
Environmental Effects on Normalized Gross Primary Productivity in Beech and Norway Spruce Forests

 ,
,  ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sites Description
2.2. Eddy Covariance and Ancillary Measurements
2.3. Soil Water Content Simulations
2.4. Data Processing and Analysis
2.4.1. Turbulent Flux Measurements
2.4.2. Estimation of Potential and Normalized GPP
- Compute the 95th percentile from the daily sum of GPP pooled over years 2012–2016 for each DOY over a 7-day window (applied iteratively);
- Outliers were detected and removed based on the percentiles method (all observations that positioned outside the interval formed by the 1 and 99 percentiles were considered as outliers).
2.4.3. Multi-Linear and Tree-Based Regression Model Analyses
2.4.4. Accuracy Test of Regression Models
3. Results
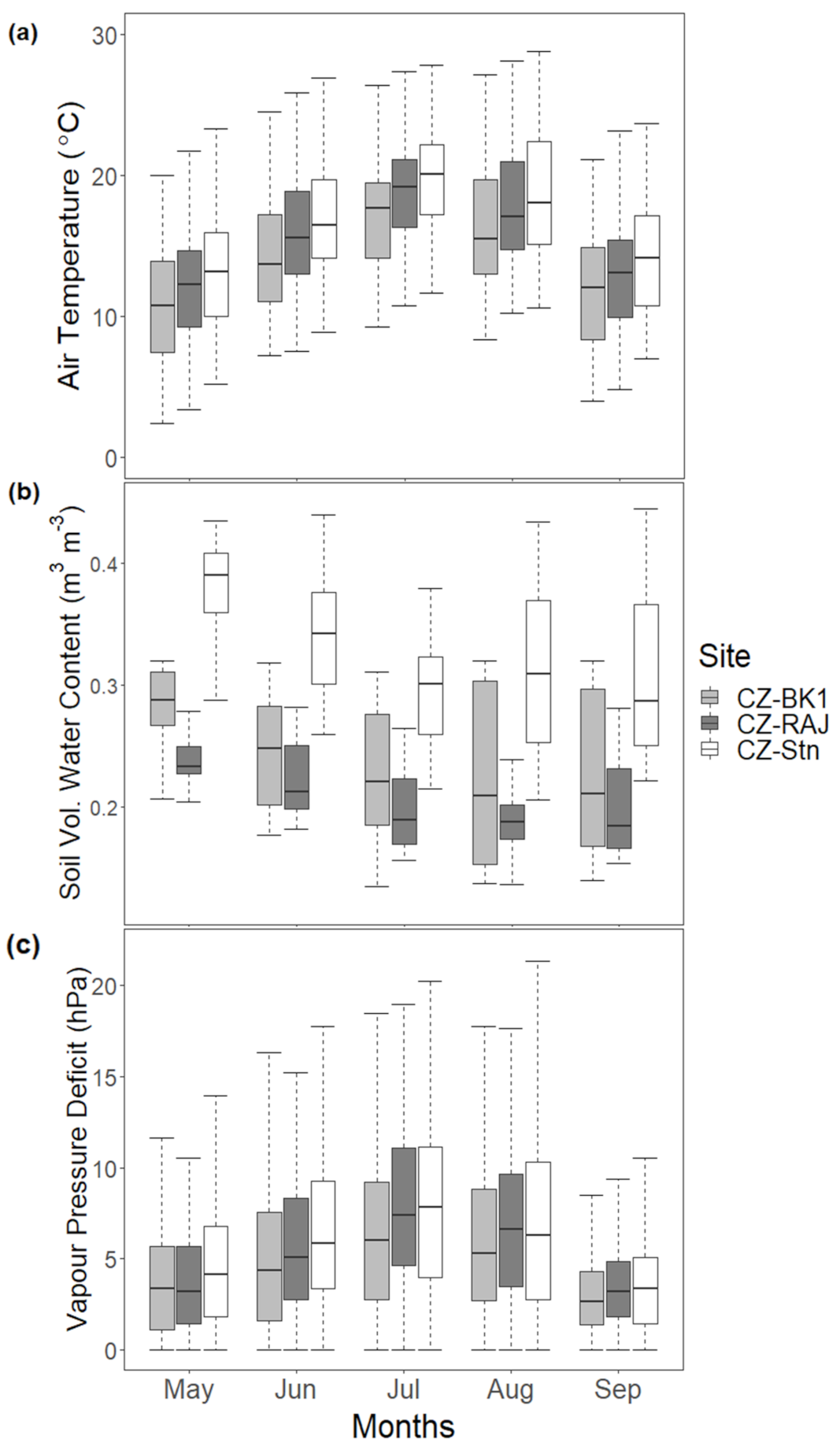
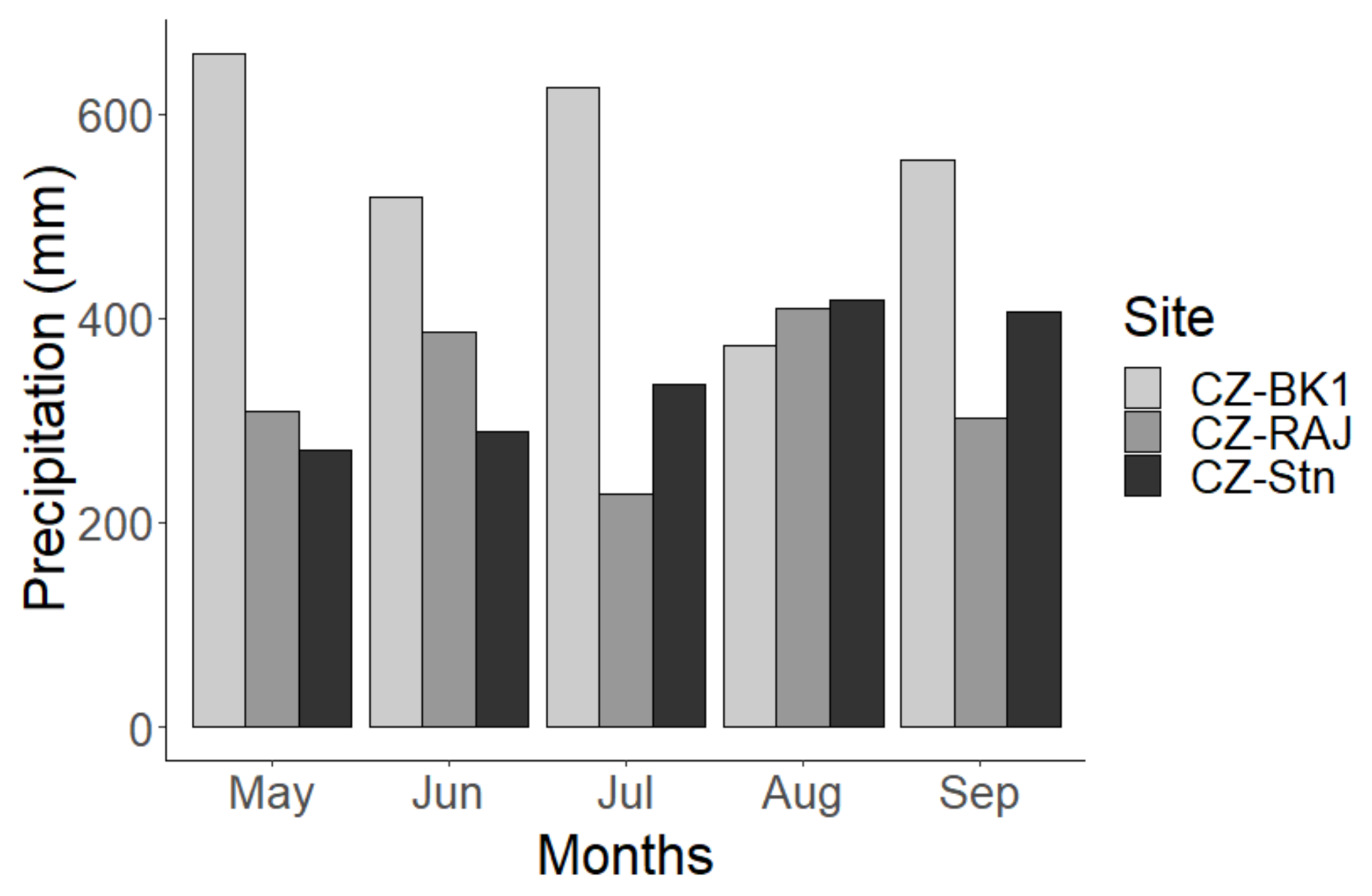
3.1. Variation in Meteorological Conditions at the Experimental Stations
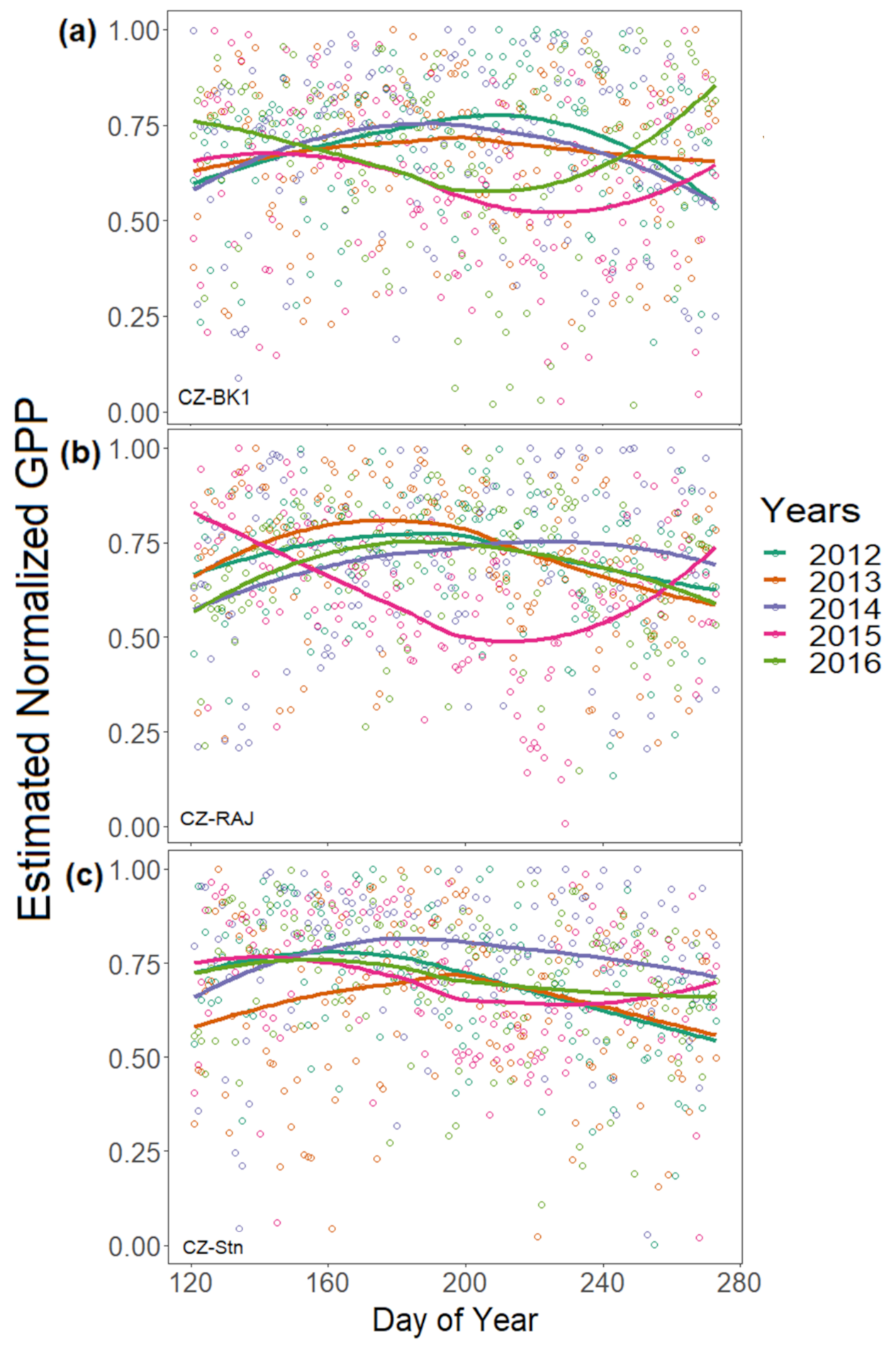
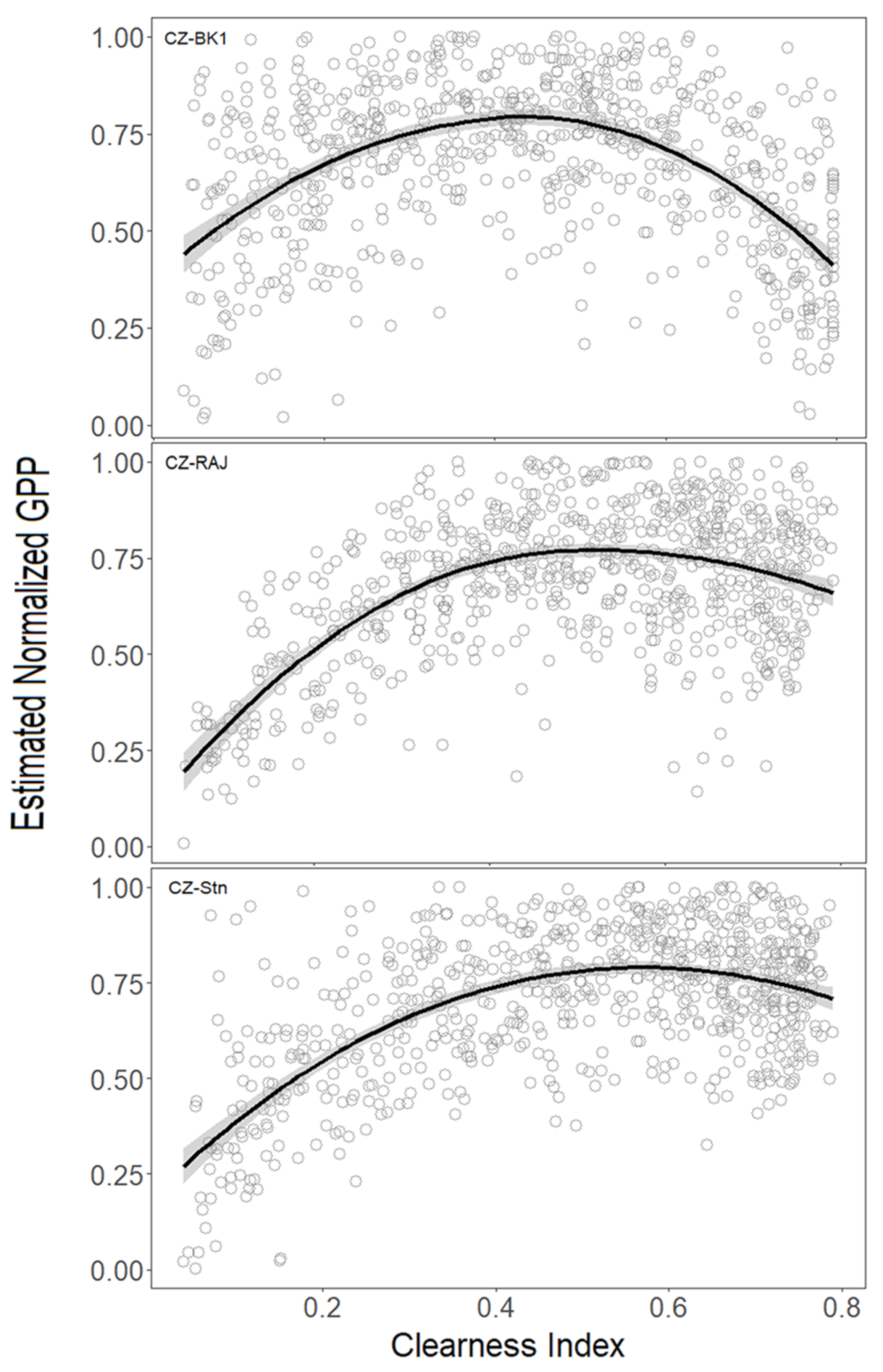
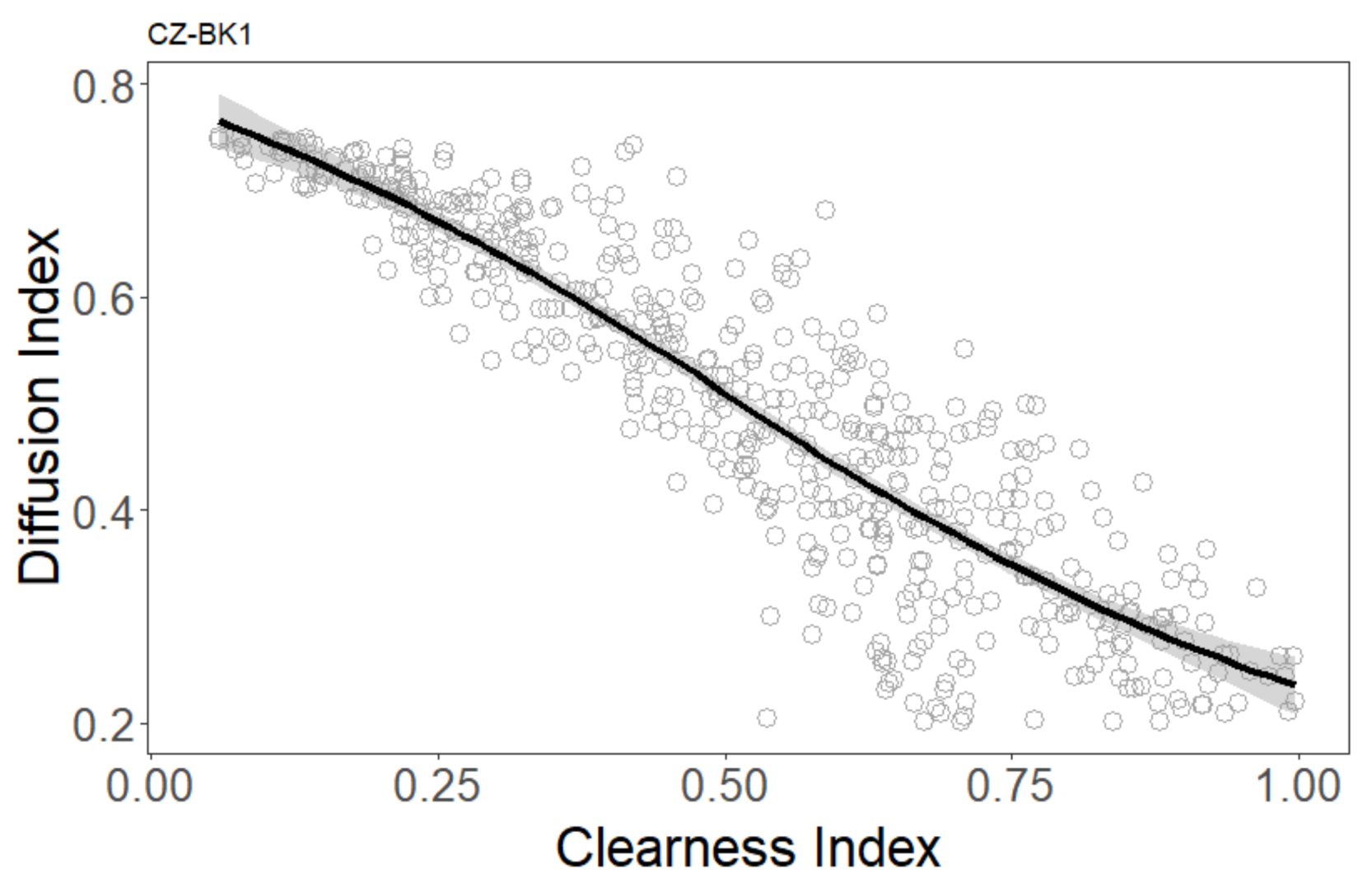
3.2. Effect of Meteorological Conditions on GPP
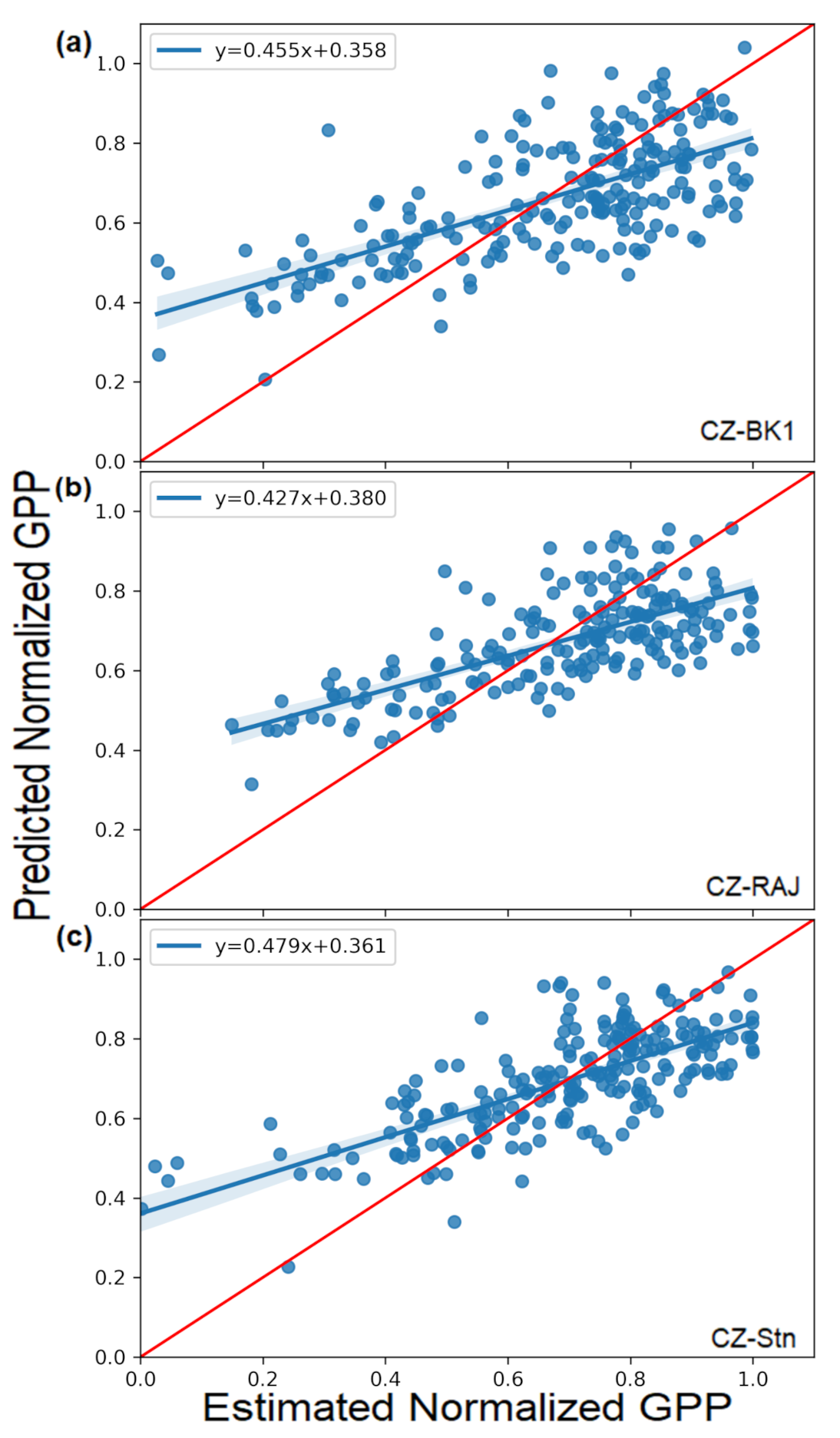
3.2.1. GPPnorm Prediction through Stepwise Multi-Linear Regression (SMLR)
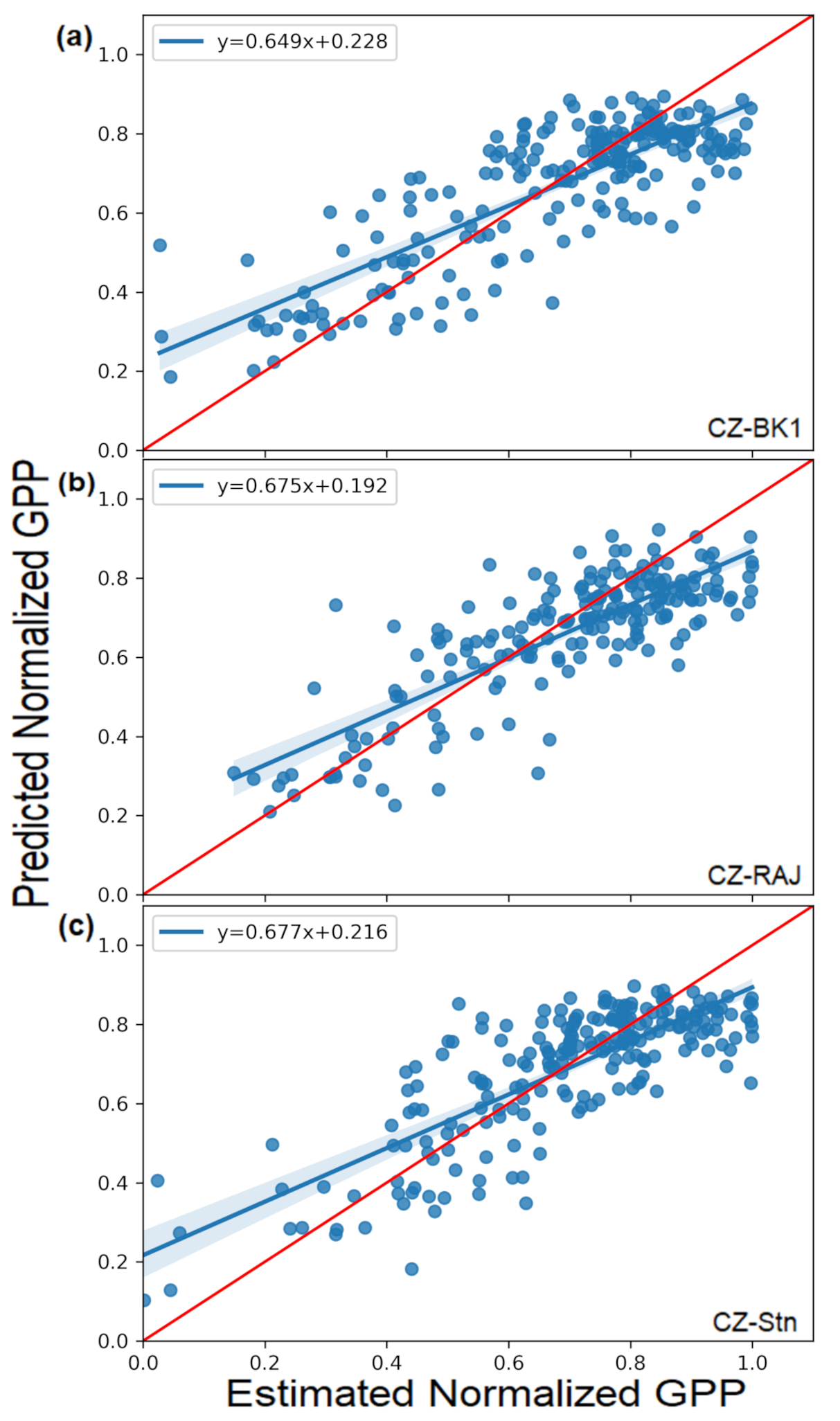
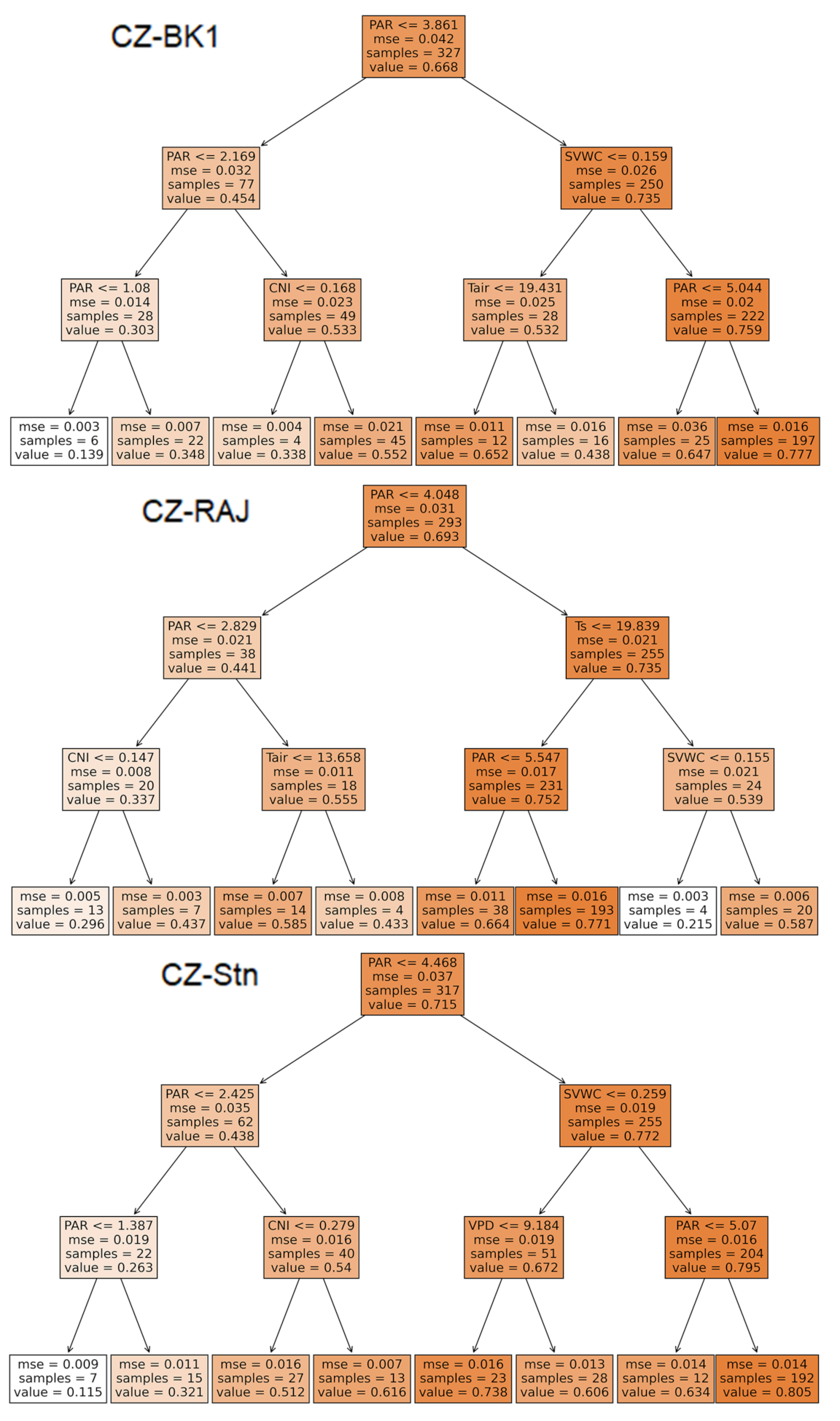
3.2.2. GPPnorm Prediction through Random Forest Analyses (RF)
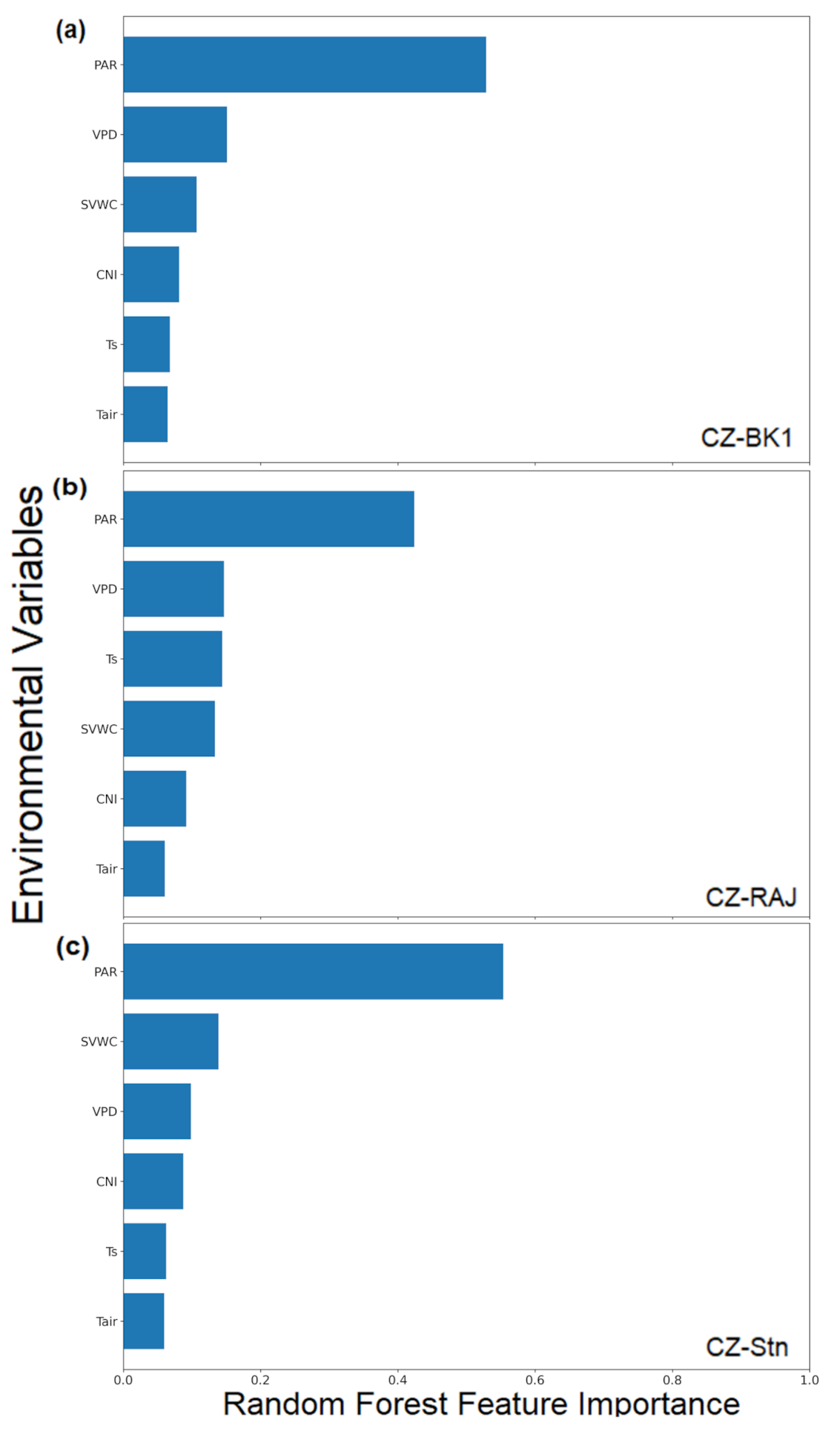
3.2.3. Importance of Environmental Variables in Random Forest Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | CZ-BK1 | CZ-RAJ | CZ-Stn |
|---|---|---|---|
| Ultrasonic Anemometer | |||
| Instrument | Gill HS-50 (Gill Instruments, UK) | Gill R3-100 (Gill Instruments, UK) but later changed to Gill HS-50 on 5 June 2015 | Gill R3-100 (Gill Instruments, UK) |
| Gas Analyser | |||
| Instrument | LI-7200 | Initially LI-7000 (IRG-0226) closed-path gas analyser, but later changed to LI-7200 on 5 June 2015 | LI-7000 closed-path gas analyser |
| Measurement Height for the Eddy covaiance set-up (m) | Initially 20.5 m, but later changed to 25 m on 7 June 2016 | 41 m | 44 m |
| Site Name | CZ-BK1 | CZ-RAJ | CZ-Stn |
|---|---|---|---|
| Air Temperature and Humidity Profile | |||
| Instrument | EMS33 temperature and humidity sensor | EMS33 temperature and humidity sensor | EMS33 temperature and humidity sensor |
| Height (m) | 2.0, 7.6, 12.6, 13.5, 14.3, 14.8, 15.4, 16.5, 18.7 | 2.0, 11.0, 23.0, 29.0, 35.0, 42.0 | 2.0, 12.0, 22.0, 28.0, 33.0, 38.0, 44.0 |
| Net Radiation | |||
| Instrument | CNR1 net radiometer (Kipp & Zonen) | CNR1 net radiometer (Kipp & Zonen) | CNR1 net radiometer (Kipp & Zonen) |
| Height (m) | Initially 20 m, then changed to 22 m in August, 2013 | 42 m | 42 m |
| Soil Temperature | |||
| Instrument | Pt 1000 | Pt 100 | Pt 1000 |
| Depth (m) | 0, 0.05, 0.10, 0.20, 0.30, 0.50 | 0, 0.05 | 0, 0.05, 0.10, 0.20, 0.30, 0.50 |
| Estimate | Standard Error | t Value | Pr (>|t|) | |
|---|---|---|---|---|
| (Intercept) | −4.581 | *** | ||
| PAR | 10.772 | < *** | ||
| PAR2 | 8.601 | < *** | ||
| SVWC2 | 1.938 | −5.649 | *** | |
| PAR:Tair | −5.536 | *** | ||
| SVWC | 5.305 | *** | ||
| Ts:PAR | 4.598 | *** | ||
| Ts2 | −4.155 | *** | ||
| VPD:SVWC | 3.972 | *** | ||
| CNI2 | −3.533 | 0.000437 *** | ||
| CNI | 3.461 | 0.000571 *** | ||
| VPD2 | −3.128 | 0.001830 *** | ||
| VPD | −2.456 | 0.014300 * | ||
| Ts | 2.367 | 0.018214 * | ||
| VPD:PAR | 2.264 | 0.023906 * | ||
| CNI:Tair | 2.128 | 0.033690 * | ||
| CNI:PAR | −2.039 | 0.041772 * | ||
| VPD:Tair | 1.976 | 0.048507 * |
| Estimate | Standard Error | t Value | Pr (> |t|) | |
|---|---|---|---|---|
| (Intercept) | 1.2416627 | 0.2834895 | 4.380 | *** |
| PAR | 0.1957506 | 0.0157165 | 12.455 | < *** |
| PAR2 | −0.0055807 | 0.0006615 | −8.436 | < *** |
| Ts:SVWC | 0.2931591 | 0.0448089 | 6.542 | *** |
| Tair2 | −0.0009479 | 0.0001752 | −5.409 | *** |
| CNI2 | −0.9090371 | 0.1592056 | −5.710 | *** |
| Ts2 | −0.0015452 | 0.0002921 | −5.290 | *** |
| SVWC:CNI | 4.0751744 | 0.8162736 | 4.992 | *** |
| Ts:Tair | 0.0016139 | 0.0003835 | 4.208 | *** |
| PAR:SVWC | −0.352378 | 0.0896162 | −3.932 | *** |
| SVWC | −8.4537169 | 2.2630082 | −3.736 | 0.000204 *** |
| VPD | −0.0364140 | 0.0102154 | −3.565 | 0.000392 *** |
| Ts | 0.0404096 | 0.0149492 | −2.703 | 0.007053 *** |
| VPD:SVWC | 0.1377881 | 0.0518878 | 2.656 | 0.008118 *** |
| SVWC2 | 10.9036492 | 5.0661812 | 2.152 | 0.031754 * |
| Estimate | Standard Error | t Value | Pr (> |t|) | |
|---|---|---|---|---|
| (Intercept) | 1.6324051 | 0.2913214 | 5.603 | *** |
| PAR | 0.1035450 | 0.0089856 | 11.523 | < *** |
| Ts:SVWC | 0.4954805 | 0.0627527 | 7.896 | *** |
| Ts | −0.1794898 | 0.0233970 | −7.671 | *** |
| CNI | 0.8213389 | 0.13378886 | 5.957 | *** |
| Tair | 0.0988082 | 0.0178532 | 5.534 | *** |
| SVWC | −6.3883130 | 1.3261064 | −4.817 | *** |
| Tair:SVWC | −0.2274474 | 0.0467787 | −4.862 | *** |
| Tair:PAR | −0.0024049 | 0.0005103 | −4.713 | *** |
| CNI:PAR | −0.0558883 | 0.0148897 | −3.753 | 0.000189 *** |
| VPD2 | −0.0008560 | 0.0002309 | −3.708 | 0.000225 *** |
| VPD:Ts | 0.0021074 | 0.0006005 | 3.510 | 0.000477 *** |
| VPD | −0.0561466 | 0.0176605 | −3.179 | 0.001540 *** |
| SVWC2 | 4.9409576 | 1.5641808 | 3.159 | 0.001651 *** |
| VPD:SVWC | 0.1048877 | 0.0339888 | 3.086 | 0.002107 *** |
| CNI2 | −0.5376705 | 0.2101539 | −2.558 | 0.010718 *** |
References
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Lal, R.; Smith, P.; Jungkunst, H.F.; Mitsch, W.J.; Lehmann, J.; Nair, P.R.; McBratney, A.B.; de Moraes Sa, J.C.; Schneider, J.; Zinn, Y.L.; et al. The carbon sequestration potential of terrestrial ecosystems. J. Soil Water Conserv. 2018, 73, 145A–152A. [Google Scholar] [CrossRef] [Green Version]
- Murthy, I.K.; Varghese, V.; Prasad, K.D. Competing Demands on Land: Implications for Carbon Sink Enhancement and Potential of Forest Sector in Karnataka to Contribute to the INDC Forest Goal of India; KVD Prasad: Telangana, India, 2019. [Google Scholar]
- Nayak, R.K.; Patel, N.R.; Dadhwal, V.K. Estimation and analysis of terrestrial net primary productivity over India by remote-sensing-driven terrestrial biosphere model. Environ. Monit. Assess. 2010, 170, 195–213. [Google Scholar] [CrossRef]
- Marek, M.V.; Janouš, D.; Taufarová, K.; Havránková, K.; Pavelka, M.; Kaplan, V.; Marková, I. Carbon exchange between ecosystems and atmosphere in the Czech Republic is affected by climate factors. Environ. Pollut. 2011, 159, 1035–1039. [Google Scholar] [CrossRef]
- Letchov, G. Carbon-use efficiency of terrestrial ecosystems under stress conditions in South East Europe (MODIS, NASA). Multidiscip. Digit. Publ. Inst. Proc. 2018, 2, 363. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.; Muûls, M.; De Preux, L.B.; Wagner, U.J. On the empirical content of carbon leakage criteria in the EU Emissions Trading Scheme. Ecol. Econ. 2014, 105, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zha, T.S.; Jia, X.; Wu, B.; Zhang, Y.Q.; Qin, S.G. Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem. Biogeosciences 2014, 11, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Pingintha, N.; Leclerc, M.Y.; Beasley, J.P., Jr.; Durden, D.; Zhang, G.; Senthong, C.; Rowland, D. Hysteresis response of daytime net ecosystem exchange during drought. Biogeosciences 2010, 7, 1159–1170. [Google Scholar] [CrossRef] [Green Version]
- Taufarová, K.; Havránková, K.; Dvorská, A.; Pavelka, M.; Urbaniak, M.; Janouš, D. Forest ecosystem as a source of CO2 during growing season: Relation to weather conditions. Int. Agrophys. 2014, 28, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Raj, R.; Bayat, B.; Lukeš, P.; Šigut, L.; Homolová, L. Analyzing Daily Estimation of Forest Gross Primary Production Based on Harmonized Landsat-8 and Sentinel-2 Product Using SCOPE Process-Based Model. Remote Sens. 2020, 12, 3773. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, Y.; Yin, L.; Wenninger, J.; Zhang, J.; Hou, G.; Zhang, E.; Uhlenbrook, S. Climatic controls on sap flow dynamics and used water sources of Salix psammophila in a semi-arid environment in northwest China. Environ. Earth Sci. 2015, 73, 289–301. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Madani, N.; Kimball, J.S.; Ballantyne, A.P.; Affleck, D.L.; Van Bodegom, P.M.; Reich, P.B.; Kattge, J.; Sala, A.; Nazeri, M.; Jones, M.O.; et al. Future global productivity will be affected by plant trait response to climate. Sci. Rep. 2018, 8, 2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Ciais, P.; Bastos, A.; Stoy, P.C.; Yang, H.; Green, J.K.; Wang, B.; Yu, K.; Huang, Y.; Knohl, A.; et al. Sensitivity of gross primary productivity to climatic drivers during the summer drought of 2018 in Europe. Philos. Trans. R. Soc. B 2020, 375, 20190747. [Google Scholar] [CrossRef]
- Urban, O.; Janouš, D.; Acosta, M.; Czerný, R.; Markova, I.; Navratil, M.; Pavelka, M.; Pokorný, R.; Šprtová, M.; Zhang, R.; et al. Ecophysiological controls over the net ecosystem exchange of mountain spruce stand. Comparison of the response in direct vs. diffuse solar radiation. Glob. Chang. Biol. 2007, 13, 157–168. [Google Scholar] [CrossRef]
- Urban, O.; Klem, K.; Ač, A.; Havránková, K.; Holišová, P.; Navrátil, M.; Zitová, M.; Kozlová, K.; Pokorný, R.; Šprtová, M.; et al. Impact of clear and cloudy sky conditions on the vertical distribution of photosynthetic CO2 uptake within a spruce canopy. Funct. Ecol. 2012, 26, 46–55. [Google Scholar] [CrossRef]
- Zhang, M.; Yu, G.R.; Zhuang, J.; Gentry, R.; Fu, Y.L.; Sun, X.M.; Zhang, L.M.; Wen, X.F.; Wang, Q.F.; Han, S.J.; et al. Effects of cloudiness change on net ecosystem exchange, light use efficiency, and water use efficiency in typical ecosystems of China. Agric. For. Meteorol. 2011, 151, 803–816. [Google Scholar] [CrossRef]
- Knohl, A.; Baldocchi, D.D. Effects of diffuse radiation on canopy gas exchange processes in a forest ecosystem. J. Abbr. 2008, 113. [Google Scholar] [CrossRef]
- Mette, T.; Dolos, K.; Meinardus, C.; Bräuning, A.; Reineking, B.; Blaschke, M.; Pretzsch, H.; Beierkuhnlein, C.; Gohlke, A.; Wellstein, C. Climatic turning point for beech and oak under climate change in Central Europe. Ecosphere 2013, 4, 1–19. [Google Scholar] [CrossRef]
- Krupková, L.; Havránková, K.; Krejza, J.; Sedlák, P.; Marek, M.V. Impact of water scarcity on spruce and beech forests. J. For. Res. 2018, 30, 899–909. [Google Scholar] [CrossRef]
- Spiecker, H.; Hansen, J.; Klimo, E.; Skovsgaard, J.; Sterba, H.; von Teuffel, K. Norway Spruce Conversion: Options and Consequences; Research Report 18: S. Brill, Leiden; European Forest Institute: Boston, MA, USA, 2004. [Google Scholar]
- Hanewinkel, M.; Hummel, S.; Cullmann, D.A. Modelling and economic evaluation of forest biome shifts under climate change in Southwest Germany. For. Ecol. Manag. 2010, 259, 710–719. [Google Scholar] [CrossRef]
- Hlásny, T.; Turčáni, M. Persisting bark beetle outbreak indicates the unsustainability of secondary Norway spruce forests: Case study from Central Europe. Ann. For. Sci. 2013, 70, 481–491. [Google Scholar] [CrossRef] [Green Version]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Jocher, G.; Fischer, M.; Šigut, L.; Pavelka, M.; Sedlák, P.; Katul, G. Assessing decoupling of above and below canopy air masses at a Norway spruce stand in complex terrain. Agric. For. Meteorol. 2020, 294, 108149. [Google Scholar] [CrossRef]
- McGloin, R.; Šigut, L.; Fischer, M.; Foltýnová, L.; Chawla, S.; Trnka, M.; Pavelka, M.; Marek, M.V. Available energy partitioning during drought at two Norway spruce forests and a European Beech forest in Central Europe. J. Geophys. Res. Atmos. 2019, 124, 3726–3742. [Google Scholar] [CrossRef]
- Sánchez, G.; Serrano, A.; Cancillo, M.L. Effect of cloudiness on solar global, solar diffuse and terrestrial downward radiation at Badajoz (Southwestern Spain). Opt. Pura Appl. 2012, 45, 33–38. [Google Scholar] [CrossRef]
- Monteith, J.; Unsworth, M. Principles of Environmental Physics: Plants, Animals, and the Atmosphere; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Thimijan, R.W.; Heins, R.D. Photometric, radiometric, and quantum light units of measure: A review of procedures for interconversion. HortScience 1983, 18, 818–822. [Google Scholar]
- Mensah, C.; Šigut, L.; Fischer, M.; Foltýnová, L.; Jocher, G.; Acosta, M.; Kowalska, N.; Kokrda, L.; Pavelka, M.; Marshall, J.D.; et al. Water requirements of short rotation poplar coppice: Experimental and modelling analyses across Europe. Agric. For. Meteorol. 2018, 250, 343–360. [Google Scholar]
- Hartig, F.; Minunno, F.; Paul, S. BayesianTools: General-Purpose MCMC and SMC Samplers and Tools for Bayesian Statistics. R Package Version 0.1.6. 2019. Available online: https://CRAN.R-project.org/package=BayesianTools (accessed on 20 March 2021).
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Brooks, S.P.; Gelman, A. General methods for monitoring convergence of iterative simulations. J. Comput. Graph. Stat. 1998, 7, 434–455. [Google Scholar]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Oren, R.; Sperry, J.S.; Katul, G.G.; Pataki, D.E.; Ewers, B.E.; Phillips, N.; Scháfer, K.V.R. Survey and synthesis of intra-and interspecific variation in stomatal sensitivity to vapour pressure deficit. Plant Cell Environ. 1999, 22, 1515–1526. [Google Scholar] [CrossRef] [Green Version]
- Aubinet, M.; Vesala, T.; Papale, D. (Eds.) Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Springer Science and Business Media: Berlin, Germany, 2012. [Google Scholar]
- Foken, T.; Leuning, R.; Oncley, S.R.; Mauder, M.; Aubinet, M. Corrections and data quality control. In Eddy Covariance; Springer: Dordrecht, The Netherlands, 2012; pp. 85–131. [Google Scholar]
- Wilczak, J.M.; Oncley, S.P.; Stage, S.A. Sonic anemometer tilt correction algorithms. Bound.-Layer Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- Moncrieff, J.B.; Jarvis, P.G.; Valentini, R. Canopy fluxes. In Methods in Ecosystem Science; Springer: New York, NY, USA, 2000; pp. 161–180. [Google Scholar]
- Ibrom, A.; Dellwik, E.; Larsen, S.E.; Pilegaard, K.I.M. On the use of the Webb-Pearman-Leuning theory for closed-path eddy correlation measurements. Tellus B Chem. Phys. Meteorol. 2007, 59, 937–946. [Google Scholar] [CrossRef]
- Horst, T.W.; Lenschow, D.H. Attenuation of scalar fluxes measured with spatially-displaced sensors. Bound.-Layer Meteorol. 2009, 130, 275–300. [Google Scholar] [CrossRef]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef] [Green Version]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- McGloin, R.; Šigut, L.; Havránková, K.; Dušek, J.; Pavelka, M.; Sedlák, P. Energy balance closure at a variety of ecosystems in Central Europe with contrasting topographies. Agric. For. Meteorol. 2018, 248, 418–431. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 22 January 2021).
- Gu, L.; Post, W.M.; Baldocchi, D.D.; Black, T.A.; Suyker, A.E.; Verma, S.B.; Vesala, T.; Wofsy, S.C. Characterizing the seasonal dynamics of plant community photosynthesis across a range of vegetation types. In Phenology of Ecosystem Processes; Springer: New York, NY, USA, 2009; pp. 35–58. [Google Scholar]
- Ripley, B. Choose a Model by AIC in a Stepwise Algorithm; R Documentation; AIC: Westborough, MA, USA, 2015. [Google Scholar]
- Liaw, A.; Wiener, M. Breiman and Cutler’s Random Forests for Classification and Regression Version (4.6–12); R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Breiman, L. Breiman and Cutler’s Random Forests for Classification and Regression: Package ’randomForest’; Institute for Statistics and Mathematics, Vienna University of Economics and Business: Vienna, Austria, 2013. [Google Scholar]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529. [Google Scholar] [CrossRef]
- Chapin, F.S.; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer Science & Business Media: Berlin, Germany, 2011. [Google Scholar]
- Ågren, G.I.; Andersson, F.O. Terrestrial Ecosystem Ecology: Principles and Applications; Cambridge University Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Leuschner, C. Die Trockenheitsempfindlichkeit der Rotbuche vor dem Hintergrund des prognostizierten Klimawandels; Walter de Gruyter GmbH & Co. KG: Berlin, Germany, 2009. [Google Scholar]
- Nikolova, P.S.; Raspe, S.; Andersen, C.P.; Mainiero, R.; Blaschke, H.; Matyssek, R.; Häberle, K.H. Effects of the extreme drought in 2003 on soil respiration in a mixed forest. Eur. J. For. Res. 2009, 128, 87–98. [Google Scholar] [CrossRef]
- Pretzsch, H.; Grams, T.; Häberle, K.H.; Pritsch, K.; Bauerle, T.; Rötzer, T. Growth and mortality of Norway spruce and European beech in monospecific and mixed- species stands under natural episodic and experimentally extended drought; Results of the KROOF throughfall exclusion experiment. Trees 2020, 34, 957–970. [Google Scholar] [CrossRef]
- Dobrovolny, L. Density and spatial distribution of beech (Fagus sylvatica L.) regeneration in Norway spruce (Picea abies (L.) Karsten) stands in the central part of the Czech Republic. iForest-Biogeosci. For. 2016, 9, 666. [Google Scholar] [CrossRef] [Green Version]
- Krejza, J.; Cienciala, E.; Světlík, J.; Bellan, M.; Noyer, E.; Horáček, P.; Štěpánek, P.; Marek, M.V. Evidence of climate-induced stress of Norway spruce along elevation gradient preceding the current dieback in Central Europe. Trees 2021, 35, 103–119. [Google Scholar] [CrossRef]
- Nishimura, P.H.; Laroque, C.P. Observed continentality in radial growth–climate relationships in a twelve site network in western Labrador, Canada. Dendrochronologia 2011, 29, 17–23. [Google Scholar] [CrossRef]
- Zang, C.; Pretzsch, H.; Rothe, A. Size-dependent responses to summer drought in Scots pine, Norway spruce and common oak. Trees 2012, 26, 557–569. [Google Scholar] [CrossRef]
- Boden, S.; Kahle, H.P.; von Wilpert, K.; Spiecker, H. Resilience of Norway spruce (Picea abies (L.) Karst) growth to changing climatic conditions in Southwest Germany. For. Ecol. Manag. 2014, 315, 12–21. [Google Scholar] [CrossRef]
- Novick, K.A.; Ficklin, D.L.; Stoy, P.C.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N.; et al. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Chang. 2016, 6, 1023–1027. [Google Scholar] [CrossRef] [Green Version]
- Sulman, B.N.; Roman, D.T.; Yi, K.; Wang, L.; Phillips, R.P.; Novick, K.A. High atmospheric demand for water can limit forest carbon uptake and transpiration as severely as dry soil. Geophys. Res. Lett. 2016, 43, 9686–9695. [Google Scholar] [CrossRef]
- Kodrik, J.; Kodrik, M. Root biomass of beech as a factor influencing the wind tree stability. J. For. Sci 2002, 48, 549–564. [Google Scholar] [CrossRef]
- Lopes de Gerenyu, V.O.; Rozanova, L.N.; Kudeyarov, V.N. Effect of Soil Temperature and Moisture on CO2 Evolution Rate of Cultivated Phaeozem: Analyses of a Long-Term Field Experiment. Plant Soil Environ.-UZPI Czech Repub. 2005, 51, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Meyers, T.; Pallardy, S.G.; Hanson, P.J.; Yang, B.; Heuer, M.; Hosman, K.P.; Riggs, J.S.; Sluss, D.; Wullschleger, S.D. Direct and indirect effects of atmospheric conditions and soil moisture on surface energy partitioning revealed by a prolonged drought at a temperate forest site. J. Geophys. Res. Atmos. 2006, 111. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Novak, M.D.; Gaumont-Guay, D.A.; Nesic, Z. Effect of soil water stress on soil respiration and its temperature sensitivity in an 18-year-old temperate Douglas-fir stand. Glob. Chang. Biol. 2008, 14, 1305–1318. [Google Scholar] [CrossRef]
- Gu, L.; Meyers, T.; Pallardy, S.G.; Hanson, P.J.; Yang, B.; Heuer, M.; Hosman, K.P.; Riggs, J.S.; Sluss, D.; Wullschleger, S.D. Climatic drivers of forest productivity in Central Europe. Agric. For. Meteorol. 2002, 234, 258–273. [Google Scholar]
- Mensah, C.; Šigut, L.; Fischer, M.; Foltýnová, L.; Jocher, G.; Acosta, M.; Kowalska, N.; Kokrda, L.; Pavelka, M.; Marshall, J.D.; et al. Assessing the Contrasting Effects of the Exceptional 2015 Drought on the Carbon Dynamics in Two Norway Spruce Forest Ecosystems. Atmosphere 2021, 12, 988. [Google Scholar] [CrossRef]
- Leuchner, M.; Hertel, C.; Rötzer, T.; Seifert, T.; Weigt, R.; Werner, H.; Menzel, A. Solar radiation as a driver for growth and competition in forest stands. In Growth and Defence in Plants; Springer: Berlin/Heidelberg, Germany, 2012; pp. 175–191. [Google Scholar]
- Čater, M.; Diaci, J. Divergent response of European beech, silver fir and Norway spruce advance regeneration to increased light levels following natural disturbance. For. Ecol. Manag. 2017, 399, 206–212. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Ač, A.; Marek, M.V.; Kalina, J.; Urban, O. Differences in pigment composition, photosynthetic rates and chlorophyll fluorescence images of sun and shade leaves of four tree species. Plant Physiol. Biochem. 2007, 45, 577–588. [Google Scholar] [CrossRef]







| Site Name | CZ-BK1 | CZ-RAJ | CZ-Stn |
|---|---|---|---|
| Location | Moravian-Silesian Beskids Mountains | Drahany Highland | White Carpathian Mountains |
| Coordinates | 493008 N, 183213 E | 492637 N, 164148 E | 490209 N, 175812 E |
| Elevation (in m a.s.l) | 875 | 625 | 540 |
| Topography | Mountainous (13 slope with SSW exposure, located close to a mountain ridge) | Hilly (5 slope with NNE exposure) | Mountainous (10 slope with WSW exposure) |
| Ecosystem Type | Coniferous evergreen forest | Coniferous evergreen forest | Deciduous broadleaf forest |
| Prevailing species | Norway spruce (Picea abies (L.) Karst.) | Norway spruce (Picea abies (L.) Karst.) | European beech (Fagus sylvatica L.) |
| Canopy height (m) | 16 (mean, as of 2015) | 33 (mean, as of 2015) | 31 (mean, as of 2015) |
| Stand age (years) | 35 (as of 2016) | 113 (as of 2016) | 115 (as of 2016) |
| Mean seasonal air temperature (May-September; C) | 14 * | 16 * | 17 * |
| Total seasonal precipitation (May-September; mm) | 2730 * | 1635 * | 1719 * |
| Seasonal sum of reference evapotranspiration (May-September; mm) | 2036 * | 2325 * | 2166 * |
| Soil type | Haplic and Entic Podzol | Modal Cambisol oligotrophic | Eutric Cambisol |
| References | [28] | [29] | [23] |
| Variants | CZ-BK1 | CZ-RAJ | CZ-Stn | |||
|---|---|---|---|---|---|---|
| SMLR | RF | SMLR | RF | SMLR | RF | |
| Pearson correlation | 0.63 | 0.76 | 0.71 | 0.81 | 0.67 | 0.75 |
| R2 | 0.40 | 0.57 | 0.49 | 0.65 | 0.45 | 0.55 |
| RMSE | 0.18 | 0.15 | 0.14 | 0.12 | 0.15 | 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mensah, C.; Šigut, L.; Fischer, M.; Foltýnová, L.; Jocher, G.; Urban, O.; Wemegah, C.S.; Nyantakyi, E.K.; Chawla, S.; Pavelka, M.; et al. Environmental Effects on Normalized Gross Primary Productivity in Beech and Norway Spruce Forests. Atmosphere 2021, 12, 1128. https://doi.org/10.3390/atmos12091128
Mensah C, Šigut L, Fischer M, Foltýnová L, Jocher G, Urban O, Wemegah CS, Nyantakyi EK, Chawla S, Pavelka M, et al. Environmental Effects on Normalized Gross Primary Productivity in Beech and Norway Spruce Forests. Atmosphere. 2021; 12(9):1128. https://doi.org/10.3390/atmos12091128
Chicago/Turabian StyleMensah, Caleb, Ladislav Šigut, Milan Fischer, Lenka Foltýnová, Georg Jocher, Otmar Urban, Cosmos Senyo Wemegah, Emmanuel K. Nyantakyi, Shilpi Chawla, Marian Pavelka, and et al. 2021. "Environmental Effects on Normalized Gross Primary Productivity in Beech and Norway Spruce Forests" Atmosphere 12, no. 9: 1128. https://doi.org/10.3390/atmos12091128