
Yield Formation Parameters of Selected Winter Wheat Genotypes in Response to Water Shortage

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Soil Material
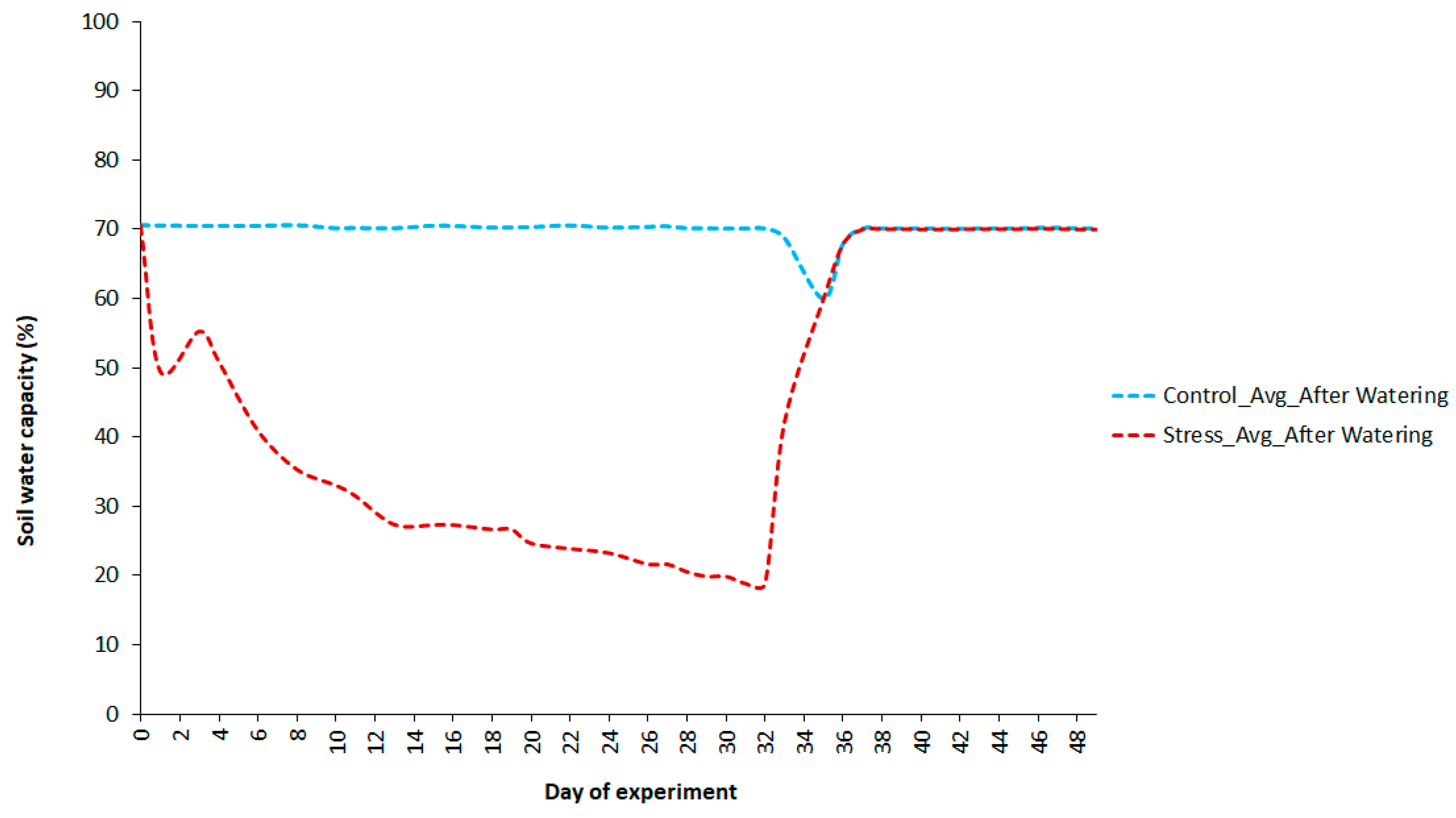
2.2. Control and Drought Stress Treatments
2.3. Data Processing and Statistical Analyses
3. Results
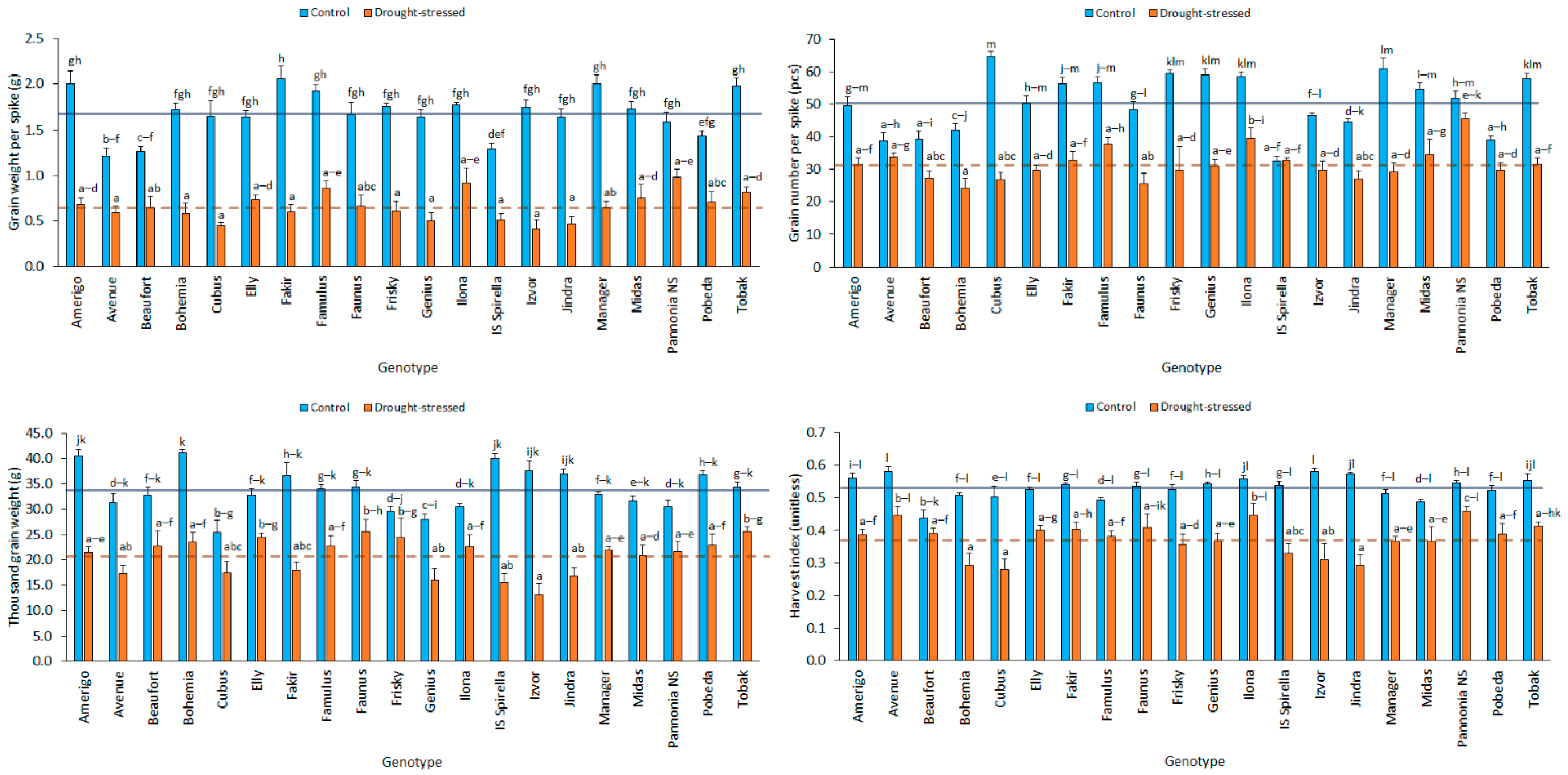
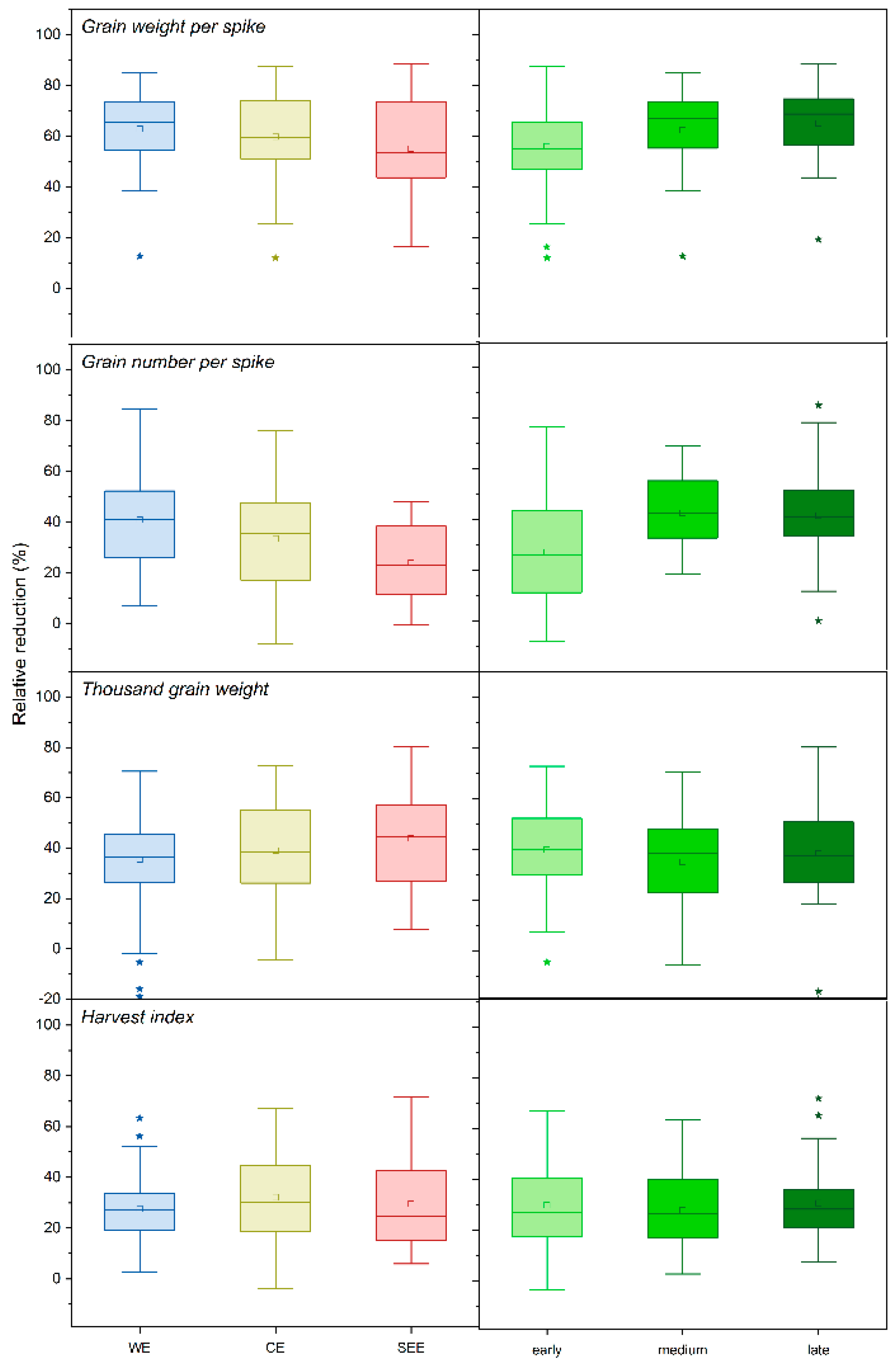
3.1. GWS of Winter Wheat Genotypes under Control and Drought Stress Treatments
3.2. GNS of Winter Wheat Genotypes under Control and Drought Stress Treatments
3.3. TGW of Winter Wheat Genotypes under Control and Drought Stress Treatments
3.4. HI of Winter Wheat Genotypes under Control and Drought Stress Treatments
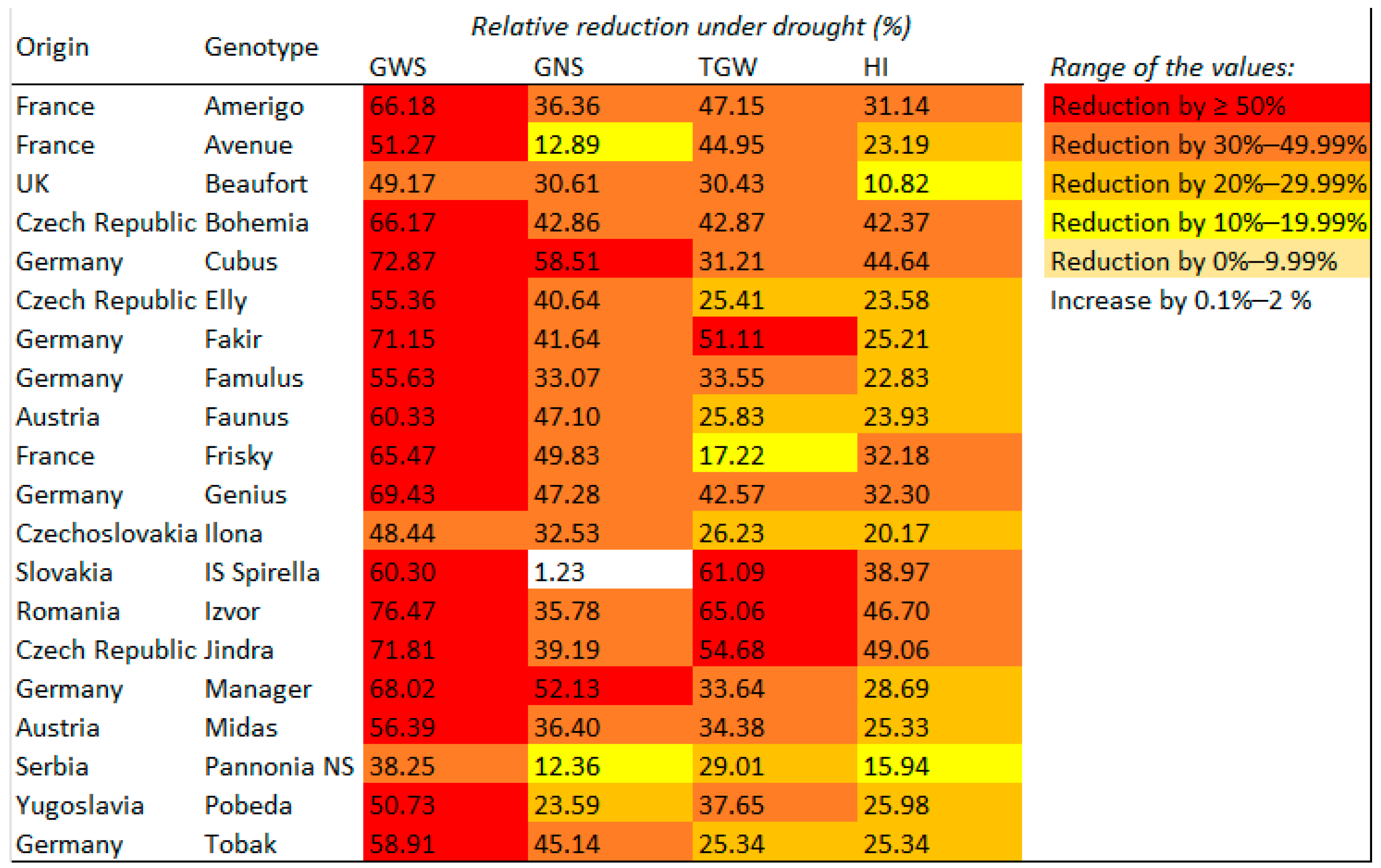
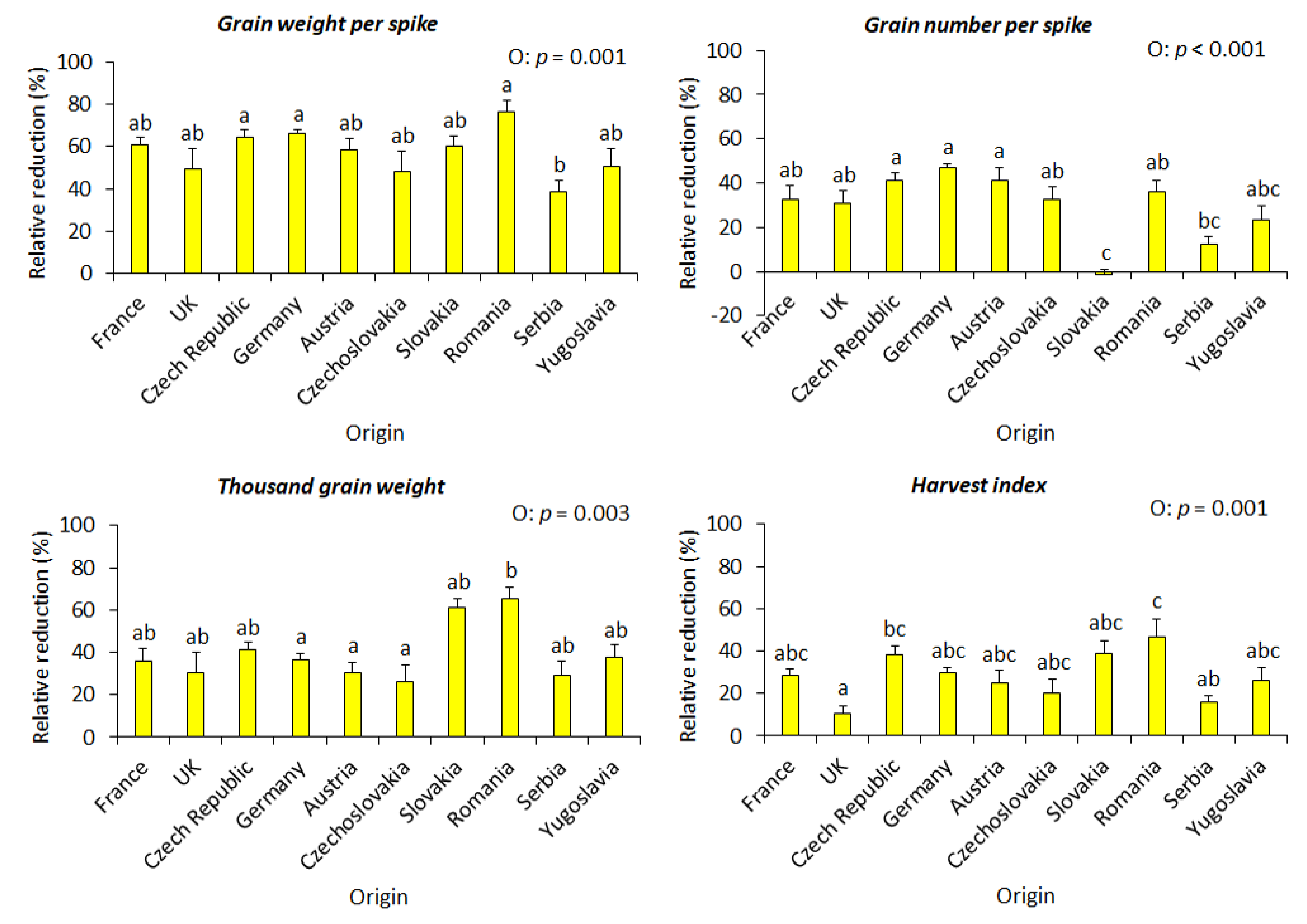
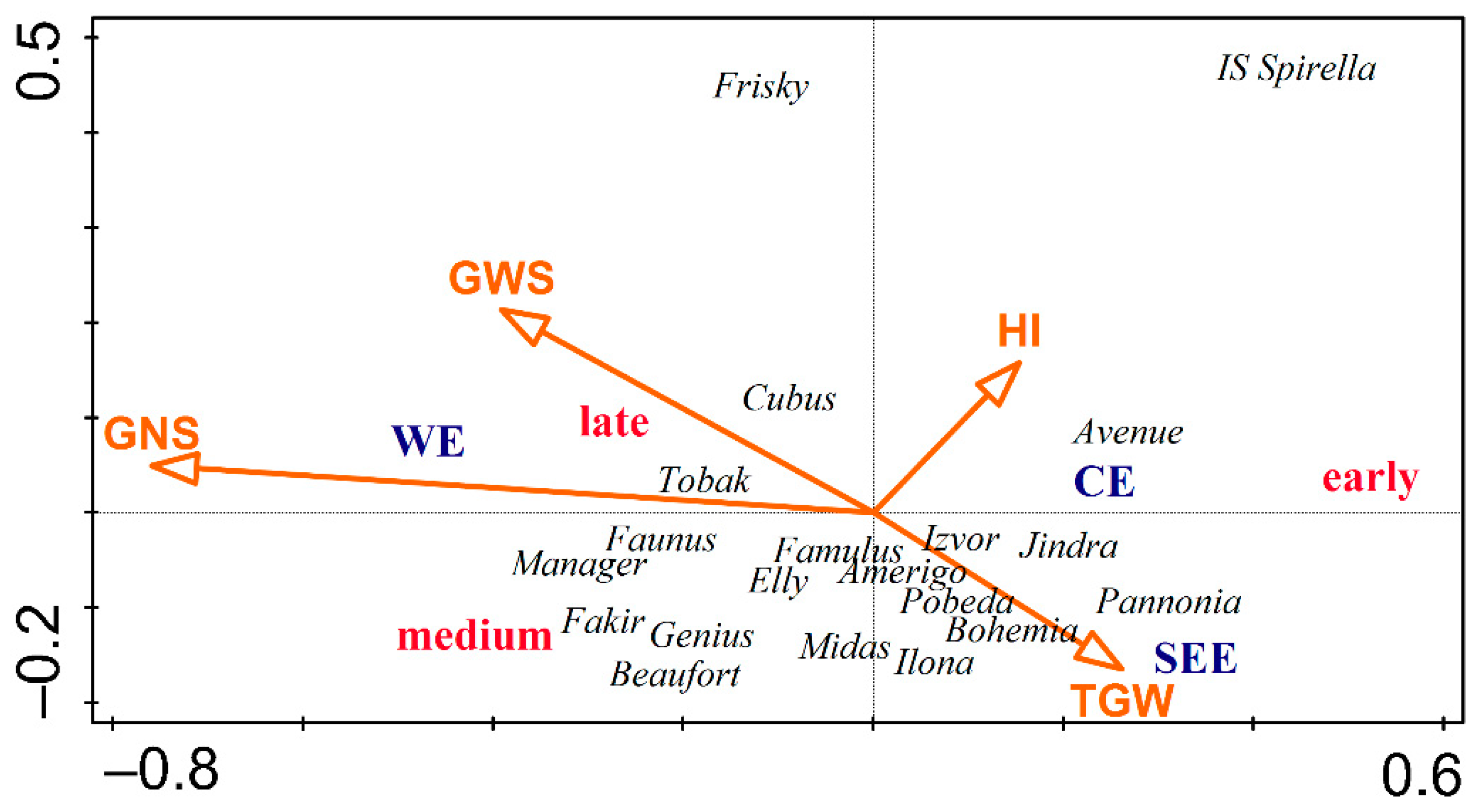
3.5. Yield Formation Parameters in Relation to the Country of Origin, Site of Origin, and Maturity
3.6. RDA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vautard, R.; Gobiet, A.; Sobolowski, S.; Kjellström, E.; Stegehuis, A.; Watkiss, P.; Mendlik, T.; Landgren, O.; Nikulin, G.; Teichmann, C.; et al. The European climate under a 2 °C global warming. Environ. Res. Lett. 2014, 9, 034006. [Google Scholar] [CrossRef]
- Jacob, D.; Kotova, L.; Teichmann, C.; Sobolowski, S.P.; Vautard, R.; Donnelly, C.; Koutroulis, A.G.; Grillakis, M.G.; Tsanis, I.K.; Damm, A.; et al. Climate impacts in Europe under +1.5 °C global warming. Earth’s Future 2018, 6, 264–285. [Google Scholar] [CrossRef] [Green Version]
- Kjellström, E.; Nikulin, G.; Strandberg, G.; Christensen, O.B.; Jacob, D.; Keuler, K.; Lenderink, G.; van Meijgaard, E.; Schär, C.; Somot, S.; et al. European climate change at global mean temperature increases of 1.5 and 2 °C above pre-industrial conditions as simulated by the EURO-CORDEX regional climate models. Earth Syst. Dynam. 2018, 9, 459–478. [Google Scholar] [CrossRef] [Green Version]
- IPCC (The Intergovernmental Panel on Climate Change). Summary for policymakers. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; World Meteorological Organization: Geneva, Switzerland, 2018; pp. 3–26. [Google Scholar]
- IPCC (The Intergovernmental Panel on Climate Change). Summary for policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 4–41. [Google Scholar]
- Brázdil, R.; Zahradníček, P.; Dobrovolný, P.; Štěpánek, P.; Trnka, M. Observed changes in precipitation during recent warming: The Czech Republic, 1961–2019. Int. J. Climatol. 2021, 41, 3881–3902. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought stress in wheat during flowering and grain-filling periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Hlaváčová, M.; Klem, K.; Rapantová, B.; Novotná, K.; Urban, O.; Hlavinka, P.; Smutná, P.; Horáková, V.; Škarpa, P.; Pohanková, E.; et al. Interactive effects of high temperature and drought stress during stem elongation, anthesis and early grain filling on the yield formation and photosynthesis of winter wheat. Field Crops Res. 2018, 221, 182–195. [Google Scholar] [CrossRef]
- Trnka, M.; Brázdil, R.; Balek, J.; Dubrovský, M.; Eitzinger, J.; Hlavinka, P.; Chuchma, F.; Možný, M.; Prášil, I.; Růžek, P.; et al. Observed changes in the agroclimatic zones in the Czech Republic between 1961 and 2019. Plant Soil Environ. 2021, 67, 154–163. [Google Scholar] [CrossRef]
- Fischer, T.; Byerlee, D.; Edmeades, G. Crop Yields and Global Food Security: Will Yield Increase Continue to Feed the World? ACIAR Monograph No. 158; Australian Centre for International Agricultural Research (ACIAR): Canberra, Australia, 2014.
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slafer, G.A.; Savin, R.; Pinochet, D.; Calderini, D.F. Chapter 3—Wheat. In Crop Physiology: Case Histories for Major Crops; Sadras, V.O., Calderini, D.F., Eds.; Academic Press (Imprint of Elsevier Inc.): London, UK, 2021; pp. 98–163. [Google Scholar]
- Vollset, S.E.; Goren, E.; Yuan, C.W.; Cao, J.; Smith, A.E.; Hsiao, T.; Bisignano, C.; Azhar, G.S.; Castro, E.; Chalek, J.; et al. Fertility, mortality, migration, and population scenarios for 195 countries and territories from 2017 to 2100: A forecasting analysis for the Global Burden of Disease Study. Lancet 2020, 396, 1285–1306. [Google Scholar] [CrossRef]
- Trnka, M.; Rötter, R.P.; Ruiz-Ramos, M.; Kersebaum, K.C.; Olesen, J.E.; Žalud, Z.; Semenov, M.A. Adverse weather conditions for European wheat production will become more frequent with climate change. Nat. Clim. Chang. 2014, 4, 637–643. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Ding, H.; Liu, D.; Liu, X.; Li, Y.; Kang, J.; Lv, J.; Wang, G. Photosynthetic and stomatal traits of spike and flag leaf of winter wheat (Triticum aestivum L.) under water deficit. Photosynthetica 2018, 56, 687–697. [Google Scholar] [CrossRef]
- de Oliveira Silva, A.; Slafer, G.A.; Fritz, A.K.; Lollato, R.P. Physiological basis of genotypic response to management in dryland wheat. Front. Plant Sci. 2020, 10, 1644. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Iizumi, T.; Nishimori, M. Global patterns of crop production losses associated with droughts from 1983 to 2009. J. Appl. Meteorol. Climatol. 2019, 58, 1233–1244. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.-A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, S.; Cheng, M.; Jiang, H.; Zhang, X.; Peng, C.; Lu, X.; Zhang, M.; Jin, J. Effect of drought on agronomic traits of rice and wheat: A meta-analysis. Int. J. Environ. Res. Public Health 2018, 15, 839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [Green Version]
- Hein, N.T.; Ciampitti, I.A.; Jagadish, S.V.K. Bottlenecks and opportunities in field-based high-throughput phenotyping for heat and drought stress. J. Exp. Bot. 2021, 72, 5102–5116. [Google Scholar] [CrossRef]
- Bodner, G.; Nakhforoosh, A.; Kaul, H.-P. Management of crop water under drought: A review. Agron. Sustain. Dev. 2015, 35, 401–442. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using Canoco 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Sarto, M.V.M.; Sarto, J.R.W.; Rampim, L.; Rosset, J.S.; Bassegio, D.; da Costa, P.F.; Inagaki, A.M. Wheat phenology and yield under drought: A review. Aust. J. Crop Sci. 2017, 11, 941–946. [Google Scholar] [CrossRef]
- Pradhan, G.P.; Prasad, P.V.V.; Fritz, A.K.; Kirkham, M.B.; Gill, B.S. Effects of drought and high temperature stress on synthetic hexaploidy wheat. Funct. Plant Biol. 2012, 39, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Raja, N.I.; Yasmeen, F.; Hussain, M.; Ejaz, M.; Shah, M.A. Impacts of heat stress on wheat: A critical review. Adv. Crop Sci. Technol. 2017, 5, 251. [Google Scholar] [CrossRef]
- Mahrookashani, A.; Siebert, S.; Hüging, H.; Ewert, F. Independent and combined effects of high temperature and drought stress around anthesis on wheat. J. Agron. Crop Sci. 2017, 203, 453–463. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Pisipati, S.R.; Momčilović, I.; Ristic, Z. Independent and combined effect of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat. J. Agron. Crop Sci. 2011, 197, 430–441. [Google Scholar] [CrossRef]
- Saedi, M.; Abdoli, M. Effect of drought stress during grain filling on yield and its components, gas exchange variables, and some physiological traits of wheat cultivars. J. Agric. Sci. Technol. 2015, 17, 885–898. [Google Scholar]
- Shamsi, K.; Kobraee, S. Bread wheat production under drought stress conditions. Ann. Biol. Res. 2011, 2, 352–358. [Google Scholar]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H. Effects of pre-anthesis drought, heat and their combination on the growth, yield and physiology of diverse wheat (Triticum aestivum L.) genotypes varying in sensitivity to heat and drought stress. Sci. Rep. 2019, 9, 6955. [Google Scholar] [CrossRef] [Green Version]
- Webber, H.; Ewert, F.; Olesen, J.E.; Müller, C.; Fronzek, S.; Ruane, A.C.; Bourgault, M.; Martre, P.; Ababaei, B.; Bindi, M.; et al. Diverging importance of drought stress for maize and winter wheat in Europe. Nat. Commun. 2018, 9, 4249. [Google Scholar] [CrossRef] [Green Version]
- Brázdil, R.; Trnka, M.; Dobrovolný, P.; Chromá, K.; Hlavinka, P.; Žalud, Z. Variability of droughts in the Czech Republic, 1881–2006. Theor. Appl. Climatol. 2009, 97, 297–315. [Google Scholar] [CrossRef]
- Řehoř, J.; Brázdil, R.; Trnka, M.; Lhotka, O.; Balek, J.; Možný, M.; Štěpánek, P.; Zahradníček, P.; Mikulová, K.; Turňa, M. Soil drought and circulation types in a longitudinal transect over central Europe. Int. J. Climatol. 2021, 41, 2834–2850. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; de Groot, S.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef] [PubMed]
- Meyre, D.; Leonardi, A.; Brisson, G.; Vartanian, N. Drought-adaptive mechanisms involved in the escape/tolerance strategies of Arabidopsis Landsberg erecta and Columbia ecotypes and their F1 reciprocal progeny. J. Plant Physiol. 2001, 158, 1145–1152. [Google Scholar] [CrossRef]
- Yashavanthakumar, K.J.; Baviskar, V.S.; Navathe, S.; Patil, R.M.; Bagwan, J.H.; Bankar, D.N.; Gite, V.D.; Gopalareddy, K.; Mishra, C.N.; Mamrutha, H.M.; et al. Impact of heat and drought stress on phenological development and yield in bread wheat. Plant Physiol. Rep. 2021, 26, 357–367. [Google Scholar] [CrossRef]
- Isidro, J.; Álvaro, F.; Royo, C.; Villegas, D.; Miralles, D.J.; García del Moral, L.F. Changes in duration of developmental phases of durum wheat caused by breeding in Spain and Italy during the 20th century and its impact on yield. Ann. Bot. 2011, 107, 1355–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Bidinger, F.R.; Witcombe, J.R. Evaluation of specific drought avoidance traits as selection criteria for improvement of drought resistance. In Drought Resistance in Cereals; Baker, F.W.G., Ed.; CAB International: Wallingford, UK, 1989; pp. 151–164. [Google Scholar]
- Fischer, R.A. The effect of water stress at various stages of development on yield processes in wheat. In Plant Response to Climatic Factors, Proceedings of the Uppsala Symposium 1970: Ecology and Conservation, Uppsala, Sweden, 15–20 September 1970; Slatyer, R.O., Ed.; UNESCO: Paris, France, 1973; Volume 5, pp. 233–241. [Google Scholar]
- Li, P.; Ma, B.; Palta, J.A.; Ding, T.; Cheng, Z.; Lv, G.; Xiong, Y. Wheat breeding highlights drought tolerance while ignores the advantages of drought avoidance: A meta-analysis. Eur. J. Agron. 2021, 122, 126196. [Google Scholar] [CrossRef]
- Levitt, J. Stress terminology. In Adaptation of Plants to Water and High Temperature Stress; Turner, N.C., Kramer, P.J., Eds.; J. Wiley: New York, NY, USA, 1980; pp. 437–439. [Google Scholar]
- Anyia, A.O.; Herzog, H. Water-use efficiency, leaf area and leaf gas exchange of cowpeas under mid-season drought. Eur. J. Agron. 2004, 20, 327–339. [Google Scholar] [CrossRef]
- Fufa, H.; Baenziger, P.S.; Beecher, B.S.; Graybosch, R.A.; Eskridge, K.M.; Nelson, L.A. Genetic improvement trends in agronomic performances and end-use quality characteristics among hard red winter wheat cultivars in Nebraska. Euphytica 2005, 144, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Staggenborg, S.A.; Ristic, Z. Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. In Response of Crops to Limited Water: Understanding and Modeling Water Stress Effects on Plant Growth Processes, Advances in Agricultural Systems Modeling; Ahuja, L.R., Reddy, V.R., Saseendran, S.A., Yu, Q., Eds.; ASA, CSSA, SSSA: Madison, WI, USA, 2008; pp. 301–355. [Google Scholar]
- Munns, R.; Richards, R.A. Recent advances in breeding wheat for drought and salt stresses. In Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops; Jenks, M.A., Hasegawa, P.M., Jain, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 565–585. [Google Scholar]
- Tariq, M.; Mahmood, A.; Mian, M.A.; Cheema, N.M.; Sabar, M.; Ihsan, M.; Rehman, A.U. Dharabi-11: A new high yielding drought and disease tolerant wheat variety. Int. J. Agric. Biol. 2013, 15, 701–706. [Google Scholar]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 2013, 610721. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Genotype—Maturity | Origin | Genotype—Maturity |
|---|---|---|---|
| Czechoslovakia */Czech Republic/Slovakia, CE | Ilona—E | Germany, WE | Cubus—M |
| Bohemia—E | Fakir ▲—L | ||
| Elly—E | Famulus—M | ||
| Jindra—E | Genius—M | ||
| IS Spirella—E | Manager—L | ||
| Tobak—L | |||
| Austria, CE | Faunus—M | ||
| Midas—E | United Kingdom, WE | Beaufort—M | |
| France, WE | Amerigo—M | Romania, SEE | Izvor—L |
| Avenue—E | |||
| Frisky—L | Yugoslavia */Serbia, SEE | Pobeda •—L | |
| Pannonia NS ▲—E |
| GWS | GNS | TGW | HI | ||
|---|---|---|---|---|---|
| Two-way ANOVA p-values | Genotype (G) | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Treatment (T) | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| G × T | <0.0001 | <0.0001 | <0.0001 | 0.0005 | |
| Relative reduction due to drought stress (%) | 61.26 | 37.61 | 38.98 | 29.71 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hlaváčová, M.; Klem, K.; Pytela, J.; Veselá, B.; Hlavinka, P.; Smutná, P.; Horáková, V.; Škarpa, P.; Trnka, M. Yield Formation Parameters of Selected Winter Wheat Genotypes in Response to Water Shortage. Agronomy 2022, 12, 831. https://doi.org/10.3390/agronomy12040831
Hlaváčová M, Klem K, Pytela J, Veselá B, Hlavinka P, Smutná P, Horáková V, Škarpa P, Trnka M. Yield Formation Parameters of Selected Winter Wheat Genotypes in Response to Water Shortage. Agronomy. 2022; 12(4):831. https://doi.org/10.3390/agronomy12040831
Chicago/Turabian StyleHlaváčová, Marcela, Karel Klem, Jaromír Pytela, Barbora Veselá, Petr Hlavinka, Pavlína Smutná, Vladimíra Horáková, Petr Škarpa, and Miroslav Trnka. 2022. "Yield Formation Parameters of Selected Winter Wheat Genotypes in Response to Water Shortage" Agronomy 12, no. 4: 831. https://doi.org/10.3390/agronomy12040831