
Species and Strain Variability among Sarcina Isolates from Diverse Mammalian Hosts
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement, Sampling, and Cultivation Analysis of Fecal Samples
2.2. Colony Isolation and Identification
2.3. Phylogenetic Studies
2.4. Data Accessibility
3. Results
3.1. Occurrence of Sarcina spp. in the Feces of Screened Mammalian Hosts
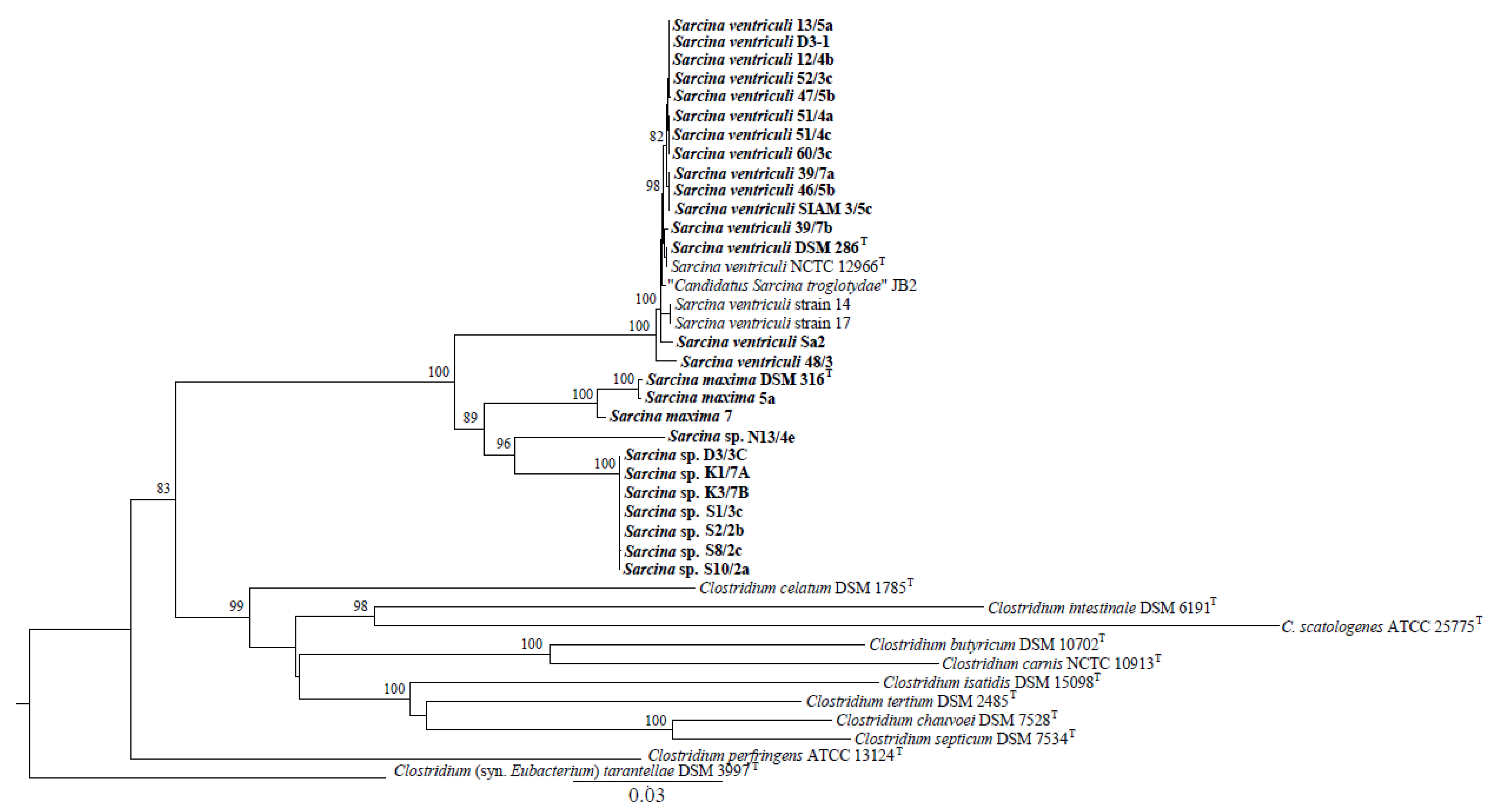
3.2. Identification and Taxonomic Classification of Isolated Sarcina Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Canale-Parola, E. Genus Sarcina. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; The Firmicutes; Springer: New York, NY, USA, 2009; Volume 3. [Google Scholar]
- Lawson, P.A.; Rainey, F.A. Proposal to restrict the genus Clostridium prazmowski to Clostridium butyricum and related species. Int. J. Syst. Evol. Microbiol. 2016, 66, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, A.; Aliuş, C.; Nica, A.E.; Antoniac, I.; Gheorghiță, D.; Grădinaru, S. Fatal outcome of gastric perforation due to infection with Sarcina spp. A case report. IDCases 2020, 19, e00711. [Google Scholar] [PubMed]
- Tintara, S.; Rice, S.; Patel, D. Sarcina Organisms: A Potential Cause of Emphysematous Gastritis in a Patient with Gastroparesis. Am. J. Gastroenterol. 2019, 114, 859. [Google Scholar] [CrossRef] [PubMed]
- Singh, K. Emphysematous Gastritis Associated with Sarcina ventriculi. Case Rep. Gastroenterol. 2019, 13, 207–213. [Google Scholar] [CrossRef]
- Alvin, M.; Al Jalbout, N. Emphysematous gastritis secondary to Sarcina ventriculi. BMJ Case Rep. 2018, 2018. [Google Scholar] [CrossRef]
- de Meij, T.G.; van Wijk, M.P.; Mookhoek, A.; Budding, A.E. Ulcerative gastritis and esophagitis in two children with Sarcina ventriculi infection. Front. Med. 2017, 4, 145. [Google Scholar] [CrossRef]
- Tartaglia, D.; Coccolini, F.; Mazzoni, A.; Strambi, S.; Cicuttin, E.; Cremonini, C.; Taddei, G.; Puglisi, A.G.; Ugolini, C.; Di Stefano, I.; et al. Sarcina Ventriculi infection: A rare but fearsome event. A Systematic Review of the Literature. Int. J. Infect. Dis. 2022, 115, 48–61. [Google Scholar] [CrossRef]
- Vatn, S.; Gunnes, G.; Nybø, K.; Juul, H.M. Possible involvement of Sarcina ventriculi in canine and equine acute gastric dilatation. Acta Vet. Scand. 2000, 41, 333–337. [Google Scholar] [CrossRef]
- Im, J.Y.; Sokol, S.; Duhamel, G.E. Gastric dilatation associated with gastric colonization with Sarcina-like bacteria in a cat with chronic enteritis. J. Am. Anim. Hosp. Assoc. 2017, 53, 321–325. [Google Scholar] [CrossRef]
- DeBey, B.M.; Blanchard, P.C.; Durfee, P.T. Abomasal bloat associated with Sarcina-like bacteria in goat kids. J. Am. Vet. Med. Assoc. 1996, 209, 1468. [Google Scholar]
- Vatn, S.; Tranulis, M.A.; Hofshagen, M. Sarcina-like bacteria, Clostridium fallax and Clostridium sordellii in lambs with abomasal bloat, haemorrhage and ulcers. J. Comp. Pathol. 2000, 122, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Owens, L.A.; Colitti, B.; Hirji, I.; Pizarro, A.; Jaffe, J.E.; Moittié, S.; Bishop-Lilly, K.A.; Estrella, L.A.; Voegtly, L.J.; Kuhn, J.H.; et al. A Sarcina bacterium linked to lethal disease in sanctuary chimpanzees in Sierra Leone. Nat. Commun. 2021, 12, 763. [Google Scholar] [CrossRef] [PubMed]
- Haroon Al Rasheed, M.R.; Kim, G.J.; Senseng, C. A Rare Case of Sarcina ventriculi of the Stomach in an Asymptomatic Patient. Int. J. Surg. Pathol. 2016, 24, 142–145. [Google Scholar] [CrossRef]
- Makovska, M.; Modrackova, N.; Bolechova, P.; Drnkova, B.; Neuzil-Bunesova, V. Antibiotic susceptibility screening of primate-associated Clostridium ventriculi. Anaerobe 2021, 69, 102347. [Google Scholar] [CrossRef]
- Bergey, D.H.G.M.; Whitman, W.B.; Parte, A.C. The actinobacteria. Bergey’s Man. Syst. Bacteriol. 2012, 5, 171–224. [Google Scholar]
- Modrackova, N.; Stovicek, A.; Burtscher, J.; Bolechova, P.; Killer, J.; Domig, K.J.; Neuzil-Bunesova, V. The bifidobacterial distribution in the microbiome of captive primates reflects parvorder and feed specialization of the host. Sci. Rep. 2021, 11, 15273. [Google Scholar] [CrossRef]
- Marcelino, L.P.; Valentini Junior, D.F.; Machado, S.M.; Schaefer, P.G.; Rivero, R.C.; Osvaldt, A.B. Sarcina ventriculi a rare pathogen. Autops. Case Rep. 2021, 11, e2021337. [Google Scholar] [CrossRef]
- Ratuapli, S.; LamHimlin, D.; Heigh, R. Gastroparesis Associated with Sarcina ventriculi Infection of the Stomach. Am. J. Gastroenterol. 2013, 108, S219. [Google Scholar] [CrossRef]
- Tuuminen, T.; Suomala, P.; Vuorinen, S. Sarcina ventriculi in blood: The first documented report since 1872. BMC Infect. Dis. 2013, 13, 169. [Google Scholar] [CrossRef]
- Bortolotti, P.; Kipnis, E.; Faure, E.; Faure, K.; Wacrenier, A.; Fauquembergue, M.; Penven, M.; Messaadi, S.; Marceau, L.; Dessein, R.; et al. Clostridium ventriculi bacteremia following acute colonic pseudo-obstruction: A case report. Anaerobe 2019, 59, 32–34. [Google Scholar] [CrossRef]
- Sauter, J.L.; Nayar, S.K.; Anders, P.D.; D’Amico, M.; Butnor, K.J.; Wilcox, R.L. Co-existence of Sarcina Organisms and Helicobacter pylori Gastritis/Duodenitis in Pediatric Siblings. J. Clin. Anat. Pathol. 2013, 1, 103. [Google Scholar] [CrossRef]
- Neuzil-Bunesova, V.; Lugli, G.A.; Modrackova, N.; Makovska, M.; Mrazek, J.; Mekadim, C.; Musilova, S.; Svobodova, I.; Spanek, R.; Ventura, M.; et al. Bifidobacterium canis sp. Nov., a novel member of the bifidobacterium pseudolongum phylogenetic group isolated from faeces of a dog (canis lupus f. familiaris). Int. J. Syst. Evol. Microbiol. 2020, 70, 5040–5047. [Google Scholar] [CrossRef]
- Bunešová, V.; Joch, M.; Musilová, S.; Rada, V. Bifidobacteria, Lactobacilli, and Short Chain Fatty Acids of Vegetarians and Omnivores. Sci. Agric. Bohem. 2017, 48, 47–54. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Information Retrieval Ltd.: London, UK, 1999; pp. c1979–c2000. [Google Scholar]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Mekadim, C.; Killer, J.; Pechar, R.; Mrazek, J. Variable regions of the glys, infb and rplb genes usable as novel genetic markers for identification and phylogenetic purposes of genera belonging to the family propionibacteriaceae. Int. J. Syst. Evol. Microbiol. 2018, 68, 2697–2705. [Google Scholar] [CrossRef]
- Edwards, G.T.; Woodger, N.G.A.; Barlow, A.M.; Bell, S.J.; Harwood, D.G.; Otter, A.; Wight, A.R. Sarcina-like bacteria associated with bloat in young lambs and calves. Vet. Rec. 2008, 163, 391–393. [Google Scholar] [CrossRef]
- Tuzcu, M.; Tuzcu, N.; Akcakavak, G.; Celik, Z. Diagnosis of Sarcina ventriculi-derived haemorrhagic abomasitis in lambs by histopathology and real-time PCR. Acta Vet. Brno 2022, 91, 227–233. [Google Scholar] [CrossRef]
- Ushida, K.; Tsuchida, S.; Ogura, Y.; Hayashi, T.; Sawada, A.; Hanya, G. Draft genome sequences of Sarcina ventriculi strains isolated from wild Japanese macaques in Yakushima Island. Genome Announc. 2016, 4, e01694-15. [Google Scholar] [CrossRef]
- Frey, J.C.; Rothman, J.M.; Pell, A.N.; Nizeyi, J.B.; Cranfield, M.R.; Angert, E.R. Fecal bacterial diversity in a wild gorilla. Appl. Environ. Microbiol. 2006, 72, 3788–3792. [Google Scholar] [CrossRef] [PubMed]
- Moeller, A.H.; Shilts, M.; Li, Y.; Rudicell, R.S.; Lonsdorf, E.V.; Pusey, A.E.; Wilson, M.L.; Hahn, B.H.; Ochman, H. Siv-induced instability of the chimpanzee gut microbiome. Cell Host Microbe 2013, 14, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.; Sun, X.; Patel, V.H.; Sanz, C.; Morgan, D.; Dantas, G. The microbiome and resistome of chimpanzees, gorillas, and humans across host lifestyle and geography. ISME J. 2020, 14, 1584–1599. [Google Scholar] [CrossRef] [PubMed]
- West, A.G.; Waite, D.W.; Deines, P.; Bourne, D.G.; Digby, A.; McKenzie, V.J.; Taylor, M.W. The microbiome in threatened species conservation. Biol. Conserv. 2019, 229, 85–98. [Google Scholar] [CrossRef]
- Wagner Mackenzie, B.; Waite, D.W.; Hoggard, M.; Douglas, R.G.; Taylor, M.W.; Biswas, K. Bacterial community collapse: A meta-analysis of the sinonasal microbiota in chronic rhinosinusitis. Environ. Microbiol. 2017, 19, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef]
- Rada, V.; Petr, J. A new selective medium for the isolation of glucose non-fermenting bifidobacteria from hen caeca. J. Microbiol. Methods 2000, 43, 127–132. [Google Scholar] [CrossRef]
- Goodwin, S.; Zeikus, J.G. Physiological adaptations of anaerobic bacteria to low pH: Metabolic control of proton motive force in Sarcina ventriculi. J. Bacteriol. 1987, 169, 2150–2157. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| PRIMATES (n = 65) | Moustached tamarin (0/2) | Cavalier King Charles spaniel (0/1) |
| Brown-mantled tamarin (0/1) | Northern Talapoin monkey (1/1) | Crossbreed dog (0/12) |
| Campbell’s Mona monkey (1/1) | Northern white-cheeked gibbon (3/4) | Czechoslovakian wolfdog (0/2) |
| Common marmoset (1/3) | Patas monkey (2/3) | Foxterier (0/1) |
| Cotton-top tamarin (0/1) | Putty-nosed monkey (1/1) | German Shepherd dog (0/28) |
| De Brazza’s monkey (1/1) | Pygmy marmoset (0/1) | Golden retriever (0/2) |
| Diana monkey (0/1) | Red-handed tamarin (0/2) | Havanese (0/3) |
| Emperor tamarin (0/4) | Ring-tailed lemur (4/4) | Labrador retriever (0/1) |
| Gibbon siamang (3/3) | Roloway monkey (1/1) | Not known (0/1) |
| Goeldi’s marmoset (0/1) | Silvery marmoset (0/4) | Samoyed (0/1) |
| Golden lion tamarin (1/8) | Yellow-cheeked crested gibbon (3/4) | Swiss shepherd (0/2) |
| Golden-bellied mangabey (0/1) | White-faced saki (0/1) | Whippet (0/1) |
| Vervet monkey (1/1) | White-headed marmoset (0/2) | OTHERS (n = 62) |
| Hamadryas baboon (0/1) | DOGS (n = 70) | Asian elephant (10/10) |
| Hamlyn’s monkey (1/1) | American staffordshire terrier (1/1) | Eastern black rhinoceros (2/2) |
| Chimpanzee (0/5) | Basenji (0/1) | Holstein-Friesian calf (2/50) |
| Lesser spot-nosed monkey (1/1) | Belgian shepherd (0/2) | |
| Lion-tailed macaque (0/1) | Border collie (0/11) |
| Isolated from | Host Location | Sarcina Isolate | 16S rRNA Identification | Similarity (%) | nts | |
|---|---|---|---|---|---|---|
| Human (vegetarian) | Homo sapiens | Prague, CZ | 7 | Sarcina maxima | 99.63 | 1356 |
| Northern white-cheeked gibbon | Nomascus leucogenys | Zoo Liberec, CZ | 40/5a | NRI | ||
| 40/5b | NRI | |||||
| 59/5a | NRI | |||||
| 60/2 | NRI | |||||
| 60/3a | NRI | |||||
| 60/3c | Sarcina ventriculi | 99.58 | 1419 | |||
| Gibon siamang | Symphalangus syndactylus | Zoo Olomouc, CZ | SIAM 3/5b | Sarcina ventriculi | 99.33 | 1347 |
| SIAM 3/5c | Sarcina ventriculi | 99.41 | 1362 | |||
| Yellow-cheeked crested gibbon | Nomascus gabriellae | Zoo Bratislava, SK | 51/4a | Sarcina ventriculi | 99.50 | 1414 |
| 51/4b | NRI | |||||
| 51/4c | Sarcina ventriculi | 99.01 | 1422 | |||
| Yellow-cheeked crested gibbon | Nomascus gabriellae | Zoo Olomouc, CZ | 46/4a | NRI | ||
| 46/4c | NRI | |||||
| 46/5b | Sarcina ventriculi | 99.64 | 1420 | |||
| 46/6 | NRI | |||||
| 47/4b | NRI | |||||
| 47/5b | Sarcina ventriculi | 99.50 | 1414 | |||
| De Brazza’s monkey | Cercopithecus neglectus | Zoo Plzeň, CZ | 39/5a | NRI | ||
| 39/6a | NRI | |||||
| 39/7a | Sarcina ventriculi | 99.56 | 1367 | |||
| 39/7b | Sarcina ventriculi/Sarcina sp. | 98.86 | 1410 | |||
| Hamlyn’s monkey | Cercopithecus hamlyni | Zoo Bojnice, SK | 55/5a | NRI | ||
| Roloway monkey | Cercopithecus roloway | Zoo Bojnice, SK | 56/4a | NRI | ||
| 56/4b | NRI | |||||
| 56/5 | NRI | |||||
| Lesser spot-nosed monkey | Cercopithecus petaurista | Zoo Bojnice, SK | 57/3 | NRI | ||
| 57/4 | NRI | |||||
| 57/4a | NRI | |||||
| 57/4b | NRI | |||||
| 57/4c | NRI | |||||
| 57/5 | NRI | |||||
| 57/5a | NRI | |||||
| 57/5b | NRI | |||||
| Vervet monkey | Chlorocebus sabaeus | Zoo Hodonín, CZ | 52/3a | Sarcina ventriculi | 99.50 | 1397 |
| 52/3b | Sarcina ventriculi | 99.56 | 1352 | |||
| 52/3c | Sarcina ventriculi | 99.58 | 1424 | |||
| Ring-tailed lemur | Lemur catta | Zoo Olomouc, CZ | 12/4b | Sarcina ventriculi | 99.56 | 1382 |
| 13/5a | Sarcina ventriculi | 99.56 | 1371 | |||
| 48/3 | Sarcina ventriculi | 99.14 | 1406 | |||
| Golden lion tamarin | Leontopithecus rosalia | Zoo Olomouc, CZ | D3-1 | Sarcina ventriculi | 99.63 | 1355 |
| Eastern black rhinoceros | Diceros bicornis michaeli | Safari park Dvůr Králové, CZ | 5a | Sarcina maxima | 100 | 1403 |
| 5b | Sarcina maxima | 99.92 | 1308 | |||
| Asian elephant | Elephas maximus | Zoo Liberec, CZ | S1/2a | Sarcina maxima/Sarcina sp. | 98.89 | 1352 |
| S1/2c | NRI | |||||
| S1/3b | NRI | |||||
| S1/3c | Sarcina maxima/Sarcina sp. | 98.74 | 1354 | |||
| S1/3d | NRI | |||||
| S10/2a | Sarcina maxima/Sarcina sp. | 98.88 | 1346 | |||
| S2/2b | Sarcina maxima/Sarcina sp. | 98.83 | 1371 | |||
| S4/2c | NRI | |||||
| S8/2c | Sarcina ventriculi/Sarcina sp. | 98.82 | 1356 | |||
| Asian elephant | Elephas maximus | Zoo Ústí nad Labem, CZ | K1/6A | NRI | ||
| K1/7A | Sarcina ventriculi | 98.95 | 1241 | |||
| K3/7B | Sarcina maxima | 98.74 | 1356 | |||
| D1/5A | NRI | |||||
| D3/3C | Sarcina maxima | 99.06 | 1273 | |||
| Holstein-Friesian calf | Bos taurus | Vražkov, CZ | Sa1 | NRI | ||
| Sa2 | Sarcina ventriculi | 99.56 | 1370 | |||
| Dog | Canis lupus f. familiaris | Prague, CZ | N13/4e | Sarcina maxima/Sarcina sp. | 98.90 | 1371 |
| Soil | DSMZ—type strain | DSM 286T | Sarcina ventriculi | 99.57 | 1392 | |
| Elephant | DSMZ—type strain | DSM 316T | Sarcina maxima | 100 | 1349 | |
| Gene | Forward/Reverse Primers (5′ → 3′) PCR Conditions | Length of the Amplified/Sequenced Fragment (Nucleotides) |
|---|---|---|
| ileS | ileSSarF: TGGACAACAACTCCGTGG/ileSSarR: TGACCACATTCACATTCCC | 699 |
| 95 °C for 5 min; 30 × (95 °C for 45 s; 56 °C for 40 s; 72 °C for 50 s); 72 °C for 6 min | ||
| pheT | pheTSarF: TGTAACGGAAAGAGAGCC/pheTSarR: ATCTAAATCAAGCTCTGCC | 570 |
| 95 °C for 5 min; 30 × (95 °C for 45 s; 53 °C for 40 s; 72 °C for 50 s); 72 °C for 6 min | ||
| pyrG | PyrGSarF: ACAGCAGCATCTTTAGG/PyrGSarR: AACTCTCCTGCTTTTCC | 480 |
| 95 °C for 5 min; 30 × (95 °C for 45 s; 53 °C for 40 s; 72 °C for 50 s); 72 °C for 6 min | ||
| rplB | rplBSarF: GGTGGTAGAAATGGTCAAGG/rplBSarR: ATCCTCTAACAGTAGGTCTGA | 516 |
| 95 °C for 5 min; 30 × (95 °C for 45 s; 56 °C for 40 s; 72 °C for 50 s); 72 °C for 6 min | ||
| rplC | rplCSarF: CCAGTAACAGTTGTAGAAGC/rplCSarR: CTTGATGGATCTGATGAAGC | 360 |
| 95 °C for 5 min; 30 × (95 °C for 45 s; 53 °C for 40 s; 72 °C for 50 s); 72 °C for 6 min | ||
| rpsC | rpsCSarF: CTCACGGACTAAGAGTTGG/rpsCSarR: TTTCTGCACCACCTAATCTACC | 444 |
| 95 °C for 5 min; 30 × (95 °C for 45 s; 56 °C for 40 s; 72 °C for 50 s); 72 °C for 6 min |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makovska, M.; Killer, J.; Modrackova, N.; Ingribelli, E.; Amin, A.; Vlkova, E.; Bolechova, P.; Neuzil-Bunesova, V. Species and Strain Variability among Sarcina Isolates from Diverse Mammalian Hosts. Animals 2023, 13, 1529. https://doi.org/10.3390/ani13091529
Makovska M, Killer J, Modrackova N, Ingribelli E, Amin A, Vlkova E, Bolechova P, Neuzil-Bunesova V. Species and Strain Variability among Sarcina Isolates from Diverse Mammalian Hosts. Animals. 2023; 13(9):1529. https://doi.org/10.3390/ani13091529
Chicago/Turabian StyleMakovska, Marie, Jiri Killer, Nikol Modrackova, Eugenio Ingribelli, Ahmad Amin, Eva Vlkova, Petra Bolechova, and Vera Neuzil-Bunesova. 2023. "Species and Strain Variability among Sarcina Isolates from Diverse Mammalian Hosts" Animals 13, no. 9: 1529. https://doi.org/10.3390/ani13091529