Cdx2 Animal Models Reveal Developmental Origins of Cancers


Laboratory of Cell Differentiation, Institute of Molecular Genetics of the Czech Academy of Sciences, v.v.i. Vídenská 1083, 4 14220 Prague, Czech Republic
Genes 2019, 10(11), 928; https://doi.org/10.3390/genes10110928
Submission received: 11 October 2019
/
Revised: 6 November 2019
/
Accepted: 13 November 2019
/
Published: 14 November 2019
(This article belongs to the Special Issue Animal Modeling in Cancer)
Abstract
:The Cdx2 homeobox gene is important in assigning positional identity during the finely orchestrated process of embryogenesis. In adults, regenerative responses to tissues damage can require a replay of these same developmental pathways. Errors in reassigning positional identity during regeneration can cause metaplasias—normal tissue arising in an abnormal location—and this in turn, is a well-recognized cancer risk factor. In animal models, a gain of Cdx2 function can elicit a posterior shift in tissue identity, modeling intestinal-type metaplasias of the esophagus (Barrett’s esophagus) and stomach. Conversely, loss of Cdx2 function can elicit an anterior shift in tissue identity, inducing serrated-type lesions expressing gastric markers in the colon. These metaplasias are major risk factors for the later development of esophageal, stomach and colon cancer. Leukemia, another cancer in which Cdx2 is ectopically expressed, may have mechanistic parallels with epithelial cancers in terms of stress-induced reprogramming. This review will address how animal models have refined our understanding of the role of Cdx2 in these common human cancers.
1. Introduction
During the development of the embryo, the specification of cellular and tissue identity is dictated according to location. This is achieved through a combination of inductive cues and cell-intrinsic genetic factors. Our current understanding of the fundamental molecular mechanisms that underly these processes, referred to as pattern formation, was initially spurred by the study of the fruit fly, Drosophila melanogaster, over a century ago [1]. In 1894 William Bateson reported a peculiar mutation in D. melanogaster, in which a leg developed in the place of antennae. This he termed “homeosis”, developmental anomalies which cause one body part to develop in the likeness of another. Genetic mutations which cause homeosis are called homeotic mutations.
Many homeotic mutations have been identified in D. melanogaster. These include the bithorax mutation, where an extra pair of wings are present instead of a pair of halteres, and the aforementioned Antennapaedia mutation, where legs developed in the place of antennae. The gene responsible for the Antennapaedia mutation would be identified almost 90 years later [2] and others soon followed. Comparative sequence analyses indicated that several homeotic genes, including the Antennapaedia gene, contained a conserved 180 nucleotide sequence—the homeobox [3,4,5]. Although many genes important for pattern formation were found to contain a homeobox sequence, homeotic transformations in D. melanogaster were only associated with those genes mapping to a single genetic locus, termed the HOM-C locus [6,7,8].
In human, the HOM-C homologues are termed the HOX clusters. Duplication events during mammalian evolution have produced four separate HOX clusters: HOXA, HOXB, HOXC and HOXD [9]. The expression of genes within both the HOX and HOM-C clusters are spatio-temporally regulated; those located at the 3’-end are expressed earlier and in more anterior regions, while those located at the 5’-end are expressed later and in more posterior regions [10,11,12,13].
Mutations in HOX genes do not cause the dramatic anatomical transformations observed in D. melanogaster, as mammalian development is much less dependent on segmental structures. Only branchial arches, the hindbrain and somites appear to develop on a truly segmental basis, and here the role of HOX genes in controlling development of these structures is well documented [13]. In the mouse, loss-of-function mutations of Hox genes often cause anterior homeotic transformations—an anterior transformation being when a segmental unit acquires the characteristics of one more rostral. Anterior transformations of the axial skeleton have been reported for several Hox null mutants, including Hoxa2 [14,15], Hoxb4 [16], Hoxc8 [17], Hoxd3 [18] and Hoxd13 [19].
An additional paralogous Hox cluster (ParaHox) also exists and, like the Hox cluster, also exhibits spatio-temporal co-linearity [20]. Both gene clusters are evolutionarily ancient, splitting from a common ancestral ProtoHox cluster prior to the split between Bilaterians and Cnidarians, i.e., before the establishment of body plans with bilateral symmetry [21]. In humans, the ParaHox cluster consists of three genes, GSH, PDX1 and CDX2.
As with many of the Hox genes, loss-of-function mutation of Cdx2 in mice is associated with anterior homeotic transformation of the axial skeleton [22]. Similarly, loss-of-function mutation of the related paralogue Cdx1 also causes anterior homeotic shifts [23] and these patterning defects become further exacerbated in Cdx1/Cdx2 compound mutants [24]. Null mutants of the third paralogue, Cdx4, do not exhibit skeletal defects, but exacerbate the axial patterning defects of both Cdx1 and Cdx2 mutants [25]. These findings illustrate not only functional overlap, but show that their collective activity is required to achieve wild-type levels of functional activity—i.e., their functional overlap does not equate to functional redundancy. As such, any genetic or environmental factors that alter Cdx protein levels can have significant effects on establishing positional identity. This is true not only during embryogenesis, but also following regenerative tissue repair in adult tissues, where the reestablishment of positional identity can be required. Incorrect reprogramming of tissue identity in adult tissues is termed metaplasia and metaplasia is increasingly recognized as a major risk factor for developing cancer. This review will focus on the function of Cdx2 and, less so, its paralogues, Cdx1 and Cdx4, and how genetically engineered mutations of these genes have provided us with animal models that have spurred our understanding of the important links between metaplasia and cancer.
2. Metaplasia is an Important Etiological Factor in Cancer
Metaplasia has long been recognized as a risk factor for cancer development and most often follows a common sequence: an environmental insult will cause tissue damage and, in the course of renewal, this tissue may transdifferentiate into a tissue type inappropriate for its location. An early recognized example is the often observed transition from a normal columnar bronchial epithelium to a squamous epithelium in the lungs of smokers, a metaplastic change that is believed to be the site of origin of lung cancers [26]. In 1985, Jonathan Slack proposed that many of these epithelial metaplasias may be analogous to homeotic transformations [27]. He proposed that epithelial stem cells may sometimes be reprogrammed back to an early ontological state and then, as normal progression proceeds, can acquire a new stable epigenetic state that is phenotypically anteriorized or posteriorized. This hypothesis was bolstered by findings showing an anterior shift in epithelial identity in the focal regions of the large intestine of Cdx2 mutant mice, occurring concomitant with anterior shifts in the axial skeleton [22,28,29]. Later studies would show that targeted overexpression of Cdx2 could induce metaplasias in the gut, in which anterior epithelial structures were replaced with posterior structures, i.e., directly analogous to a posterior homeotic shift [30,31,32,33,34] (Table 1, Figure 1). Thus, Cdx2 insufficiency was associated with shifts in the opposite direction to conditions where Cdx2 was overexpressed. Nevertheless, in both cases, the shifts were associated with cancer progression pathways. As will be discussed in the following sections, these animal models have been valuable in furthering our understanding of cancers of the esophagus, stomach and colon, as well as its less understood oncogenic role in leukemia.
Environmental insults can induce reprogramming of the gut epithelium. These metaplasias and metaplasia-like (i.e., serrated polyps in the colon) alterations can confer risk for subsequent development of cancers of the gastrointestinal tract.
3. Esophageal Cancer
3.1. Barrett’s Esophagus is a Major Risk Factor for Human Esophageal Adenocarcinoma
Barrett’s esophagus is a metaplastic change of normal squamous esophageal epithelium to an abnormal columnar epithelia with gastric and intestinal features [36,37]. The metaplasia arises as a consequence of the epithelial damage and inflammatory response wrought by chronic acid reflux. Patients diagnosed with Barrett’s esophagus have an approximate 100-fold increased risk of developing esophageal adenocarcinoma [38] and, because this metaplastic transformation appears irreversible, preventative measures have relied on controlling acid reflux, primarily through the use of proton pump inhibitors (PPIs) [39]. A recent clinical trial has shown that patients with existing Barrett’s esophagus can still reduce their risk of developing adenocarcinoma by taking PPIs [40].
Not being present in the normal esophagus, CDX2 expression is a biomarker for Barrett’s esophagus [41,42,43,44]. Moreover, CDX2 expression can often be found in esophageal squamous epithelia inflamed by acid reflux, suggesting that its expression precedes the metaplastic transformation [41,45]. CDX2 is a direct transcriptional target of the key inflammatory mediator NF-κB [46]. Thus, it is likely that the onset of CDX2 expression is due to activated NF-κB, which has also been shown to be present in pre-metaplastic inflamed squamous epithelia [47,48]. CDX2 expression is maintained if the metaplasia advances to an adenocarcinoma, but expression diminishes as the cancer loses epithelial morphology [49,50].
3.2. Animal Models Reveal Functional Roles for Cdx2 in Barrett’s Esophagus
A keratin 14 promoter was used to force Cdx2 overexpression to the squamous epithelia of mouse esophagus [31]. This was sufficient to induce metaplastic changes in the esophagus that resembled Barrett’s esophagus, but lacked the intestinal goblet cells that are characteristic of the disease in humans. The transition from squamous to columnar epithelia was also associated with a decrease in barrier function, leading to the hypothesis that the transformed epithelia could, in turn, be more sensitive to reflux esophagitis, reinforcing the transition [31]. A similar model was generated in zebrafish, using a keratin 5 promoter, to drive expression of cdx1b [30]. Like the transgenic mouse model, the zebrafish model exhibited similar metaplastic changes in the esophagus but, once again, without any appearance of goblet cells. More recently, a discrete transitional columnar epithelium was found to exist at the junction of the stomach and esophagus [34] and may represent the true source of Barrett’s esophagus. Keratin 7 was identified as a specific marker of this transitional epithelium and, by employing a keratin 7 promoter to confine inducible Cdx2 overexpression to this cell-type, a metaplasia that included goblet cells was observed [34]. Currently, this compound transgenic model (Krt7rtTA; otet-CDX2-T2A-mCherry) represents the best animal model for replicating Barrett’s esophagus.
4. Stomach Cancer
4.1. CDX2 and the Metaplastic Origins of Human Stomach Cancer
The current model for human gastric carcinogenesis, proposed in 1992 [51], follows a similar course to the model for esophageal carcinogenesis discussed earlier, in that an initial pro-inflammatory stimulus will lead to inflammation (gastritis), followed by intestinal metaplasia, and, ultimately, to adenocarcinoma. In the stomach, the major environmental stimulus initiating this pathway, and thereby conferring the risk of cancer development, is chronic infection with Helicobacter pylori [52].
Two major types of metaplastic lineages have been identified adjacent to cancers of the stomach: an intestinal-type metaplasia with the characteristic presence of goblet cells [53] and a spasmolytic polypeptide-expressing metaplasia (SPEM) which expresses trefoil factor 2 (TFF2), then designated as spasmolytic polypeptide [54]. SPEM exhibits similarities to glands of the antrum (the caudal-most region of the stomach) [55,56]. SPEM may represent a reparative response to acute gastritis, while goblet-cell intestinal metaplasia may require a chronic inflammatory environment.
CDX2 expression is detected in gastric intestinal metaplasia but not in normal gastric mucosa [57,58,59]. CDX2 could also be detected in chronic gastritis without evidence of metaplasia, suggesting that the onset of CDX2 expression preceded the metaplastic change [60]. As the metaplasias progresses to carcinoma, CDX2 levels are often reduced [58].
4.2. Animal Models Reveal Functional Roles for Cdx2 in Stomach Cancer
Cdx2 overexpression in the gastric mucosa of transgenic mice using the parietal cell-specific H+/K+-ATPase subunit b promoter resulted in gastric intestinal-type metaplasia that spontaneously developed into adenocarcinomas [61,62]. Another line, employing the Foxa3 promoter to drive Cdx2 overexpression, also exhibited intestinal-type metaplasia but progression to adenocarcinoma was not observed [33]. The significant overlap in phenotype is somewhat surprising, as Foxa3 is expressed during embryonic development while the H+/K+-ATPase promoter is only active postnatally in the acid-producing parietal cells. It is possible that the phenotype may be indirectly influenced by parietal cell loss, as gastrin knockout mice, which exhibit impairment of stomach acid production, also exhibit intestinal-type metaplasia that eventually progresses to stomach cancer [63,64].
Modelling the action of the major risk factor for stomach cancer, H. pylori infection, has proven more difficult in mice, as this is heavily mouse strain-dependent [65] and produce SPEM rather than intestinal-type metaplasia [66]. For replicating the human disease, the Mongolian gerbil has been superior, recapitulating upon H. pylori infection the progression from gastritis to intestinal metaplasia and, eventually, to gastric cancer [67,68,69].
5. Colon Cancer
5.1. CDX2 is a Tumour Suppressor in Human Colorectal Cancer
Most colorectal cancers arise from an epithelial-derived adenomatous precursor lesion that, with further mutations in oncogenes and tumor suppressor genes, can clonally progress to carcinoma [70]. This adenoma–carcinoma pathway is most often initiated by activating mutations of the WNT pathway [71]. An alternative pathway, broadly termed the ‘serrated pathway’, maintains epithelial gland morphology and mucin production in the benign precursor lesions. It has been estimated that 20%–30% of colorectal cancers arise by this alternative pathway, although classification can be difficult as the cancer progresses and loses its serrated morphology [72,73]. This pathway is most often associated with activating mutations in the BRAF oncogene [72,74], and is considered to follow a more aggressive course than the conventional pathway [75]. Loss of CDX2 expression has recently emerged as a biomarker for colon cancers arising via the serrated pathway, often coinciding with activated BRAF mutations [76,77]. It also has been identified as a prognostic marker in stage II colon cancer, where it was suggested that patients with CDX-negative cancers would benefit from adjuvant chemotherapy, rather than the common practice of treating all stage II patients with surgery alone [78].
Are serrated pathway cancers derived from metaplastic changes in the colonic epithelium? Suggestive of an anteriorization of epithelial identity is the fact that these cancers often express gastric epithelial markers, including mucin 2 (MUC2), MUC5AC, MUC6 and annexin A10 (ANXA10) [79,80,81]. Perhaps more compelling is that loss of CDX2 expression is associated with a gain in PDX1 expression, the ParaHox gene responsible for patterning the midgut [77].
5.2. Animal Models Reveal Functional Roles for Cdx2 in Colorectal Cancer
A possible role for Cdx2 in colon cancer was initially suggested based on the knockout phenotype in mice; heterozygous mice had numerous tumors, although they never spontaneously advanced to carcinoma [22,29]. Upon closer examination, these tumors consisted of small foci of histologically normal forestomach epithelia that were surrounded in successive order by cardia, corpus, antrum and small intestine epithelia [82]. This observation was ascribed as a heterotopia, analogous to a metaplastic transformation, only with its origins occurring during embryological development instead of as a consequence of mucosal injury and repair. To model the latter, a Cre-ERT transgene under the Cyp1a1 promoter, was used to achieve mosaic inactivation of a Cdx2fl/fl allele, thus allowing the study of Cdx2 deficient lesions in the context of wild-type mucosa [83]. The Cdx2 deficient lesions were found to express a number of gastric genes but did not form normal gastric mucosa, presumably because of incompatible mesenchymal signaling [83]. Under current classifications, these lesions could be interpreted as “serrated”. Could they therefore be susceptible to transformation via activating mutations of BRAF?
Mouse models combining Cdx2 inactivation and oncogenic BRAF (BRAFV600E) activation were recently described and indeed, this led to invasive carcinogenesis along the serrated pathway [77,84]. Tamoxifen-regulated Cre protein (CreERT2) was used to inducibly inactivate loxP-containing alleles of Cdx2 (Cdx2fl/fl) or to inducibly activate an oncogenic BRAF allele (BRAFV600E) in the adult intestinal epithelium. Mutation of either allele individually had little to no effect on median survival; however, their combined mutation resulted in progression to carcinoma. Immunohistochemical analyses of tumors revealed ectopic expression of typical serrated pathway markers such as annexin A10 and mucin 5AC [77].
Mouse models have also provided information that loss of Cdx2 expression can influence not only the serrated pathway, but also the classical adenoma–carcinoma pathway. The classical pathway is associated with activating mutations of the Wnt signaling pathway, most predominantly through loss of function of the Wnt-signaling inhibitor, Apc [71]. Mutations in the human APC gene are causative for the cancer syndrome Familial Adenomatous Polyposis (FAP), as well as for sporadic cancers arising predominantly in the distal colon [85]. FAP can be modeled in mice carrying mutations of the Apc gene, including the truncated mutant Apc∆716 [86], but tumors in mice arise predominantly in the small intestine. When the Apc∆716 mutant allele is combined with the Cdx2+/− heterozygous mutation, there is a large increase in the number of adenomatous polyps in the distal colon, more closely reflecting the tumor distribution in human FAP [87].
More recently, it was reported that the tumor-promoting effect of Cdx2 deficiency on the classical adenoma–carcinoma pathway may be non-cell autonomous [35]. This discovery was enabled by a complex mouse model, where mosaic inactivation of a Cdx2fl/fl allele was combined with a mutant Apc+/Δ14 allele to drive adenoma formation and a conditionally activated fluorescent reporter allele (tdTomato) to trace cells that underwent Cre-mediated recombination. As expected, adenomas contained high levels of nuclear β-catenin, a measure of hyperactive Wnt signaling arising due to the loss of heterozygosity of the Apc tumor suppressor allele. However, these cells were never red (Cdx2 negative). The Cdx2-negative cells were not contributing to the adenoma, but instead created an environment that promoted neoplasia of Cdx2-positive cells—i.e., Cdx2 was acting as a “non-cell-autonomous tumor suppressor” [35].
Previous studies had shown that Cdx2+/− mice are more susceptible to DSS-induced colitis [88]. Perhaps the permissive environment is pro-inflammatory. Indeed, NF-κB, a key mediator for inflammatory responses, was activated only in the Cdx2-positive cells that were adjacent to the regions of incomplete metaplasia [35]. These activated cells also expressed high levels of nitric oxide synthase (iNOS), indicating that these cells were under increased nitrosative and oxidative stress and therefore more susceptible to DNA damage. Supporting this hypothesis, treatment with the iNOS inhibitor aminoguanidine reduced the tumor load in mice carrying a mutant Cdx2 allele (Apc+/Δ14; Cdx2+/−), while having no effect on mice with only wild-type Cdx2 alleles (Apc+/Δ14; Cdx2+/+) [35].
6. Leukemia
6.1. CDX2 is a Proto-Oncogene in Human AML
A possible role for CDX2 in human acute myeloid leukemia (AML) was first suggested following the identification of a novel chromosomal rearrangement, t (12; 13)(p13;q12), in a patient with AML. The rearrangement yielded an ets variant gene 6–CDX2 (ETV6–CDX2) fusion protein [89] and, as ETV6 is an important regulator of HSC survival and is frequently affected by translocations [90,91], it was thought that this fusion may be oncogenic. However, when the fusion protein was transduced into murine hematopoietic progenitors, it caused only minor myeloproliferation, and not transplantable AML [92]. It is now accepted that it was the full-length CDX2 protein, driven from an alternative ETV6 promoter, that was leukemogenic. Indeed, transduction of full-length CDX2 into murine hematopoietic progenitors does result in transplantable lethal AML [92].
Up to 89% of AML cases, and up to 81% of acute lymphoblastic leukemia (ALL) cases, express CDX2 [93,94,95,96] and at least for ALL, CDX2 expression levels were directly associated with the aggressiveness of the disease [93,95]. Thus CDX2 is one of the most frequently expressed proto-oncogenes in human leukemia.
Known downstream targets of CDX2, namely the HOX genes, had also been identified as proto-oncogenes in AML [97,98]. Forced expression of Hoxa9 or Hoxa10 are also capable of inducing rapid AML in mice [99,100], and aberrant expression of human HOX genes, including HOXA9, correlates with clinical measures of disease burden [101,102,103,104].
During hematopoiesis, HOX genes of the A and B cluster are highly expressed in normal murine and human hematopoietic stem and committed progenitor cells, and become silenced during the course of normal differentiation [105,106]. Bone marrow from Hoxa9 deficient mice has a profound deficiency in the number of hematopoietic stem cells and progenitors [107,108]. On the other hand, CDX2 is not detected in hematopoietic stem or progenitor cells from healthy subjects, neither in human nor in mouse [109]. Also, there were no significant effects on hematopoiesis in knockout mouse models of any of the CDX genes [22,25,109,110]. Thus, although CDX2 and the HOX genes have similar roles in leukemogenesis, the similarities are not readily apparent in regards to the process of normal hematopoiesis. A true functional role would only be revealed as a result of important scientific discoveries in zebrafish.
6.2. Cdx Genes are Required for Normal Hematopoiesis in Zebrafish
The first indication that Cdx genes may have a functional role in hematopoiesis came from studies in zebrafish, when the causative mutation underlying the autosomal recessive mutation kugelig (kgg) was identified in the cdx4 gene [111]. Homozygous kgg embryos die early in development (day 5 to 10 post fertilization) with severe tail defects and a prominent reduction in hemoglobin-staining erythroid cells. This phenotype was consistent with the expression pattern of cdx4, which became restricted to the posterior end of the embryo during early somitogenesis, prior to the emergence of the posterior blood islands. Furthermore, the in vivo injection of cdx4 mRNA was able to rescue hematopoiesis in these kgg mutants [111].
Zebrafish contain a duplication of the Cdx1 gene (cdx1a and cdx1b), while lacking a Cdx2 orthologue. Therefore, although they contain the same number of Cdx genes as in mammals, they lack the prototypical ParaHox cluster. Nevertheless, like in mammals, the zebrafish cohort of Cdx genes does exhibit some degree of functional redundancy. Indeed, morpholino-mediated knockdown of cdx1a in kgg mutant fish exacerbates the phenotype, causing a complete failure to specify blood [112].
The hematopoiesis defect in cdx4 mutant zebrafish is reminiscent of anterior homeotic transformation of the axial skeleton observed in mouse loss of function mutants [22,110], as there appeared to be a posterior shift in the boundary between anteriorly localized hemangioblasts, fated to develop into endothelial cells and the posteriorly localized hemangioblasts, fated to develop both blood and endothelial cells [111]. Both populations are labelled with scl (tal1), which coexpresses with cdx4 in the posterior blood islands [111]. Even though scl overexpression is able to expand hematopoietic cell numbers when overexpressed in wild-type zebrafish embryos [113], it was incapable of rescuing hematopoiesis in cdx4 mutant kgg embryos [111]. Thus, the hematopoietic defect did not seem to be due to an overt lack in the number of scl+ hemangioblast progenitors, but rather a failure to pattern these progenitors to favor differentiation towards the erythrocyte lineage.
An evolutionarily conserved role of Cdx genes in regulating the expression of Hox genes [110,114,115] appears to underly the failure to pattern scl+ hemangioblasts in zebrafish. Indeed, kgg mutants exhibit large alterations in hox expression patterns, which can be restored upon ectopic cdx4 expression [111]. Overexpression of several of the most downregulated hox genes (hoxb6b, hoxb7a and hoxa9a) successfully rescues hematopoiesis in kgg mutants [111], and overexpression of hoxa9a (but not hoxb7a) rescues the complete hematopoietic failure observed upon combined cdx1a and cdx4 deficiency [112].
6.3. Cdx Genes Control Mammalian Hematopoiesis
The implications from these studies in zebrafish were that a functional role for mammalian CDX genes may be masked by functional redundancies and that the CDX genes may exert their function not at the level of hematopoetic stem cells, but by pre-patterning their early mesodermal progenitors during embryogenesis. This possibility could be simply assessed by the in vitro differentiation of embryonic stem cell lines, since differentiation towards hematopoietic lineages involves a transition through a hemangioblast intermediate.
In vitro differentiation of single Cdx gene deficient murine embryonic stem cell lines resulted in only minor reductions in the numbers of multipotential blood progenitor colonies [116]. Knockdown of either Cdx1 or Cdx2 by RNA interference in a Cdx4-deficient background resulted in more severe reductions, while combined knockdown of both Cdx1 and Cdx2 in the Cdx4-deficient background resulted in an almost complete failure of hematopoiesis [116]. In embryos where Cdx2 was conditionally inactivated in a Cdx1−/− background, there were defects in primitive hematopoiesis as well as yolk sac vascularization [117]. Thus a previously unrecognized role for Cdx genes in hematopoiesis was made evident when all genes in the family were inactivated.
The role of Cdx genes in pre-patterning early presomitic mesodermal progenitors, which will later give rise to hematopoietic lineages, can first of all be inferred by their ability to pattern the somitic mesoderm, resulting in the prototypical anterior homeotic transformation of the vertebrae [22,110]. Also, upon in vitro differentiation of human and mouse embryonic stem cells, Cdx gene expression peaks at the same time as hemangioblasts are specified and, if inducibly overexpressed during this time window, strongly enhances the production of hematopoietic progenitors [118,119,120]. The effect on hemangioblast production is likely the result of both a decreased amount of posterior unsegmented mesoderm [121] and an anterior shift in patterning the mesoderm [122]. In zebrafish, the tbx5a-expressing anterior cardiogenic mesoderm was expanded in cdx1a/4 mutants [122]. Similarly, in mice, ectopic Tbx5 expression was observed in the yolk sac of Cdx1/2 compound conditional null mutants at the expense of hematopoietic markers [123]. Current evidence supports a mechanism of action for Cdx genes in which they stably repress cardiac loci in early Mesp1+ mesoderm by directly recruiting the SWI/SNF epigenetic silencing complex [123]. Thus, the expression of Cdx biases these progenitors to hematopoietic lineages at the expense of cardiac lineages.
7. Summary
Metaplasias, long recognized as a cancer risk factor, have been suggested to be analogous to developmental homeosis, where normal tissues develop in an abnormal location [27]. Homeobox genes, including Cdx2, are important factors in conferring positional identity to developing tissues, whether during embryogenesis or during the regenerative process following tissue injury. Animal models in which Cdx2 overexpression is targeted to the esophagus show Barrett’s metaplasia (Barrett’s esophagus), characterized by the presence of intestinal-type epithelia in place of normal squamous epithelia. [30,31,34]. Similarly, targeted overexpression of Cdx2 in the stomach also causes metaplasia, with a posteriorization of epithelial identity [32,33]. These tissue alterations model pre-neoplastic metaplasias that are common in humans. Conversely, loss of Cdx2 in the colon causes metaplasia-like alterations, in which epithelia are misallocated towards an identity characteristic of more anterior structures [22,28,29], and this has provided important insights into understanding the progression of human serrated-type colonic tumors.
It is easy to recognize epithelial metaplasias, as any change in the reacquiring of positional identity in an epithelial stem cell will be conferred as a change in phenotype in its regionally constrained cell progeny. However, this is not the case for another cancer in which Cdx2 is ectopically expressed—leukemia. Nevertheless, it is possible that the same sequence of events is occurring in leukemia—chronic inflammatory damage triggering a regenerative response, which results in the acquisition of a more “posteriorized” epigenetic state.
While the importance of CDX2 in human cancer pathology is indisputable, its functional role has been more difficult to define. It has been designated, somewhat contradictorily, as both as an oncogene and a tumor suppressor. But, unlike prototypical oncogenes and tumor suppressor genes, there is no strong statistical evidence for cancer-associated mutations or loss of heterozygosity. The issue is that these terms describe cell-intrinsic functions, while the core function of CDX2, as a designator of positional identity, is, by definition, relativistic. Therefore, a true understanding of its role in cancer progression must be context-dependent. Indeed, the conceptual advances in this field, as discussed in this review, have been driven almost exclusively by the judicious use of animal models.
Funding
This work was funded by the Ministry of Education, Youth and Sports of the Czech Republic under the National Sustainability Program I project LO1419 and GACR 17-16959S.
Acknowledgments
I thank Felix Beck and Trevor Epp for helpful comments on this review manuscript and for proofreading.
Conflicts of Interest
The author declares no conflict of interest.
References
- Bateson, W. Materials for the Study of Variation: Treated with Especial Regard to Discontinuity in the Origin of Species; Macmillan: London, UK; New York, NY, USA, 1894; p. 598. [Google Scholar]
- Garber, R.L.; Kuroiwa, A.; Gehring, W.J. Genomic and cDNA clones of the homeotic locus Antennapedia in Drosophila. EMBO J. 1983, 2, 2027–2036. [Google Scholar] [CrossRef]
- McGinnis, W.; Garber, R.L.; Wirz, J.; Kuroiwa, A.; Gehring, W.J. A homologous protein-coding sequence in Drosophila homeotic genes and its conservation in other metazoans. Cell 1984, 37, 403–408. [Google Scholar] [CrossRef]
- McGinnis, W.; Hart, C.P.; Gehring, W.J.; Ruddle, F.H. Molecular cloning and chromosome mapping of a mouse DNA sequence homologous to homeotic genes of Drosophila. Cell 1984, 38, 675–680. [Google Scholar] [CrossRef]
- Scott, M.P.; Weiner, A.J. Structural relationships among genes that control development: Sequence homology between the Antennapedia, Ultrabithorax, and fushi tarazu loci of Drosophila. Proc. Natl. Acad. Sci. USA 1984, 81, 4115–4119. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Nusslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef]
- Wakimoto, B.T.; Kaufman, T.C. Analysis of larval segmentation in lethal genotypes associated with the antennapedia gene complex in Drosophila melanogaster. Dev. Biol. 1981, 81, 51–64. [Google Scholar] [CrossRef]
- Kappen, C.; Ruddle, F.H. Evolution of a regulatory gene family: HOM/HOX genes. Curr. Opin. Genet. Dev. 1993, 3, 931–938. [Google Scholar] [CrossRef]
- Duboule, D.; Morata, G. Colinearity and functional hierarchy among genes of the homeotic complexes. Trends Genet. 1994, 10, 358–364. [Google Scholar] [CrossRef]
- Graham, A.; Papalopulu, N.; Krumlauf, R. The murine and Drosophila homeobox gene complexes have common features of organization and expression. Cell 1989, 57, 367–378. [Google Scholar] [CrossRef]
- Izpisua-Belmonte, J.C.; Falkenstein, H.; Dolle, P.; Renucci, A.; Duboule, D. Murine genes related to the Drosophila AbdB homeotic genes are sequentially expressed during development of the posterior part of the body. EMBO J. 1991, 10, 2279–2289. [Google Scholar] [CrossRef]
- Krumlauf, R. Hox genes in vertebrate development. Cell 1994, 78, 191–201. [Google Scholar] [CrossRef]
- Gendron-Maguire, M.; Mallo, M.; Zhang, M.; Gridley, T. Hoxa-2 mutant mice exhibit homeotic transformation of skeletal elements derived from cranial neural crest. Cell 1993, 75, 1317–1331. [Google Scholar] [CrossRef]
- Rijli, F.M.; Mark, M.; Lakkaraju, S.; Dierich, A.; Dolle, P.; Chambon, P. A homeotic transformation is generated in the rostral branchial region of the head by disruption of Hoxa-2, which acts as a selector gene. Cell 1993, 75, 1333–1349. [Google Scholar] [CrossRef]
- Ramirez-Solis, R.; Zheng, H.; Whiting, J.; Krumlauf, R.; Bradley, A. Hoxb-4 (Hox-2.6) mutant mice show homeotic transformation of a cervical vertebra and defects in the closure of the sternal rudiments. Cell 1993, 73, 279–294. [Google Scholar] [CrossRef]
- Le Mouellic, H.; Lallemand, Y.; Brulet, P. Homeosis in the mouse induced by a null mutation in the Hox-3.1 gene. Cell 1992, 69, 251–264. [Google Scholar] [CrossRef]
- Condie, B.G.; Capecchi, M.R. Mice homozygous for a targeted disruption of Hoxd-3 (Hox-4.1) exhibit anterior transformations of the first and second cervical vertebrae, the atlas and the axis. Development 1993, 119, 579–595. [Google Scholar]
- Dolle, P.; Dierich, A.; LeMeur, M.; Schimmang, T.; Schuhbaur, B.; Chambon, P.; Duboule, D. Disruption of the Hoxd-13 gene induces localized heterochrony leading to mice with neotenic limbs. Cell 1993, 75, 431–441. [Google Scholar] [CrossRef]
- Brooke, N.M.; Garcia-Fernandez, J.; Holland, P.W. The ParaHox gene cluster is an evolutionary sister of the Hox gene cluster. Nature 1998, 392, 920–922. [Google Scholar] [CrossRef]
- Chourrout, D.; Delsuc, F.; Chourrout, P.; Edvardsen, R.B.; Rentzsch, F.; Renfer, E.; Jensen, M.F.; Zhu, B.; de Jong, P.; Steele, R.E.; et al. Minimal ProtoHox cluster inferred from bilaterian and cnidarian Hox complements. Nature 2006, 442, 684–687. [Google Scholar] [CrossRef]
- Chawengsaksophak, K.; James, R.; Hammond, V.E.; Kontgen, F.; Beck, F. Homeosis and intestinal tumours in Cdx2 mutant mice. Nature 1997, 386, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, V.; Meyer, B.; Evans, G.S. The murine Cdx1 gene product localises to the proliferative compartment in the developing and regenerating intestinal epithelium. Differentiation 1998, 64, 11–18. [Google Scholar] [CrossRef]
- Van den Akker, E.; Forlani, S.; Chawengsaksophak, K.; de Graaff, W.; Beck, F.; Meyer, B.I.; Deschamps, J. Cdx1 and Cdx2 have overlapping functions in anteroposterior patterning and posterior axis elongation. Development 2002, 129, 2181–2193. [Google Scholar] [PubMed]
- Van Nes, J.; de Graaff, W.; Lebrin, F.; Gerhard, M.; Beck, F.; Deschamps, J. The Cdx4 mutation affects axial development and reveals an essential role of Cdx genes in the ontogenesis of the placental labyrinth in mice. Development 2006, 133, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, O.; Stout, A.P.; Hammond, E.C.; Garfinkel, L. Changes in bronchial epithelium in relation to cigarette smoking and in relation to lung cancer. N. Engl. J. Med. 1961, 265, 253–267. [Google Scholar] [CrossRef]
- Slack, J.M. Homoeotic transformations in man: Implications for the mechanism of embryonic development and for the organization of epithelia. J. Theor. Biol. 1985, 114, 463–490. [Google Scholar] [CrossRef]
- Beck, F.; Chawengsaksophak, K.; Waring, P.; Playford, R.J.; Furness, J.B. Reprogramming of intestinal differentiation and intercalary regeneration in Cdx2 mutant mice. Proc. Natl. Acad. Sci. USA 1999, 96, 7318–7323. [Google Scholar] [CrossRef]
- Tamai, Y.; Nakajima, R.; Ishikawa, T.; Takaku, K.; Seldin, M.F.; Taketo, M.M. Colonic hamartoma development by anomalous duplication in Cdx2 knockout mice. Cancer Res. 1999, 59, 2965–2970. [Google Scholar]
- Hu, B.; Chen, H.; Liu, X.; Zhang, C.; Cole, G.J.; Lee, J.A.; Chen, X. Transgenic overexpression of cdx1b induces metaplastic changes of gene expression in zebrafish esophageal squamous epithelium. Zebrafish 2013, 10, 218–227. [Google Scholar] [CrossRef]
- Kong, J.; Crissey, M.A.; Funakoshi, S.; Kreindler, J.L.; Lynch, J.P. Ectopic Cdx2 expression in murine esophagus models an intermediate stage in the emergence of Barrett’s esophagus. PLoS ONE 2011, 6, e18280. [Google Scholar] [CrossRef]
- Mutoh, H.; Satoh, K.; Kita, H.; Sakamoto, H.; Hayakawa, H.; Yamamoto, H.; Isoda, N.; Tamada, K.; Ido, K.; Sugano, K. Cdx2 specifies the differentiation of morphological as well as functional absorptive enterocytes of the small intestine. Int J. Dev. Biol. 2005, 49, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Silberg, D.G.; Sullivan, J.; Kang, E.; Swain, G.P.; Moffett, J.; Sund, N.J.; Sackett, S.D.; Kaestner, K.H. Cdx2 ectopic expression induces gastric intestinal metaplasia in transgenic mice. Gastroenterology 2002, 122, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Li, H.; Zhang, Y.; Yang, Y.; Lu, R.; Liu, K.; Lin, S.; Lan, X.; Wang, H.; Wu, H.; et al. Transitional basal cells at the squamous-columnar junction generate Barrett’s oesophagus. Nature 2017, 550, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Balbinot, C.; Armant, O.; Elarouci, N.; Marisa, L.; Martin, E.; De Clara, E.; Onea, A.; Deschamps, J.; Beck, F.; Freund, J.N.; et al. The Cdx2 homeobox gene suppresses intestinal tumorigenesis through non-cell-autonomous mechanisms. J. Exp. Med. 2018, 215, 911–926. [Google Scholar] [CrossRef]
- McDonald, S.A.; Lavery, D.; Wright, N.A.; Jansen, M. Barrett oesophagus: Lessons on its origins from the lesion itself. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 50–60. [Google Scholar] [CrossRef]
- Spechler, S.J.; Souza, R.F. Barrett’s esophagus. N. Engl. J. Med. 2014, 371, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.P.; Hardie, L.J. Reflux, Barrett’s oesophagus and adenocarcinoma: Burning questions. Nat. Rev. Cancer 2003, 3, 676–684. [Google Scholar] [CrossRef]
- El-Serag, H.B.; Aguirre, T.V.; Davis, S.; Kuebeler, M.; Bhattacharyya, A.; Sampliner, R.E. Proton pump inhibitors are associated with reduced incidence of dysplasia in Barrett’s esophagus. Am. J. Gastroenterol. 2004, 99, 1877–1883. [Google Scholar] [CrossRef]
- Jankowski, J.A.Z.; de Caestecker, J.; Love, S.B.; Reilly, G.; Watson, P.; Sanders, S.; Ang, Y.; Morris, D.; Bhandari, P.; Brooks, C.; et al. Esomeprazole and aspirin in Barrett’s oesophagus (AspECT): A randomised factorial trial. Lancet 2018, 392, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Eda, A.; Osawa, H.; Satoh, K.; Yanaka, I.; Kihira, K.; Ishino, Y.; Mutoh, H.; Sugano, K. Aberrant expression of CDX2 in Barrett’s epithelium and inflammatory esophageal mucosa. J. Gastroenterol. 2003, 38, 14–22. [Google Scholar] [CrossRef]
- Groisman, G.M.; Amar, M.; Meir, A. Expression of the intestinal marker Cdx2 in the columnar-lined esophagus with and without intestinal (Barrett’s) metaplasia. Mod. Pathol. 2004, 17, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.W.; Frierson, H.F., Jr.; Moskaluk, C.A. Cdx2 as a marker of epithelial intestinal differentiation in the esophagus. Am. J. Surg. Pathol. 2003, 27, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Vallbohmer, D.; DeMeester, S.R.; Peters, J.H.; Oh, D.S.; Kuramochi, H.; Shimizu, D.; Hagen, J.A.; Danenberg, K.D.; Danenberg, P.V.; DeMeester, T.R.; et al. Cdx-2 expression in squamous and metaplastic columnar epithelia of the esophagus. Dis Esophagus 2006, 19, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Moons, L.M.; Bax, D.A.; Kuipers, E.J.; Van Dekken, H.; Haringsma, J.; Van Vliet, A.H.; Siersema, P.D.; Kusters, J.G. The homeodomain protein CDX2 is an early marker of Barrett’s oesophagus. J. Clin. Pathol 2004, 57, 1063–1068. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Domon-Dell, C.; Wang, Q.; Chung, D.H.; Di Cristofano, A.; Pandolfi, P.P.; Freund, J.N.; Evers, B.M. PTEN and TNF-alpha regulation of the intestinal-specific Cdx-2 homeobox gene through a PI3K, PKB/Akt, and NF-kappaB-dependent pathway. Gastroenterology 2002, 123, 1163–1178. [Google Scholar] [CrossRef]
- Huo, X.; Agoston, A.T.; Dunbar, K.B.; Cipher, D.J.; Zhang, X.; Yu, C.; Cheng, E.; Zhang, Q.; Pham, T.H.; Tambar, U.K.; et al. Hypoxia-inducible factor-2alpha plays a role in mediating oesophagitis in GORD. Gut 2017, 66, 1542–1554. [Google Scholar] [CrossRef] [Green Version]
- O’Riordan, J.M.; Abdel-latif, M.M.; Ravi, N.; McNamara, D.; Byrne, P.J.; McDonald, G.S.; Keeling, P.W.; Kelleher, D.; Reynolds, J.V. Proinflammatory cytokine and nuclear factor kappa-B expression along the inflammation-metaplasia-dysplasia-adenocarcinoma sequence in the esophagus. Am. J. Gastroenterol. 2005, 100, 1257–1264. [Google Scholar] [CrossRef]
- Moskaluk, C.A.; Zhang, H.; Powell, S.M.; Cerilli, L.A.; Hampton, G.M.; Frierson, H.F., Jr. Cdx2 protein expression in normal and malignant human tissues: An immunohistochemical survey using tissue microarrays. Mod. Pathol. 2003, 16, 913–919. [Google Scholar] [CrossRef]
- Werling, R.W.; Yaziji, H.; Bacchi, C.E.; Gown, A.M. CDX2, a highly sensitive and specific marker of adenocarcinomas of intestinal origin: An immunohistochemical survey of 476 primary and metastatic carcinomas. Am. J. Surg. Pathol. 2003, 27, 303–310. [Google Scholar] [CrossRef]
- Correa, P. Human gastric carcinogenesis: A multistep and multifactorial process--First American Cancer Society Award Lecture on Cancer Epidemiology and Prevention. Cancer Res. 1992, 52, 6735–6740. [Google Scholar]
- Correa, P.; Houghton, J. Carcinogenesis of Helicobacter pylori. Gastroenterology 2007, 133, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Okamoto, S.; Yamamoto, S.; Matsumura, N.; Yamaguchi, S.; Yamakido, M.; Taniyama, K.; Sasaki, N.; Schlemper, R.J. Helicobacter pylori infection and the development of gastric cancer. N. Engl. J. Med. 2001, 345, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.H.; Lee, J.R.; Joshi, V.; Playford, R.J.; Poulsom, R.; Wright, N.A.; Goldenring, J.R. Identification of a metaplastic cell lineage associated with human gastric adenocarcinoma. Lab. Investig. 1999, 79, 639–646. [Google Scholar]
- Nomura, S.; Baxter, T.; Yamaguchi, H.; Leys, C.; Vartapetian, A.B.; Fox, J.G.; Lee, J.R.; Wang, T.C.; Goldenring, J.R. Spasmolytic polypeptide expressing metaplasia to preneoplasia in H. felis-infected mice. Gastroenterology 2004, 127, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Yamaguchi, H.; Ogawa, M.; Wang, T.C.; Lee, J.R.; Goldenring, J.R. Alterations in gastric mucosal lineages induced by acute oxyntic atrophy in wild-type and gastrin-deficient mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G362–G375. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.; Silva, E.; Santos-Silva, F.; Silberg, D.G.; Wang, J.; De Bolos, C.; David, L. Expression of intestine-specific transcription factors, CDX1 and CDX2, in intestinal metaplasia and gastric carcinomas. J. Pathol. 2003, 199, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.Q.; Yamamoto, H.; Akiyama, Y.; Tanaka, H.; Takizawa, T.; Koike, M.; Kenji Yagi, O.; Saitoh, K.; Takeshita, K.; Iwai, T.; et al. Ectopic expression of homeodomain protein CDX2 in intestinal metaplasia and carcinomas of the stomach. Cancer Lett. 2002, 176, 47–55. [Google Scholar] [CrossRef]
- Mizoshita, T.; Inada, K.; Tsukamoto, T.; Kodera, Y.; Yamamura, Y.; Hirai, T.; Kato, T.; Joh, T.; Itoh, M.; Tatematsu, M. Expression of Cdx1 and Cdx2 mRNAs and relevance of this expression to differentiation in human gastrointestinal mucosa—with special emphasis on participation in intestinal metaplasia of the human stomach. Gastric Cancer 2001, 4, 185–191. [Google Scholar] [CrossRef]
- Eda, A.; Osawa, H.; Yanaka, I.; Satoh, K.; Mutoh, H.; Kihira, K.; Sugano, K. Expression of homeobox gene CDX2 precedes that of CDX1 during the progression of intestinal metaplasia. J. Gastroenterol. 2002, 37, 94–100. [Google Scholar] [CrossRef]
- Mutoh, H.; Hakamata, Y.; Sato, K.; Eda, A.; Yanaka, I.; Honda, S.; Osawa, H.; Kaneko, Y.; Sugano, K. Conversion of gastric mucosa to intestinal metaplasia in Cdx2-expressing transgenic mice. Biochem. Biophys. Res. Commun. 2002, 294, 470–479. [Google Scholar] [CrossRef]
- Mutoh, H.; Sakurai, S.; Satoh, K.; Osawa, H.; Hakamata, Y.; Takeuchi, T.; Sugano, K. Cdx1 induced intestinal metaplasia in the transgenic mouse stomach: Comparative study with Cdx2 transgenic mice. Gut 2004, 53, 1416–1423. [Google Scholar] [CrossRef] [PubMed]
- Friis-Hansen, L.; Rieneck, K.; Nilsson, H.O.; Wadstrom, T.; Rehfeld, J.F. Gastric inflammation, metaplasia, and tumor development in gastrin-deficient mice. Gastroenterology 2006, 131, 246–258. [Google Scholar] [CrossRef]
- Zavros, Y.; Eaton, K.A.; Kang, W.; Rathinavelu, S.; Katukuri, V.; Kao, J.Y.; Samuelson, L.C.; Merchant, J.L. Chronic gastritis in the hypochlorhydric gastrin-deficient mouse progresses to adenocarcinoma. Oncogene 2005, 24, 2354–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakagami, T.; Dixon, M.; O’Rourke, J.; Howlett, R.; Alderuccio, F.; Vella, J.; Shimoyama, T.; Lee, A. Atrophic gastric changes in both Helicobacter felis and Helicobacter pylori infected mice are host dependent and separate from antral gastritis. Gut 1996, 39, 639–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.C.; Goldenring, J.R.; Dangler, C.; Ito, S.; Mueller, A.; Jeon, W.K.; Koh, T.J.; Fox, J.G. Mice lacking secretory phospholipase A2 show altered apoptosis and differentiation with Helicobacter felis infection. Gastroenterology 1998, 114, 675–689. [Google Scholar] [CrossRef]
- Honda, S.; Fujioka, T.; Tokieda, M.; Satoh, R.; Nishizono, A.; Nasu, M. Development of Helicobacter pylori-induced gastric carcinoma in Mongolian gerbils. Cancer Res. 1998, 58, 4255–4259. [Google Scholar]
- Watanabe, H.; Fujii, I.; Terada, Y. Induction of intestinal metaplasia in the rat gastric mucosa by local X-irradiation. Pathol Res. Pract 1980, 170, 104–114. [Google Scholar] [CrossRef]
- Zheng, Q.; Chen, X.Y.; Shi, Y.; Xiao, S.D. Development of gastric adenocarcinoma in Mongolian gerbils after long-term infection with Helicobacter pylori. J. Gastroenterol. Hepatol. 2004, 19, 1192–1198. [Google Scholar] [CrossRef]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Powell, S.M.; Zilz, N.; Beazer-Barclay, Y.; Bryan, T.M.; Hamilton, S.R.; Thibodeau, S.N.; Vogelstein, B.; Kinzler, K.W. APC mutations occur early during colorectal tumorigenesis. Nature 1992, 359, 235–237. [Google Scholar] [CrossRef]
- Bettington, M.; Walker, N.; Clouston, A.; Brown, I.; Leggett, B.; Whitehall, V. The serrated pathway to colorectal carcinoma: Current concepts and challenges. Histopathology 2013, 62, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Langner, C. Serrated and non-serrated precursor lesions of colorectal cancer. Dig. Dis. 2015, 33, 28–37. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, E.M.F.; Wang, X.; Jansen, M.; Fessler, E.; Trinh, A.; de Rooij, L.P.; de Jong, J.H.; de Boer, O.J.; van Leersum, R.; Bijlsma, M.F.; et al. Poor-prognosis colon cancer is defined by a molecularly distinct subtype and develops from serrated precursor lesions. Nat. Med. 2013, 19, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Solano, J.; Conesa-Zamora, P.; Trujillo-Santos, J.; Makinen, M.J.; Perez-Guillermo, M. Tumour budding and other prognostic pathological features at invasive margins in serrated colorectal adenocarcinoma: A comparative study with conventional carcinoma. Histopathology 2011, 59, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Landau, M.S.; Kuan, S.F.; Chiosea, S.; Pai, R.K. BRAF-mutated microsatellite stable colorectal carcinoma: An aggressive adenocarcinoma with reduced CDX2 and increased cytokeratin 7 immunohistochemical expression. Hum. Pathol. 2014, 45, 1704–1712. [Google Scholar] [CrossRef]
- Sakamoto, N.; Feng, Y.; Stolfi, C.; Kurosu, Y.; Green, M.; Lin, J.; Green, M.E.; Sentani, K.; Yasui, W.; McMahon, M.; et al. BRAF(V600E) cooperates with CDX2 inactivation to promote serrated colorectal tumorigenesis. Elife 2017, 6. [Google Scholar] [CrossRef]
- Dalerba, P.; Sahoo, D.; Paik, S.; Guo, X.; Yothers, G.; Song, N.; Wilcox-Fogel, N.; Forgo, E.; Rajendran, P.S.; Miranda, S.P.; et al. CDX2 as a Prognostic Biomarker in Stage II and Stage III Colon Cancer. N. Engl. J. Med. 2016, 374, 211–222. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, K.J.; Rhee, Y.Y.; Bae, J.M.; Cho, N.Y.; Lee, H.S.; Kang, G.H. Gastric-type expression signature in serrated pathway-associated colorectal tumors. Hum. Pathol. 2015, 46, 643–656. [Google Scholar] [CrossRef]
- Tsai, J.H.; Lin, Y.L.; Cheng, Y.C.; Chen, C.C.; Lin, L.I.; Tseng, L.H.; Cheng, M.L.; Liau, J.Y.; Jeng, Y.M. Aberrant expression of annexin A10 is closely related to gastric phenotype in serrated pathway to colorectal carcinoma. Mod. Pathol. 2015, 28, 268–278. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.D.; Clendenning, M.; Williamson, E.; Pearson, S.A.; Walters, R.J.; Nagler, B.; Packenas, D.; Win, A.K.; Hopper, J.L.; Jenkins, M.A.; et al. Expression of MUC2, MUC5AC, MUC5B, and MUC6 mucins in colorectal cancers and their association with the CpG island methylator phenotype. Mod. Pathol. 2013, 26, 1642–1656. [Google Scholar] [CrossRef] [Green Version]
- Beck, F.; Chawengsaksophak, K.; Luckett, J.; Giblett, S.; Tucci, J.; Brown, J.; Poulsom, R.; Jeffery, R.; Wright, N.A. A study of regional gut endoderm potency by analysis of Cdx2 null mutant chimaeric mice. Dev. Biol. 2003, 255, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Stringer, E.J.; Duluc, I.; Saandi, T.; Davidson, I.; Bialecka, M.; Sato, T.; Barker, N.; Clevers, H.; Pritchard, C.A.; Winton, D.J.; et al. Cdx2 determines the fate of postnatal intestinal endoderm. Development 2012, 139, 465–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, K.; Pellon-Cardenas, O.; Sirihorachai, V.R.; Warder, B.N.; Kothari, O.A.; Perekatt, A.O.; Fokas, E.E.; Fullem, R.L.; Zhou, A.; Thackray, J.K.; et al. Degree of Tissue Differentiation Dictates Susceptibility to BRAF-Driven Colorectal Cancer. Cell Rep. 2017, 21, 3833–3845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groden, J.; Thliveris, A.; Samowitz, W.; Carlson, M.; Gelbert, L.; Albertsen, H.; Joslyn, G.; Stevens, J.; Spirio, L.; Robertson, M.; et al. Identification and characterization of the familial adenomatous polyposis coli gene. Cell 1991, 66, 589–600. [Google Scholar] [CrossRef]
- Fodde, R.; Edelmann, W.; Yang, K.; van Leeuwen, C.; Carlson, C.; Renault, B.; Breukel, C.; Alt, E.; Lipkin, M.; Khan, P.M.; et al. A targeted chain-termination mutation in the mouse Apc gene results in multiple intestinal tumors. Proc. Natl. Acad. Sci. USA 1994, 91, 8969–8973. [Google Scholar] [CrossRef] [Green Version]
- Aoki, K.; Tamai, Y.; Horiike, S.; Oshima, M.; Taketo, M.M. Colonic polyposis caused by mTOR-mediated chromosomal instability in Apc+/Delta716 Cdx2+/− compound mutant mice. Nat. Genet. 2003, 35, 323–330. [Google Scholar] [CrossRef]
- Calon, A.; Gross, I.; Lhermitte, B.; Martin, E.; Beck, F.; Duclos, B.; Kedinger, M.; Duluc, I.; Domon-Dell, C.; Freund, J.N. Different effects of the Cdx1 and Cdx2 homeobox genes in a murine model of intestinal inflammation. Gut 2007, 56, 1688–1695. [Google Scholar] [CrossRef]
- Chase, A.; Reiter, A.; Burci, L.; Cazzaniga, G.; Biondi, A.; Pickard, J.; Roberts, I.A.; Goldman, J.M.; Cross, N.C. Fusion of ETV6 to the caudal-related homeobox gene CDX2 in acute myeloid leukemia with the t(12;13)(p13;q12). Blood 1999, 93, 1025–1031. [Google Scholar] [CrossRef]
- Bohlander, S.K. ETV6: A versatile player in leukemogenesis. Semin Cancer Biol. 2005, 15, 162–174. [Google Scholar] [CrossRef]
- Hock, H.; Meade, E.; Medeiros, S.; Schindler, J.W.; Valk, P.J.; Fujiwara, Y.; Orkin, S.H. Tel/Etv6 is an essential and selective regulator of adult hematopoietic stem cell survival. Genes Dev. 2004, 18, 2336–2341. [Google Scholar] [CrossRef]
- Rawat, V.P.; Cusan, M.; Deshpande, A.; Hiddemann, W.; Quintanilla-Martinez, L.; Humphries, R.K.; Bohlander, S.K.; Feuring-Buske, M.; Buske, C. Ectopic expression of the homeobox gene Cdx2 is the transforming event in a mouse model of t(12;13)(p13;q12) acute myeloid leukemia. Proc. Natl. Acad. Sci. USA 2004, 101, 817–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoene, S.; Rawat, V.P.; Heilmeier, B.; Hoster, E.; Metzeler, K.H.; Herold, T.; Hiddemann, W.; Gokbuget, N.; Hoelzer, D.; Bohlander, S.K.; et al. The homeobox gene CDX2 is aberrantly expressed and associated with an inferior prognosis in patients with acute lymphoblastic leukemia. Leukemia 2009, 23, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Scholl, C.; Bansal, D.; Dohner, K.; Eiwen, K.; Huntly, B.J.; Lee, B.H.; Rucker, F.G.; Schlenk, R.F.; Bullinger, L.; Dohner, H.; et al. The homeobox gene CDX2 is aberrantly expressed in most cases of acute myeloid leukemia and promotes leukemogenesis. J. Clin. Invest. 2007, 117, 1037–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedt, T.; Ebinger, M.; Salih, H.R.; Tomiuk, J.; Handgretinger, R.; Kanz, L.; Grunebach, F.; Lengerke, C. Aberrant expression of the homeobox gene CDX2 in pediatric acute lymphoblastic leukemia. Blood 2009, 113, 4049–4051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawat, V.P.; Thoene, S.; Naidu, V.M.; Arseni, N.; Heilmeier, B.; Metzeler, K.; Petropoulos, K.; Deshpande, A.; Quintanilla-Martinez, L.; Bohlander, S.K.; et al. Overexpression of CDX2 perturbs HOX gene expression in murine progenitors depending on its N-terminal domain and is closely correlated with deregulated HOX gene expression in human acute myeloid leukemia. Blood 2008, 111, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, H.J.; Largman, C. Homeobox genes in normal hematopoiesis and leukemia. Blood 1992, 80, 2445–2453. [Google Scholar] [CrossRef]
- Lawrence, H.J.; Sauvageau, G.; Ahmadi, N.; Lopez, A.R.; LeBeau, M.M.; Link, M.; Humphries, K.; Largman, C. Stage-and lineage-specific expression of the HOXA10 homeobox gene in normal and leukemic hematopoietic cells. Exp. Hematol. 1995, 23, 1160–1166. [Google Scholar]
- Thorsteinsdottir, U.; Kroon, E.; Jerome, L.; Blasi, F.; Sauvageau, G. Defining roles for HOX and MEIS1 genes in induction of acute myeloid leukemia. Mol. Cell Biol. 2001, 21, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Thorsteinsdottir, U.; Sauvageau, G.; Humphries, R.K. Hox homeobox genes as regulators of normal and leukemic hematopoiesis. Hematol. Oncol. Clin. N. Am. 1997, 11, 1221–1237. [Google Scholar] [CrossRef]
- Golub, T.R.; Slonim, D.K.; Tamayo, P.; Huard, C.; Gaasenbeek, M.; Mesirov, J.P.; Coller, H.; Loh, M.L.; Downing, J.R.; Caligiuri, M.A.; et al. Molecular classification of cancer: Class discovery and class prediction by gene expression monitoring. Science 1999, 286, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Haferlach, C.; Mecucci, C.; Schnittger, S.; Kohlmann, A.; Mancini, M.; Cuneo, A.; Testoni, N.; Rege-Cambrin, G.; Santucci, A.; Vignetti, M.; et al. AML with mutated NPM1 carrying a normal or aberrant karyotype show overlapping biologic, pathologic, immunophenotypic, and prognostic features. Blood 2009, 114, 3024–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krivtsov, A.V.; Armstrong, S.A. MLL translocations, histone modifications and leukaemia stem-cell development. Nat. Rev. Cancer 2007, 7, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.L.; Licht, J.D. HOX deregulation in acute myeloid leukemia. J. Clin. Investig. 2007, 117, 865–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineault, N.; Helgason, C.D.; Lawrence, H.J.; Humphries, R.K. Differential expression of Hox, Meis1, and Pbx1 genes in primitive cells throughout murine hematopoietic ontogeny. Exp. Hematol. 2002, 30, 49–57. [Google Scholar] [CrossRef]
- Sauvageau, G.; Lansdorp, P.M.; Eaves, C.J.; Hogge, D.E.; Dragowska, W.H.; Reid, D.S.; Largman, C.; Lawrence, H.J.; Humphries, R.K. Differential expression of homeobox genes in functionally distinct CD34+ subpopulations of human bone marrow cells. Proc. Natl. Acad. Sci. USA 1994, 91, 12223–12227. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, H.J.; Christensen, J.; Fong, S.; Hu, Y.L.; Weissman, I.; Sauvageau, G.; Humphries, R.K.; Largman, C. Loss of expression of the Hoxa-9 homeobox gene impairs the proliferation and repopulating ability of hematopoietic stem cells. Blood 2005, 106, 3988–3994. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, H.J.; Helgason, C.D.; Sauvageau, G.; Fong, S.; Izon, D.J.; Humphries, R.K.; Largman, C. Mice bearing a targeted interruption of the homeobox gene HOXA9 have defects in myeloid, erythroid, and lymphoid hematopoiesis. Blood 1997, 89, 1922–1930. [Google Scholar] [CrossRef]
- Lengerke, C.; Daley, G.Q. Patterning definitive hematopoietic stem cells from embryonic stem cells. Exp. Hematol. 2005, 33, 971–979. [Google Scholar] [CrossRef]
- Subramanian, V.; Meyer, B.I.; Gruss, P. Disruption of the murine homeobox gene Cdx1 affects axial skeletal identities by altering the mesodermal expression domains of Hox genes. Cell 1995, 83, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Davidson, A.J.; Ernst, P.; Wang, Y.; Dekens, M.P.; Kingsley, P.D.; Palis, J.; Korsmeyer, S.J.; Daley, G.Q.; Zon, L.I. cdx4 mutants fail to specify blood progenitors and can be rescued by multiple hox genes. Nature 2003, 425, 300–306. [Google Scholar] [CrossRef]
- Davidson, A.J.; Zon, L.I. The caudal-related homeobox genes cdx1a and cdx4 act redundantly to regulate hox gene expression and the formation of putative hematopoietic stem cells during zebrafish embryogenesis. Dev. Biol. 2006, 292, 506–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gering, M.; Rodaway, A.R.; Gottgens, B.; Patient, R.K.; Green, A.R. The SCL gene specifies haemangioblast development from early mesoderm. EMBO J. 1998, 17, 4029–4045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charite, J.; de Graaff, W.; Consten, D.; Reijnen, M.J.; Korving, J.; Deschamps, J. Transducing positional information to the Hox genes: Critical interaction of cdx gene products with position-sensitive regulatory elements. Development 1998, 125, 4349–4358. [Google Scholar]
- Hunter, C.P.; Harris, J.M.; Maloof, J.N.; Kenyon, C. Hox gene expression in a single Caenorhabditis elegans cell is regulated by a caudal homolog and intercellular signals that inhibit wnt signaling. Development 1999, 126, 805–814. [Google Scholar] [PubMed]
- Wang, Y.; Yabuuchi, A.; McKinney-Freeman, S.; Ducharme, D.M.; Ray, M.K.; Chawengsaksophak, K.; Archer, T.K.; Daley, G.Q. Cdx gene deficiency compromises embryonic hematopoiesis in the mouse. Proc. Natl. Acad. Sci. USA 2008, 105, 7756–7761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooke-Bisschop, T.; Savory, J.G.A.; Foley, T.; Ringuette, R.; Lohnes, D. Essential roles for Cdx in murine primitive hematopoiesis. Dev. Biol. 2017, 422, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Lengerke, C.; Grauer, M.; Niebuhr, N.I.; Riedt, T.; Kanz, L.; Park, I.H.; Daley, G.Q. Hematopoietic development from human induced pluripotent stem cells. Ann. N. Y. Acad. Sci. 2009, 1176, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengerke, C.; McKinney-Freeman, S.; Naveiras, O.; Yates, F.; Wang, Y.; Bansal, D.; Daley, G.Q. The cdx-hox pathway in hematopoietic stem cell formation from embryonic stem cells. Ann. N. Y. Acad. Sci. 2007, 1106, 197–208. [Google Scholar] [CrossRef]
- Lengerke, C.; Schmitt, S.; Bowman, T.V.; Jang, I.H.; Maouche-Chretien, L.; McKinney-Freeman, S.; Davidson, A.J.; Hammerschmidt, M.; Rentzsch, F.; Green, J.B.; et al. BMP and Wnt specify hematopoietic fate by activation of the Cdx-Hox pathway. Cell Stem Cell 2008, 2, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Chawengsaksophak, K.; de Graaff, W.; Rossant, J.; Deschamps, J.; Beck, F. Cdx2 is essential for axial elongation in mouse development. Proc. Natl. Acad. Sci. USA 2004, 101, 7641–7645. [Google Scholar] [CrossRef] [Green Version]
- Lengerke, C.; Wingert, R.; Beeretz, M.; Grauer, M.; Schmidt, A.G.; Konantz, M.; Daley, G.Q.; Davidson, A.J. Interactions between Cdx genes and retinoic acid modulate early cardiogenesis. Dev. Biol. 2011, 354, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, T.E.; Hess, B.; Savory, J.G.A.; Ringuette, R.; Lohnes, D. Role of Cdx factors in early mesodermal fate decisions. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
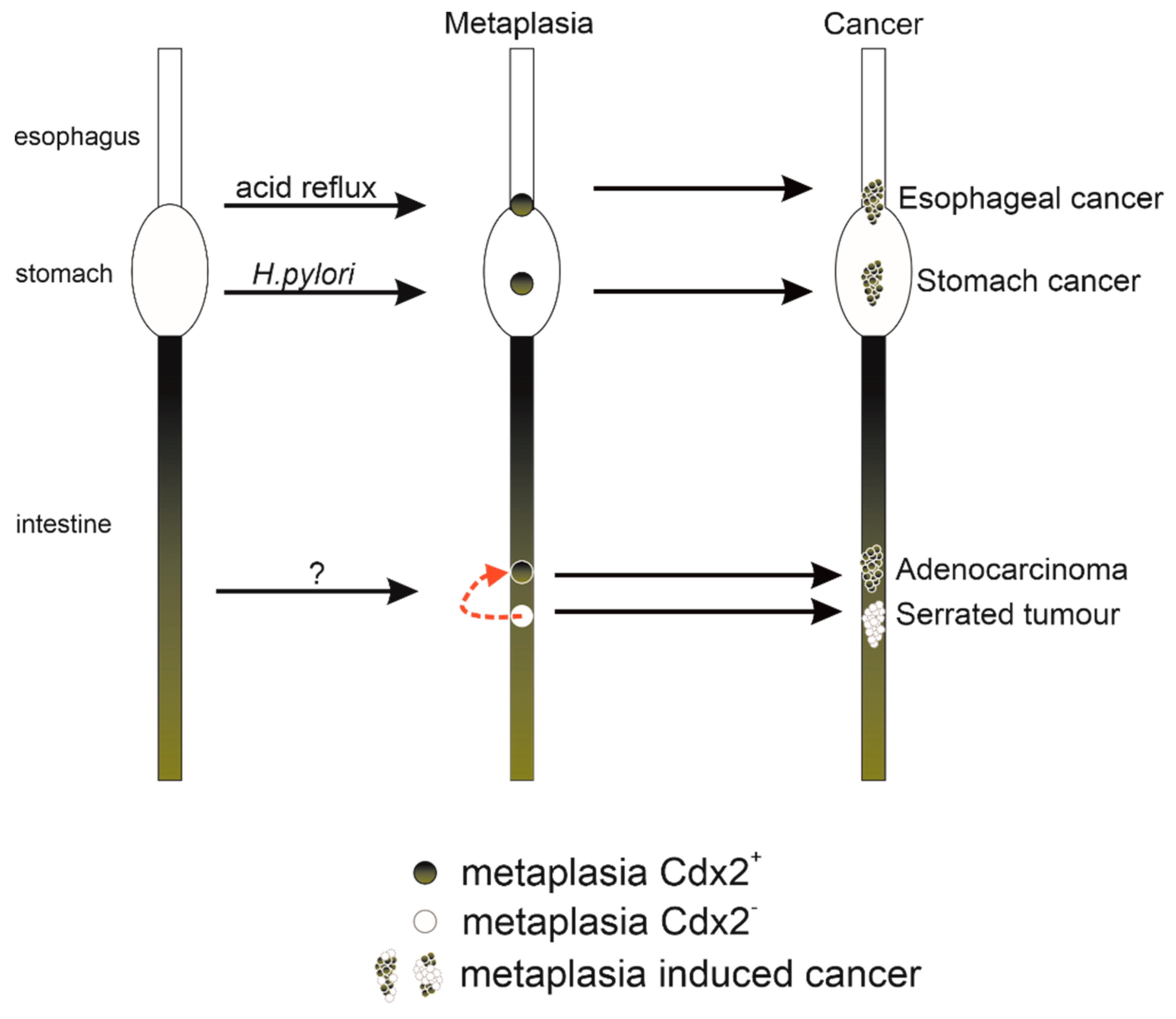
Figure 1.
Simplified schematic diagram depicting how metaplasia caused by the alteration of Cdx2 expression can progress to cancer.
Figure 1.
Simplified schematic diagram depicting how metaplasia caused by the alteration of Cdx2 expression can progress to cancer.


{kind=link}
Table 1.
Animal cancer models generated through the genetic manipulation of Cdx genes.
| Mutation | Phenotype | Reference |
|---|---|---|
| Cdx2KO | Homozygotes: preimplantation lethality Heterozygotes: anterior homeotic shift of vertebrae, nondysplastic colonic tumors often containing metaplastic/heterotopic foci with gastric features | [22,28,29] |
| Cdx2CKO; Apc+/Δ14 | Mixed tumors with adenomatous and serrated features | [35] |
| Tg(Foxa3–Cdx2) | Metaplasia in stomach | [33] |
| Tg(Atp4a–Cdx2) | Metaplasia in stomach | [32] |
| Tg(K14–Cdx2) | Non-intestinal type metaplasia in esophagus | [31] |
| Tg(Krt7rtTA); Tg(otet-Cdx2) | Intestinal type metaplasia in esophagus | [34] |
| Tg(krt5:cdx1b–EGFP) 1 | Metaplasia in esophagus | [30] |
1 Transgenic zebrafish model.
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chawengsaksophak, K. Cdx2 Animal Models Reveal Developmental Origins of Cancers. Genes 2019, 10, 928. https://doi.org/10.3390/genes10110928
AMA Style
Chawengsaksophak K. Cdx2 Animal Models Reveal Developmental Origins of Cancers. Genes. 2019; 10(11):928. https://doi.org/10.3390/genes10110928
Chicago/Turabian StyleChawengsaksophak, Kallayanee. 2019. "Cdx2 Animal Models Reveal Developmental Origins of Cancers" Genes 10, no. 11: 928. https://doi.org/10.3390/genes10110928
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.