Mashavave, T.; Suvarna, A.; van den Buuse, M.; Sarnyai, Z. Effects of beta-hydroxybutyrate administration on MK-801-induced schizophrenia-like behaviour in mice. Psychopharmacology 2020. [Google Scholar] [CrossRef]
- Geyer, M.A.; Krebs-Thomson, K.; Braff, D.L.; Swerdlow, N.R. Pharmacological Studies of Prepulse Inhibition Models of Sensorimotor Gating Deficits in Schizophrenia: A Decade in Review. Psychopharmacology 2001, 156, 117–154. [Google Scholar] [CrossRef]
- Bukanova, J.V.; Solntseva, E.I.; Kolbaev, S.N.; Kudova, E. Modulation of GABA and glycine receptors in rat pyramidal hippocampal neurones by 3α5β-pregnanolone derivatives. Neurochem. Int. 2018. [Google Scholar] [CrossRef]
- Qaiser, M.Z.; Dolman, D.E.M.; Begley, D.J.; Abbott, N.J.; Cazacu-Davidescu, M.; Corol, D.I.; Fry, J.P. Uptake and metabolism of sulphated steroids by the blood–brain barrier in the adult male rat. J. Neurochem. 2017. [Google Scholar] [CrossRef] [Green Version]
- Romeo, E.; Cheney, D.L.; Zivkovic, I.; Costa, E.; Guidotti, A. Mitochondrial diazepam-binding inhibitor receptor complex agonists antagonize dizocilpine amnesia: Putative role for allopregnanolone. J. Pharmacol. Exp. Ther. 1994, 270, 89–96. [Google Scholar]
- Grillon, C.; Duncko, R.; Covington, M.F.; Kopperman, L.; Kling, M.A. Acute Stress Potentiates Anxiety in Humans. Biol. Psychiatry 2007, 62, 1183–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reznikov, L.R.; Grillo, C.A.; Piroli, G.G.; Pasumarthi, R.K.; Reagan, L.P.; Fadel, J. Acute stress-mediated increases in extracellular glutamate levels in the rat amygdala: Differential effects of antidepressant treatment. Eur. J. Neurosci. 2007, 25, 3109–3114. [Google Scholar] [CrossRef]
- Yuen, E.Y.; Liu, W.; Karatsoreos, I.N.; Feng, J.; McEwen, B.S.; Yan, Z. Acute stress enhances glutamatergic transmission in prefrontal cortex and facilitates working memory. Proc. Natl. Acad. Sci. USA 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treccani, G.; Musazzi, L.; Perego, C.; Milanese, M.; Nava, N.; Bonifacino, T.; Lamanna, J.; Malgaroli, A.; Drago, F.; Racagni, G.; et al. Stress and corticosterone increase the readily releasable pool of glutamate vesicles in synaptic terminals of prefrontal and frontal cortex. Mol. Psychiatry 2014, 19, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, G.S.; Sazma, M.A.; Yonelinas, A.P. The effects of acute stress on core executive functions: A meta-analysis and comparison with cortisol. Neurosci. Biobehav. Rev. 2016, 68, 651–668. [Google Scholar] [CrossRef] [Green Version]
- Biggio, G.; Concas, A.; Follesa, P.; Sanna, E.; Serra, M. Stress, ethanol, and neuroactive steroids. Pharmacol. Ther. 2007, 116, 140–171. [Google Scholar] [CrossRef] [Green Version]
- Barbaccia, M.L.; Roscetti, G.; Trabucchi, M.; Mostallino, M.C.; Concas, A.; Purdy, R.H.; Biggio, G. Time-dependent changes in rat brain neuroactive steroid concentrations and gabaa receptor function after acute stress. Neuroendocrinology 1996. [Google Scholar] [CrossRef]
- Grunze, H.C.R.; Rainnie, D.G.; Hasselmo, M.E.; Barkai, E.; Hearn, E.F.; McCarley, R.W.; Greene, R.W. NMDA-dependent modulation of CA1 local circuit inhibition. J. Neurosci. 1996, 16, 2034–2043. [Google Scholar] [CrossRef] [Green Version]
- Homayoun, H.; Moghaddam, B. NMDA receptor hypofunction produces opposite effects on prefrontal cortex interneurons and pyramidal neurons. J. Neurosci. 2007. [Google Scholar] [CrossRef] [PubMed]
- Kudo, K.; Tachikawa, E.; Kashimoto, T. Inhibition by pregnenolone sulfate of nicotinic acetylcholine response in adrenal chromaffin cells. Eur. J. Pharmacol. 2002, 456, 19–27. [Google Scholar] [CrossRef]
- Shirakawa, H.; Katsuki, H.; Kume, T.; Kaneko, S.; Akaike, A. Pregnenolone sulphate attenuates AMPA cytotoxicity on rat cortical neurons. Eur. J. Neurosci. 2005, 21, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Boero, G.; Porcu, P.; Morrow, A.L. Pleiotropic actions of allopregnanolone underlie therapeutic benefits in stress-related disease. Neurobiol. Stress 2020, 12, 100203. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holubova, K.; Chvojkova, M.; Hrcka Krausova, B.; Vyklicky, V.; Kudova, E.; Chodounska, H.; Vyklicky, L.; Vales, K. Pitfalls of NMDA Receptor Modulation by Neuroactive Steroids. The Effect of Positive and Negative Modulation of NMDA Receptors in an Animal Model of Schizophrenia. Biomolecules 2021, 11, 1026. https://doi.org/10.3390/biom11071026
Holubova K, Chvojkova M, Hrcka Krausova B, Vyklicky V, Kudova E, Chodounska H, Vyklicky L, Vales K. Pitfalls of NMDA Receptor Modulation by Neuroactive Steroids. The Effect of Positive and Negative Modulation of NMDA Receptors in an Animal Model of Schizophrenia. Biomolecules. 2021; 11(7):1026. https://doi.org/10.3390/biom11071026
Chicago/Turabian StyleHolubova, Kristina, Marketa Chvojkova, Barbora Hrcka Krausova, Vojtech Vyklicky, Eva Kudova, Hana Chodounska, Ladislav Vyklicky, and Karel Vales. 2021. "Pitfalls of NMDA Receptor Modulation by Neuroactive Steroids. The Effect of Positive and Negative Modulation of NMDA Receptors in an Animal Model of Schizophrenia" Biomolecules 11, no. 7: 1026. https://doi.org/10.3390/biom11071026