
Culture Isolate of Rickettsia felis from a Tick


1
Department of Rickettsiology, Biomedical Research Center, Slovak Academy of Sciences, Dúbravská cesta 9, 845 05 Bratislava, Slovakia
2
Laboratory of Molecular Structure Characterization, Institute of Microbiology, Czech Academy of Sciences, Vídeňská 1038, 142 20 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Int. J. Environ. Res. Public Health 2022, 19(7), 4321; https://doi.org/10.3390/ijerph19074321
Submission received: 28 January 2022
/
Revised: 28 March 2022
/
Accepted: 31 March 2022
/
Published: 4 April 2022
(This article belongs to the Special Issue Ticks and Tick-Borne Diseases: Persistent and Emerging Threats to Human Health)
Abstract
:Although the cat flea, Ctenocephalides felis, has been identified as the primary vector of Rickettsia felis, additional flea, tick, mite, and louse species have also been associated with this bacterium by molecular means; however, the role of these arthropods in the transmission of R. felis has not been clarified. Here, we succeeded in culture isolation of R. felis from a host-seeking castor bean tick, Ixodes ricinus, the most common tick in Slovakia. The bacterial isolation was performed on XTC-2 cells at 28 °C using the shell-vial technique. An evaluation of the growth properties was performed for both the XTC-2 and Vero cell lines. We observed R. felis in the infected host cells microscopically by Gimenez staining and immunofluorescence assay. The R. felis isolate was purified by gradient ultracentrifugation and visualized by electron microscopy. Fragments of the genes gltA, ompA, ompB, htrA, rpoB, sca4, rffE, and rrs were amplified and compared with the corresponding sequences of the type strain URRWXCal2 and other R. felis culture -isolated strains. We did not detect any nucleotide polymorphisms; however, plasmid pRFδ, characteristic of the standard strain, was absent in our isolate. Herein, we describe the first successful isolation and characterization of a tick-derived R. felis strain “Danube”, obtained from an I. ricinus nymph.
1. Introduction
The obligate intracellular bacterium, Rickettsia felis, is a vector-borne pathogen and the causative agent of flea-borne spotted fever [1]. Similar to other rickettsioses (e.g., murine typhus, rickettsialpox, and Mediterranean spotted fever), typical symptoms include fever, headache, maculopapular rash, fatigue, myalgia, arthralgia, elevation of liver enzymes, and eschar at the bite site [2,3]. The disease is considered clinically mild, but in older patients and in individuals with delayed diagnosis, it may cause severe illness with gastrointestinal and neurologic signs [4,5,6], including some cases of meningoencephalitis being fatal [7].
The distribution of R. felis appears to be cosmopolitan, as the organism has been detected in various countries worldwide, with an increasing number of human case reports [8,9,10]. In Eurasia, R. felis was detected for the first time in southern Spain in the cat flea Ctenocephalides felis, the biological vector and reservoir of this bacterium [11]. Although R. felis is primarily identified in the cat flea, it has also been associated with other arthropods [12,13]. By molecular techniques, R. felis has been detected in different flea species, mosquitoes, mites, and ticks (both Ixodid and Argasid). These tick species include Rhipicephalus sanguineus in Brazil, Spain, Chile, China, and the Philippines [14,15,16,17,18,19,20,21]; Rhipicephalus bursa in Turkey [22]; Rhipicephalus turanicus in Italy [23]; Haemaphysalis flava, Haemaphysalis kitasatoe, and Ixodes ovatus in Japan [24]; Haemaphysalis sulcata in Croatia [25]; Haemaphysalis leporispalustris in the United States [26]; Haemaphysalis sp. and Rhipicephalus microplus in Malaysia [27]; Heamaphysalis bancrofti in Australia [28]; Amblyomma maculatum in the United States [29]; Amblyomma cajennense, Amblyomma humerale, Amblyomma ovale, and Amblyomma sculptum in Brazil [14,30,31,32]; Ixodes granulatus in Taiwan [33]; Ixodes hexagonus in Italy [34]; Ixodes ricinus in Germany, France, Spain, Romania, and Serbia [35,36,37,38,39,40]; Dermacentor nitens in Brazil and Cuba [21,41]; and Dermacentor variabilis and Carios capensis in the United States [42,43] (Table S1). The previous reports of the presence of R. felis in hard and soft ticks are not necessarily related to a vectorial role, and to date, the role of ticks in the ecology and transmission of R. felis has not been clarified. Its presence in ticks could be the result of an ingested blood meal from a bacteremic vertebrate host and a subsequent vertical transmission or from an infected flea via cofeeding transmission [12]. In Slovakia, the presence of R. felis was recently confirmed in Ctenophthalmus solutus fleas collected from Apodemus agrarius [44,45].
After the initial discovery of R. felis (historically known as the “ELB agent”) in cat flea tissues by electron microscopy [46], several attempts were made to establish a sustained bacterial culture, but that constituted a challenging task [47,48,49,50]. Finally, the propagation of R. felis, isolated from commercial fleas maintained by Flea Data, Inc. (Freeville, New York, NY, USA), was successful [51]. This isolate, named Marseille-URRWXCal2, is the reference strain of R. felis [52].
Seven R. felis culture isolates have been reported up to now (Table 1). The growth of R. felis in vitro has been accomplished using several different cell lines: amphibian cells (XTC-2) [51,53,54], tick cells (ISE6) [55,56], mosquito cells (Aa23, Sua5B, C6/36) [57,58,59], fly cells (S2) [60], and mammalian cells (Vero) with special medium supplementation [51,52,61]. A common characteristic of the effective propagation of R. felis is incubation at lower temperatures, i.e., ≤32 °C [52,60,62]. All culture isolates, except the most recent one from Australia and the unique LSU-Lb strain, were obtained from commercial/laboratory colony cat fleas or cat fleas collected from dogs. In addition, Stevenson and colleagues isolated R. felis from Anomiopsyllus nudata fleas (collected from Neotoma albigula in the United States), but unfortunately, the authors could not establish a sustained culture of rickettsiae [63].
In the current study, we describe the successful isolation and cultivation of R. felis from a questing I. ricinus nymph by the shell-vial technique. The isolate named “Danube” was established in the amphibian XTC-2 cells line, and the partial characterization of its biological properties by molecular biology techniques and microscopy are discussed here.
2. Materials and Methods
2.1. Collection of Ticks
For culture isolation, host-seeking I. ricinus nymphs were collected in the borough of Podunajské Biskupice (48.07° N, 17.12° E, altitude about 130 m), in the south-eastern part of the Bratislava region (Figure 1). The area lies in the geographic zone of the Danube lowland, a grassland with edge habitats forested by Quercus, Carpinus, Populus, and Salix, and characterized by a moderate climate. The collection area is influenced by anthropic rural environmental activities (hunting and farming). The Bratislava region is also characterized by a high level of biodiversity of wild vertebrate species. All ticks were collected by blanket dragging from vegetation, morphologically identified according to a standard taxonomic key [64], and stored at −80°C until further investigation. No specific permissions were required for questing tick collection in this location as the region is not a protected area. The field study did not involve any endangered or protected species.
2.2. Isolation of Rickettsia from Ticks
Isolation of rickettsiae was performed as previously described [65] with minor modifications. Briefly, nymphs were thawed, sterilized by immersion in iodinated alcohol for 10 min, rinsed with sterile water for 10 min, and dried on sterile filter paper under a laminar flow hood. Nymphs were longitudinally cut, one half of the tick was triturated in 1 mL of Leibovitz’s medium (L-15; Lonza, Basel, Switzerland), and the mixture was placed into a shell-vial containing a monolayer of amphibian epithelial XTC-2 cells [66] (the cell line was obtained from the laboratories of the Reference Center for Rickettsioses, Faculty of Medicine, Marseille, France). All ticks were processed individually. The shell-vials were centrifuged at 700× g for 20 min, and the supernatant was replaced with 1 mL of supplemented Leibovitz’s medium containing 2.5% fetal bovine serum (FBS; HyClone, Logan, UT, USA), 8% tryptose phosphate broth (TPB; Sigma-Aldrich, St. Louis, MO, USA), and 2 mM L-glutamine (Lonza). After 7 days of incubation at 28 °C, scraped XTC-2 cells were applied to a microscope slide and stained by the method of Gimenez [67] to monitor the infection status of host cells by rickettsiae. Supernatant with detached cells from the infected shell-vials was transferred into cell culture flasks with a growth area of 25 cm2 (Corning Inc., Corning, NY, USA). Fresh supplemented Leibovitz’s medium was added, and the flasks were incubated at 28 °C in the absence of CO2.
2.3. Cultivation and Purification of Isolated R. felis
The bacteria were sub-cultured without trypsinization in the XTC-2 cell line with supplemented Leibovitz’s medium containing 2.5% FBS, 8% TPB, and 2 mM L-glutamine at 28 °C, and subsequently in the Vero cell line (ATCC CCL-81) with supplemented RPMI 1640 medium containing 25 mM HEPES, 2 mM L-glutamine, and 2.5% FBS at 32 °C and 34 °C in a 5% CO2 atmosphere. Host cells were examined daily for cytopathic effect formation due to rickettsial infection, and the medium was replaced weekly to ensure adequate nutrition. When cells showed infection with apparent cytopathic effect (≥90%), aliquots were used to establish new infections in XTC-2 cell monolayers or were frozen at −80 °C, and stored for future reference. The presence of R. felis in the cell culture was verified by Gimenez staining using an Eclipse Ni-U microscope (Nikon, Tokyo, Japan), and photographs of stained slides were taken using a Coolpix P330 camera (Nikon, Tokyo, Japan). Only contrast was adjusted on the images.
The propagated isolate of R. felis in the XTC-2 cell line was purified by a modified isopycnic density gradient centrifugation protocol [68,69]. Briefly, infected cells were harvested from flasks and pelleted by centrifugation at 1000× g for 5 min at 4 °C. The cell culture medium was removed, and cold K36 buffer (0.1 M KCl, 0.015 M NaCl, 0.05 M potassium phosphate buffer, pH 7.0) was added to each sample. Host cell lysis was achieved by sonication, and rickettsiae were separated from host cell debris by centrifugation at 1000× g for 5 min at 4 °C (Hettich, Tuttlingen, Germany). Supernatants containing bacteria were transferred to new tubes and centrifuged at 22,000× g for 30 min at 4 °C. Pelleted rickettsiae were resuspended in SPG buffer (0.218 M sucrose, 3.76 mM KH2PO4, 7.1 mM K2HPO4, and 4.9 mM potassium glutamate) and overlaid onto a discontinuous Renografin gradient (Ultravist 370, Bayer, Leverkusen, Germany) made with three concentrations: 42%, 36%, and 30% solution. Sample tubes were then centrifuged at 120,000× g for 90 min at 4 °C. After ultracentrifugation (Figure S1), viable rickettsiae were collected into clean tubes and centrifuged at 22,000× g for 30 min at 4 °C. The bacterial pellet was resuspended in cold SPG buffer and stored at −80 °C for further experiments.
2.4. Monitoring of Rickettsial Intracellular Growth
The cell density of viable XTC-2 cells was enumerated using trypan blue staining in a Bürker Counting Chamber, and afterward, cells were seeded into 12-well tissue culture plates (Greiner Bio-One, Kremsmünster, Austria). The monolayers of host cells were infected with R. felis at a multiplicity of infection (MOI) equal to 10. The infection was initiated from the frozen stocks of purified rickettsiae stored in SPG buffer. The concentration of bacteria was determined as described previously [70]. On each day post-infection (dpi) up to one week, infected XTC-2 cells were scraped into culture media and centrifugated at 16,000× g for 15 min at 4 °C. The supernatants were discarded, and the pelleted samples were stored at −80 °C until further nucleic acid extraction. Two separate biological replicates were carried out for each collection time point.
2.5. RNA Extraction and Reverse-Transcriptase Quantitative PCR (RT-qPCR)
Total RNA from infected XTC-2 cells was extracted using the RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Extracted RNA was treated with RNase-Free DNase Set (Qiagen) and purified using the RNeasy MinElute Cleanup Kit (Qiagen). The quality and quantity of the extracted RNA samples were evaluated with a nanophotometer (Implen, Westlake Village, CA, USA), and the subsequent reverse transcription was performed on 1 µg of isolated RNA using the First Strand cDNA Synthesis Kit with random hexamer primers (Thermo Scientific, Waltham, MA, USA). No-reverse transcriptase controls (reverse transcription without the enzyme) and negative controls (ultrapure water instead of a template) were included for cDNA synthesis and RT-qPCR. All samples were stored at −20 °C until further processing.
To determine rickettsial transcript copy numbers of the rpsL gene (encoding ribosomal S12 protein), we performed RT-qPCR as published previously [70]. The PCR reactions were carried out in a CFX 96 Real-Time system C 1000 Thermal Cycler (Bio-Rad, Hercules, CA, USA). The components of the PCR mixture included Maxima Probe/ROX qPCR Master Mix (Thermo Scientific, Waltham, MA, USA), 0.4 µM of each primer, 0.4 µM of TaqMan probe, ultrapure nuclease-free water, and 12.5 ng of cDNA template, or serial dilution of standards. The amplification conditions were as follows: an initial denaturation step at 95 °C for 10 min, followed by 40 cycles of denaturation at 95 °C for 20 s, annealing, and elongation at 58 °C for 40 s, with fluorescence acquisition in single mode. Each sample was tested in technical triplicate. To generate a standard curve, we utilized a 10-fold serial dilution of the amplified and purified PCR product of the gene rpsL from Rickettsia akari strain MK (Kaplan, ATCC VR-148) [71].
2.6. DNA Extraction
Total genomic DNA was extracted from the remaining half-ticks and rickettsial culture samples using the QIAamp DNA Mini kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Plasmid DNA from purified R. felis was extracted with QIAprep Spin Miniprep Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. Extracted DNA samples were eluted in ultrapure nuclease-free water (Thermo Scientific, Waltham, MA, USA). The quality and quantity of DNA were evaluated using a nanophotometer (Implen, Westlake Village, CA, USA), and the samples were stored at −20 °C until further use. To avoid contamination, DNA extraction, PCR setup, and agarose gel analysis were carried out in separate rooms.
2.7. Molecular Identification by PCR and Sequencing
The samples from the nymphs, infected cell lines, and purified bacteria were tested by conventional PCR carried out in a GenePro Thermal Cycler (Bioer Technology, Hangzhou, China) and/or Labcycler (Sensoquest, Göttingen, Germany). Primer sets amplifying fragments of the rickettsial genes rrs [72,73], sca4 [74], htrA [24], gltA [51,75,76], ompA [55,77], ompB [78], rpoB [52,79], and rffE [71] are listed in Table S2. The PCR mixture included FIREPol Master Mix Ready to Load (Solis BioDyne), 0.25 µM of each primer, ultrapure nuclease-free water, and 100 ng of extracted DNA template. The thermocycling parameters were those published previously (Table S2). The culture was also checked for any R. helvetica contamination using species-specific primers (rhelv.26f and rhelv.356r) targeting the 23S rRNA coding gene [80]. Positive controls (genomic DNA from Rickettsia slovaca strain B-13, ATCC VR-1639 and/or Rickettsia helvetica strain C9P9, ATCC VR-1375), and no template controls were included in all PCRs.
To amplify a portion of the R. felis pRF and pRFδ plasmid sequences, we used oligonucleotides published previously [81]. The PCR reaction was composed of Platinum Taq DNA High Fidelity Polymerase (1 U per reaction; Invitrogen, Waltham, MA, USA), High Fidelity PCR Buffer, 2.0 mM MgSO4, 0.2 mM of each dNTP, 0.4 µM of each primer, ultrapure nuclease-free water, and 2 µL of template DNA.
The amplified PCR products were separated by electrophoresis in 1% agarose gels and visualized with SYBR Safe DNA gel stain (Invitrogen, Waltham, MA, USA) using a Benchtop 3UV Transilluminator (UVP, Upland, CA, USA). The DNA fragments were cleaned up with ExoSAP-IT (Thermo Scientific, Waltham, MA, USA) and sequenced at least twice, in both forward and reverse directions (Microsynth Seqlab, Göttingen, Germany). The obtained sequences were aligned with BioEdit Sequence Alignment Editor software [82] and compared with those of Rickettsia spp. available in GenBank by means of the Basic Local Alignment Search Tool (http://www.ncbi.nlm.nih.gov/BLAST/, 24 March 2022).
2.8. Immunofluorescence Assay (IFA)
Immunofluorescence detection of R. felis in the infected host cells was performed as previously described [83]. Confluent XTC-2 and Vero cells were infected with an MOI 25. After 72 h, host cells were fixed with 4% paraformaldehyde in cytoskeleton-stabilizing PHEM buffer [84], permeabilized with 0.1% Triton X-100 in PBS, and blocked with 5% milk in PBS. Cells were then incubated with a rabbit polyclonal primary antibody against Rickettsia conorii (1:100; prepared in our experimental animal facility at the Biomedical Research Center, Slovak Academy of Sciences, Bratislava, Slovakia). For the localization of intracellular bacteria, we applied a goat anti-rabbit IgG (H + L) polyclonal secondary antibody conjugated with Rhodamine Red-X (Invitrogen, reference number R6394, lot 1402199, made in the USA). To visualize filamentous actin, coverslips were probed with Alexa Fluor 488-tagged phalloidin (Invitrogen, reference number A12379, lot 1378369, made in the United States). Finally, samples were mounted with Vectashield Medium (Vector Laboratories, Burlingame, CA, USA) containing DAPI and analyzed using a Zeiss LSM 510 META confocal fluorescent microscope. Brightness and contrast were adjusted by LSM Image browser software (Zeiss, Jena, Germany).
2.9. Electron Microscopy
For negative staining, 5 μL of the sample was placed onto glow-discharge activated [85] carbon/formvar grids. After 30 s of adsorption, the grids were negatively stained with 1% ammonium molybdate and 0.1% trehalose for 30 s. The grids were air-dried and examined in an FEI/Philips CM100 electron microscope (FEI).
For scanning electron microscopy, the purified samples of R. felis isolate were fixed with 3% glutaraldehyde in SP buffer (3.76 mM KH2PO4, 7.1 mM K2HPO4, 0.218 M sucrose) overnight at 4 °C. The washed cells were then allowed to sediment overnight onto poly-l-lysine-treated circular coverslips or silicon wafers at 4 °C. The coverslips and the wafers with attached bacteria were dehydrated through an alcohol series and critical point dried from liquid CO2 in a K850 Critical Point Dryer (Quorum Technologies, Lewes, United Kingdom). The dried samples were sputter-coated with 3 nm of platinum in a Q150T Turbo-Pumped Sputter Coater (Quorum Technologies, Lewes, United Kingdom). The final samples were examined in an FEI Nova NanoSEM scanning electron microscope (FEI) at 5 kV using CBS and TLD detectors.
2.10. Statistical Analysis
For evaluation of rickettsial growth in host cells, one-way analysis of variance (ANOVA) was performed, followed by Dunnett’s multiple comparison post-hoc test when an overall significance (p-value of <0.05) was found. Analyses were performed using Prism 9 software (GraphPad Software, San Diego, CA, USA).
3. Results and Discussion
3.1. Initial Isolation of R. felis from an I. ricinus Nymph
Sixty unfed I. ricinus nymphs, collected from vegetation by flagging, were processed in the experiment. After surface sterilization, nymphs were longitudinally cut in half, and one half was used for culture isolation and the second half for an initial molecular screening. The presence of rickettsiae was confirmed in 20% of samples by conventional PCR, using the primers CS877p and CS1258n. In parallel, we focused on a positive culture isolate that we recovered from a tested I. ricinus nymph. By Gimenez staining, we detected intracellular, rod-shaped bacteria in the host cells cultured in the shell-vial 7 days after inoculation. Following an apparent cytopathic effect in XTC-2 cells three weeks after inoculation, we harvested and sub-cultured the infected cells into a confluent monolayer of XTC-2 cells, first in 25 cm2 and later in 75 cm2 tissue culture flasks to establish a sustained bacterial culture. In order to identify the species of the isolated rickettsiae, we tested samples from infected XTC-2 cells by PCR using genus-specific primers, amplifying the fragments of the gltA and rffE genes, coding the citrate synthase and UDP-N-acetylglucosamine 2-epimerase, respectively (Figure 2). Next, the amplification products were sequenced (#ON053296, #ON053294), and the isolated rickettsiae were confirmed as R. felis, with 100% identity to the previously reported gene fragment sequences of the type strain URRWXCal2 and significant nucleotide differences from other rickettsiae. The isolate was named the “Danube” strain (for the tick collection site in the Danube lowland).
The use of the XTC-2 cell line has proven effective in the isolation of other R. felis strains, including the type strain and the culture isolates reported from Australia (Table 1). Our laboratory successfully employed the shell-vial technique previously to isolate other intracellular microorganisms from arthropods, for instance, Rickettsia monacensis IRS3 [86], R. slovaca [87], Diplorickettsia massiliensis [88], R. helvetica [65], Arsenophonus nasoniae [89], and Rickettsia raoultii [90].
For further studies, we purified the rickettsiae collected from the infected XTC-2 cells by renografin density gradient centrifugation (Figure S1). As R. helvetica is the most prevalent rickettsia in I. ricinus ticks in Slovakia [91,92,93,94], we performed species-specific amplification of the 23S rRNA gene to confirm the absence of R. helvetica in the cultured R. felis isolate Danube (Figure S2). Rickettsia helvetica could not be detected in the purified sample of R. felis by PCR, whereas the corresponding primer pair amplified the positive control DNA.
3.2. Propagation of Rickettsiae in XTC-2 and Vero Cell Lines
The R. felis isolate adapted well to propagation in an XTC-2 cell-containing culture flask after sub-culturing from the shell-vial. We observed an evident cytopathic effect in infected host cells (similar to the standard strain URRWXCal2 [52]), and the bacterial proliferation was also confirmed by Gimenez staining (Figure 3). Next, we attempted to infect the mammalian Vero cells with our R. felis isolate at 32 °C and 34 °C. Rickettsia felis Danube was able to infect Vero cells, inducing round cytopathic foci at 32 °C, similar to those of the infected XTC-2 cell monolayers (Figure 3). Comparably to the reference strain [51], R. felis Danube replicated better in XTC-2 cells than in the mammalian cell line. In Vero cells maintained at 34 °C, the number of propagating bacteria gradually decreased. A similar observation was reported during the early cultivation of the “ELB” agent in Vero cells at 34 °C [47].
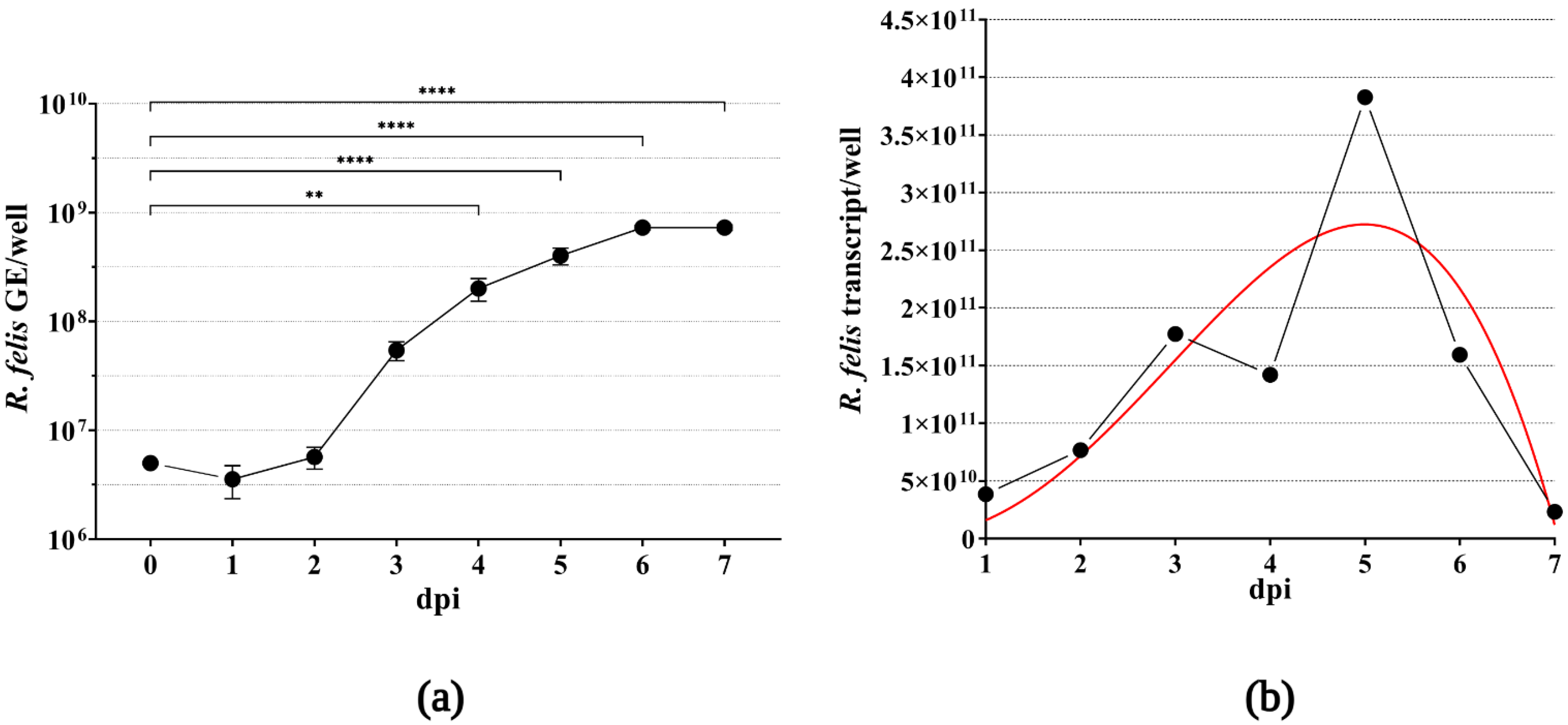
To extend the knowledge regarding the growth characteristics of R. felis Danube in XTC-2 cells, we propagated the bacteria in 12-well plates to estimate the multiplication rate. Samples collected every 24 h after infection were subjected to qPCR, and the mean number of rickettsial genome equivalents was calculated for each well, based on the amplification of the rickettsial single-copy, house-keeping gene rpsL compared to the standard calibration curve. As displayed in Figure 4a, R. felis proliferated rapidly in host cells, with a statistically significant increase in the mean value of the bacterial genome equivalents over the time course of our experiment. The growth curve showed an initial lag phase (1–2 dpi), followed by an exponential increase in R. felis genome copy numbers (3–6 dpi). We observed the highest multiplication rate at 6 dpi, followed by a stationary phase.
To demonstrate the viability of rickettsiae in the infected XTC-2 cells, we extracted RNA from the parallel cultivation wells and quantified the synthesized complementary DNA by qPCR, in the same manner as the extracted DNA samples. Similar to the rickettsial growth kinetics based on the estimated numbers of genomic equivalents, we observed an increase in the mean transcript numbers of the R. felis rpsL gene (Figure 4b). Following the highest transcript number of the ribosomal gene at 5 dpi, we observed a decrease in rickettsial cDNA quantity, which corresponds to the heavy host cell detachment due to bacterial burden observed by light microscopy (data not shown).
3.3. Molecular Characterization of the R. felis Isolate Danube
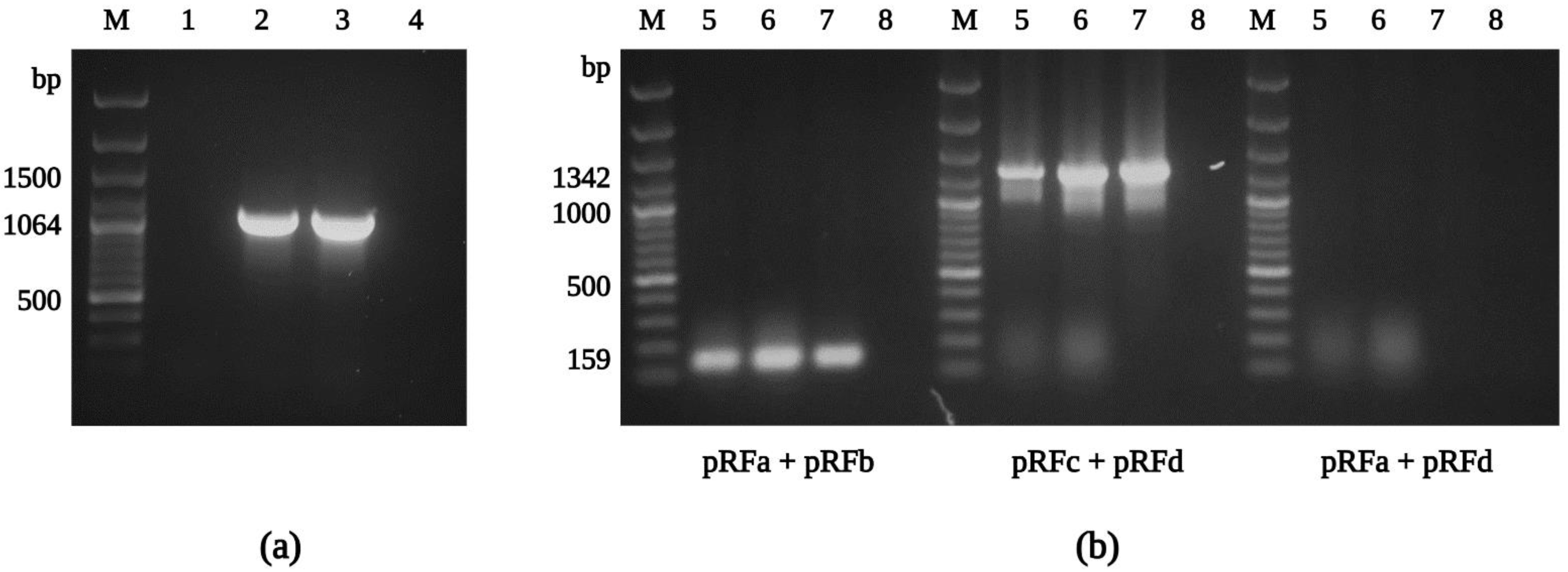
In addition to confirming the species identity and assessing the nucleotide variability of conserved rickettsial genes in R. felis Danube, fragments of the ompA and ompB (outer membrane proteins A and B), sca4 (surface cell antigen 4), gltA, htrA (17 kDa surface antigen), rpoB (RNA polymerase subunit β), and rrs (16S rRNA) genes were amplified by conventional PCR, including R. felis-specific primer sets [55,77] (Figure 5a). The subsequent sequence analysis confirmed the isolate Danube as R. felis, with 99–100% similarity to those sequences obtained from other culture-isolated strains of R. felis (Table S3). Partial sequences of R. felis Danube generated in this study were deposited into GenBank and assigned accession numbers ON053297, ON053298 (gltA), ON053299, ON053306 (ompA), ON053301-ON053303 (ompB), ON053304 (sca4), ON053294 (htrA), ON053305 (rpoB), and ON053300 (rrs).
The complete genome of the R. felis strain URRWXCal2 was previously described, and unique characteristics, including the presence of two conjugative plasmids, pRF and pRFδ, were revealed [95]. Recently, additional strains of R. felis, LSU and LSU-Lb, were subjected to genome and plasmid content analysis [96]. In our study, we obtained positive results for the presence of the pRF plasmid using the primer sets pRFa and pRFb, and pRFc and pRFd (Figure 5b), and the sequences amplified in the current study (#ON053307) were identical to those reported for R. felis previously. However, we could not detect the pRFδ plasmid sequence by PCR (primers pRFa and pRFd) in any of our R. felis Danube samples. Similar findings were described for the R. felis strains LSU and LSU-Lb [55,56], and the R. felis genotype RF2125 endemic in Archaeopsylla erinacei fleas from Algeria [97], confirming plasmid content variation between different R. felis strains [81].
3.4. Microscopy
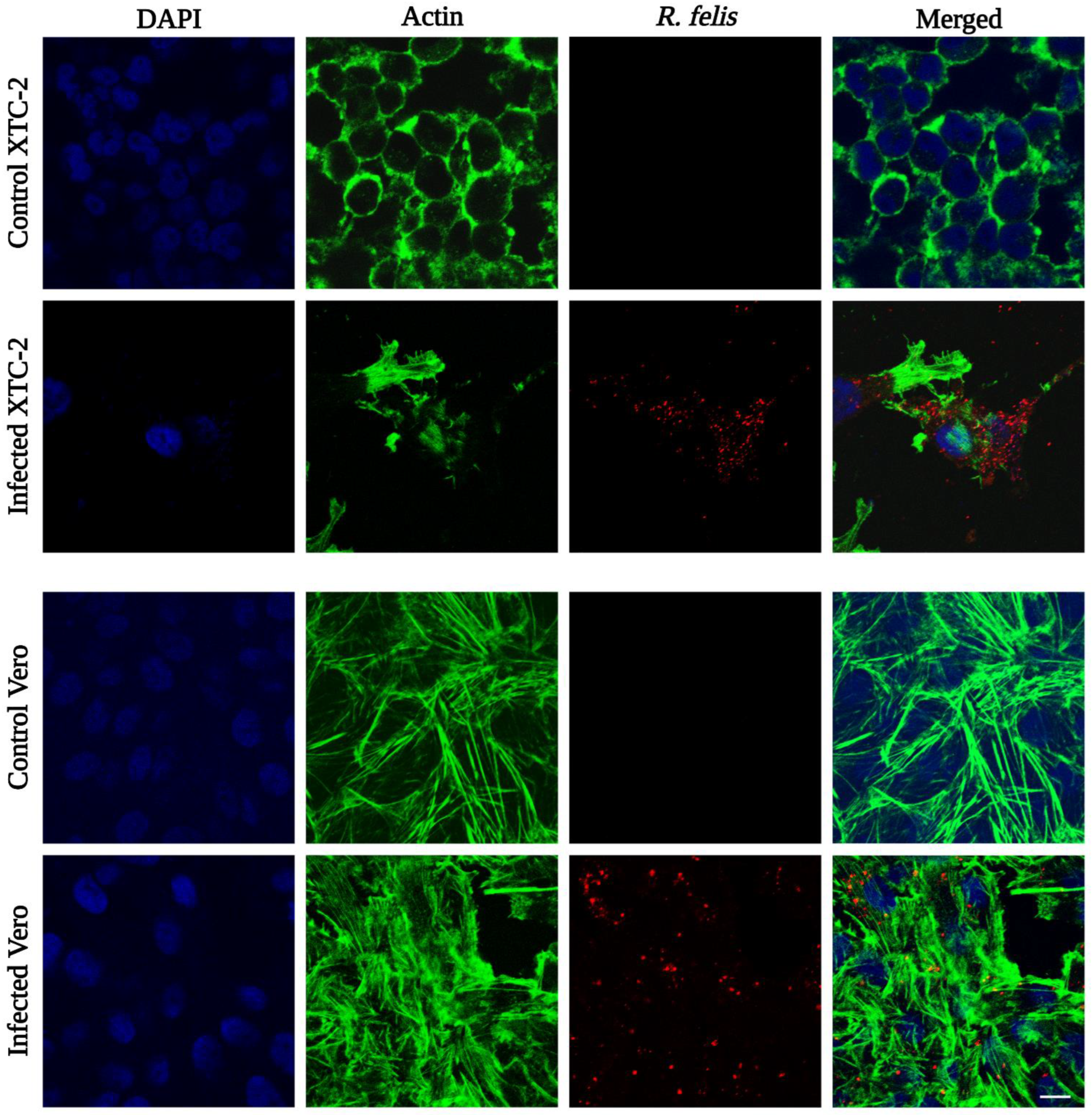
Rickettsia felis Danube was observed in infected XTC-2 and Vero cells by indirect immunofluorescence assay. To visualize rickettsiae, we applied anti-R. conorii rabbit serum as the primary antibody, followed by a rhodamine-conjugated secondary antibody. Serologically, spotted fever group rickettsia antibodies are cross-reactive with R. felis antigens [98,99]. We labeled the actin filaments of host cells with conjugated phalloidin, and cell nuclei were counterstained with DAPI. Confocal microscopy demonstrated that rickettsiae were present in the cytoplasm of the infected host cells (Figure 6), similar to the initial isolate “ELB” and the reference strain [49,51]. Unlike the standard strain, but similar to R. felis LSU, we did not observe evident intracellular actin polymerization by IFA [55].
To visualize the ultrastructure of R. felis Danube, we examined the purified rickettsiae using transmission and scanning electron microscopy (Figure 7). The morphology of the isolate (appearing as a typical-shaped coccobacillus) was characteristic of rickettsiae. The size of the detected microorganisms was 0.3 µm in width and 0.9 µm in length. We repeatedly observed bacteria undergoing binary fission. Even after comprehensive purification, occasional host cell debris could be seen in our samples.
3.5. A Possible Role of Ticks in the Ecology of R. felis
The use of molecular biology and cell culture techniques to uncover microorganisms in arthropod disease vectors has enabled new discoveries. The list of circulating pathogens that can be found in Slovakia has broadly expanded in the past decade. Although successful cultivation of R. felis has been achieved before (Table 1), this is the first cultivation of a tick-derived strain, and the first isolate originated from Europe. Our study revealed that ticks in Slovakia, alongside fleas [44,45], can harbor viable, infectious R. felis.
In the present study, R. felis was isolated from a questing I. ricinus nymph, collected from vegetation in spring. We assume that a larval tick was most likely infected the year before in summer, molted, and then overwintered with the rickettsial pathogen. The dormancy phenomenon (diapause) in Central Europe is part of the I. ricinus life cycle [100]. Similar to our finding, R. felis was identified in a questing I. ricinus nymph in France [36], indicating transstadial transmission of the bacteria.
The acquisition of rickettsiae by ticks occurs via feeding on a rickettsiaemic vertebrate host or by cofeeding transmission between arthropods. Small rodents, insectivores, and birds are important hosts for the immature stages of Ixodid ticks, including the castor bean tick [101]. Rickettsia felis DNA has been detected in ear tissues of Apodemus sylvaticus, Apodemus flavicollis, and Myodes glareolus in Germany [102,103]. In addition, hedgehogs can carry R. felis-positive ticks and fleas and may act as a reservoir host for pathogens [104,105,106]. Recently, R. felis was also identified in a I. ricinus nymph removed from a common blackbird (Turdus merula) in Romania [39].
Immature ticks can compensate for the absence of birds and small mammals by feeding on larger animals [101]. Feral and/or companion cats and dogs; domesticated animals such as cattle, horses, and sheep; as well as wildlife (all present in the tick collection area in Slovakia) can be reservoirs of vector-borne zoonotic Rickettsia spp., such as R. felis (Table S1), and may facilitate the horizontal transmission of these pathogens.
Ticks can harbor a number of bacterial species within the Rickettsia genus and are capable of transmitting these microorganisms to humans during a bite. Lejal and colleagues detected R. felis in the salivary glands of unfed, adult I. ricinus ticks by microfluidic real-time PCR [37]. A new publication from Serbia examining ticks feeding on humans and their blood samples revealed a case study of R. felis infection in an elderly patient parasitized by an adult I. ricinus female tick [40]. However, to date, culture isolation of R. felis from a clinical sample has not been achieved [3].
Much work still lies ahead to fully understand the complex ecology of R. felis. Laboratory experiments are necessary to clarify the vector competence of ticks in the transmission route of R. felis. Further investigation is needed regarding transovarial [107] and transstadial transmission of this microorganism to subsequent life stages of ticks. Also, additional evidence is required to verify the potential role of ticks in the transmission of R. felis to vertebrate hosts, including humans.
4. Conclusions
The present study described the successful isolation of a tick-derived R. felis strain obtained from an unfed I. ricinus nymph. The isolation was performed in XTC-2 cells at 28 °C by the shell-vial technique. Evaluation of bacterial growth was conducted for both XTC-2 and Vero cell lines. We observed R. felis in infected host cells by Gimenez staining and IFA. The new R. felis isolate was purified by gradient ultracentrifugation and visualized by electron microscopy. For genetic analysis, fragments of the genes gltA, ompA, ompB, htrA, rpoB, sca4, rrfE, and rrs were sequenced and compared with the corresponding sequences of the R. felis type strain URRWXCal2 and the other reported culture isolates. We did not detect any nucleotide variability; however, the plasmid pRFδ was not present in our isolate. Our findings demonstrated that ticks harbor viable R. felis in Slovakia. Further studies regarding the role of ticks in the ecology of R. felis are warranted.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijerph19074321/s1, Figure S1: Centrifugation tubes with R. felis isolate Danube purified from XTC-2 cells by isopycnic density gradient ultracentrifugation; Figure S2: PCR confirmation of the absence of R. helvetica contamination in the purified isolate Danube; Table S1: Molecular detection of R. felis in ticks; Table S2: Oligonucleotides used in the study for conventional PCR and sequencing of the R. felis strain Danube; Table S3: Sequence similarities of selected rickettsial gene fragments of the Slovak isolate Danube with other R. felis strains.
Author Contributions
Conceptualization, M.D. and Z.S.; methodology M.D. and Z.S.; validation M.D., O.B. and Z.S.; formal analysis, M.D.; investigation, M.D., O.B. and Z.S.; resources Ľ.Š. and Z.S.; data curation M.D., O.B. and Z.S.; writing—original draft preparation M.D.; writing—review and editing M.D., O.B., Ľ.Š. and Z.S.; visualization M.D.; supervision Ľ.Š. and Z.S.; funding acquisition, Z.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Slovak Research and Development Agency: SRDA-19-0066, and SRDA-19-0519; and Scientific Grant Agency of the Ministry of Education of the Slovak Republic, and the Slovak Academy of Sciences (VEGA): 2/0010/19 and 2/0021/21.
Institutional Review Board Statement
The usage of animals in the experiment was approved by the Ethical Committee of the Biomedical Research Center, Slovak Academy of Sciences, and the State Veterinary and Food Administration of the Slovak Republic (permit number: RO-2977-3/2020-220). The experiments were performed under standard conditions in the experimental animal facility of the Biomedical Research Center, Slovak Academy of Sciences.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We would like to thank Gabriela Flores Ramirez and Fernando Zuñiga Navarrete for their helpful comments on molecular biology methods. We thank Lucia Csáderová for the technical help with confocal microscopy. We also thank Meghan Hermance for proofreading our manuscript. The Graphical Abstract and the figures were created with BioRender.com (accessed on 27 January 2022).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schriefer, M.E.; Sacci, J.B.; Dumler, J.S.; Bullen, M.G.; Azad, A.F. Identification of a novel rickettsial infection in a patient diagnosed with murine typhus. J. Clin. Microbiol. 1994, 32, 949–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdad, M.Y.; Stenos, J.; Graves, S. Rickettsia felis, an emerging flea-transmitted human pathogen. Emerg. Health Threat. J. 2011, 4, 7168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanton, L.S.; Walker, D.H. Flea-Borne Rickettsioses and Rickettsiae. Am. J. Trop. Med. Hyg. 2017, 96, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Azad, A.F.; Radulovic, S.; Higgins, J.A.; Noden, B.H.; Troyer, J.M. Flea-borne rickettsioses: Ecologic considerations. Emerg. Infect. Dis. 1997, 3, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Zavala-Velázquez, J.E.; Ruiz-Sosa, J.A.; Sánchez-Elias, R.A.; Becerra-Carmona, G.; Walker, D.H. Rickettsia felis rickettsiosis in Yucatán. Lancet 2000, 356, 1079–1080. [Google Scholar] [CrossRef]
- Lindblom, A.; Severinson, K.; Nilsson, K. Rickettsia felis infection in Sweden: Report of two cases with subacute meningitis and review of the literature. Scand. J. Infect. Dis. 2010, 42, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Mawuntu, A.H.P.; Johar, E.; Anggraeni, R.; Feliana, F.; Bernadus, J.B.B.; Safari, D.; Yudhaputri, F.A.; Dhenni, R.; Dewi, Y.P.; Kato, C.; et al. Rickettsia felis identified in two fatal cases of acute meningoencephalitis. PLoS Negl. Trop. Dis. 2020, 14, e0007893. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Osorio, C.E.; Zavala-Velázquez, J.E.; Arias León, J.J.; Zavala-Castro, J.E. Rickettsia felis as emergent global threat for humans. Emerg. Infect. Dis. 2008, 14, 1019–1023. [Google Scholar] [CrossRef]
- Angelakis, E.; Mediannikov, O.; Parola, P.; Raoult, D. Rickettsia felis: The Complex Journey of an Emergent Human Pathogen. Trends Parasitol. 2016, 32, 554–564. [Google Scholar] [CrossRef]
- Parola, P. Rickettsia felis: From a rare disease in the USA to a common cause of fever in sub-Saharan Africa. Clin. Microbiol. Infect. 2011, 17, 996–1000. [Google Scholar] [CrossRef] [Green Version]
- Márquez, F.J.; Muniain, M.A.; Pérez, J.M.; Pachón, J. Presence of Rickettsia felis in the cat flea from southwestern Europe. Emerg. Infect. Dis. 2002, 8, 89–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reif, K.E.; Macaluso, K.R. Ecology of Rickettsia felis: A review. J. Med. Entomol. 2009, 46, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.D.; Macaluso, K.R. Rickettsia felis, an Emerging Flea-Borne Rickettsiosis. Curr. Trop. Med. Rep. 2016, 3, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, L.D.; Freitas, R.N.; Mafra, C.L.; Neves, C.V.; Figueira, F.C.; Labruna, M.B.; Gennari, S.M.; Walker, D.H.; Galvão, M.A. Characterization of Rickettsia spp. circulating in a silent peri-urban focus for Brazilian spotted fever in Caratinga, Minas Gerais, Brazil. Cad. Saude Publica 2006, 22, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, K.A.; Oliveira, L.S.; Dias, C.C.; Silva, A.; Almeida, M.R.; Almada, G.; Bouyer, D.H.; Galvão, M.A.; Mafra, C. Molecular identification of Rickettsia felis in ticks and fleas from an endemic area for Brazilian Spotted Fever. Mem. Inst. Oswaldo Cruz 2008, 103, 191–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo, A.; Olmeda, A.S.; Escudero, R.; Jado, I.; Valcárcel, F.; Casado-Nistal, M.A.; Rodríguez-Vargas, M.; Gil, H.; Anda, P. Tick-borne zoonotic bacteria in ticks collected from central Spain. Am. J. Trop. Med. Hyg. 2009, 81, 67–74. [Google Scholar] [CrossRef]
- Abarca, K.; López, J.; Acosta-Jamett, G.; Martínez-Valdebenito, C. Rickettsia felis in Rhipicephalus sanguineus from two distant Chilean cities. Vector Borne Zoonotic Dis. 2013, 13, 607–609. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, G.; Kelly, P.; Zhang, Z.; Wei, L.; Yu, D.; Kayizha, S.; Wang, C. First report of Rickettsia felis in China. BMC Infect. Dis. 2014, 14, 682. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.L.; Colella, V.; Greco, G.; Fang, F.; Nurcahyo, W.; Hadi, U.K.; Venturina, V.; Tong, K.B.Y.; Tsai, Y.L.; Taweethavonsawat, P.; et al. Molecular detection of pathogens in ticks and fleas collected from companion dogs and cats in East and Southeast Asia. Parasites Vectors 2020, 13, 420. [Google Scholar] [CrossRef]
- Campos, S.D.E.; Cunha, N.C.D.; Machado, C.S.C.; Telleria, E.L.; Cordeiro, M.D.; Fonseca, A.H.D.; Toma, H.K.; Santos, J.P.C.D.; Almosny, N.R.P. Rickettsial pathogens circulating in urban districts of Rio de Janeiro, without report of human Brazilian Spotted Fever. Rev. Bras. Parasitol. Vet. 2020, 29, e014220. [Google Scholar] [CrossRef]
- de Oliveira, J.C.P.; Reckziegel, G.H.; Ramos, C.A.D.N.; Giannelli, A.; Alves, L.C.; de Carvalho, G.A.; Ramos, R.A.N. Detection of Rickettsia felis in ectoparasites collected from domestic animals. Exp. Appl. Acarol. 2020, 81, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Gargili, A.; Palomar, A.M.; Midilli, K.; Portillo, A.; Kar, S.; Oteo, J.A. Rickettsia species in ticks removed from humans in Istanbul, Turkey. Vector Borne Zoonotic Dis. 2012, 12, 938–941. [Google Scholar] [CrossRef] [PubMed]
- Raele, D.A.; Galante, D.; Pugliese, N.; Salandra, G.; Cafiero, M.A. Spotted fever group rickettsiae associated with ixodid ticks in wild environment in Southern Italy. Microbiologyopen 2018, 7, e00527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikura, M.; Ando, S.; Shinagawa, Y.; Matsuura, K.; Hasegawa, S.; Nakayama, T.; Fujita, H.; Watanabe, M. Phylogenetic analysis of spotted fever group rickettsiae based on gltA, 17-kDa, and rOmpA genes amplified by nested PCR from ticks in Japan. Microbiol. Immunol. 2003, 47, 823–832. [Google Scholar] [CrossRef] [Green Version]
- Duh, D.; Punda-Polić, V.; Trilar, T.; Petrovec, M.; Bradarić, N.; Avsic-Zupanc, T. Molecular identification of Rickettsia felis-like bacteria in Haemaphysalis sulcata ticks collected from domestic animals in southern Croatia. Ann. N. Y. Acad. Sci. 2006, 1078, 347–351. [Google Scholar] [CrossRef]
- Roth, T.; Lane, R.S.; Foley, J. A Molecular Survey for Francisella tularensis and Rickettsia spp. in Haemaphysalis leporispalustris (Acari: Ixodidae) in Northern California. J. Med. Entomol. 2017, 54, 492–495. [Google Scholar] [CrossRef]
- Khoo, J.J.; Chen, F.; Kho, K.L.; Shanizza, A.I.A.; Lim, F.S.; Tan, K.K.; Chang, L.Y.; AbuBakar, S. Bacterial community in Haemaphysalis ticks of domesticated animals from the Orang Asli communities in Malaysia. Ticks Tick Borne Dis. 2016, 7, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Chalada, M.J.; Stenos, J.; Vincent, G.; Barker, D.; Bradbury, R.S. A Molecular Survey of Tick-Borne Pathogens from Ticks Collected in Central Queensland, Australia. Vector Borne Zoonotic Dis. 2018, 18, 151–163. [Google Scholar] [CrossRef]
- Jiang, J.; Stromdahl, E.Y.; Richards, A.L. Detection of Rickettsia parkeri and Candidatus Rickettsia andeanae in Amblyomma maculatum Gulf Coast ticks collected from humans in the United States. Vector Borne Zoonotic Dis. 2012, 12, 175–182. [Google Scholar] [CrossRef]
- Soares, H.S.; Barbieri, A.R.; Martins, T.F.; Minervino, A.H.; de Lima, J.T.; Marcili, A.; Gennari, S.M.; Labruna, M.B. Ticks and rickettsial infection in the wildlife of two regions of the Brazilian Amazon. Exp. Appl. Acarol. 2015, 65, 125–140. [Google Scholar] [CrossRef]
- Bitencourth, K.; Amorim, M.; DE Oliveira, S.V.; Caetano, R.L.; Voloch, C.M.; Gazêta, G.S. Amblyomma sculptum: Genetic diversity and rickettsias in the Brazilian Cerrado biome. Med. Vet. Entomol. 2017, 31, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Bitencourth, K.; Amorim, M.; de Oliveira, S.V.; Voloch, C.M.; Gazêta, G.S. Genetic diversity, population structure and rickettsias in Amblyomma ovale in areas of epidemiological interest for spotted fever in Brazil. Med. Vet. Entomol. 2019, 33, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Tsui, P.Y.; Tsai, K.H.; Weng, M.H.; Hung, Y.W.; Liu, Y.T.; Hu, K.Y.; Lien, J.C.; Lin, P.R.; Shaio, M.F.; Wang, H.C.; et al. Molecular detection and characterization of spotted fever group rickettsiae in Taiwan. Am. J. Trop. Med. Hyg. 2007, 77, 883–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascucci, I.; Di Domenico, M.; Curini, V.; Cocco, A.; Averaimo, D.; D’Alterio, N.; Cammà, C. Diversity of Rickettsia in Ticks Collected in Abruzzi and Molise Regions (Central Italy). Microorganisms 2019, 7, 696. [Google Scholar] [CrossRef] [Green Version]
- Dobler, G.; Wölfel, R. Typhus and other rickettsioses: Emerging infections in Germany. Dtsch. Arztebl. Int. 2009, 106, 348–354. [Google Scholar] [CrossRef]
- Lejal, E.; Marsot, M.; Chalvet-Monfray, K.; Cosson, J.F.; Moutailler, S.; Vayssier-Taussat, M.; Pollet, T. A three-years assessment of Ixodes ricinus-borne pathogens in a French peri-urban forest. Parasites Vectors 2019, 12, 551. [Google Scholar] [CrossRef] [Green Version]
- Lejal, E.; Moutailler, S.; Šimo, L.; Vayssier-Taussat, M.; Pollet, T. Tick-borne pathogen detection in midgut and salivary glands of adult Ixodes ricinus. Parasites Vectors 2019, 12, 152. [Google Scholar] [CrossRef] [Green Version]
- Remesar, S.; Díaz, P.; Portillo, A.; Santibáñez, S.; Prieto, A.; Díaz-Cao, J.M.; López, C.M.; Panadero, R.; Fernández, G.; Díez-Baños, P.; et al. Prevalence and molecular characterization of Rickettsia spp. in questing ticks from north-western Spain. Exp. Appl. Acarol. 2019, 79, 267–278. [Google Scholar] [CrossRef]
- Borşan, S.D.; Ionică, A.M.; Galon, C.; Toma-Naic, A.; Peştean, C.; Sándor, A.D.; Moutailler, S.; Mihalca, A.D. High Diversity, Prevalence, and Co-infection Rates of Tick-Borne Pathogens in Ticks and Wildlife Hosts in an Urban Area in Romania. Front. Microbiol. 2021, 12, 645002. [Google Scholar] [CrossRef]
- Banović, P.; Díaz-Sánchez, A.A.; Galon, C.; Simin, V.; Mijatović, D.; Obregón, D.; Moutailler, S.; Cabezas-Cruz, A. Humans infested with Ixodes ricinus are exposed to a diverse array of tick-borne pathogens in Serbia. Ticks Tick Borne Dis. 2021, 12, 101609. [Google Scholar] [CrossRef]
- Díaz-Sánchez, A.A.; Chilton, N.B.; Roblejo-Arias, L.; Fonseca-Rodríguez, O.; Marrero-Perera, R.; Diyes, C.P.; Yunik, M.E.M.; Lobo-Rivero, E.; Corona-González, B. Molecular detection and identification of spotted fever group rickettsiae in ticks collected from horses in Cuba. Med. Vet. Entomol. 2021, 35, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Stanley, H.; Rhodes, D.V.L. Presence of Rickettsia Species in Ticks Collected from Companion Animals in Northeastern Georgia, United States. Vet. Sci. 2021, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.K.; Loftis, A.D.; Sanders, F.; Spinks, M.D.; Wills, W.; Denison, A.M.; Dasch, G.A. Borrelia, Coxiella, and Rickettsia in Carios capensis (Acari: Argasidae) from a brown pelican (Pelecanus occidentalis) rookery in South Carolina, USA. Exp. Appl. Acarol. 2006, 39, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Špitalská, E.; Boldiš, V.; Mošanský, L.; Sparagano, O.; Stanko, M. Rickettsia species in fleas collected from small mammals in Slovakia. Parasitol. Res. 2015, 114, 4333–4339. [Google Scholar] [CrossRef] [PubMed]
- Heglasová, I.; Víchová, B.; Stanko, M. Detection of Rickettsia spp. in Fleas Collected from Small Mammals in Slovakia, Central Europe. Vector Borne Zoonotic Dis. 2020, 20, 652–656. [Google Scholar] [CrossRef]
- Adams, J.R.; Schmidtmann, E.T.; Azad, A.F. Infection of colonized cat fleas, Ctenocephalides felis (Bouché), with a rickettsia-like microorganism. Am. J. Trop. Med. Hyg. 1990, 43, 400–409. [Google Scholar] [CrossRef]
- Radulovic, S.; Higgins, J.A.; Jaworski, D.C.; Dasch, G.A.; Azad, A.F. Isolation, cultivation, and partial characterization of the ELB agent associated with cat fleas. Infect. Immun. 1995, 63, 4826–4829. [Google Scholar] [CrossRef] [Green Version]
- Radulovic, S.; Higgins, J.A.; Jaworski, D.C.; Azad, A.F. In vitro and in vivo antibiotic susceptibilities of ELB rickettsiae. Antimicrob. Agents Chemother. 1995, 39, 2564–2566. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.A.; Radulovic, S.; Schriefer, M.E.; Azad, A.F. Rickettsia felis: A new species of pathogenic rickettsia isolated from cat fleas. J. Clin. Microbiol. 1996, 34, 671–674. [Google Scholar] [CrossRef] [Green Version]
- Bouyer, D.H.; Stenos, J.; Crocquet-Valdes, P.; Moron, C.G.; Popov, V.L.; Zavala-Velazquez, J.E.; Foil, L.D.; Stothard, D.R.; Azad, A.F.; Walker, D.H. Rickettsia felis: Molecular characterization of a new member of the spotted fever group. Int. J. Syst. Evol. Microbiol. 2001, 51, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Raoult, D.; La Scola, B.; Enea, M.; Fournier, P.E.; Roux, V.; Fenollar, F.; Galvao, M.A.; de Lamballerie, X. A flea-associated Rickettsia pathogenic for humans. Emerg. Infect. Dis. 2001, 7, 73–81. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Meconi, S.; Fenollar, F.; Rolain, J.M.; Roux, V.; Raoult, D. Emended description of Rickettsia felis (Bouyer et al. 2001), a temperature-dependent cultured bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52, 2035–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hii, S.F.; Abdad, M.Y.; Kopp, S.R.; Stenos, J.; Rees, R.L.; Traub, R.J. Seroprevalence and risk factors for Rickettsia felis exposure in dogs from Southeast Queensland and the Northern Territory, Australia. Parasites Vectors 2013, 6, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng-Nguyen, D.; Hii, S.F.; Hoang, M.T.; Nguyen, V.T.; Rees, R.; Stenos, J.; Traub, R.J. Domestic dogs are mammalian reservoirs for the emerging zoonosis flea-borne spotted fever, caused by Rickettsia felis. Sci. Rep. 2020, 10, 4151. [Google Scholar] [CrossRef] [PubMed]
- Pornwiroon, W.; Pourciau, S.S.; Foil, L.D.; Macaluso, K.R. Rickettsia felis from cat fleas: Isolation and culture in a tick-derived cell line. Appl. Environ. Microbiol. 2006, 72, 5589–5595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thepparit, C.; Sunyakumthorn, P.; Guillotte, M.L.; Popov, V.L.; Foil, L.D.; Macaluso, K.R. Isolation of a rickettsial pathogen from a non-hematophagous arthropod. PLoS ONE 2011, 6, e16396. [Google Scholar] [CrossRef] [PubMed]
- Horta, M.C.; Labruna, M.B.; Durigon, E.L.; Schumaker, T.T. Isolation of Rickettsia felis in the mosquito cell line C6/36. Appl. Environ. Microbiol. 2006, 72, 1705–1707. [Google Scholar] [CrossRef] [Green Version]
- Hun, L.; Troyo, A.; Taylor, L.; Barbieri, A.M.; Labruna, M.B. First report of the isolation and molecular characterization of Rickettsia amblyommii and Rickettsia felis in Central America. Vector Borne Zoonotic Dis. 2011, 11, 1395–1397. [Google Scholar] [CrossRef]
- Sakamoto, J.M.; Azad, A.F. Propagation of arthropod-borne Rickettsia spp. in two mosquito cell lines. Appl. Environ. Microbiol. 2007, 73, 6637–6643. [Google Scholar] [CrossRef] [Green Version]
- Luce-Fedrow, A.; Macaluso, K.R.; Richards, A.L. Growth of Rickettsia felis in Drosophila melanogaster S2 cells. Vector Borne Zoonotic Dis. 2014, 14, 101–110. [Google Scholar] [CrossRef]
- Saisongkorh, W.; El Karkouri, K.; Patrice, J.Y.; Bernard, A.; Rolain, J.M.; Raoult, D. Tryptose phosphate broth improves Rickettsia felis replication in mammalian cells. FEMS Immunol. Med. Microbiol. 2012, 64, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, F.; Pons, I.; Pla, J.; Nogueras, M.M. Shell-vial culture and real-time PCR applied to Rickettsia typhi and Rickettsia felis detection. World J. Microbiol. Biotechnol. 2015, 31, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, H.L.; Labruna, M.B.; Montenieri, J.A.; Kosoy, M.Y.; Gage, K.L.; Walker, D.H. Detection of Rickettsia felis in a New World flea species, Anomiopsyllus nudata (Siphonaptera: Ctenophthalmidae). J. Med. Entomol. 2005, 42, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Nosek, J.; Sixl, W. Central-European ticks (Ixodoidea). Key for determination. Mitt. Abt. Zool. Landesmus. Joanneum 1972, 1, 61–92. [Google Scholar]
- Sekeyová, Z.; Mediannikov, O.; Subramanian, G.; Kowalczewska, M.; Quevedo-Diaz, M.; Kocianová, E.; Raoult, D. Isolation of rickettsia helvetica from ticks in Slovakia. Acta Virol. 2012, 56, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Pudney, M.; Varma, M.G.; Leake, C.J. Establishment of a cell line (XTC-2) from the South African clawed toad, Xenopus laevis. Experientia 1973, 29, 466–467. [Google Scholar] [CrossRef]
- Giménez, D.F. Staining Rickettsiae in Yolk-Sac Cultures. Stain Technol. 1964, 39, 135–140. [Google Scholar] [CrossRef]
- Ammerman, N.C.; Beier-Sexton, M.; Azad, A.F. Laboratory maintenance of Rickettsia rickettsii. Curr. Protoc. Microbiol. 2008, 11, 3A.5.1–3A.5.21. [Google Scholar] [CrossRef] [Green Version]
- Weiss, E.; Coolbaugh, J.C.; Williams, J.C. Separation of viable Rickettsia typhi from yolk sac and L cell host components by renografin density gradient centrifugation. Appl. Microbiol. 1975, 30, 456–463. [Google Scholar] [CrossRef]
- Bohacsova, M.; Filipcik, P.; Opattova, A.; Valarikova, J.; Diaz, M.Q.; Skultety, L.; Novak, M.; Sekeyova, Z. Survival of rat cerebrocortical neurons after rickettsial infection. Microbes Infect. 2015, 17, 845–849. [Google Scholar] [CrossRef]
- Valarikova, J.; Sekeyova, Z.; Skultety, L.; Bohacsova, M.; Quevedo-Diaz, M. New way of purification of pathogenic rickettsiae reducing health risks. Acta Virol. 2016, 60, 206–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, V.; Raoult, D. Phylogenetic analysis of the genus Rickettsia by 16S rDNA sequencing. Res. Microbiol. 1995, 146, 385–396. [Google Scholar] [CrossRef]
- Al Masalma, M.; Armougom, F.; Scheld, W.M.; Dufour, H.; Roche, P.H.; Drancourt, M.; Raoult, D. The expansion of the microbiological spectrum of brain abscesses with use of multiple 16S ribosomal DNA sequencing. Clin. Infect. Dis. 2009, 48, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Sekeyova, Z.; Roux, V.; Raoult, D. Phylogeny of Rickettsia spp. inferred by comparing sequences of ’gene D’, which encodes an intracytoplasmic protein. Int. J. Syst. Evol. Microbiol. 2001, 51, 1353–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labruna, M.B.; Whitworth, T.; Horta, M.C.; Bouyer, D.H.; McBride, J.W.; Pinter, A.; Popov, V.; Gennari, S.M.; Walker, D.H. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of São Paulo, Brazil, where Brazilian spotted fever is endemic. J. Clin. Microbiol. 2004, 42, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Rydkina, E.; Eremeeva, M.; Raoult, D. Citrate synthase gene comparison, a new tool for phylogenetic analysis, and its application for the rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavala-Castro, J.E.; Small, M.; Keng, C.; Bouyer, D.H.; Zavala-Velázquez, J.; Walker, D.H. Transcription of the Rickettsia felis ompA gene in naturally infected fleas. Am. J. Trop. Med. Hyg. 2005, 73, 662–666. [Google Scholar] [CrossRef]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Drancourt, M.; Raoult, D. Characterization of mutations in the rpoB gene in naturally rifampin-resistant Rickettsia species. Antimicrob. Agents Chemother. 1999, 43, 2400–2403. [Google Scholar] [CrossRef] [Green Version]
- Boretti, F.S.; Perreten, A.; Meli, M.L.; Cattori, V.; Willi, B.; Wengi, N.; Hornok, S.; Honegger, H.; Hegglin, D.; Woelfel, R.; et al. Molecular Investigations of Rickettsia helvetica infection in dogs, foxes, humans, and Ixodes ticks. Appl. Environ. Microbiol. 2009, 75, 3230–3237. [Google Scholar] [CrossRef] [Green Version]
- Fournier, P.E.; Belghazi, L.; Robert, C.; Elkarkouri, K.; Richards, A.L.; Greub, G.; Collyn, F.; Ogawa, M.; Portillo, A.; Oteo, J.A.; et al. Variations of plasmid content in Rickettsia felis. PLoS ONE 2008, 3, e2289. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Danchenko, M.; Csaderova, L.; Fournier, P.E.; Sekeyova, Z. Optimized fixation of actin filaments for improved indirect immunofluorescence staining of rickettsiae. BMC Res. Notes 2019, 12, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobue, K.; Fujio, Y.; Kanda, K. Tumor promoter induces reorganization of actin filaments and calspectin (fodrin or nonerythroid spectrin) in 3T3 cells. Proc. Natl. Acad. Sci. USA 1988, 85, 482–486. [Google Scholar] [CrossRef] [Green Version]
- Benada, O.; Pokorný, V. Modification of the Polaron sputter-coater unit for glow-discharge activation of carbon support films. J. Electron. Microsc. Tech. 1990, 16, 235–239. [Google Scholar] [CrossRef]
- Sekeyová, Z.; Fournier, P.E.; Rehácek, J.; Raoult, D. Characterization of a new spotted fever group rickettsia detected in Ixodes ricinus (Acari: Ixodidae) collected in Slovakia. J. Med. Entomol. 2000, 37, 707–713. [Google Scholar] [CrossRef]
- Boldis, V.; Strus, J.; Kocianová, E.; Tusek-Znidaric, M.; Stefanidesová, K.; Schwarzová, K.; Kúdelová, M.; Sekeyová, Z.; Spitalská, E. Life cycle of Rickettsia slovaca in L929 cell line studied by quantitative real-time PCR and transmission electron microscopy. FEMS Microbiol. Lett. 2009, 293, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Mediannikov, O.; Sekeyová, Z.; Birg, M.L.; Raoult, D. A novel obligate intracellular gamma-proteobacterium associated with ixodid ticks, Diplorickettsia massiliensis, Gen. Nov., sp. Nov. PLoS ONE 2010, 5, e11478. [Google Scholar] [CrossRef]
- Mediannikov, O.; Subramanian, G.; Sekeyova, Z.; Bell-Sakyi, L.; Raoult, D. Isolation of Arsenophonus nasoniae from Ixodes ricinus ticks in Slovakia. Ticks Tick Borne Dis. 2012, 3, 367–370. [Google Scholar] [CrossRef]
- Spitalská, E.; Stefanidesová, K.; Kocianová, E.; Boldiš, V. Rickettsia slovaca and Rickettsia raoultii in Dermacentor marginatus and Dermacentor reticulatus ticks from Slovak Republic. Exp. Appl. Acarol. 2012, 57, 189–197. [Google Scholar] [CrossRef]
- Špitalská, E.; Boldiš, V.; Derdáková, M.; Selyemová, D.; Rusňáková Taragel’ová, V. Rickettsial infection in Ixodes ricinus ticks in urban and natural habitats of Slovakia. Ticks Tick Borne Dis. 2014, 5, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Svehlová, A.; Berthová, L.; Sallay, B.; Boldiš, V.; Sparagano, O.A.; Spitalská, E. Sympatric occurrence of Ixodes ricinus, Dermacentor reticulatus and Haemaphysalis concinna ticks and Rickettsia and Babesia species in Slovakia. Ticks Tick Borne Dis. 2014, 5, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Špitalská, E.; Stanko, M.; Mošanský, L.; Kraljik, J.; Miklisová, D.; Mahríková, L.; Bona, M.; Kazimírová, M. Seasonal analysis of Rickettsia species in ticks in an agricultural site of Slovakia. Exp. Appl. Acarol. 2016, 68, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Berthová, L.; Slobodník, V.; Slobodník, R.; Olekšák, M.; Sekeyová, Z.; Svitálková, Z.; Kazimírová, M.; Špitalská, E. The natural infection of birds and ticks feeding on birds with Rickettsia spp. and Coxiella burnetii in Slovakia. Exp. Appl. Acarol. 2016, 68, 299–314. [Google Scholar] [CrossRef]
- Ogata, H.; Renesto, P.; Audic, S.; Robert, C.; Blanc, G.; Fournier, P.E.; Parinello, H.; Claverie, J.M.; Raoult, D. The genome sequence of Rickettsia felis identifies the first putative conjugative plasmid in an obligate intracellular parasite. PLoS Biol. 2005, 3, e248. [Google Scholar] [CrossRef]
- Gillespie, J.J.; Driscoll, T.P.; Verhoeve, V.I.; Utsuki, T.; Husseneder, C.; Chouljenko, V.N.; Azad, A.F.; Macaluso, K.R. Genomic diversification in strains of Rickettsia felis Isolated from different arthropods. Genome Biol. Evol. 2014, 7, 35–56. [Google Scholar] [CrossRef]
- Bitam, I.; Parola, P.; De La Cruz, K.D.; Matsumoto, K.; Baziz, B.; Rolain, J.M.; Belkaid, M.; Raoult, D. First molecular detection of Rickettsia felis in fleas from Algeria. Am. J. Trop. Med. Hyg. 2006, 74, 532–535. [Google Scholar] [CrossRef]
- Znazen, A.; Rolain, J.M.; Hammami, A.; Jemaa, M.B.; Raoult, D. Rickettsia felis infection, Tunisia. Emerg. Infect. Dis. 2006, 12, 138–140. [Google Scholar] [CrossRef]
- Fang, R.; Raoult, D. Antigenic classification of Rickettsia felis by using monoclonal and polyclonal antibodies. Clin. Diagn. Lab. Immunol. 2003, 10, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S.; Kahl, O.; Lane, R.S.; Levin, M.L.; Tsao, J.I. Diapause in ticks of the medically important Ixodes ricinus species complex. Ticks Tick-Borne Dis. 2016, 7, 992–1003. [Google Scholar] [CrossRef] [Green Version]
- Matuschka, F.R.; Fischer, P.; Musgrave, K.; Richter, D.; Spielman, A. Hosts on which nymphal Ixodes ricinus most abundantly feed. Am. J. Trop. Med. Hyg. 1991, 44, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Spierling, N.G.; Heuser, E.; Kling, C.; Schmidt, S.; Rosenfeld, U.M.; Reil, D.; Imholt, C.; Jacob, J.; Ulrich, R.G.; et al. High prevalence of Rickettsia helvetica in wild small mammal populations in Germany. Ticks Tick Borne Dis. 2018, 9, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Schex, S.; Dobler, G.; Riehm, J.; Müller, J.; Essbauer, S. Rickettsia spp. in wild small mammals in Lower Bavaria, South-Eastern Germany. Vector Borne Zoonotic Dis. 2011, 11, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Riley, P.Y.; Chomel, B.B. Hedgehog zoonoses. Emerg. Infect. Dis. 2005, 11, 1–5. [Google Scholar] [CrossRef]
- Jahfari, S.; Ruyts, S.C.; Frazer-Mendelewska, E.; Jaarsma, R.; Verheyen, K.; Sprong, H. Melting pot of tick-borne zoonoses: The European hedgehog contributes to the maintenance of various tick-borne diseases in natural cycles urban and suburban areas. Parasites Vectors 2017, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- Marié, J.L.; Davoust, B.; Socolovschi, C.; Raoult, D.; Parola, P. Molecular detection of rickettsial agents in ticks and fleas collected from a European hedgehog (Erinaceus europaeus) in Marseilles, France. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 77–79. [Google Scholar] [CrossRef]
- Harris, E.K.; Verhoeve, V.I.; Banajee, K.H.; Macaluso, J.A.; Azad, A.F.; Macaluso, K.R. Comparative vertical transmission of Rickettsia by Dermacentor variabilis and Amblyomma maculatum. Ticks Tick Borne Dis. 2017, 8, 598–604. [Google Scholar] [CrossRef]
Figure 1.
Tick sampling location in Slovakia. On the left: a political map of Europe indicating the location of Slovakia (in light green). Ticks were collected in the Bratislava district (marked with a triangle) by flagging method (maps from www.vidiani.com, accessed on 27 January 2022, licensed under CC-BY 3.0, desaturated from the original). On the right: I. ricinus nymph collected from the local vegetation (SMZ1500 Stereomicroscope).
Figure 1.
Tick sampling location in Slovakia. On the left: a political map of Europe indicating the location of Slovakia (in light green). Ticks were collected in the Bratislava district (marked with a triangle) by flagging method (maps from www.vidiani.com, accessed on 27 January 2022, licensed under CC-BY 3.0, desaturated from the original). On the right: I. ricinus nymph collected from the local vegetation (SMZ1500 Stereomicroscope).

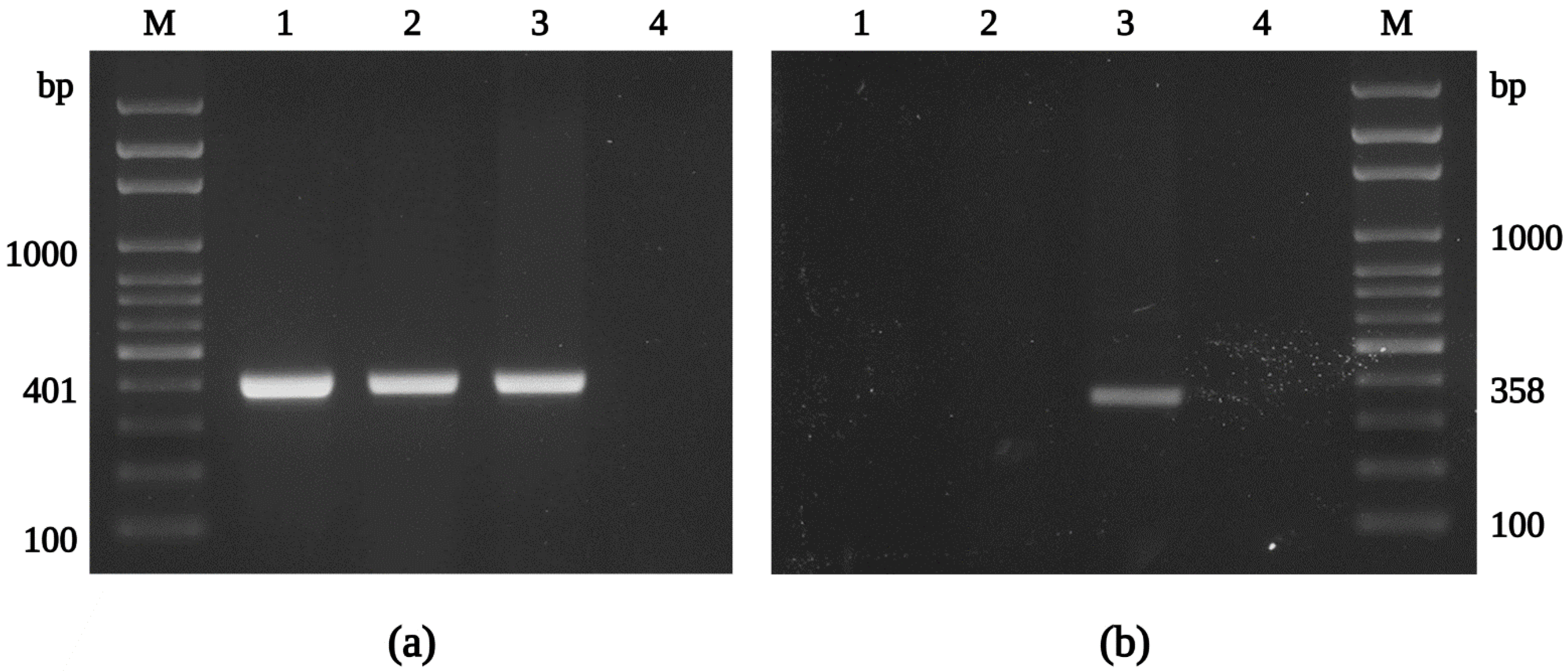
Figure 2.
Identification of the isolate Danube by PCR. Agarose gel electrophoresis of PCR products of the genes gltA (a) and rffE (b), amplified with primers CS-78 and CS-323, MQ32 and MQ33, respectively. M: molecular marker; 1: R. helvetica C9P9; 2: R. helvetica IR16; 3: R. felis Danube; 4: negative control. Note: primer set MQ32, MQ33 does not amplify R. helvetica.
Figure 2.
Identification of the isolate Danube by PCR. Agarose gel electrophoresis of PCR products of the genes gltA (a) and rffE (b), amplified with primers CS-78 and CS-323, MQ32 and MQ33, respectively. M: molecular marker; 1: R. helvetica C9P9; 2: R. helvetica IR16; 3: R. felis Danube; 4: negative control. Note: primer set MQ32, MQ33 does not amplify R. helvetica.

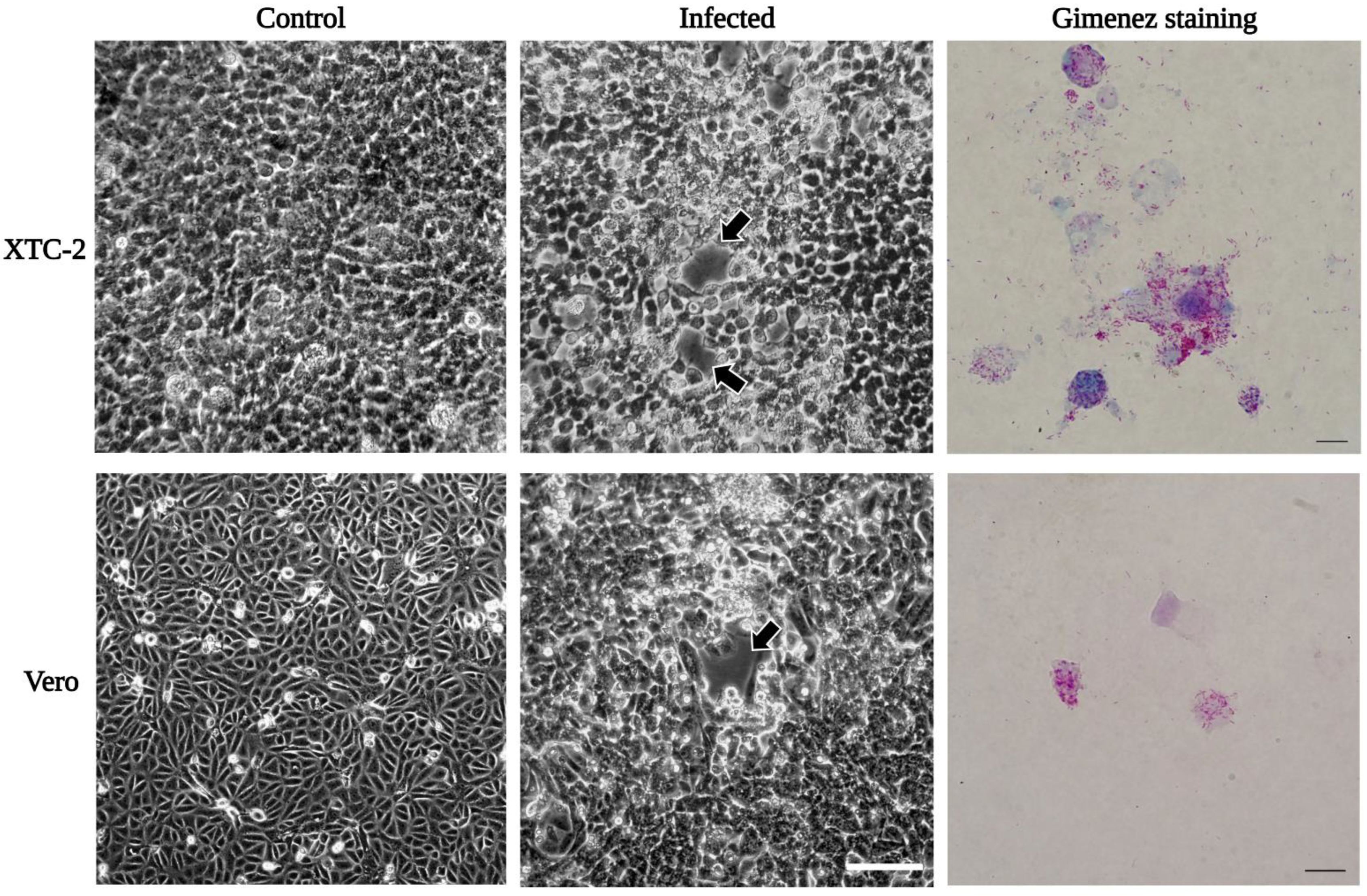
Figure 3.
Growth of R. felis Danube in XTC-2 and Vero cells. Rickettsiae were propagated in XTC-2 and Vero cells at 28 °C and 32 °C, respectively. We observed an apparent cytopathic effect (plaque formation, black arrows) in the infected host cells at 6 dpi in XTC-2 and 14 dpi in Vero cells (Zeiss Axiovert 40 CFL trinocular inverted phase-contrast microscope; scale bar: 100 µm). Rickettsia felis stained magenta in contrast to host cells in blue by Gimenez technique (Leica DM 4500B microscope; scale bar 10 µm).
Figure 3.
Growth of R. felis Danube in XTC-2 and Vero cells. Rickettsiae were propagated in XTC-2 and Vero cells at 28 °C and 32 °C, respectively. We observed an apparent cytopathic effect (plaque formation, black arrows) in the infected host cells at 6 dpi in XTC-2 and 14 dpi in Vero cells (Zeiss Axiovert 40 CFL trinocular inverted phase-contrast microscope; scale bar: 100 µm). Rickettsia felis stained magenta in contrast to host cells in blue by Gimenez technique (Leica DM 4500B microscope; scale bar 10 µm).

Figure 4.
Graphical representation of the rickettsial rate of intracellular growth in XTC-2 cells. Amphibian host cells incubated in 12-well plates and approaching confluence were infected with MOI 10 rickettsiae. Infected cells from culture wells were harvested in 24 h intervals. To demonstrate the growth kinetics of R. felis Danube, the approximate mean numbers of bacterial genome equivalents (GE) were calculated based on the amplification of the rpsL gene by qPCR. Standard errors of the mean were calculated from two biological replicates. For statistical analysis, one-way ANOVA was performed, followed by Dunnett’s multiple comparison test (** p < 0.01, **** p < 0.0001) (a). To confirm viability of rickettsiae, the estimated transcript numbers of the same gene were evaluated per well, as described in Materials and Methods. The red line represents nonlinear regression, Beta growth then decay (b).
Figure 4.
Graphical representation of the rickettsial rate of intracellular growth in XTC-2 cells. Amphibian host cells incubated in 12-well plates and approaching confluence were infected with MOI 10 rickettsiae. Infected cells from culture wells were harvested in 24 h intervals. To demonstrate the growth kinetics of R. felis Danube, the approximate mean numbers of bacterial genome equivalents (GE) were calculated based on the amplification of the rpsL gene by qPCR. Standard errors of the mean were calculated from two biological replicates. For statistical analysis, one-way ANOVA was performed, followed by Dunnett’s multiple comparison test (** p < 0.01, **** p < 0.0001) (a). To confirm viability of rickettsiae, the estimated transcript numbers of the same gene were evaluated per well, as described in Materials and Methods. The red line represents nonlinear regression, Beta growth then decay (b).

Figure 5.
Agarose gel electrophoresis of PCR amplicons of the ompA gene and plasmids from R. felis Danube. The partial sequence of the gene ompA was amplified using R. felis-specific primers Rf190.1790fw and Rf190.2857rev (a). The pRF plasmid was detected using the primer pairs pRFa-pRFb (expected size 159 bp) and pRFc-pRFd (expected size 1342 bp), but not pRFδ (primers pRFa and pRFd; expected size 1168 bp) (b). M: molecular marker; 1: uninfected XTC-2 cells; 2: infected XTC-2 cells; 3: purified R. felis Danube; 4: negative control; 5: infected XTC-2 cells; 6: purified R. felis Danube; 7: extracted plasmid DNA from purified R. felis Danube; 8: negative control.
Figure 5.
Agarose gel electrophoresis of PCR amplicons of the ompA gene and plasmids from R. felis Danube. The partial sequence of the gene ompA was amplified using R. felis-specific primers Rf190.1790fw and Rf190.2857rev (a). The pRF plasmid was detected using the primer pairs pRFa-pRFb (expected size 159 bp) and pRFc-pRFd (expected size 1342 bp), but not pRFδ (primers pRFa and pRFd; expected size 1168 bp) (b). M: molecular marker; 1: uninfected XTC-2 cells; 2: infected XTC-2 cells; 3: purified R. felis Danube; 4: negative control; 5: infected XTC-2 cells; 6: purified R. felis Danube; 7: extracted plasmid DNA from purified R. felis Danube; 8: negative control.

Figure 6.
Detection of R. felis Danube by immunofluorescence assay. Infected XTC-2 and Vero cells were fixed with 4% paraformaldehyde in PHEM buffer, and rickettsiae were labeled with rabbit polyclonal antibody against R. conorii, followed by rhodamine-conjugated goat anti-rabbit antibody (red signal). Actin filaments of host cells were labeled with conjugated phalloidin (green signal), and cell nuclei were stained with DAPI (blue signal). Uninfected cells treated with both primary and secondary antibodies were used as negative controls (scale bar 10 µm).
Figure 6.
Detection of R. felis Danube by immunofluorescence assay. Infected XTC-2 and Vero cells were fixed with 4% paraformaldehyde in PHEM buffer, and rickettsiae were labeled with rabbit polyclonal antibody against R. conorii, followed by rhodamine-conjugated goat anti-rabbit antibody (red signal). Actin filaments of host cells were labeled with conjugated phalloidin (green signal), and cell nuclei were stained with DAPI (blue signal). Uninfected cells treated with both primary and secondary antibodies were used as negative controls (scale bar 10 µm).

Figure 7.
Visualization of purified R. felis Danube by electron microscopy. Negative staining of rickettsial binary fission captured by transmission electron microscopy (a). Scanning electron microscopy of purified and glutaraldehyde-fixed R. felis sedimented onto circular coverslips and silicon wafer (b).
Figure 7.
Visualization of purified R. felis Danube by electron microscopy. Negative staining of rickettsial binary fission captured by transmission electron microscopy (a). Scanning electron microscopy of purified and glutaraldehyde-fixed R. felis sedimented onto circular coverslips and silicon wafer (b).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Rickettsia felis culture-isolated strains.
| Strain | Year | Origin | Cell Culture | Reference |
|---|---|---|---|---|
| URRWXCal2 | 2001 | Cat flea of the colony maintained by Flea Data, Inc., in the United States | Xenopus laevis cell line XTC-2 (28 °C) | Raoult D. et al. Emerg Infect Dis 7(1): 73–81 [51] |
| Pedreira | 2006 | Cat flea collected from a naturally infested farm dog, in Brazil | Aedes albopictus cell line C6/36 (25 °C) | Horta M.C. et al. Appl Environ Microbiol 72(2): 1705–1707 [57] |
| LSU | 2006 | Cat flea of the laboratory colony maintained at the Louisiana State University, in the United States | Ixodes scapularis cell line ISE6 (32 °C) | Pornwiroon W. et al. Appl Environ Microbiol 72(8): 5589–5595 [55] |
| LSU-Lb | 2011 | Booklouse Liposcelis bostrychophila from shredded corn cobs, in the United States | Ixodes scapularis cell line ISE6 (32 °C) | Thepparit C. et al. PLoS ONE 6(1): e16396 [56] |
| CfCR(SJ) | 2011 | Cat flea collected from a domestic dog, in Costa Rica | Aedes albopictus cell line C6/36 (28 °C) | Hun L. et al. Vector Borne Zoonotic Dis 11(10): 1395–1397 [58] |
| N/A * | 2013 | Cat flea collected from a pound dog, and the flea laboratory colony maintained at the University of Queensland, in Australia | Xenopus laevis cell line XTC-2 (28 °C) | Hii S.F. et al. Parasit Vectors 6:159 [53] |
| N/A * | 2020 | Domestic dog infected with R. felis in a laboratory setting, in Australia | Xenopus laevis cell line XTC-2 (28 °C) | Ng-Nguyen D. et al. Sci Rep 10: 4151 [54] |
| Danube | This study | Questing Ixodes ricinus nymph, in Slovakia | Xenopus laevis cell line XTC-2 (28 °C) | This study |
* N/A—not available.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Danchenko, M.; Benada, O.; Škultéty, Ľ.; Sekeyová, Z. Culture Isolate of Rickettsia felis from a Tick. Int. J. Environ. Res. Public Health 2022, 19, 4321. https://doi.org/10.3390/ijerph19074321
AMA Style
Danchenko M, Benada O, Škultéty Ľ, Sekeyová Z. Culture Isolate of Rickettsia felis from a Tick. International Journal of Environmental Research and Public Health. 2022; 19(7):4321. https://doi.org/10.3390/ijerph19074321
Chicago/Turabian StyleDanchenko, Monika, Oldřich Benada, Ľudovít Škultéty, and Zuzana Sekeyová. 2022. "Culture Isolate of Rickettsia felis from a Tick" International Journal of Environmental Research and Public Health 19, no. 7: 4321. https://doi.org/10.3390/ijerph19074321
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.