
Structural Characterization of Mono- and Dimethylphosphatidylethanolamines from Various Organisms Using a Complex Analytical Strategy including Chiral Chromatography

 , , and
, , and 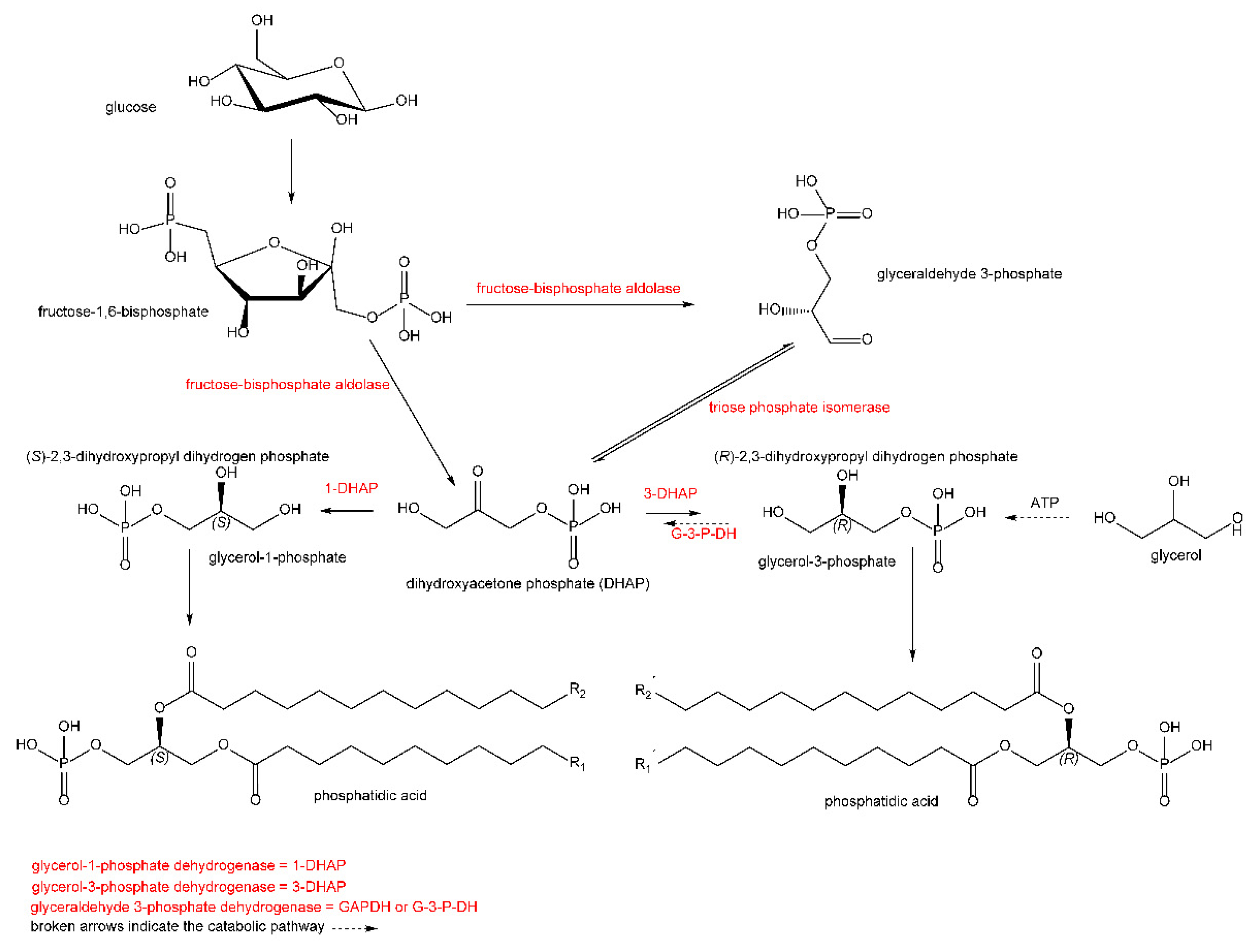{kind=link}
{kind=link}
{kind=link}
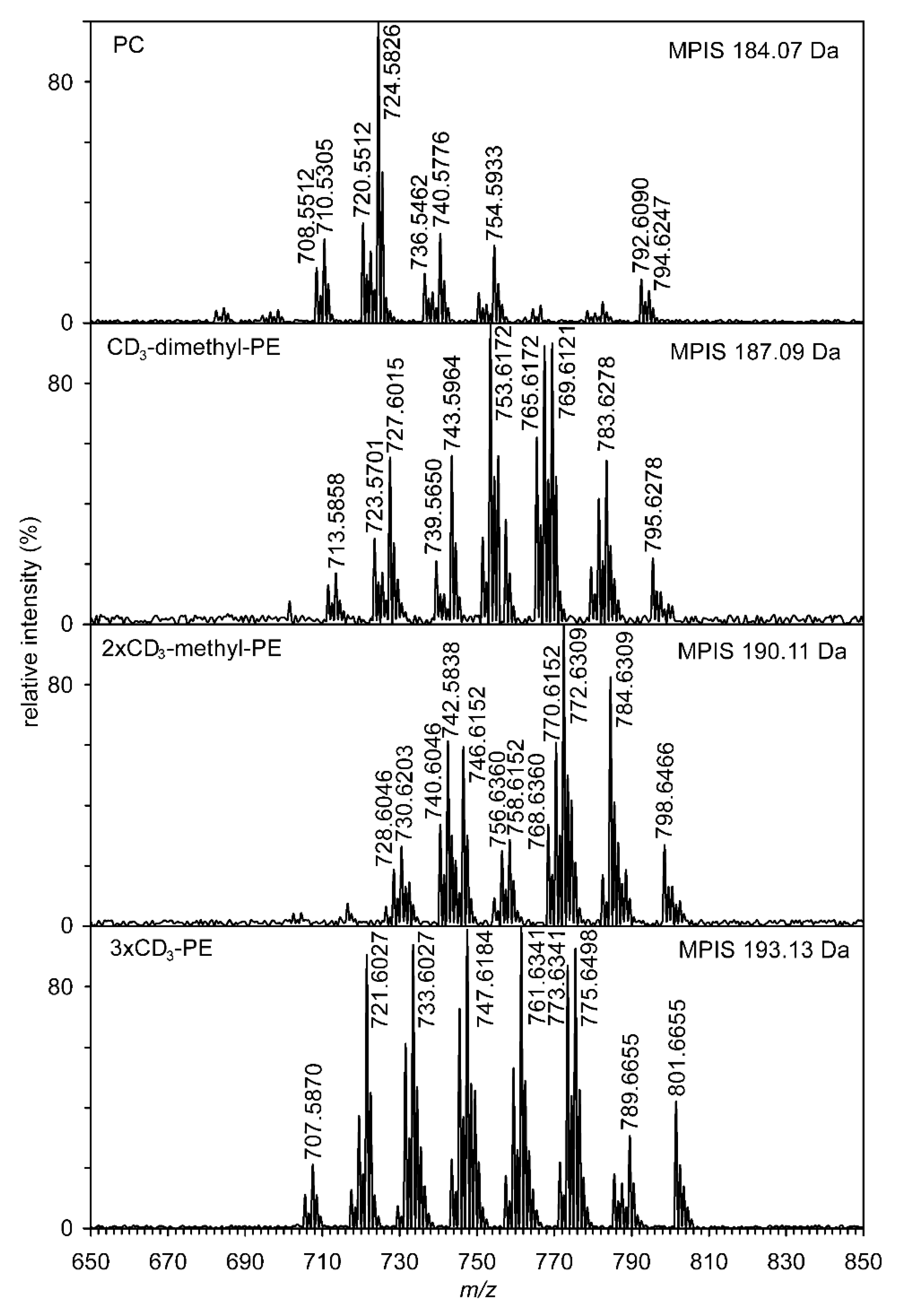{kind=link}
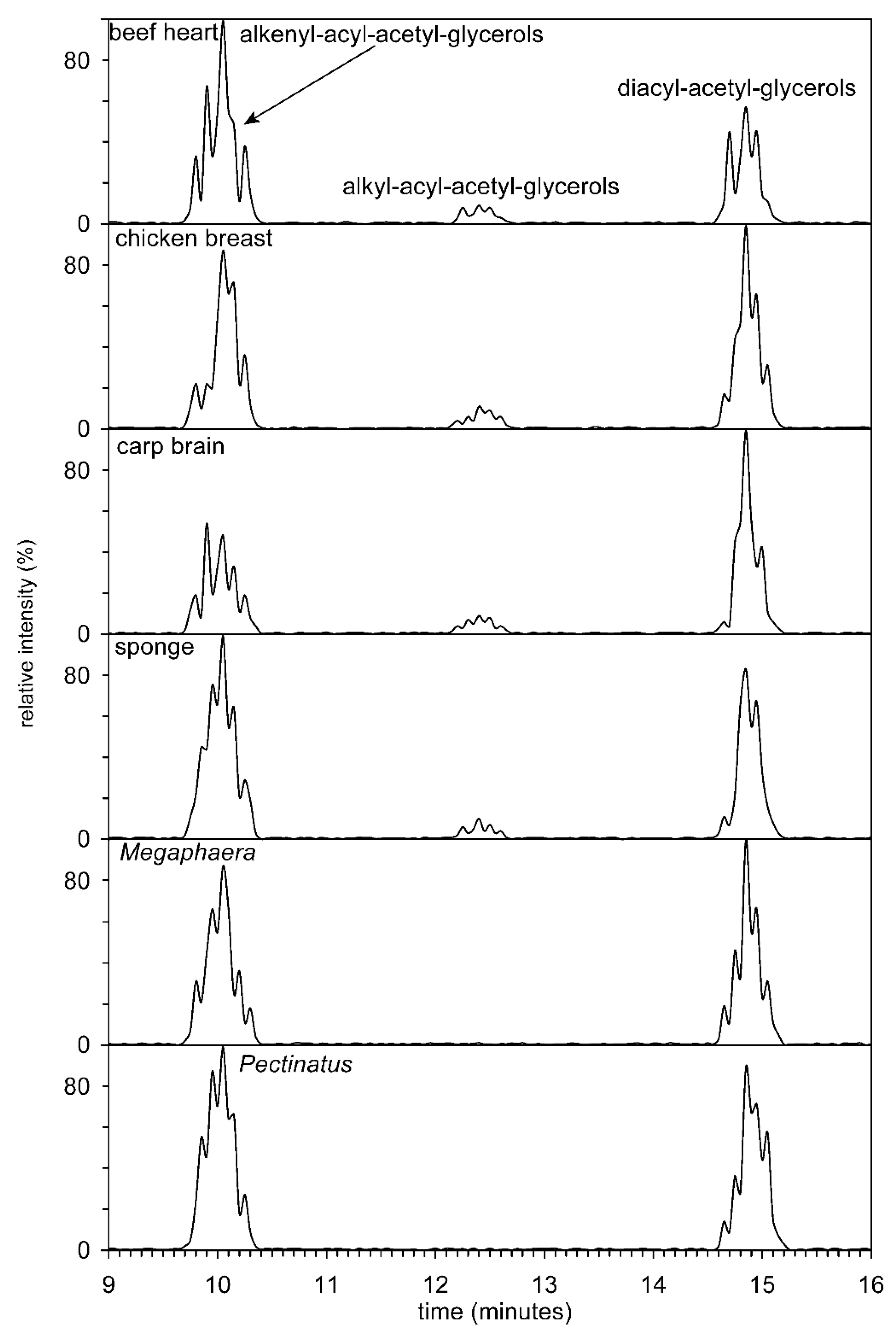{kind=link}
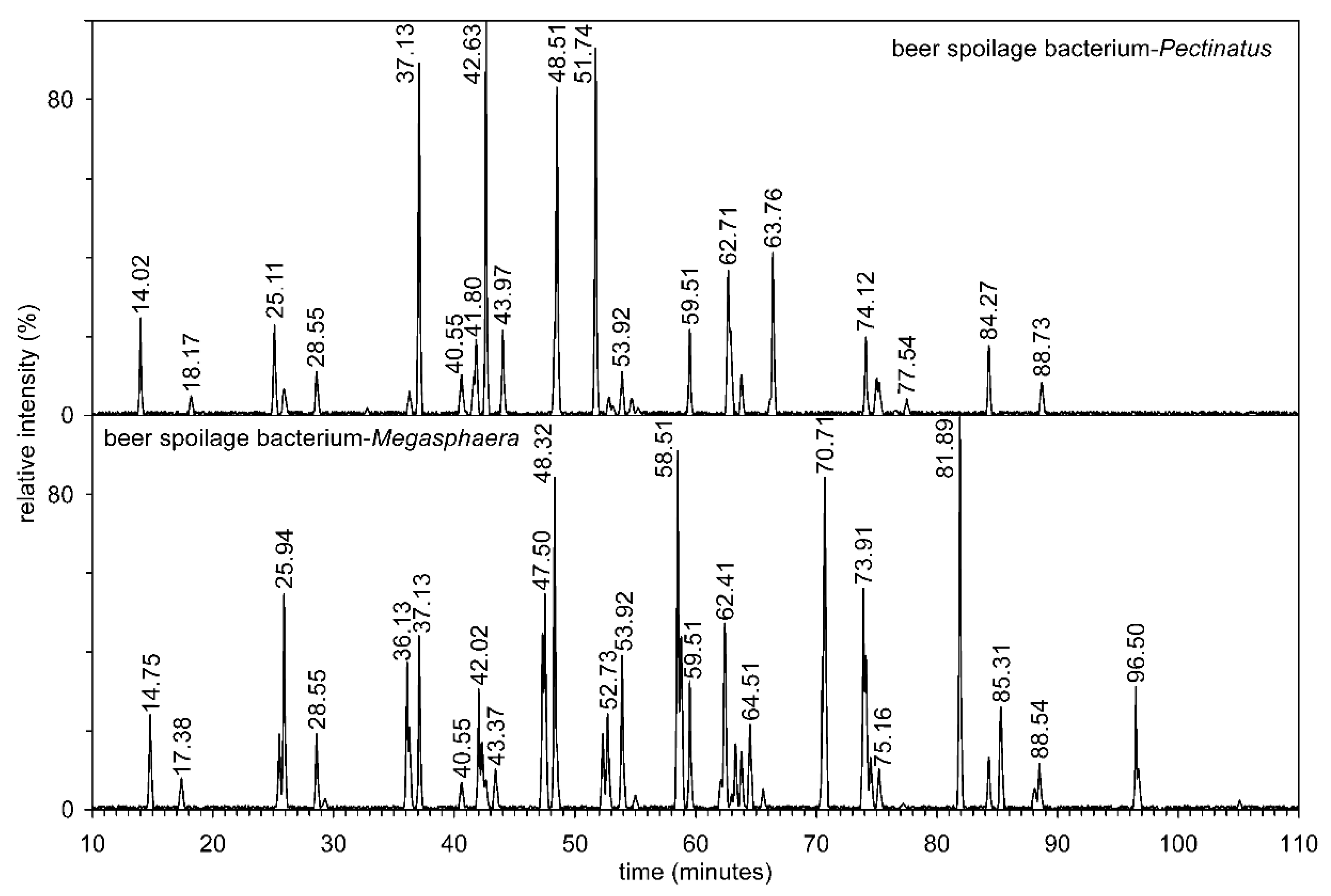{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Standards
2.2. Sample Preparation of Organisms
2.3. Extraction of Lipids
2.4. Semipreparative HILIC-ESI-MS
2.5. Analysis of Aldehydes and Fatty Acids from Phospolipids
2.6. Labeling with Deuterated CD3I
2.7. Shotgun Lipidomics
2.8. Phospholipase C Hydrolysis
2.9. Separation of Subclasses—Alkyl-Acyl, Alkenyl-Acyl, and Diacylglycerols
2.10. Separation and Identification of Acetyldiradylglycerols by RP-HPLC/MS2
2.11. Chiral Analysis of Acetyldiradylacetyl Glycerols by HPLC-ESI-MS2
2.12. Lipid Nomenclature
3. Results and Discussion
3.1. Analysis of Fatty Acids and Aldehydes
3.2. Hydrophilic Interaction Liquid Chromatography-HILIC
3.3. Identification of Methyl-PE and Dimethyl-PE
3.4. Separation of Subclasses–Alkyl-Acyl, Alkenyl-Acyl and Diacylglycerols
3.5. Analysis of Ether Lipids by RP-HPLC
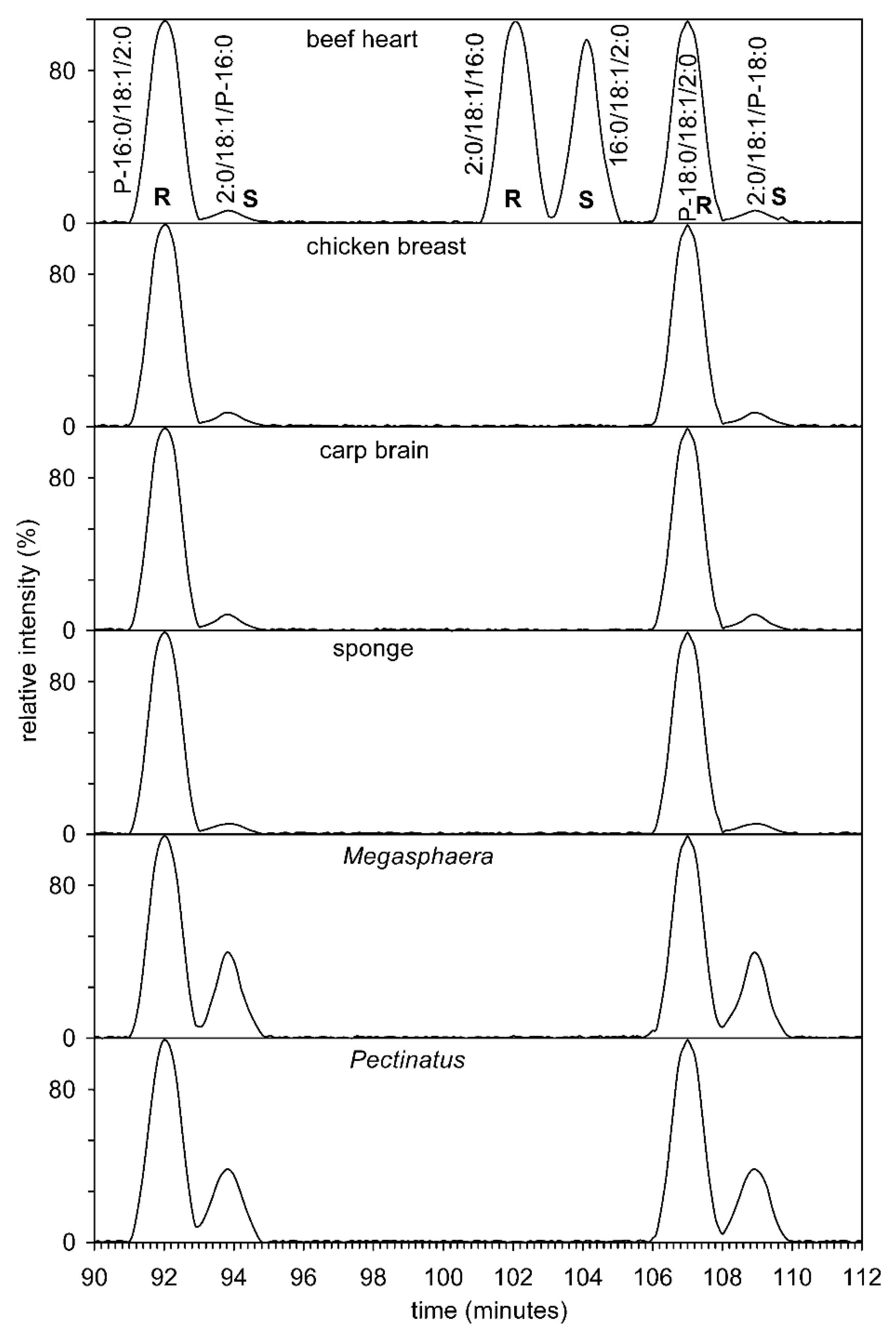
3.6. Analysis by Chiral HPLC
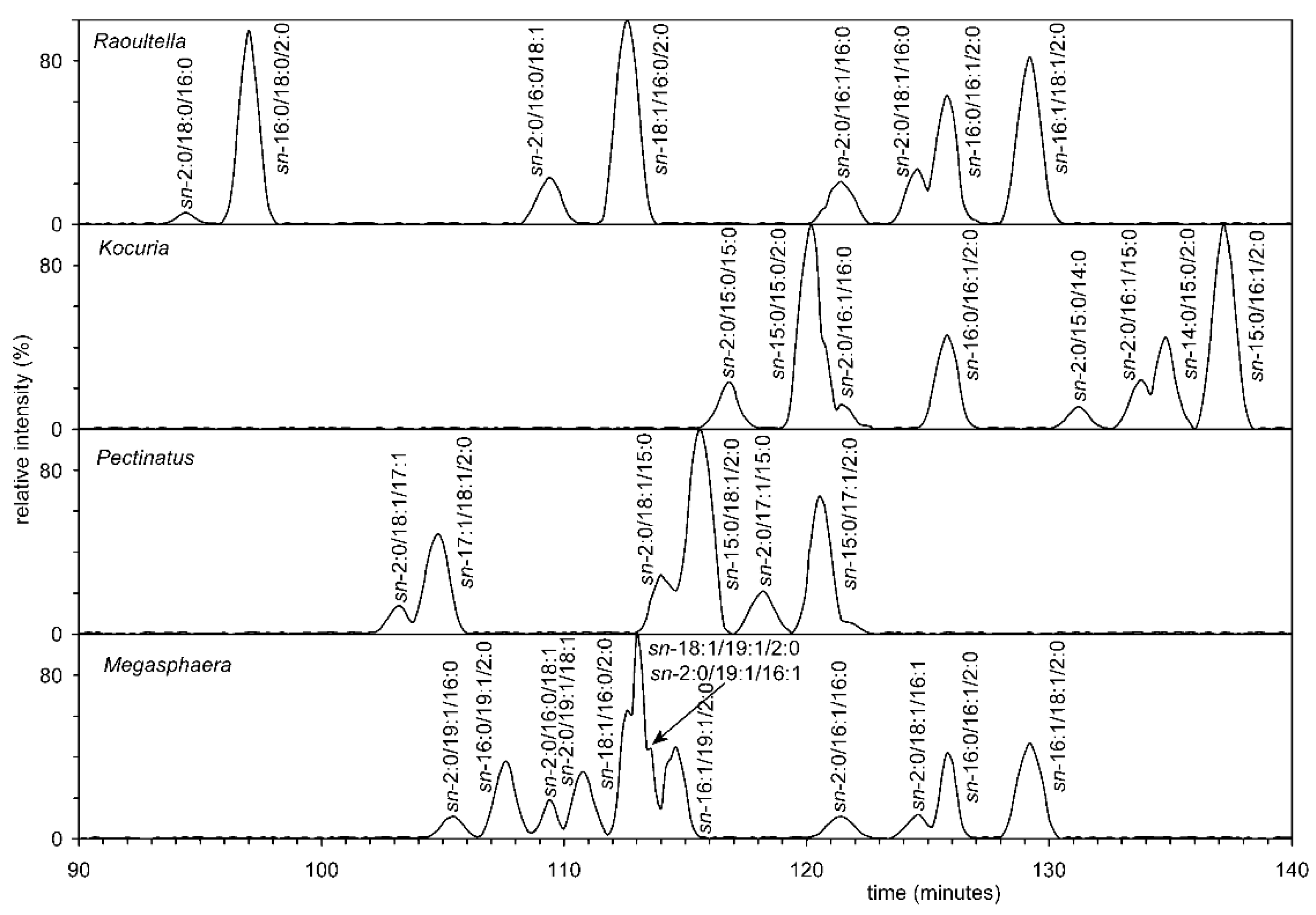
3.7. Analysis of all Four Bacteria by Chiral HPLC
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, M.; Wang, C.; Han, R.H.; Han, X. Novel advances in shotgun lipidomics for biology and medicine. Prog. Lipid Res. 2016, 61, 83–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, L.K.; Vance, J.E.; Vance, D.E. Phosphatidylcholine biosynthesis and lipoprotein metabolism. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2012, 1821, 754–761. [Google Scholar] [CrossRef]
- McMaster, C.R. From yeast to humans-roles of the Kennedy pathway for phosphatidylcholine synthesis. FEBS Lett. 2018, 592, 1256–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y. Headgroup biosynthesis of phosphatidylcholine and phosphatidylethanolamine in seed plants. Prog. Lipid Res. 2021, 82, 101091. [Google Scholar] [CrossRef] [PubMed]
- Geiger, O.; Lopez-Lara, I.M.; Sohlenkamp, C. Phosphatidylcholine biosynthesis and function in bacteria. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 503–513. [Google Scholar] [CrossRef]
- Vitova, M.; Palyzova, A.; Rezanka, T. Plasmalogens-ubiquitous molecules occurring widely, from anaerobic bacteria to humans. Prog. Lipid Res. 2021, 83, 101111. [Google Scholar] [CrossRef]
- Yao, J.W.; Rock, C.O. Phosphatidic acid synthesis in bacteria. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Athenstaedt, K.; Daum, G. Biosynthesis of phosphatidic acid in lipid particles and endoplasmic reticulum of Saccharomyces cerevisiae. J. Bacteriol. 1997, 179, 7611–7616. [Google Scholar] [CrossRef] [Green Version]
- Koga, Y.; Morii, H. Biosynthesis of ether-type polar lipids in archaea and evolutionary considerations. Microbiol. Mol. Biol. Rev. 2007, 71, 97–120. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, L.; von Meijenfeldt, F.A.B.; Westbye, A.B.; Yadav, S.; Hopmans, E.C.; Dutilh, B.E.; Damste, J.S.S. Bridging the membrane lipid divide: Bacteria of the FCB group superphylum have the potential to synthesize archaeal ether lipids. ISME J. 2021, 15, 168–182. [Google Scholar] [CrossRef]
- Tang, D.Q.; Zou, L.; Yin, X.X.; Ong, C.N. HILIC-MS for metabolomics: An attractive and complementary approach to RPLC-MS. Mass Spectrom. Rev. 2016, 35, 574–600. [Google Scholar] [CrossRef] [PubMed]
- Granafei, S.; Azzone, P.; Spinelli, V.A.; Losito, I.; Palmisano, F.; Cataldi, T.R. Hydrophilic interaction and reversed phase mixed-mode liquid chromatography coupled to high resolution tandem mass spectrometry for polar lipids analysis. J. Chromatogr. A 2016, 1477, 47–55. [Google Scholar] [CrossRef]
- Schwudke, D.; Oegema, J.; Burton, L.; Entchev, E.; Hannich, J.T.; Ejsing, C.S.; Kurzchalia, T.; Shevchenko, A. Lipid profiling by multiple precursor and neutral loss scanning driven by the data-dependent acquisition. Anal. Chem. 2006, 78, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Bilgin, M.; Markgraf, D.F.; Duchoslav, E.; Knudsen, J.; Jensen, O.N.; de Kroon, A.I.P.M.; Ejsing, C.S. Quantitative profiling of PE, MMPE, DMPE, and PC lipid species by multiple precursor ion scanning: A tool for monitoring PE metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Schneiter, R.; Brugger, B.; Sandhoff, R.; Zellnig, G.; Leber, A.; Lampl, M.; Athenstaedt, K.; Hrastnik, C.; Eder, S.; Daum, G.; et al. Electrospray ionization tandem mass spectrometry (ESI-MS/MS) analysis of the lipid molecular species composition of yeast subcellular membranes reveals acyl chain-based sorting/remodeling of distinct molecular species en route to the plasma membrane. J. Cell Biol. 1999, 146, 741–754. [Google Scholar] [CrossRef]
- Hein, E.M.; Hayen, H. Comparative lipidomic profiling of S. cerevisiae and four other Hemiascomycetous yeasts. Metabolites 2012, 2, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Moore, E.K.; Hopmans, E.C.; Rijpstra, W.I.C.; Villanueva, L.; Dedysh, S.N.; Kulichevskaya, I.S.; Wienk, H.; Schoutsen, F.; Damste, J.S.S. Novel mono-, di-, and trimethylornithine membrane lipids in Northern Wetland Planctomycetes. Appl. Environ. Microbiol. 2013, 79, 6874–6884. [Google Scholar] [CrossRef] [Green Version]
- Basconcillo, L.S.; Zaheer, R.; Finan, T.M.; McCarry, B.E. A shotgun lipidomics approach in Sinorhizobium meliloti as a tool in functional genomics. J. Lipid Res. 2009, 50, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.Z.; Grunler, J.; Piao, S.F.; Sindelar, P.J. Separation and quantitation of phospholipids and their ether analogues by high-performance liquid chromatography. Anal. Biochem. 2001, 297, 137–143. [Google Scholar] [CrossRef]
- Qin, D.H.; Byun, H.S.; Bittman, R. Synthesis of plasmalogen via 2,3-bis-O-(4 ’-methoxybenzyl)-sn-glycerol. J. Am. Chem. Soc. 1999, 121, 662–668. [Google Scholar] [CrossRef]
- Koch, J.; Lackner, K.; Wohlfarter, Y.; Sailer, S.; Zschocke, J.; Werner, E.R.; Watschinger, K.; Keller, M.A. Unequivocal mapping of molecular ether lipid species by LC-MS/MS in plasmalogen-deficient mice. Anal. Chem. 2020, 92, 11268–11276. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Ventura, G.; Sardanelli, A.M.M.; Savino, L.; Losito, I.; De Michele, G.; Palmisano, F.; Cataldi, T.R.I. Searching for potential lipid biomarkers of Parkinson’s disease in Parkin-mutant human skin fibroblasts by HILIC-ESI-MS/MS: Preliminary findings. Int. J. Mol. Sci. 2019, 20, 3341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezanka, T.; Siristova, L.; Matoulkova, D.; Sigler, K. Hydrophilic interaction liquid chromatography: ESI-MS/MS of plasmalogen phospholipids from Pectinatus bacterium. Lipids 2011, 46, 765–780. [Google Scholar] [CrossRef] [PubMed]
- Cifkova, E.; Holcapek, M.; Lisa, M. Nontargeted lipidomic characterization of porcine organs using hydrophilic interaction liquid chromatography and off-line two-dimensional liquid chromatography-electrospray ionization mass spectrometry. Lipids 2013, 48, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Berkecz, R.; Tomosi, F.; Kormoczi, T.; Szegedi, V.; Horvath, J.; Janaky, T. Comprehensive phospholipid and sphingomyelin profiling of different brain regions in mouse model of anxiety disorder using online two-dimensional (HILIC/RP)-LC/MS method. J. Pharm. Biomed. Anal. 2018, 149, 308–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhoenacker, D.; Ruben t’Kindt, F.D.; Pat Sandra, K.S. Unraveling the complexity of lipidomes by multiple heart-cutting Q-TOF LC/MS with the Agilent 1290 Infinity 2D-LC solution. Agil. Technol. 2019, 1–14. [Google Scholar]
- Pham, T.H.; Zaeem, M.; Fillier, T.A.; Nadeem, M.; Vidal, N.P.; Manful, C.; Cheema, S.; Cheema, M.; Thomas, R.H. Targeting modified lipids during routine lipidomics analysis using HILIC and C30 reverse phase liquid chromatography coupled to mass spectrometry. Sci. Rep. 2019, 9, 5048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, Y.; Horrocks, L.A. Separation of alkenylacyl, alkylacyl, and diacyl analogs and their molecular species by high-performance liquid-chromatography. J. Lipid Res. 1983, 24, 1268–1275. [Google Scholar] [CrossRef]
- Vitova, M.; Stranska, M.; Palyzova, A.; Rezanka, T. Detailed structural characterization of cardiolipins from various biological sources using a complex analytical strategy comprising fractionation, hydrolysis and chiral chromatography. J. Chromatogr. A 2021, 1648, 462185. [Google Scholar] [CrossRef] [PubMed]
- Palyzova, A.; Rezanka, T. Separation and identification of diacylglycerols containing branched chain fatty acids by liquid chromatography-mass spectrometry. J. Chromatogr. A 2021, 1635, 461708. [Google Scholar] [CrossRef]
- Palyzova, A.; Rezanka, T. Enantiomeric separation of triacylglycerols containing fatty acids with a ring (cyclofatty acids). J. Chromatogr. A 2020, 1622, 461103. [Google Scholar] [CrossRef] [PubMed]
- Palyzova, A.; Rezanka, T. Separation of triacylglycerols containing allenic and acetylenic fatty acids by enantiomeric liquid chromatography-mass spectrometry. J. Chromatogr. A 2020, 1623, 461161. [Google Scholar] [CrossRef] [PubMed]
- Palyzova, A.; Maresova, H.; Novak, J.; Zahradnik, J.; Rezanka, T. Effect of the anti-inflammatory drug diclofenac on lipid composition of bacterial strain Raoultella sp. KDF8. Folia Microbiol. 2020, 65, 763–773. [Google Scholar] [CrossRef]
- Vitova, M.; Goecke, F.; Sigler, K.; Rezanka, T. Lipidomic analysis of the extremophilic red alga Galdieria sulphuraria in response to changes in pH. Algal Res. Biomass Biofuels Bioprod. 2016, 13, 218–226. [Google Scholar] [CrossRef]
- Vitova, M.; Bisova, K.; Hlavova, M.; Zachleder, V.; Rucki, M.; Cizkova, M. Glutathione peroxidase activity in the selenium-treated alga Scenedesmus quadricauda. Aquat. Toxicol. 2011, 102, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Rezanka, T.; Kolouchova, I.; Gharwalova, L.; Palyzova, A.; Sigler, K. Identification and characterization of phospholipids with very long chain fatty acids in brewer’s yeast. Lipids 2017, 52, 1007–1017. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar]
- Kiene, R.P.; Visscher, P.T.; Keller, M.D.; Kirst, G.O. Biosynthetic pathways for phosphatidylsulfocholine, the sulfonium analogue of phosphatidylcholine, in Diatoms. In Biological and Environmental Chemistry of DMSP and Related Sulfonium Compounds; Springer: Boston, MA, USA, 1996; pp. 109–119. [Google Scholar]
- Alves, S.P.; Santos-Silva, J.; Cabrita, A.R.J.; Fonseca, A.J.M.; Bessa, R.J.B. Detailed dimethylacetal and fatty acid composition of rumen content from lambs fed lucerne or concentrate supplemented with soybean oil. PLoS ONE 2013, 8, e58386. [Google Scholar] [CrossRef] [Green Version]
- Morrison, W.R.; Smith, L.M. Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride-methanol. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [CrossRef]
- Dobson, G.; Itabashi, Y.; Christie, W.W.; Robertson, G.W. Liquid chromatography with particle-beam electron-impact mass spectrometry of diacylglycerol nicotinates. Chem. Phys. Lipids 1998, 97, 27–39. [Google Scholar] [CrossRef]
- Destaillats, F.; Angers, P. One-step methodology for the synthesis of FA picolinyl esters from intact lipids. J. Am. Oil Chem. Soc. 2002, 79, 253–256. [Google Scholar] [CrossRef]
- Rezanka, T.; Vitova, M.; Lukavsky, J.; Sigler, K. Lipidomic study of precursors of Endocannabinoids in freshwater Bryozoan Pectinatella magnifica. Lipids 2018, 53, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Rezanka, T.; Rozentsvet, O. A Lipid composition of 3 macrophytes from the caspian sea. Phytochem 1993, 33, 1015–1019. [Google Scholar] [CrossRef]
- Rezanka, T.; Podojil, M. Identification of wax esters of the fresh water green alga Chlorella kessleri by gas-chromatography mass spectrometry. J. Chromatogr. 1986, 362, 399–406. [Google Scholar] [CrossRef]
- Yokota, K.; Kanamoto, R.; Kito, M. Composition of cardiolipin molecular species in Escherichia coli. J. Bacteriol. 1980, 141, 1047–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.N.; Siegenthaler, P.A. Phosphatidylglycerol molecular species of photosynthetic membranes analyzed by high-performance liquid chromatography: Theoretical considerations. Lipids 1996, 31, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Itabashi, Y.; Myher, J.J.; Kuksis, A. Determination of positional distribution of short-chain fatty acids in bovine milk fat on chiral columns. J. Am. Oil Chem. Soc. 1993, 70, 1177–1181. [Google Scholar] [CrossRef]
- Limb, J.K.; Kim, Y.H.; Han, S.Y.; Jhon, G.J. Isolation and characterization of monoacetyldiglycerides from bovine udder. J. Lipid Res. 1999, 40, 2169–2176. [Google Scholar] [CrossRef]
- Kalo, P.; Kemppinen, A.; Ollilainen, V.; Kuksis, A. Regiospecific determination of short-chain triacylglycerols in butterfat by normal-phase HPLC with on-line electrospray-tandem mass spectrometry. Lipids 2004, 39, 915–928. [Google Scholar] [CrossRef]
- Palyzová, A.; Cajthaml, T.; Řezanka, T. Separation of regioisomers and enantiomers of triacylglycerols containing branched fatty acids (iso and/or anteiso). Electrophoresis 2021, 42, 1832–1843. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Vizcaino, J.A.; Kofeler, H.; Trotzmuller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J.O. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, D.J.; Tiffany, J.M. Comparison of derivatives for the characterization of branched long-chain alcohols and 1,2-diols by mass spectrometry. Biomed. Mass Spectrom. 1984, 11, 353–359. [Google Scholar] [CrossRef]
- Vetter, W.; Meister, W. Nicotinates as derivatives for the mass spectrometry investigation of long chain alcohols. Org. Mass Spectrom. 1981, 16, 118–122. [Google Scholar] [CrossRef]
- Peterson, B.L.; Cummings, B.S. A review of chromatographic methods for the assessment of phospholipids in biological samples. Biomed. Chromatogr. 2006, 20, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Kim, G.H.; Wei, F.; Chen, H.; Altarejos, J.; Han, X. Improved method for quantitative analysis of methylated phosphatidylethanolamine species and its application for analysis of diabetic-mouse liver samples. Anal. Bioanal. Chem. 2015, 407, 5021–5032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murgia, S.; Mele, S.; Monduzzi, M. Quantitative characterization of phospholipids in milk fat via 31P NMR using a monophasic solvent mixture. Lipids 2003, 38, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Carman, G.M.; Zeimetz, G.M. Regulation of phospholipid biosynthesis in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 1996, 271, 13293–13296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ejsing, C.S.; Sampaio, J.L.; Surendranath, V.; Duchoslav, E.; Ekroos, K.; Klemm, R.W.; Simons, K.; Shevchenko, A. Global analysis of the yeast lipidome by quantitative shotgun mass spectrometry. Proc. Natl. Acad. Sci. USA 2009, 106, 2136–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.D.; Summers, P.S.; Weretilnyk, E.A. Phosphocholine synthesis in spinach: Characterization of phosphoethanolamine N-methyltransferase. Physiol. Plant. 2000, 108, 286–294. [Google Scholar] [CrossRef]
- Hirashima, T.; Toyoshima, M.; Moriyama, T.; Sato, N. Evolution of the phosphatidylcholine biosynthesis pathways in green algae: Combinatorial diversity of methyltransferases. J. Mol. Evol. 2018, 86, 68–76. [Google Scholar] [CrossRef]
- Aveiro, S.S.; Melo, T.; Figueiredo, A.; Domingues, P.; Pereira, H.; Maia, I.B.; Silva, J.; Domingues, M.R.; Nunes, C.; Moreira, A.S.P. The polar lipidome of cultured Emiliania huxleyi: A source of bioactive lipids with relevance for biotechnological applications. Biomolecules 2020, 10, 1434. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.R.; Wilmes, P.; Bowen, B.P.; Northen, T.R.; Banfield, J.F. Deuterium-exchange metabolomics identifies N-methyl lyso phosphatidylethanolamines as abundant lipids in acidophilic mixed microbial communities. Metabolomics 2012, 8, 566–578. [Google Scholar] [CrossRef] [Green Version]
- Choma, A.; Komaniecka, I. The polar lipid composition of Mesorhizobium ciceri. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2003, 1631, 188–196. [Google Scholar] [CrossRef]
- Yamashita, S.; Abe, A.; Nakagawa, K.; Kinoshita, M.; Miyazawa, T. Separation and detection of plasmalogen in marine invertebrates by high-performance liquid chromatography with evaporative light-scattering detection. Lipids 2014, 49, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Jenkins, C.M.; Dilthey, B.; Gross, R.W. Multidimensional mass spectrometry-based shotgun lipidomics analysis of vinyl ether diglycerides. Anal. Bioanal. Chem. 2015, 407, 5199–5210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, Y.K.; Park, E.J.; Lee, C.W.; Joo, H.T.; Farkas, T. Ether lipid composition and molecular species alterations in carp brain (Cyprinus carpio L.) during normoxic temperature acclimation. Neurochem. Res. 1997, 22, 1257–1264. [Google Scholar] [CrossRef]
- Yamashita, S.; Kanno, S.; Honjo, A.; Otoki, Y.; Nakagawa, K.; Kinoshita, M.; Miyazawa, T. Analysis of plasmalogen species in foodstuffs. Lipids 2016, 51, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Cahn, R.S.; Ingold, C.; Prelog, V. Specification of molecular chirality. Angew. Chem. Int. Ed. 1966, 5, 385–415. [Google Scholar] [CrossRef]
- Rezanka, T.; Kolouchova, I.; Nedbalova, L.; Sigler, K. Enantiomeric separation of triacylglycerols containing very long chain fatty acids. J. Chromatogr. A 2018, 1557, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.; Buchet, R. Symmetry breaking of phospholipids. Symmetry 2020, 12, 1488. [Google Scholar] [CrossRef]
- Vance, D.; Walkey, C.; Agellon, L. Why has phosphatidylethanolamine N-methyltransferase survived in evolution? Biochem. Soc. Trans. 1998, 26, 337–340. [Google Scholar] [CrossRef]
- Kleetz, J.; Vasilopoulos, G.; Czolkoss, S.; Aktas, M.; Narberhaus, F. Recombinant and endogenous ways to produce methylated phospholipids in Escherichia coli. Appl. Microb. Cell Physiol. 2021, 105, 8837–8851. [Google Scholar] [CrossRef]
- Wachterhauser, G. From pre-cells to Eukarya-a tale of two lipids. Mol. Microbiol. 2003, 47, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Shimada, H.; Yamagishi, A. Stability of heterochiral hybrid membrane made of bacterial sn-G3P lipids and archaeal sn-G1P lipids. Biochemistry 2011, 50, 4114–4120. [Google Scholar] [CrossRef]
- Caforio, A.; Siliakus, M.F.; Exterkate, M.; Jain, S.; Jumde, V.R.; Andringa, R.L.H.; Kengen, S.W.M.; Minnaard, A.J.; Driessen, A.J.M.; van der Oost, J. Converting Escherichia coli into an archaebacterium with a hybrid heterochiral membrane. Proc. Natl. Acad. Sci. USA 2018, 115, 3704–3709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezanka, T. Polyunsaturated and unusual fatty acids from slime moulds. Phytochemistry 1993, 33, 1441–1444. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rezanka, T.; Bychek, I.A. Fatty acids and phospholipids from lichens of the order Lecanorales. Phytochemistry 1992, 31, 851–853. [Google Scholar] [CrossRef]
- Farquhar, J.W. Identification and gas-liquid chromatographic behavior of plasmalogen aldehydes and their acetal, alcohol, and acetylated alcohol derivatives. J. Lipid Res. 1962, 3, 21–30. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Řezanka, T.; Palyzová, A.; Vítová, M.; Brányik, T.; Kulišová, M.; Irena, J.K. Structural Characterization of Mono- and Dimethylphosphatidylethanolamines from Various Organisms Using a Complex Analytical Strategy including Chiral Chromatography. Symmetry 2022, 14, 616. https://doi.org/10.3390/sym14030616
Řezanka T, Palyzová A, Vítová M, Brányik T, Kulišová M, Irena JK. Structural Characterization of Mono- and Dimethylphosphatidylethanolamines from Various Organisms Using a Complex Analytical Strategy including Chiral Chromatography. Symmetry. 2022; 14(3):616. https://doi.org/10.3390/sym14030616
Chicago/Turabian StyleŘezanka, Tomáš, Andrea Palyzová, Milada Vítová, Tomáš Brányik, Markéta Kulišová, and Jarošová Kolouchová Irena. 2022. "Structural Characterization of Mono- and Dimethylphosphatidylethanolamines from Various Organisms Using a Complex Analytical Strategy including Chiral Chromatography" Symmetry 14, no. 3: 616. https://doi.org/10.3390/sym14030616