
Vaccinia Virus Expressing Interferon Regulatory Factor 3 Induces Higher Protective Immune Responses against Lethal Poxvirus Challenge in Atopic Organism

 , and
, and
Abstract
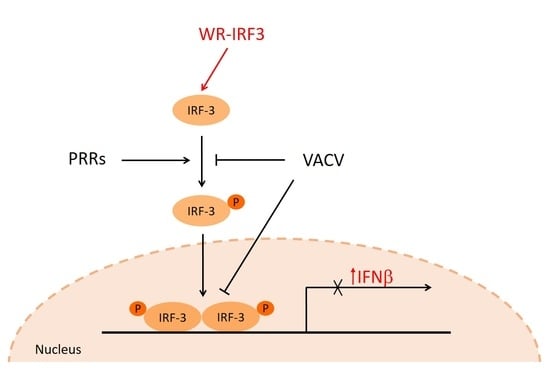
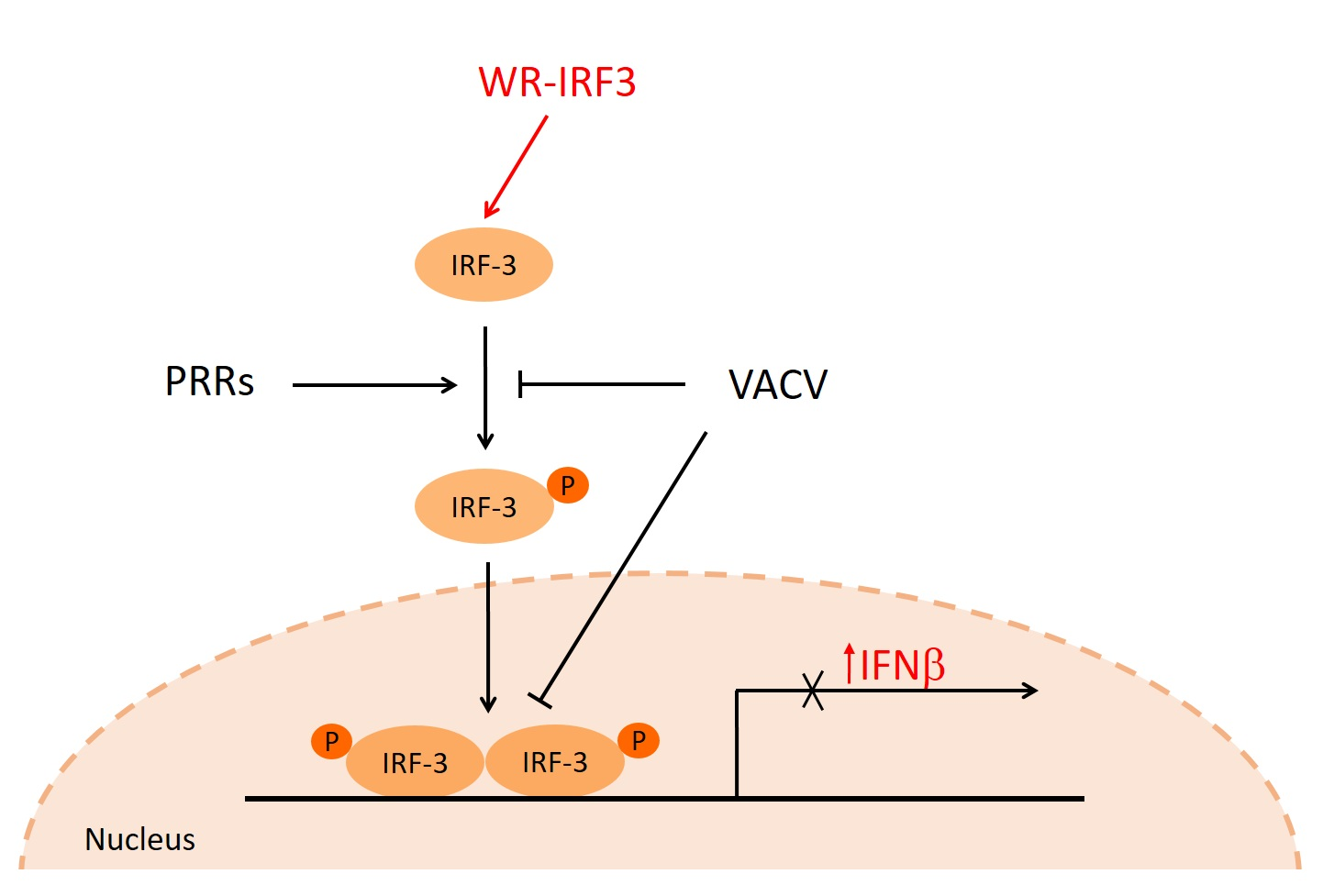
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Chemicals
2.3. Cells
2.4. Viruses
2.5. Animals and Infection
2.6. Determination of Virus Titer
2.7. Determination of mRNA Levels
2.8. Western Blot Analysis
2.9. Fluorescence Microscopy
2.10. Statistical Analyses
3. Results
3.1. Expression of Mouse Recombinant IRF-3 in Cell Lines and Its Effects
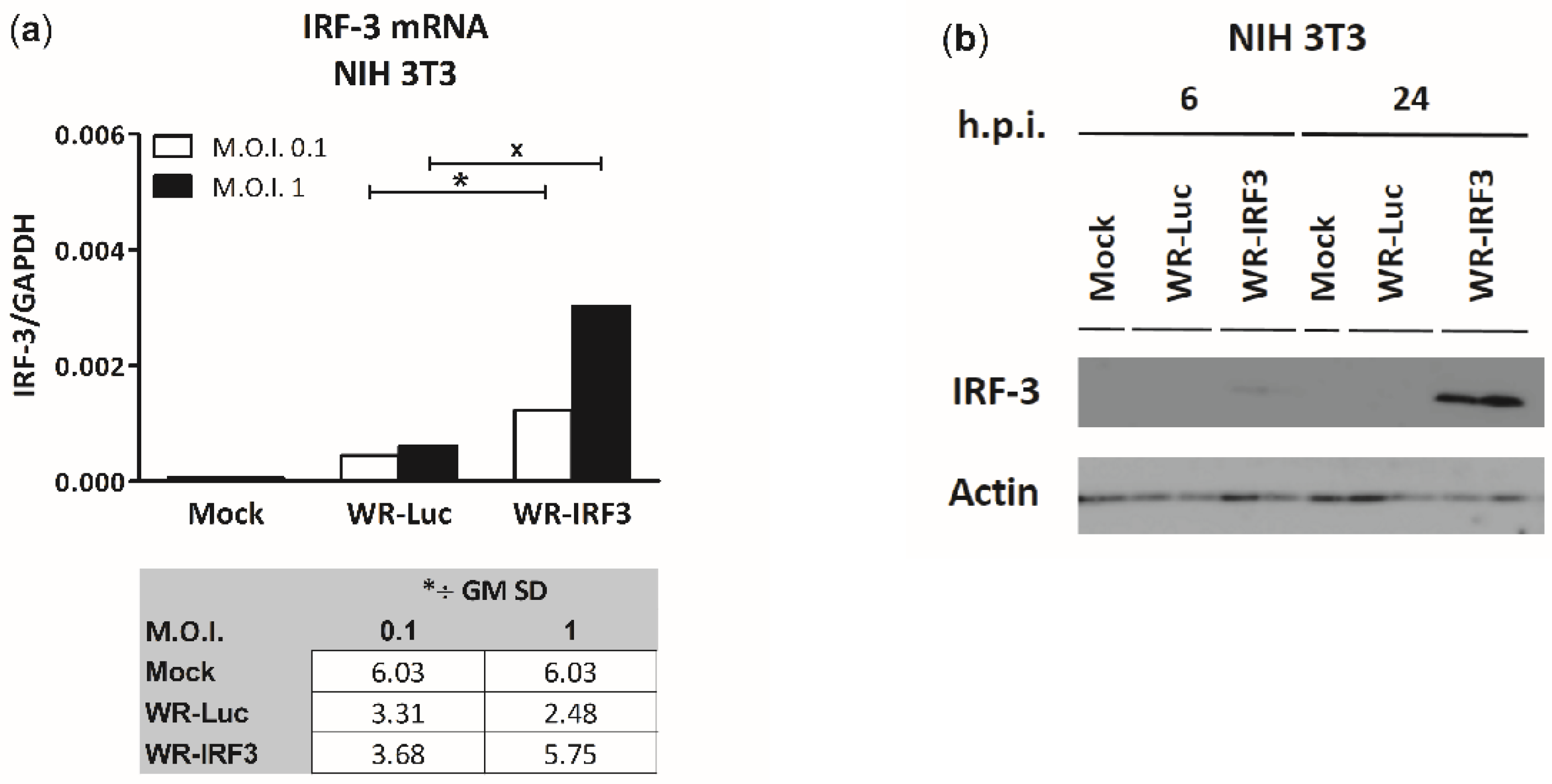
3.1.1. Expression of IRF-3 by WR-IRF3
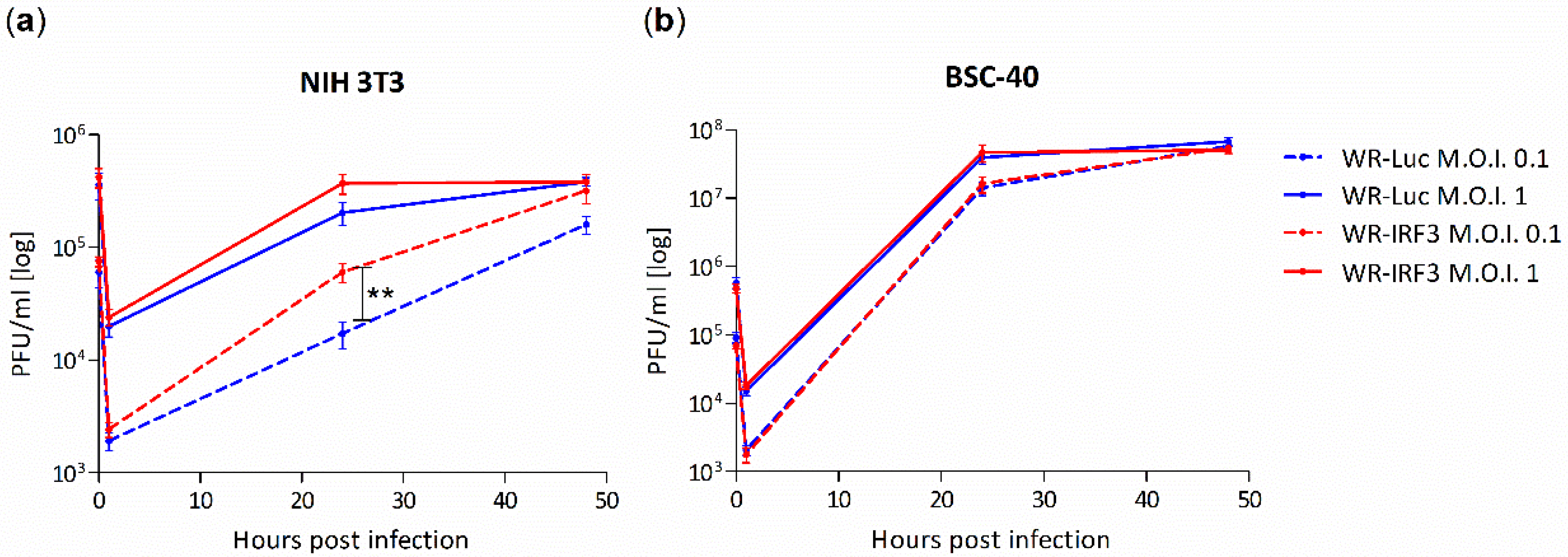
3.1.2. Growth of WR-IRF3 Is Increased in Mouse Cells
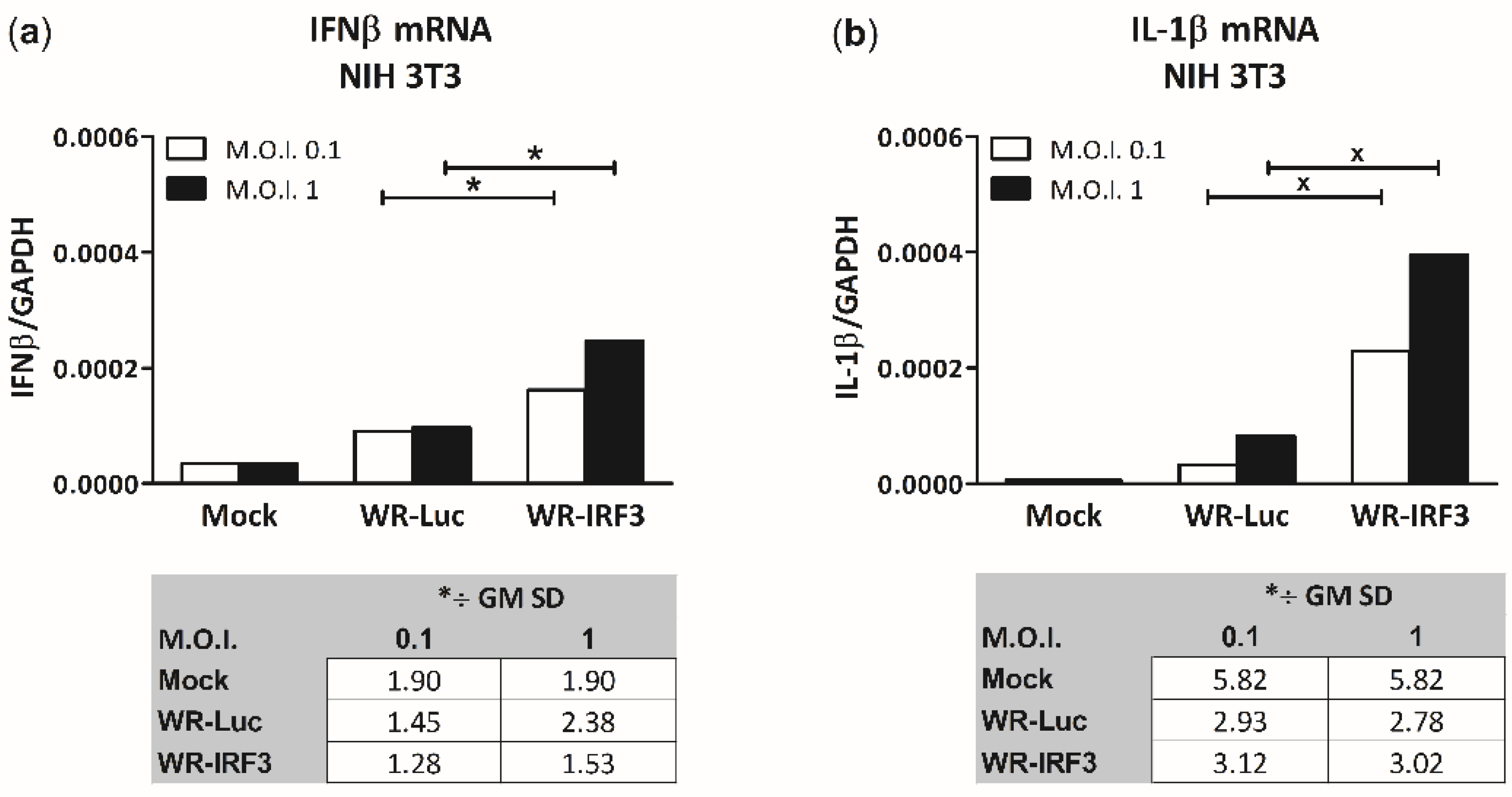
3.1.3. WR-IRF3 Induces Increased Levels of IFNβ and IL-1β mRNA
3.1.4. WR-IRF3 Induces Increased Apoptosis in Mouse Cells
3.2. Experiments with Mice
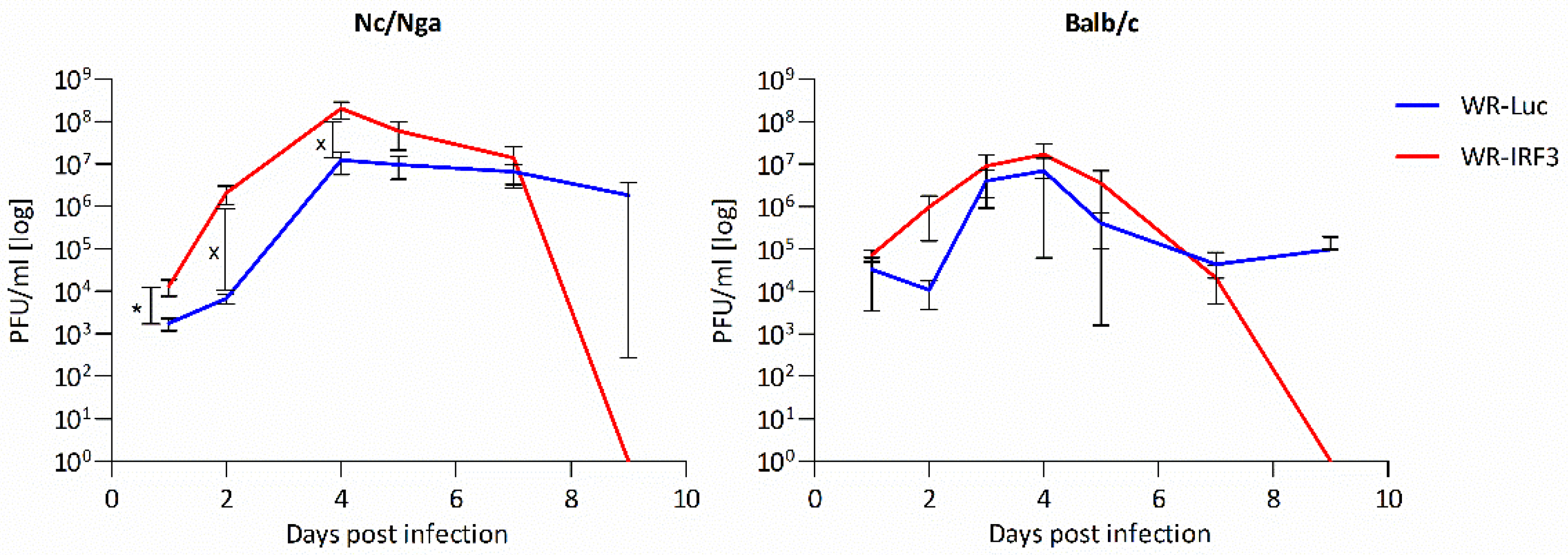
3.2.1. Growth of WR-IRF3 in Mice and Its Effects
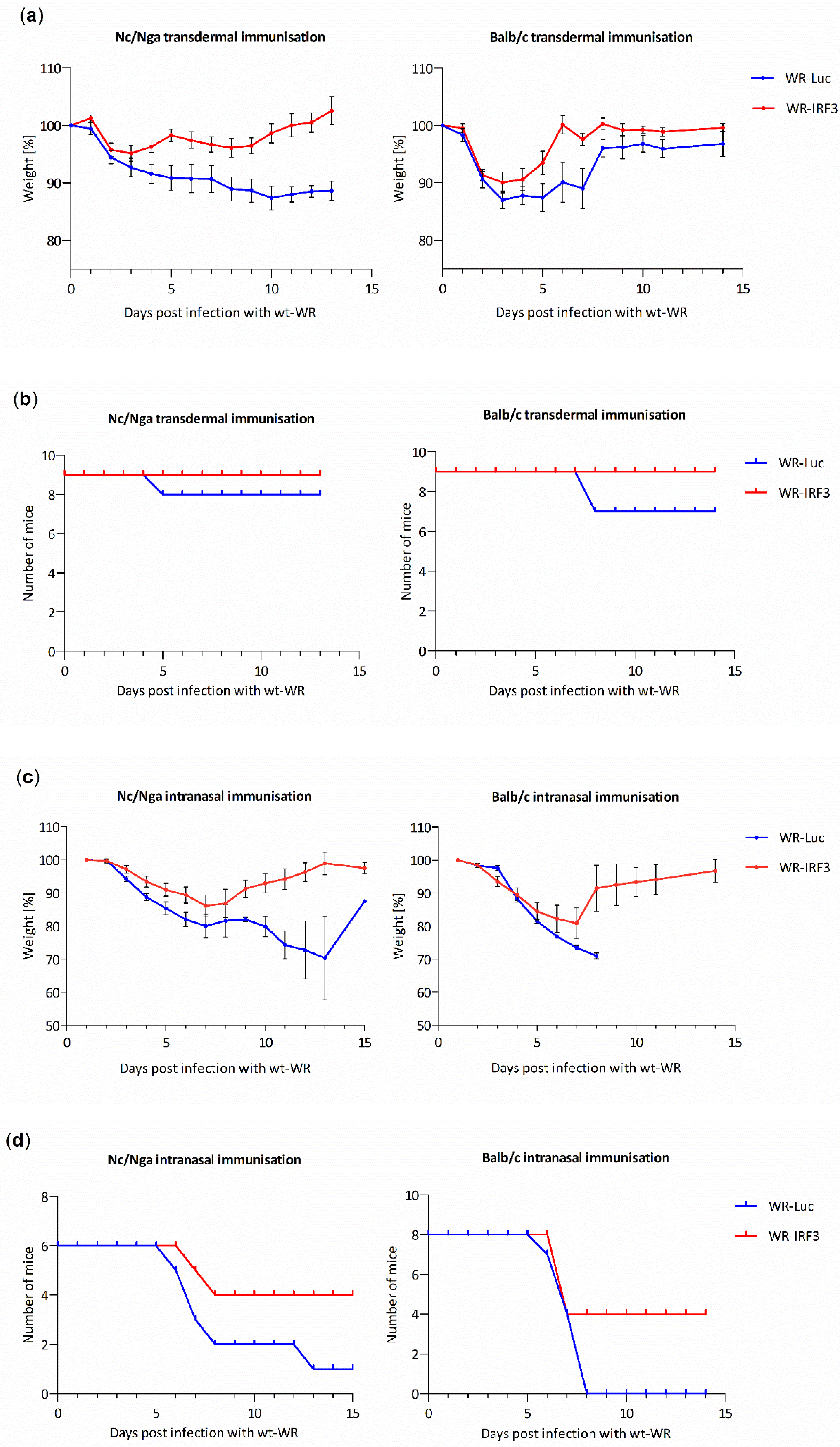
3.2.2. WR-IRF3 Induces Higher Protective Immunity against Lethal Poxvirus Challenge
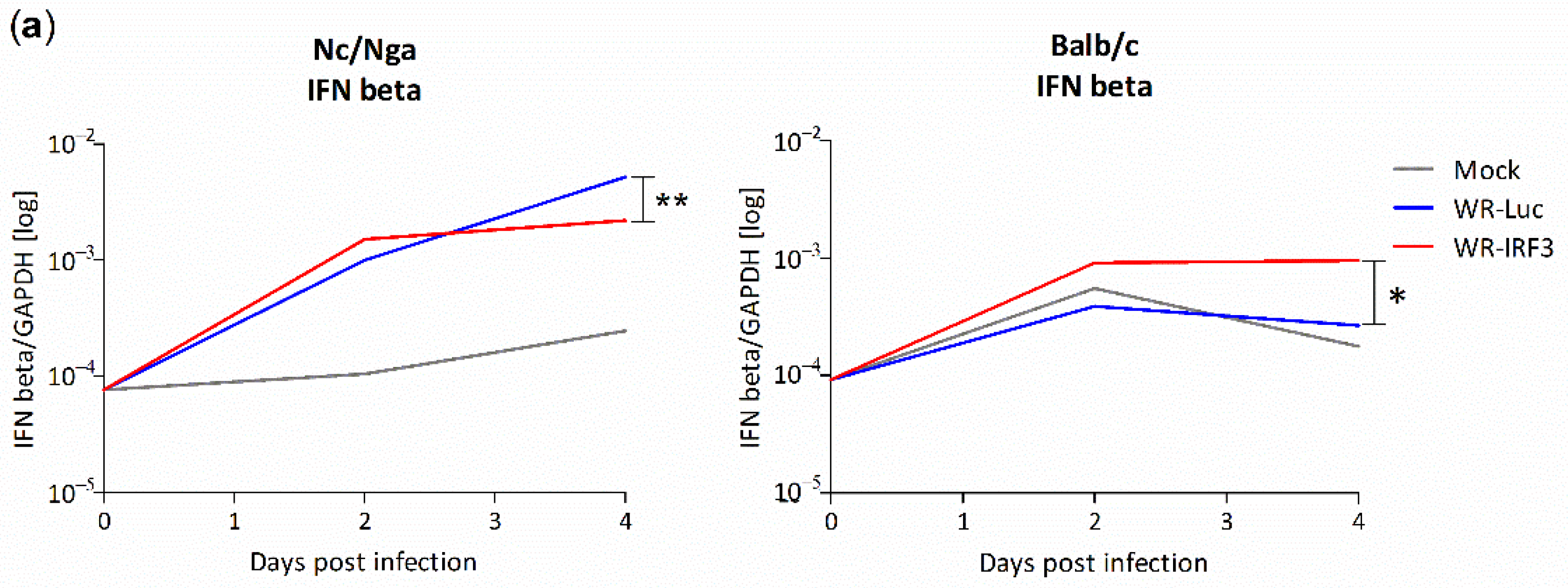
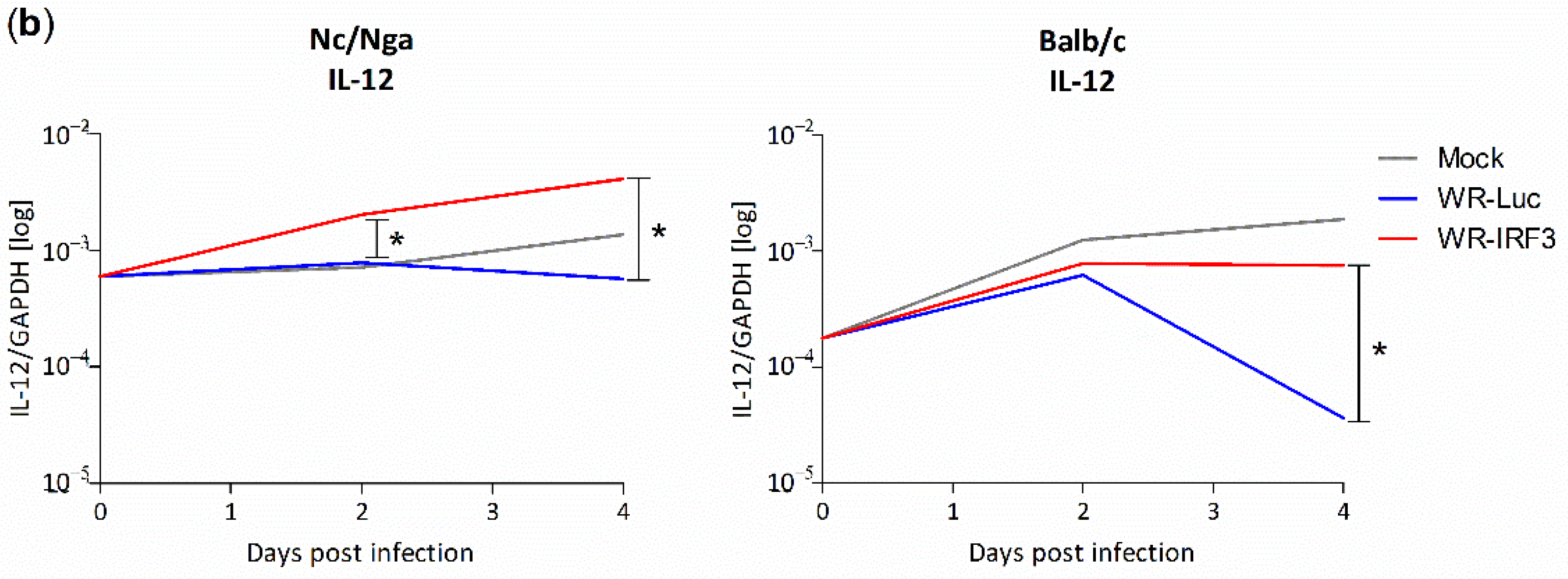
3.2.3. Changes in Interferon and Cytokine mRNA Levels in Mouse Skin Lesions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Artenstein, A.W. New generation smallpox vaccines: A review of preclinical and clinical data. Rev. Med. Virol. 2008, 18, 217–231. [Google Scholar] [CrossRef]
- Jacobs, B.L.; Langland, J.O.; Kibler, K.V.; Denzler, K.L.; White, S.D.; Holechek, S.A.; Wong, S.; Huynh, T.; Baskin, C.R. Vaccinia virus vaccines: Past, present and future. Antiviral Res. 2009, 84, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Engler, R.J.; Kenner, J.; Leung, D.Y. Smallpox vaccination: Risk considerations for patients with atopic dermatitis. J. Allergy Clin. Immunol. 2002, 110, 357–365. [Google Scholar] [CrossRef]
- Knitlova, J.; Hajkova, V.; Voska, L.; Elsterova, J.; Obrova, B.; Melkova, Z. Development of eczema vaccinatum in atopic mouse models and efficacy of MVA vaccination against lethal poxviral infection. PLoS ONE 2014, 9, e114374. [Google Scholar] [CrossRef] [Green Version]
- Werfel, T.; Allam, J.P.; Biedermann, T.; Eyerich, K.; Gilles, S.; Guttman-Yassky, E.; Hoetzenecker, W.; Knol, E.; Simon, H.U.; Wollenberg, A.; et al. Cellular and molecular immunologic mechanisms in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2016, 138, 336–349. [Google Scholar] [CrossRef] [Green Version]
- Mansouri, Y.; Guttman-Yassky, E. Immune Pathways in Atopic Dermatitis, and Definition of Biomarkers through Broad and Targeted Therapeutics. J. Clin. Med. 2015, 4, 858–873. [Google Scholar] [CrossRef] [Green Version]
- Wollenberg, A.; Wagner, M.; Gunther, S.; Towarowski, A.; Tuma, E.; Moderer, M.; Rothenfusser, S.; Wetzel, S.; Endres, S.; Hartmann, G. Plasmacytoid dendritic cells: A new cutaneous dendritic cell subset with distinct role in inflammatory skin diseases. J. Investig. Dermatol. 2002, 119, 1096–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, Y.; Tomimori, Y.; Yumoto, K.; Hasegawa, S.; Ando, T.; Tagaya, Y.; Crotty, S.; Kawakami, T. Inhibition of NK cell activity by IL-17 allows vaccinia virus to induce severe skin lesions in a mouse model of eczema vaccinatum. J. Exp. Med. 2009, 206, 1219–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, M.D.; Wollenberg, A.; Gallo, R.L.; Flaig, M.; Streib, J.E.; Wong, C.; Pavicic, T.; Boguniewicz, M.; Leung, D.Y. Cathelicidin deficiency predisposes to eczema herpeticum. J. Allergy Clin. Immunol. 2006, 117, 836–841. [Google Scholar] [CrossRef] [Green Version]
- Howell, M.D.; Gallo, R.L.; Boguniewicz, M.; Jones, J.F.; Wong, C.; Streib, J.E.; Leung, D.Y. Cytokine milieu of atopic dermatitis skin subverts the innate immune response to vaccinia virus. Immunity 2006, 24, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, L.S.; Goleva, E.; Hall, C.; Leung, D.Y. T regulatory cells in atopic dermatitis and subversion of their activity by superantigens. J. Allergy Clin. Immunol. 2004, 113, 756–763. [Google Scholar] [CrossRef]
- Kohara, Y.; Tanabe, K.; Matsuoka, K.; Kanda, N.; Matsuda, H.; Karasuyama, H.; Yonekawa, H. A major determinant quantitative-trait locus responsible for atopic dermatitis-like skin lesions in NC/Nga mice is located on Chromosome 9. Immunogenetics 2001, 53, 15–21. [Google Scholar] [CrossRef]
- Suto, H.; Matsuda, H.; Mitsuishi, K.; Hira, K.; Uchida, T.; Unno, T.; Ogawa, H.; Ra, C. NC/Nga mice: A mouse model for atopic dermatitis. Int. Arch. Allergy Immunol. 1999, 120 (Suppl. 1), 70–75. [Google Scholar] [CrossRef]
- Smith, G.L.; Benfield, C.T.O.; Maluquer de Motes, C.; Mazzon, M.; Ember, S.W.J.; Ferguson, B.J.; Sumner, R.P. Vaccinia virus immune evasion: Mechanisms, virulence and immunogenicity. J. Gen. Virol. 2013, 94, 2367–2392. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.L.; Talbot-Cooper, C.; Lu, Y. How Does Vaccinia Virus Interfere With Interferon? Adv. Virus Res. 2018, 100, 355–378. [Google Scholar] [CrossRef] [PubMed]
- Unterholzner, L.; Sumner, R.P.; Baran, M.; Ren, H.; Mansur, D.S.; Bourke, N.M.; Randow, F.; Smith, G.L.; Bowie, A.G. Vaccinia virus protein C6 is a virulence factor that binds TBK-1 adaptor proteins and inhibits activation of IRF3 and IRF7. PLoS Pathog. 2011, 7, e1002247. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.J.; Benfield, C.T.O.; Ren, H.; Lee, V.H.; Frazer, G.L.; Strnadova, P.; Sumner, R.P.; Smith, G.L. Vaccinia virus protein N2 is a nuclear IRF3 inhibitor that promotes virulence. J. Gen. Virol. 2013, 94, 2070–2081. [Google Scholar] [CrossRef] [PubMed]
- Liskova, J.; Knitlova, J.; Honner, R.; Melkova, Z. Apoptosis and necrosis in vaccinia virus-infected HeLa G and BSC-40 cells. Virus Res. 2011, 160, 40–50. [Google Scholar] [CrossRef]
- Benfield, C.T.O.; Ren, H.; Lucas, S.J.; Bahsoun, B.; Smith, G.L. Vaccinia virus protein K7 is a virulence factor that alters the acute immune response to infection. J. Gen. Virol. 2013, 94, 1647–1657. [Google Scholar] [CrossRef]
- Mansur, D.S.; Maluquer de Motes, C.; Unterholzner, L.; Sumner, R.P.; Ferguson, B.J.; Ren, H.; Strnadova, P.; Bowie, A.G.; Smith, G.L. Poxvirus targeting of E3 ligase beta-TrCP by molecular mimicry: A mechanism to inhibit NF-kappaB activation and promote immune evasion and virulence. PLoS Pathog. 2013, 9, e1003183. [Google Scholar] [CrossRef] [Green Version]
- Kalbacova, M.; Spisakova, M.; Liskova, J.; Melkova, Z. Lytic infection with vaccinia virus activates caspases in a Bcl-2-inhibitable manner. Virus Res. 2008, 135, 53–63. [Google Scholar] [CrossRef]
- Melkova, Z.; Esteban, M. Interferon-gamma severely inhibits DNA synthesis of vaccinia virus in a macrophage cell line. Virology 1994, 198, 731–735. [Google Scholar] [CrossRef]
- Kalbacova, M.; Vrbacky, M.; Humlova, Z.; Melkova, Z. Protooncogene Bcl-2 induces apoptosis in several cell lines. Folia Biol. (Praha) 2002, 48, 15–27. [Google Scholar] [PubMed]
- Rodriguez, J.F.; Rodriguez, D.; Rodriguez, J.R.; McGowan, E.B.; Esteban, M. Expression of the firefly luciferase gene in vaccinia virus: A highly sensitive gene marker to follow virus dissemination in tissues of infected animals. Proc. Natl. Acad. Sci. USA 1988, 85, 1667–1671. [Google Scholar] [CrossRef] [Green Version]
- Mackett, M.; Smith, G.L.; Moss, B. General method for production and selection of infectious vaccinia virus recombinants expressing foreign genes. J. Virol. 1984, 49, 857–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spisakova, M.; Cizek, Z.; Melkova, Z. Ethacrynic and alpha-lipoic acids inhibit vaccinia virus late gene expression. Antiviral Res. 2009, 81, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Joklik, W.K. The preparation and characteristics of highly purified radioactively labelled poxvirus. Biochim. Biophys Acta 1962, 61, 290–301. [Google Scholar] [CrossRef]
- Castellaneta, A.; Yoshida, O.; Kimura, S.; Yokota, S.; Geller, D.A.; Murase, N.; Thomson, A.W. Plasmacytoid dendritic cell-derived IFN-alpha promotes murine liver ischemia/reperfusion injury by induction of hepatocyte IRF-1. Hepatology 2014, 60, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Cogswell, J.P.; Godlevski, M.M.; Wisely, G.B.; Clay, W.C.; Leesnitzer, L.M.; Ways, J.P.; Gray, J.G. NF-kappa B regulates IL-1 beta transcription through a consensus NF-kappa B binding site and a nonconsensus CRE-like site. J. Immunol. 1994, 153, 712–723. [Google Scholar]
- Chattopadhyay, S.; Marques, J.T.; Yamashita, M.; Peters, K.L.; Smith, K.; Desai, A.; Williams, B.R.; Sen, G.C. Viral apoptosis is induced by IRF-3-mediated activation of Bax. EMBO J. 2010, 29, 1762–1773. [Google Scholar] [CrossRef] [Green Version]
- Gherardi, M.M.; Ramirez, J.C.; Rodriguez, D.; Rodriguez, J.R.; Sano, G.; Zavala, F.; Esteban, M. IL-12 delivery from recombinant vaccinia virus attenuates the vector and enhances the cellular immune response against HIV-1 Env in a dose-dependent manner. J. Immunol. 1999, 162, 6724–6733. [Google Scholar]
- von Sonnenburg, F.; Perona, P.; Darsow, U.; Ring, J.; von Krempelhuber, A.; Vollmar, J.; Roesch, S.; Baedeker, N.; Kollaritsch, H.; Chaplin, P. Safety and immunogenicity of modified vaccinia Ankara as a smallpox vaccine in people with atopic dermatitis. Vaccine 2014, 32, 5696–5702. [Google Scholar] [CrossRef]
- Greenberg, R.N.; Hurley, M.Y.; Dinh, D.V.; Mraz, S.; Vera, J.G.; von Bredow, D.; von Krempelhuber, A.; Roesch, S.; Virgin, G.; Arndtz-Wiedemann, N.; et al. A Multicenter, Open-Label, Controlled Phase II Study to Evaluate Safety and Immunogenicity of MVA Smallpox Vaccine (IMVAMUNE) in 18-40 Year Old Subjects with Diagnosed Atopic Dermatitis. PLoS ONE 2015, 10, e0138348. [Google Scholar] [CrossRef] [Green Version]
- Pittman, P.R.; Hahn, M.; Lee, H.S.; Koca, C.; Samy, N.; Schmidt, D.; Hornung, J.; Weidenthaler, H.; Heery, C.R.; Meyer, T.P.H.; et al. Phase 3 Efficacy Trial of Modified Vaccinia Ankara as a Vaccine against Smallpox. N. Engl. J. Med. 2019, 381, 1897–1908. [Google Scholar] [CrossRef]
- Yokote, H.; Shinmura, Y.; Kanehara, T.; Maruno, S.; Kuranaga, M.; Matsui, H.; Hashizume, S. Safety of attenuated smallpox vaccine LC16m8 in immunodeficient mice. Clin. Vaccine Immunol. 2014, 21, 1261–1266. [Google Scholar] [CrossRef] [Green Version]
- Danon, Y.L.; Sutter, G. Use of the LC16m8 Smallpox Vaccine in Immunocompromised Individuals Is Still Too Risky. Clin. Vaccine Immunol. 2015, 22, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.; Barber, G.N. STING signaling and host defense against microbial infection. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ni, G.; Ma, Z.; Damania, B. cGAS and STING: At the intersection of DNA and RNA virus-sensing networks. PLoS Pathog. 2018, 14, e1007148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glanz, A.; Chakravarty, S.; Varghese, M.; Kottapalli, A.; Fan, S.; Chakravarti, R.; Chattopadhyay, S. Transcriptional and Non-Transcriptional Activation, Posttranslational Modifications, and Antiviral Functions of Interferon Regulatory Factor 3 and Viral Antagonism by the SARS-Coronavirus. Viruses 2021, 13, 575. [Google Scholar] [CrossRef] [PubMed]
- Servant, M.J.; ten Oever, B.; LePage, C.; Conti, L.; Gessani, S.; Julkunen, I.; Lin, R.; Hiscott, J. Identification of distinct signaling pathways leading to the phosphorylation of interferon regulatory factor 3. J. Biol. Chem. 2001, 276, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.L.; Scott, D.E.; Bray, M. Eczema vaccinatum. Clin. Infect. Dis. 2012, 54, 832–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thanos, D.; Maniatis, T. Virus induction of human IFN beta gene expression requires the assembly of an enhanceosome. Cell 1995, 83, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Panne, D.; Maniatis, T.; Harrison, S.C. An atomic model of the interferon-beta enhanceosome. Cell 2007, 129, 1111–1123. [Google Scholar] [CrossRef] [Green Version]
- Garoufalis, E.; Kwan, I.; Lin, R.; Mustafa, A.; Pepin, N.; Roulston, A.; Lacoste, J.; Hiscott, J. Viral induction of the human beta interferon promoter: Modulation of transcription by NF-kappa B/rel proteins and interferon regulatory factors. J. Virol. 1994, 68, 4707–4715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, S.; Rottach, A.; Lotz-Havla, A.S.; Laux, V.; Muschaweckh, A.; Gersting, S.W.; Muntau, A.C.; Hopfner, K.P.; Jin, L.; Vanness, K.; et al. Rad50-CARD9 interactions link cytosolic DNA sensing to IL-1beta production. Nat. Immunol. 2014, 15, 538–545. [Google Scholar] [CrossRef] [Green Version]
- Aarreberg, L.D.; Wilkins, C.; Ramos, H.J.; Green, R.; Davis, M.A.; Chow, K.; Gale, M., Jr. Interleukin-1beta Signaling in Dendritic Cells Induces Antiviral Interferon Responses. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Barber, G.N. Cytosolic-DNA-mediated, STING-dependent proinflammatory gene induction necessitates canonical NF-kappaB activation through TBK1. J. Virol. 2014, 88, 5328–5341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yum, S.; Li, M.; Fang, Y.; Chen, Z.J. TBK1 recruitment to STING activates both IRF3 and NF-kappaB that mediate immune defense against tumors and viral infections. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Fang, R.; Wang, C.; Jiang, Q.; Lv, M.; Gao, P.; Yu, X.; Mu, P.; Zhang, R.; Bi, S.; Feng, J.M.; et al. NEMO-IKKbeta Are Essential for IRF3 and NF-kappaB Activation in the cGAS-STING Pathway. J. Immunol. 2017, 199, 3222–3233. [Google Scholar] [CrossRef] [PubMed]
- Guarda, G.; Braun, M.; Staehli, F.; Tardivel, A.; Mattmann, C.; Forster, I.; Farlik, M.; Decker, T.; Du Pasquier, R.A.; Romero, P.; et al. Type I interferon inhibits interleukin-1 production and inflammasome activation. Immunity 2011, 34, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Castiglia, V.; Piersigilli, A.; Ebner, F.; Janos, M.; Goldmann, O.; Dambock, U.; Kroger, A.; Weiss, S.; Knapp, S.; Jamieson, A.M.; et al. Type I Interferon Signaling Prevents IL-1beta-Driven Lethal Systemic Hyperinflammation during Invasive Bacterial Infection of Soft Tissue. Cell Host Microbe 2016, 19, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Tarassishin, L.; Loudig, O.; Bauman, A.; Shafit-Zagardo, B.; Suh, H.S.; Lee, S.C. Interferon regulatory factor 3 inhibits astrocyte inflammatory gene expression through suppression of the proinflammatory miR-155 and miR-155*. Glia 2011, 59, 1911–1922. [Google Scholar] [CrossRef] [Green Version]
- Kettle, S.; Alcami, A.; Khanna, A.; Ehret, R.; Jassoy, C.; Smith, G.L. Vaccinia virus serpin B13R (SPI-2) inhibits interleukin-1beta-converting enzyme and protects virus-infected cells from TNF- and Fas-mediated apoptosis, but does not prevent IL-1beta-induced fever. J. Gen. Virol. 1997, 78 Pt 3, 677–685. [Google Scholar] [CrossRef]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1beta secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Hornung, V.; Ablasser, A.; Charrel-Dennis, M.; Bauernfeind, F.; Horvath, G.; Caffrey, D.R.; Latz, E.; Fitzgerald, K.A. AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature 2009, 458, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Gerlic, M.; Faustin, B.; Postigo, A.; Yu, E.C.; Proell, M.; Gombosuren, N.; Krajewska, M.; Flynn, R.; Croft, M.; Way, M.; et al. Vaccinia virus F1L protein promotes virulence by inhibiting inflammasome activation. Proc. Natl. Acad. Sci. USA 2013, 110, 7808–7813. [Google Scholar] [CrossRef] [Green Version]
- Isaacs, A.; Lindenmann, J. Virus interference. I. The interferon. Proc. R Soc. Lond B Biol. Sci. 1957, 147, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Melkova, Z.; Yan, W.; Williams, B.R.; Hovanessian, A.G.; Esteban, M. The interferon-induced double-stranded RNA-activated human p68 protein kinase potently inhibits protein synthesis in cultured cells. Virology 1993, 192, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Rivas, C.; Gil, J.; Melkova, Z.; Esteban, M.; Diaz-Guerra, M. Vaccinia virus E3L protein is an inhibitor of the interferon (i.f.n.)-induced 2-5A synthetase enzyme. Virology 1998, 243, 406–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, J.R.; Rodriguez, D.; Esteban, M. Interferon treatment inhibits early events in vaccinia virus gene expression in infected mice. Virology 1991, 185, 929–933. [Google Scholar] [CrossRef]
- Liu, G.; Zhai, Q.; Schaffner, D.J.; Wu, A.; Yohannes, A.; Robinson, T.M.; Maland, M.; Wells, J.; Voss, T.G.; Bailey, C.; et al. Prevention of lethal respiratory vaccinia infections in mice with interferon-alpha and interferon-gamma. FEMS Immunol. Med. Microbiol. 2004, 40, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Ferguson, T.; Zitvogel, L.; Kroemer, G. Immunogenic and tolerogenic cell death. Nat. Rev. Immunol. 2009, 9, 353–363. [Google Scholar] [CrossRef]
- Kepp, O.; Senovilla, L.; Galluzzi, L.; Panaretakis, T.; Tesniere, A.; Schlemmer, F.; Madeo, F.; Zitvogel, L.; Kroemer, G. Viral subversion of immunogenic cell death. Cell Cycle 2009, 8, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Humlova, Z.; Vokurka, M.; Esteban, M.; Melkova, Z. Vaccinia virus induces apoptosis of infected macrophages. J. Gen. Virol. 2002, 83, 2821–2832. [Google Scholar] [CrossRef] [Green Version]
- Baixeras, E.; Cebrian, A.; Albar, J.P.; Salas, J.; Martinez, A.C.; Vinuela, E.; Revilla, Y. Vaccinia virus-induced apoptosis in immature B lymphocytes: Role of cellular Bcl-2. Virus Res. 1998, 58, 107–113. [Google Scholar] [CrossRef]
- Engelmayer, J.; Larsson, M.; Subklewe, M.; Chahroudi, A.; Cox, W.I.; Steinman, R.M.; Bhardwaj, N. Vaccinia virus inhibits the maturation of human dendritic cells: A novel mechanism of immune evasion. J. Immunol. 1999, 163, 6762–6768. [Google Scholar] [PubMed]
- Heylbroeck, C.; Balachandran, S.; Servant, M.J.; DeLuca, C.; Barber, G.N.; Lin, R.; Hiscott, J. The IRF-3 transcription factor mediates Sendai virus-induced apoptosis. J. Virol. 2000, 74, 3781–3792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apelbaum, A.; Yarden, G.; Warszawski, S.; Harari, D.; Schreiber, G. Type I interferons induce apoptosis by balancing cFLIP and caspase-8 independent of death ligands. Mol. Cell Biol. 2013, 33, 800–814. [Google Scholar] [CrossRef] [Green Version]
- Vestergaard, C.; Yoneyama, H.; Murai, M.; Nakamura, K.; Tamaki, K.; Terashima, Y.; Imai, T.; Yoshie, O.; Irimura, T.; Mizutani, H.; et al. Overproduction of Th2-specific chemokines in NC/Nga mice exhibiting atopic dermatitis-like lesions. J. Clin. Investig. 1999, 104, 1097–1105. [Google Scholar] [CrossRef]
- Komastu, T.; Ireland, D.D.; Reiss, C.S. IL-12 and viral infections. Cytokine Growth Factor Rev. 1998, 9, 277–285. [Google Scholar] [CrossRef]
- Fujikura, Y.; Kudlackova, P.; Vokurka, M.; Krijt, J.; Melkova, Z. The effect of nitric oxide on vaccinia virus-encoded ribonucleotide reductase. Nitric Oxide 2009, 20, 114–121. [Google Scholar] [CrossRef]
- Melkova, Z.; Esteban, M. Inhibition of vaccinia virus DNA replication by inducible expression of nitric oxide synthase. J. Immunol. 1995, 155, 5711–5718. [Google Scholar] [PubMed]
- Karupiah, G.; Xie, Q.W.; Buller, R.M.; Nathan, C.; Duarte, C.; MacMicking, J.D. Inhibition of viral replication by interferon-gamma-induced nitric oxide synthase. Science 1993, 261, 1445–1448. [Google Scholar] [CrossRef]
- Huang, S.; Hendriks, W.; Althage, A.; Hemmi, S.; Bluethmann, H.; Kamijo, R.; Vilcek, J.; Zinkernagel, R.M.; Aguet, M. Immune response in mice that lack the interferon-gamma receptor. Science 1993, 259, 1742–1745. [Google Scholar] [CrossRef]
- Ma, W.T.; Yao, X.T.; Peng, Q.; Chen, D.K. The protective and pathogenic roles of IL-17 in viral infections: Friend or foe? Open Biol. 2019, 9, 190109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, A.R.; Freyschmidt, E.J.; Burton, O.T.; Koleoglou, K.J.; Oyoshi, M.K.; Oettgen, H.C. IL-10 suppresses IL-17-mediated dermal inflammation and reduces the systemic burden of Vaccinia virus in a mouse model of eczema vaccinatum. Clin. Immunol. 2014, 150, 153–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patera, A.C.; Pesnicak, L.; Bertin, J.; Cohen, J.I. Interleukin 17 modulates the immune response to vaccinia virus infection. Virology 2002, 299, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Shiohara, T.; Hayakawa, J.; Mizukawa, Y. Animal models for atopic dermatitis: Are they relevant to human disease? J. Dermatol. Sci. 2004, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Achdout, H.; Lustig, S.; Israely, T.; Erez, N.; Politi, B.; Tamir, H.; Israeli, O.; Waner, T.; Melamed, S.; Paran, N. Induction, treatment and prevention of eczema vaccinatum in atopic dermatitis mouse models. Vaccine 2017, 35, 4245–4254. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.; Gao, P.S.; Grigoryev, D.N.; Rafaels, N.M.; Streib, J.E.; Howell, M.D.; Taylor, P.A.; Boguniewicz, M.; Canniff, J.; Armstrong, B.; et al. Human atopic dermatitis complicated by eczema herpeticum is associated with abnormalities in IFN-gamma response. J. Allergy Clin. Immunol. 2011, 127, 965–973.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward Primer 5′-3′ | Reverse Primer 5′-3′ |
|---|---|---|
| Mu GAPDH | CGG TGC TGA GTA TGT CGT GGA | GGC AGA AGG GGC GGA GAT GA |
| Mu IFNα 1 | GCA ACC CTC CTA GAC TCA TTC T | CCA GCA GGG CGT CTT CCT |
| Mu IFNβ | GCA CTG GGT GGA ATG AGA CT | AGT GGA GAG CAG TTG AGG ACA |
| Mu IFNγ | TGG CAT AGA TGT GGA AGA AAA GAG | TGC AGG ATT TTC ATG TCA CCA |
| Mu IL-1β | TCC ATT GAG GTG GAG AGC TT | GGA TGA GGA CAT GAG CAC CT |
| Mu IL-12 2 | TGT CAA TCA CGC TAC CTC CTC | GTC CAG AGA CTG GAA TGA CCC |
| Mu IL-17A | GGA CTC TCC ACC GCA ATG AA | TTT CCC TCC GCA TTG ACA CA |
| Mu IL-4 | CTT GGA AGC CCT ACA GAC GAG | GGA GAT GGA TGT GCC AAA CG |
| Mu IL-10 | AGG CGC TGT CAT CGA TTT CTC | GCC TTG TAG ACA CCT TGG TCT T |
| Mu TGFβ1 | CAC TGA TAC GCC TGA GTG GC | TCG AAA GCC CTG TAT TCC GT |
| Mu TGFβ3 | GCA AGA ATC TGC CCA CAA GG | CCA TTG GGC TGA AAG GTG TG |
| Mu TNFα | GAT CGG TCC CCA AAG GGA TG | TGA GGG TCT GGG CCA TAG AA |
| Mu AIM2 | AGG CAG TGG GAA CAA GAC AG | GAA ACC TTC CTG ACG CCA CC |
| Mu iNOS 3 | ACG AGA CGG ATA GGC AGA GA | GCA CAT GCA AGG AAG GGA AC |
| Mu IRF-3 | TAG GCT GGC TGT TGG AGA TGT | CCA GGT CTT CCA GCA GAC ACT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pilna, H.; Hajkova, V.; Knitlova, J.; Liskova, J.; Elsterova, J.; Melkova, Z. Vaccinia Virus Expressing Interferon Regulatory Factor 3 Induces Higher Protective Immune Responses against Lethal Poxvirus Challenge in Atopic Organism. Viruses 2021, 13, 1986. https://doi.org/10.3390/v13101986
Pilna H, Hajkova V, Knitlova J, Liskova J, Elsterova J, Melkova Z. Vaccinia Virus Expressing Interferon Regulatory Factor 3 Induces Higher Protective Immune Responses against Lethal Poxvirus Challenge in Atopic Organism. Viruses. 2021; 13(10):1986. https://doi.org/10.3390/v13101986
Chicago/Turabian StylePilna, Hana, Vera Hajkova, Jarmila Knitlova, Jana Liskova, Jana Elsterova, and Zora Melkova. 2021. "Vaccinia Virus Expressing Interferon Regulatory Factor 3 Induces Higher Protective Immune Responses against Lethal Poxvirus Challenge in Atopic Organism" Viruses 13, no. 10: 1986. https://doi.org/10.3390/v13101986