
The Ameliorating Effects of n-3 Polyunsaturated Fatty Acids on Liver Steatosis Induced by a High-Fat Methionine Choline-Deficient Diet in Mice
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
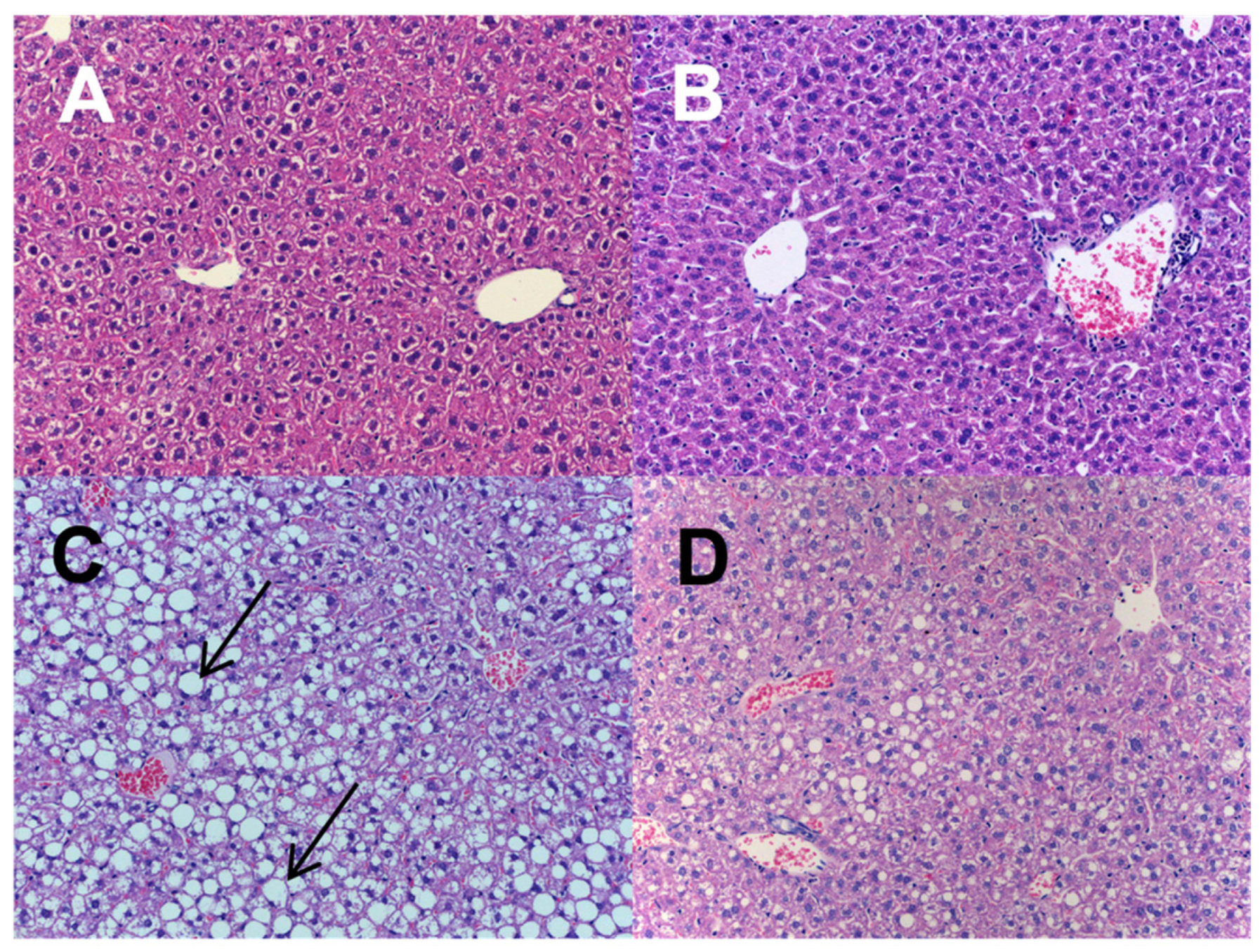
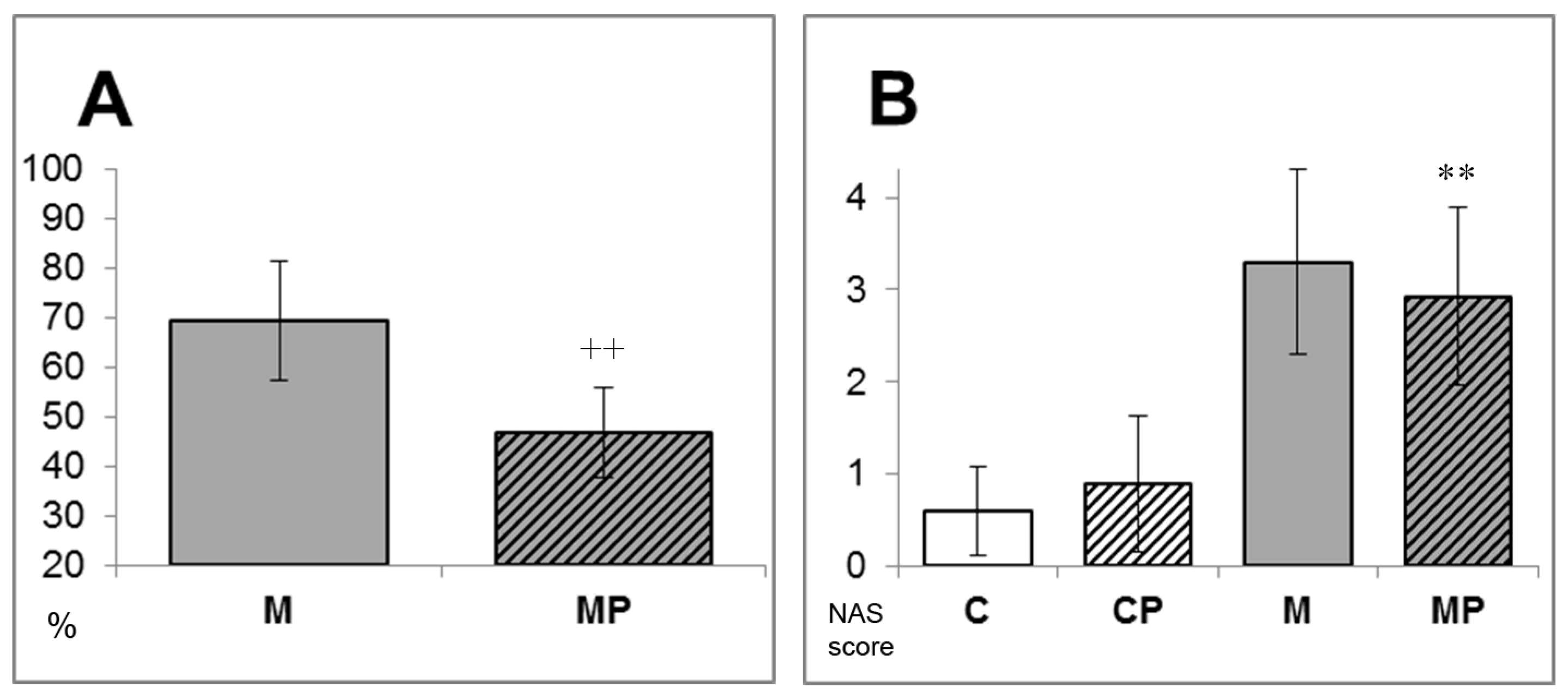
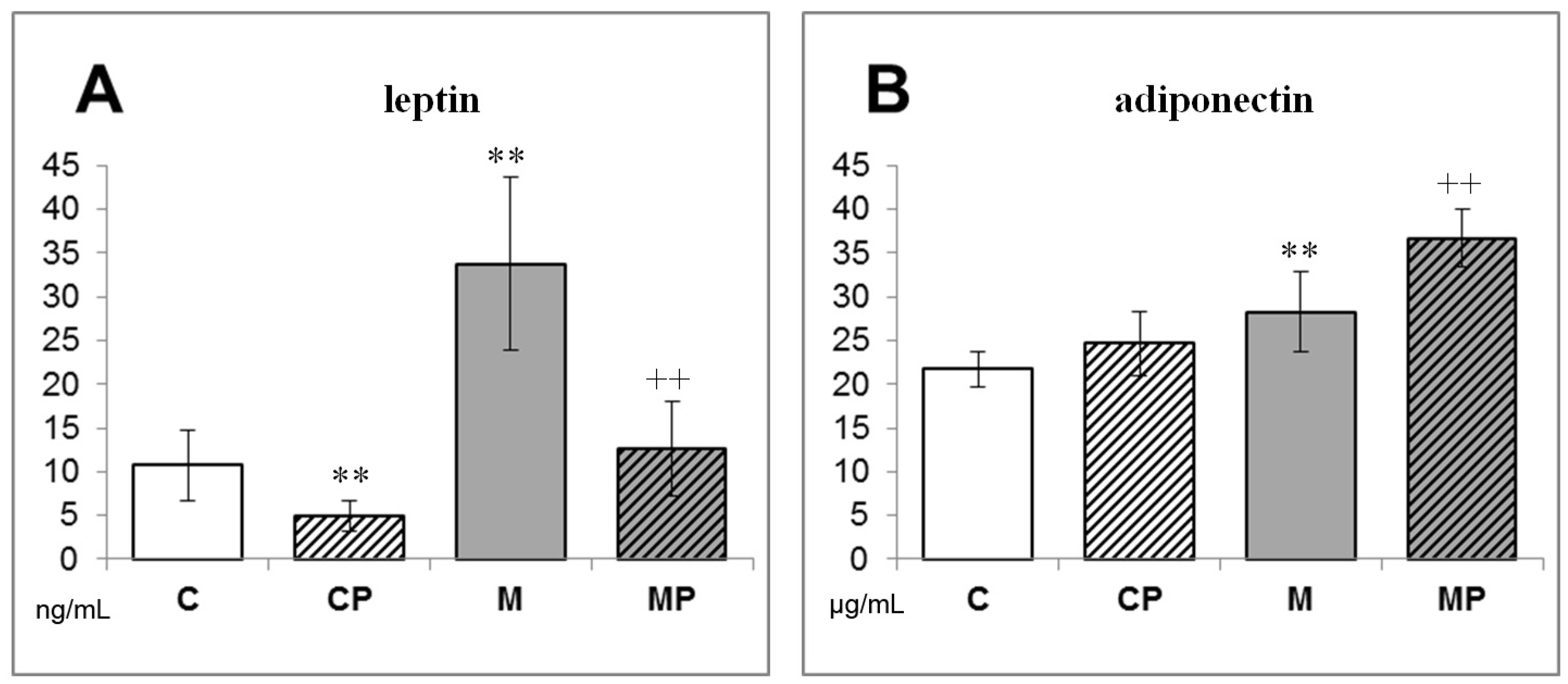
2.1. The Effect of n-3-PUFAs on NAFLD Development
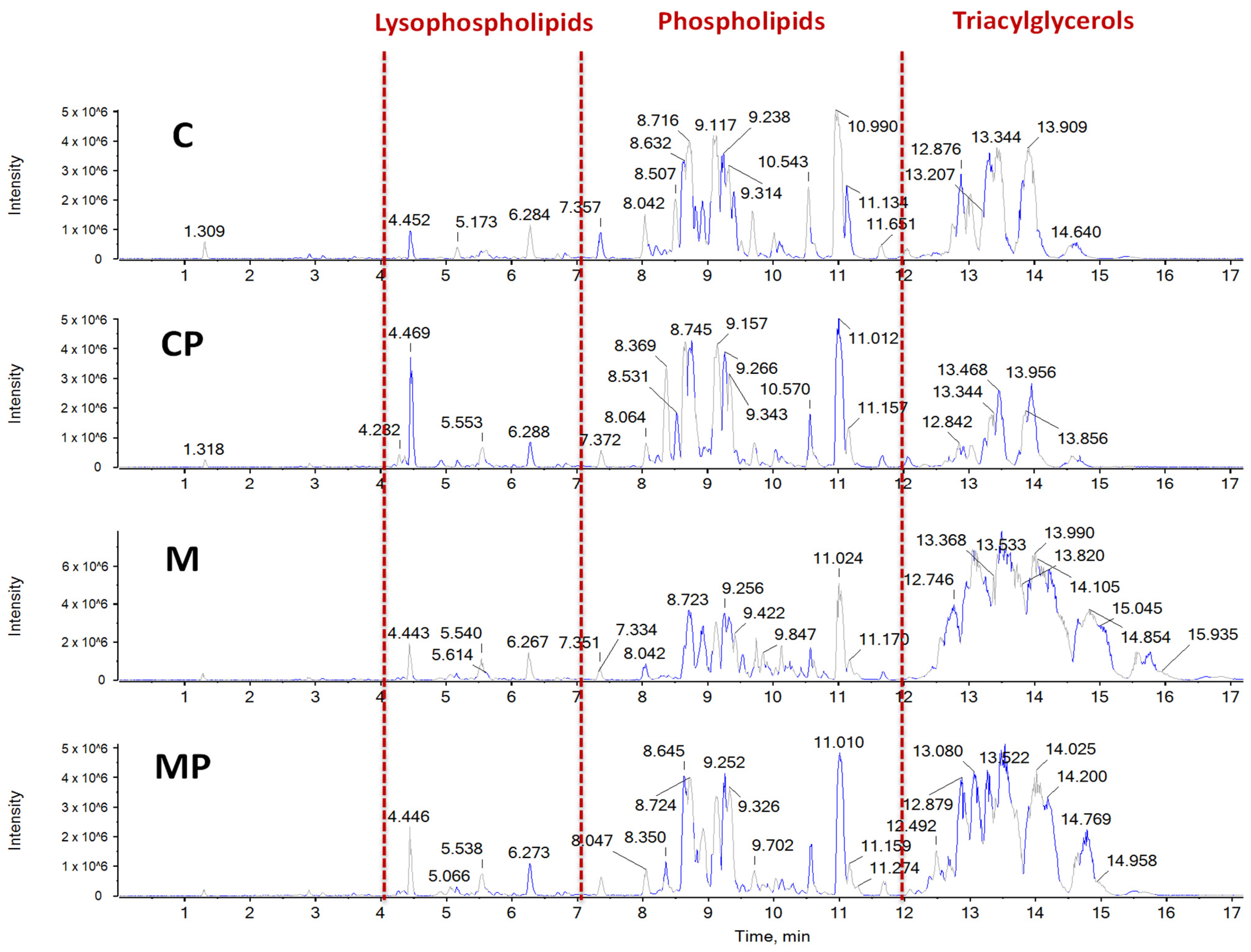
2.2. The Effect of n-3-PUFAs on Liver Lipidome
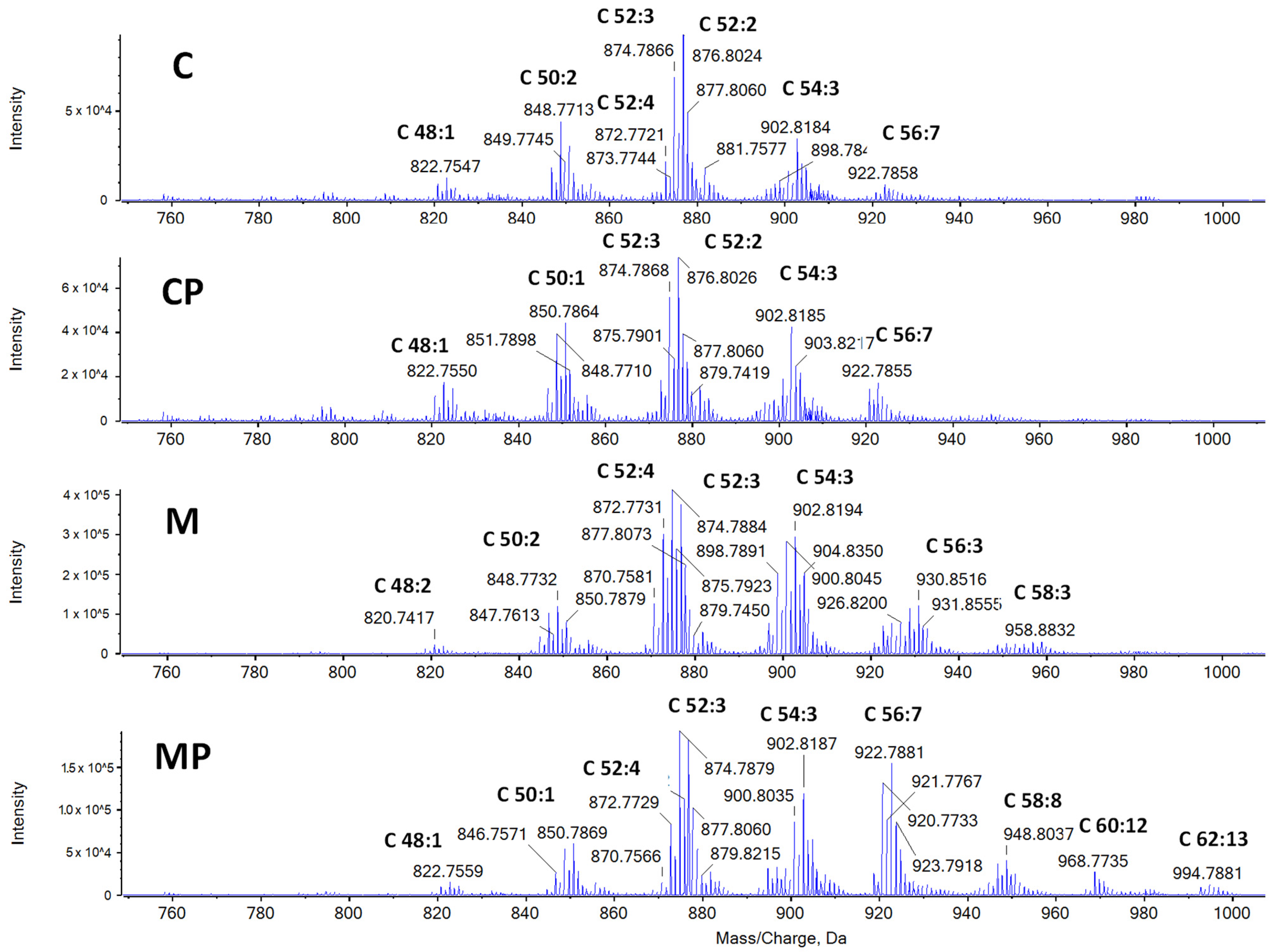
2.2.1. The Effect of n-3-PUFAs on Liver TAGs
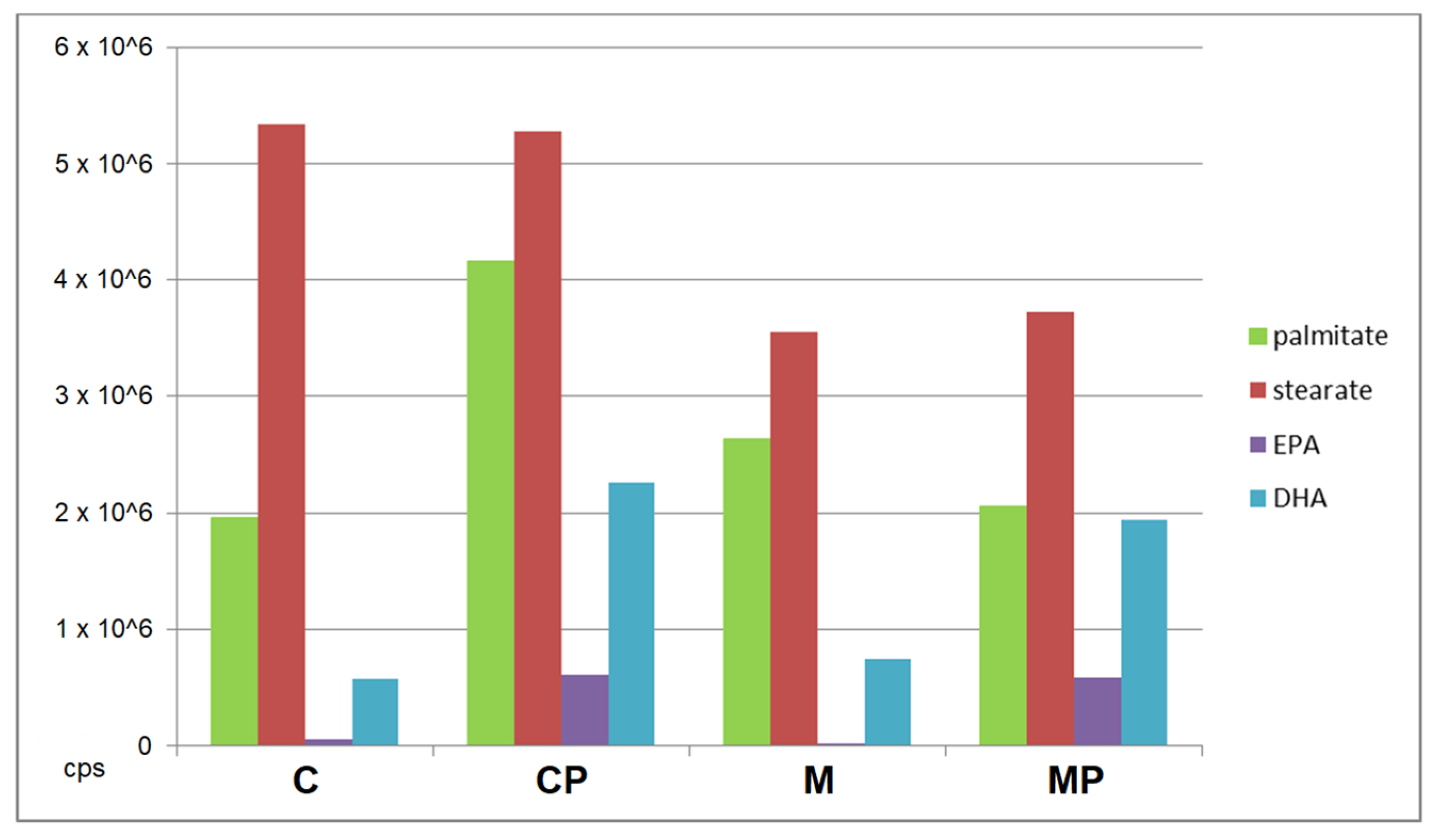
2.2.2. The Effect of n-3-PUFAs on Liver Free FAs
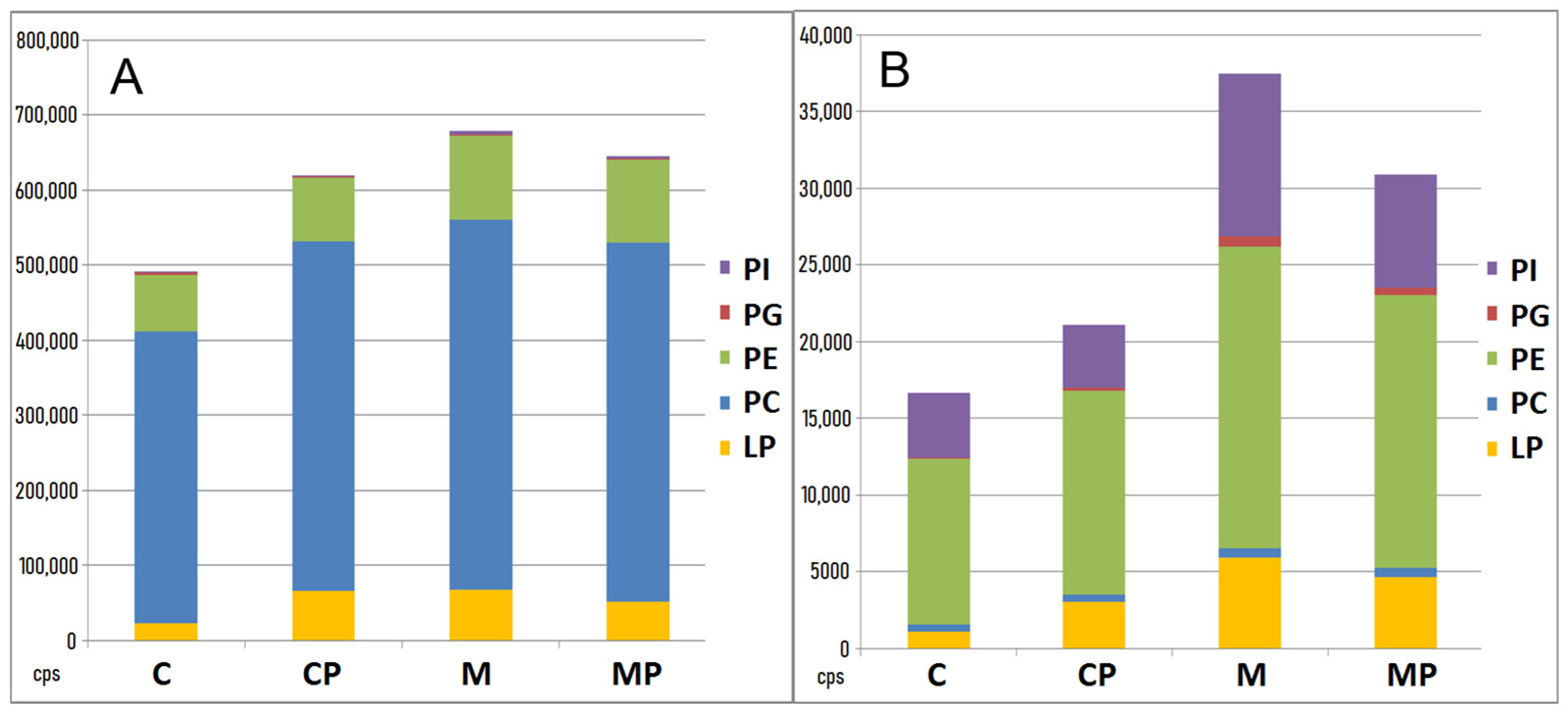
2.2.3. The Effect of n-3-PUFAs on Liver Phospholipids
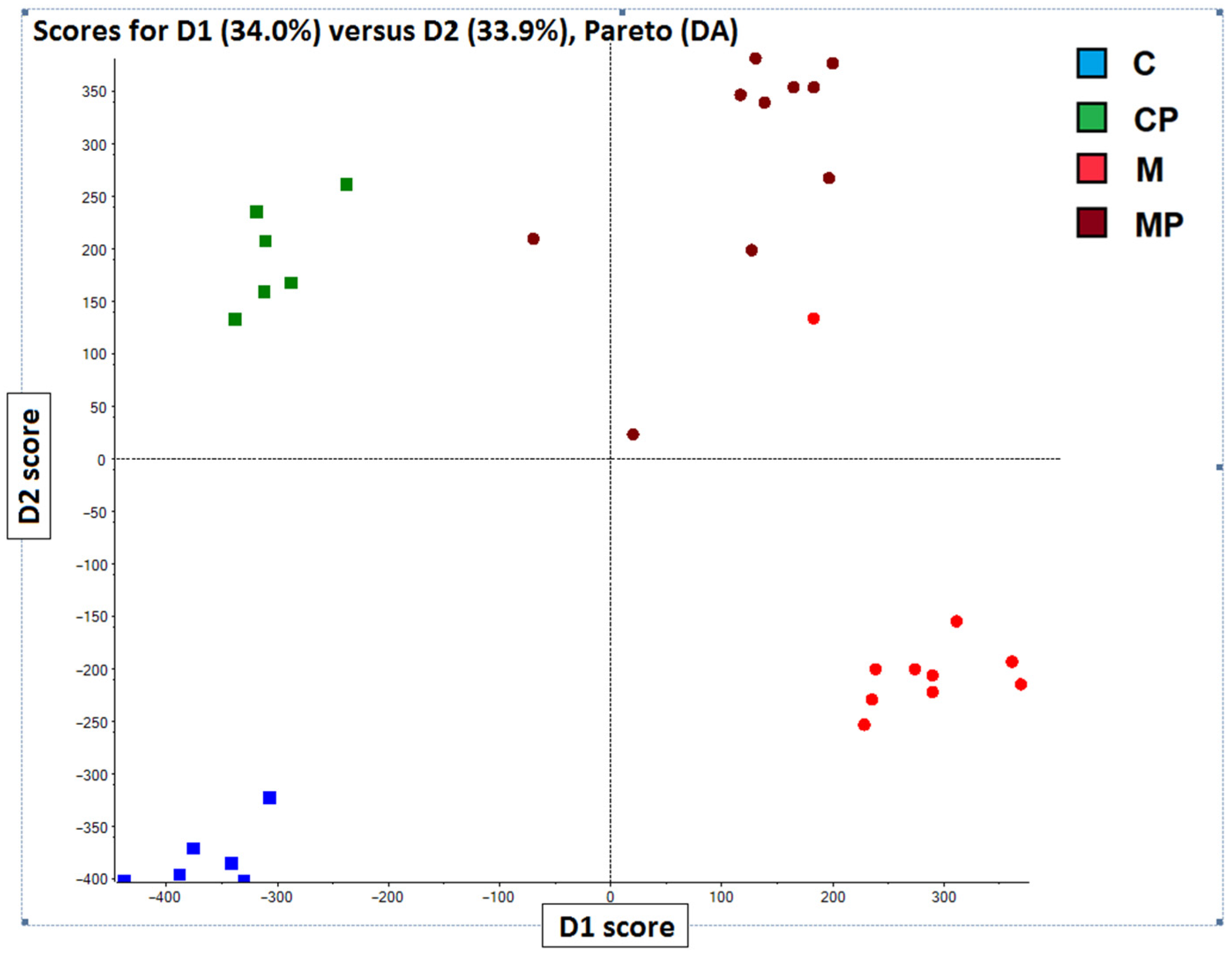
2.3. Chemometric Analysis PCA-DA of Chromatographic and Spectral Lipidomic Datasets Generated by Metabolic Fingerprinting
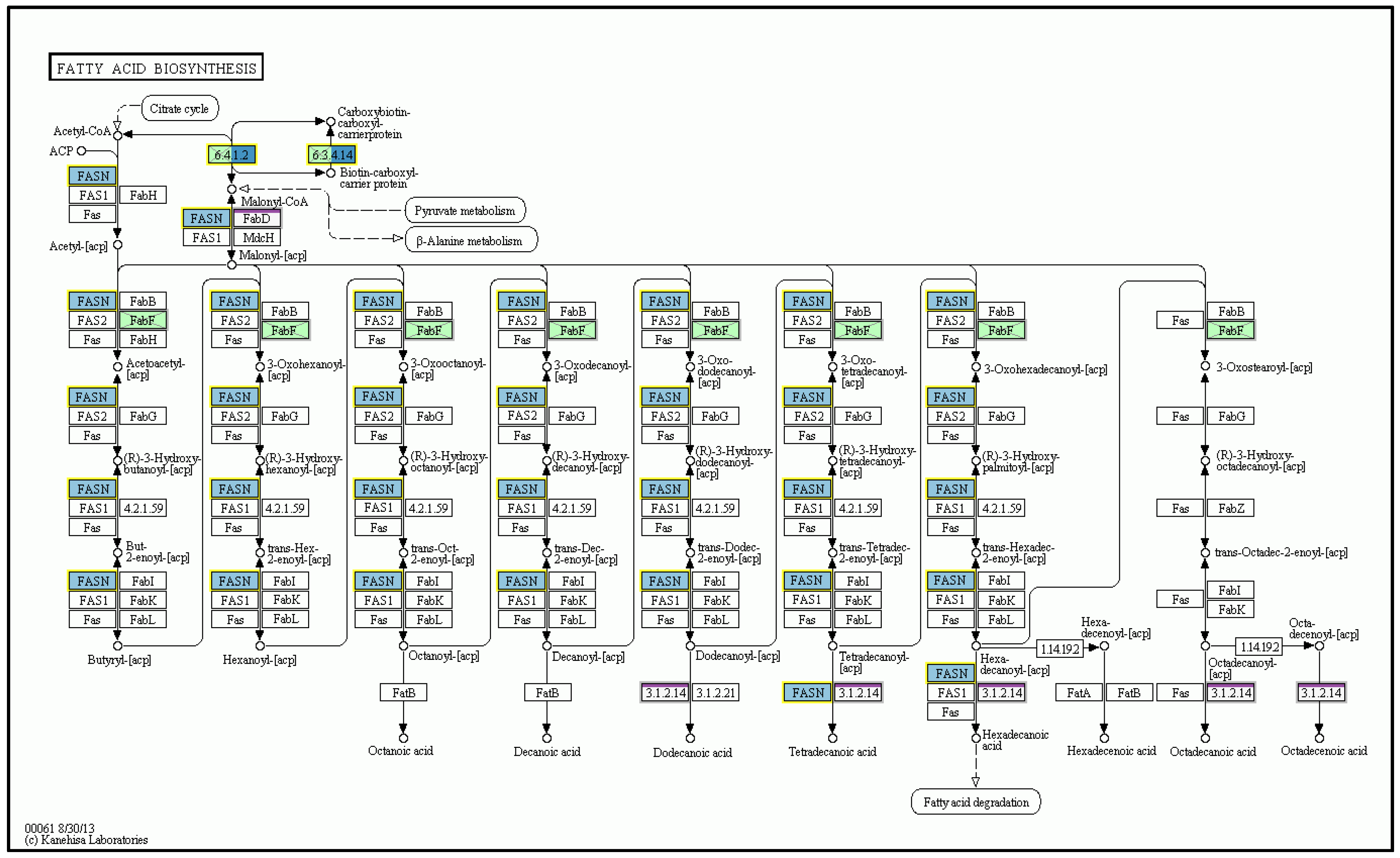
2.4. The Effect of n-3-PUFAs on Expression of Specific Genes in the Liver Tissue
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Experimental Design
4.3. Induction of Liver Steatosis
4.4. N-3-PUFA Administration
4.5. General Tissue Preparation
4.6. Serum Biochemistry
4.7. Liver Histology
4.8. Determination of Adipokines
4.9. Serum FA Composition
4.10. Lipidomic Analysis
4.11. Transcription Profiling of the Liver Tissues
4.12. Statistical Analysis and Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Marra, F.; Svegliati-Baroni, G. Lipotoxicity and the gut-liver axis in NASH pathogenesis. J. Hepatol. 2018, 68, 280–295. [Google Scholar] [CrossRef]
- Allard, J.P.; Aghdassi, E.; Mohammed, S.; Raman, M.; Avand, G.; Arendt, B.M.; Jalali, P.; Kandasamy, T.; Prayitno, N.; Sherman, M.; et al. Nutritional assessment and hepatic fatty acid composition in non-alcoholic fatty liver disease (NAFLD): A cross-sectional study. J. Hepatol. 2008, 48, 300–307. [Google Scholar] [CrossRef]
- Araya, J.; Rodrigo, R.; Videla, L.A.; Thielemann, L.; Orellana, M.; Pettinelli, P.; Poniachik, J. Increase in long-chain polyunsaturated fatty acid n-6/n-3 ratio in relation to hepatic steatosis in patients with non-alcoholic fatty liver disease. Clin. Sci. 2004, 106, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Gentile, C.L.; Pagliassotti, M.J. The role of fatty acids in the development and progression of nonalcoholic fatty liver disease. J. Nutr. Biochem. 2008, 19, 567–576. [Google Scholar] [CrossRef]
- Day, C.P.; James, O.F. Steatohepatitis: A tale of two “hits”? Gastroenterology 1998, 114, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Wanless, I.R.; Lentz, J.S. Fatty liver hepatitis (steatohepatitis) and obesity: An autopsy study with analysis of risk factors. Hepatology 1990, 12, 1106–1110. [Google Scholar] [CrossRef] [PubMed]
- Koek, G.H.; Liedorp, P.R.; Bast, A. The role of oxidative stress in non-alcoholic steatohepatitis. Clin. Chim. Acta 2011, 412, 1297–1305. [Google Scholar] [CrossRef]
- Ibrahim, S.H.; Gores, G.J. Who pulls the trigger: JNK activation in liver lipotoxicity? J. Hepatol. 2012, 56, 17–19. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; De Michieli, F.; Cassader, M.; Rizzetto, M.; Durazzo, M.; Faga, E.; Silli, B.; Pagano, G. Dietary habits and their relations to insulin resistance and postprandial lipemia in nonalcoholic steatohepatitis. Hepatology 2003, 37, 909–916. [Google Scholar] [CrossRef]
- Videla, L.A.; Hernandez-Rodas, M.C.; Metherel, A.H.; Valenzuela, R. Influence of the nutritional status and oxidative stress in the desaturation and elongation of n-3 and n-6 polyunsaturated fatty acids: Impact on non-alcoholic fatty liver disease. Prostaglandins Leukot. Essent. Fatty Acids 2022, 181, 102441. [Google Scholar] [CrossRef]
- Malinska, H.; Oliyarnyk, O.; Hubova, M.; Zidek, V.; Landa, V.; Simakova, M.; Mlejnek, P.; Kazdova, L.; Kurtz, T.W.; Pravenec, M. Increased liver oxidative stress and altered PUFA metabolism precede development of non-alcoholic steatohepatitis in SREBP-1a transgenic spontaneously hypertensive rats with genetic predisposition to hepatic steatosis. Mol. Cell. Biochem. 2010, 335, 119–125. [Google Scholar] [CrossRef]
- Xin, Y.N.; Xuan, S.Y.; Zhang, J.H.; Zheng, M.H.; Guan, H.S. Omega-3 polyunsaturated fatty acids: A specific liver drug for non-alcoholic fatty liver disease (NAFLD). Med. Hypotheses 2008, 71, 820–821. [Google Scholar] [CrossRef]
- Di Minno, M.N.; Russolillo, A.; Lupoli, R.; Ambrosino, P.; Di Minno, A.; Tarantino, G. Omega-3 fatty acids for the treatment of non-alcoholic fatty liver disease. World J. Gastroenterol. 2012, 18, 5839–5847. [Google Scholar] [CrossRef]
- Sekiya, M.; Yahagi, N.; Matsuzaka, T.; Najima, Y.; Nakakuki, M.; Nagai, R.; Ishibashi, S.; Osuga, J.; Yamada, N.; Shimano, H. Polyunsaturated fatty acids ameliorate hepatic steatosis in obese mice by SREBP-1 suppression. Hepatology 2003, 38, 1529–1539. [Google Scholar] [CrossRef]
- Kuda, O.; Jelenik, T.; Jilkova, Z.; Flachs, P.; Rossmeisl, M.; Hensler, M.; Kazdova, L.; Ogston, N.; Baranowski, M.; Gorski, J.; et al. n-3 fatty acids and rosiglitazone improve insulin sensitivity through additive stimulatory effects on muscle glycogen synthesis in mice fed a high-fat diet. Diabetologia 2009, 52, 941–951. [Google Scholar] [CrossRef] [PubMed]
- EFSA Assesses Safety of Long-Chain Omega-3 Fatty Acids. Available online: https://www.efsa.europa.eu/en/press/news/120727 (accessed on 27 July 2012).
- El-Badry, A.M.; Graf, R.; Clavien, P.A. Omega 3-Omega 6: What is right for the liver? J. Hepatol. 2007, 47, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Jump, D.B.; Lytle, K.A.; Depner, C.M.; Tripathy, S. Omega-3 polyunsaturated fatty acids as a treatment strategy for nonalcoholic fatty liver disease. Pharmacol. Ther. 2018, 181, 108–125. [Google Scholar] [CrossRef] [PubMed]
- Khadge, S.; Sharp, J.G.; Thiele, G.M.; McGuire, T.R.; Klassen, L.W.; Duryee, M.J.; Britton, H.C.; Dafferner, A.J.; Beck, J.; Black, P.N.; et al. Dietary omega-3 and omega-6 polyunsaturated fatty acids modulate hepatic pathology. J. Nutr. Biochem. 2018, 52, 92–102. [Google Scholar] [CrossRef]
- Kang, J.X. Balance of omega-6/omega-3 essential fatty acids is important for health. The evidence from gene transfer studies. World Rev. Nutr. Diet. 2005, 95, 93–102. [Google Scholar]
- Salehi-Sahlabadi, A.; Sadat, S.; Beigrezaei, S.; Pourmasomi, M.; Feizi, A.; Ghiasvand, R.; Hadi, A.; Clark, C.C.T.; Miraghajani, M. Dietary patterns and risk of non-alcoholic fatty liver disease. BMC Gastroenterol. 2021, 21, 41. [Google Scholar] [CrossRef]
- Rossmeisl, M.; Jilkova, Z.M.; Kuda, O.; Jelenik, T.; Medrikova, D.; Stankova, B.; Kristinsson, B.; Haraldsson, G.G.; Svensen, H.; Stoknes, I.; et al. Metabolic effects of n-3 PUFA as phospholipids are superior to triglycerides in mice fed a high-fat diet: Possible role of endocannabinoids. PLoS ONE 2012, 7, e38834. [Google Scholar] [CrossRef] [PubMed]
- Roche, H.M.; Gibney, M.J. Effect of long-chain n-3 polyunsaturated fatty acids on fasting and postprandial triacylglycerol metabolism. Am. J. Clin. Nutr. 2000, 71, 232S–237S. [Google Scholar] [CrossRef] [PubMed]
- Itariu, B.K.; Zeyda, M.; Hochbrugger, E.E.; Neuhofer, A.; Prager, G.; Schindler, K.; Bohdjalian, A.; Mascher, D.; Vangala, S.; Schranz, M.; et al. Long-chain n-3 PUFAs reduce adipose tissue and systemic inflammation in severely obese nondiabetic patients: A randomized controlled trial. Am. J. Clin. Nutr. 2012, 96, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, J.A.; Katz, R. n-3 long-chain polyunsaturated fatty acids in type 2 diabetes: A review. J. Am. Diet. Assoc. 2005, 105, 428–440. [Google Scholar] [CrossRef]
- Flachs, P.; Rossmeisl, M.; Bryhn, M.; Kopecky, J. Cellular and molecular effects of n-3 polyunsaturated fatty acids on adipose tissue biology and metabolism. Clin. Sci. 2009, 116, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Flachs, P.; Mohamed-Ali, V.; Horakova, O.; Rossmeisl, M.; Hosseinzadeh-Attar, M.J.; Hensler, M.; Ruzickova, J.; Kopecky, J. Polyunsaturated fatty acids of marine origin induce adiponectin in mice fed a high-fat diet. Diabetologia 2006, 49, 394–397. [Google Scholar] [CrossRef]
- Sanderson, L.M.; de Groot, P.J.; Hooiveld, G.J.; Koppen, A.; Kalkhoven, E.; Muller, M.; Kersten, S. Effect of synthetic dietary triglycerides: A novel research paradigm for nutrigenomics. PLoS ONE 2008, 3, e1681. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Martinez-Arranz, I.; Bruzzone, C.; Noureddin, M.; Gil-Redondo, R.; Minchole, I.; Bizkarguenaga, M.; Arretxe, E.; Iruarrizaga-Lejarreta, M.; Fernandez-Ramos, D.; Lopitz-Otsoa, F.; et al. Metabolic subtypes of patients with NAFLD exhibit distinctive cardiovascular risk profiles. Hepatology 2022, 76, 1121–1134. [Google Scholar] [CrossRef]
- Mato, J.M.; Alonso, C.; Noureddin, M.; Lu, S.C. Biomarkers and subtypes of deranged lipid metabolism in non-alcoholic fatty liver disease. World J. Gastroenterol. 2019, 25, 3009–3020. [Google Scholar] [CrossRef]
- Anjani, K.; Lhomme, M.; Sokolovska, N.; Poitou, C.; Aron-Wisnewsky, J.; Bouillot, J.L.; Lesnik, P.; Bedossa, P.; Kontush, A.; Clement, K.; et al. Circulating phospholipid profiling identifies portal contribution to NASH signature in obesity. J. Hepatol. 2015, 62, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.W.; Arendt, B.M.; Hillyer, L.M.; Fung, S.K.; McGilvray, I.; Guindi, M.; Allard, J.P. Plasma phospholipids and fatty acid composition differ between liver biopsy-proven nonalcoholic fatty liver disease and healthy subjects. Nutr. Diabetes 2016, 6, e220. [Google Scholar] [CrossRef] [PubMed]
- Puri, P.; Baillie, R.A.; Wiest, M.M.; Mirshahi, F.; Choudhury, J.; Cheung, O.; Sargeant, C.; Contos, M.J.; Sanyal, A.J. A lipidomic analysis of nonalcoholic fatty liver disease. Hepatology 2007, 46, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Parker, H.M.; Johnson, N.A.; Burdon, C.A.; Cohn, J.S.; O’Connor, H.T.; George, J. Omega-3 supplementation and non-alcoholic fatty liver disease: A systematic review and meta-analysis. J. Hepatol. 2012, 56, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Oddy, W.H.; Herbison, C.E.; Jacoby, P.; Ambrosini, G.L.; O’Sullivan, T.A.; Ayonrinde, O.T.; Olynyk, J.K.; Black, L.J.; Beilin, L.J.; Mori, T.A.; et al. The Western dietary pattern is prospectively associated with nonalcoholic fatty liver disease in adolescence. Am. J. Gastroenterol. 2013, 108, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Faber, K.N.; de Meijer, V.E.; Blokzijl, H.; Moshage, H. How does hepatic lipid accumulation lead to lipotoxicity in non-alcoholic fatty liver disease? Hepatol. Int. 2021, 15, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Rada, P.; Gonzalez-Rodriguez, A.; Garcia-Monzon, C.; Valverde, A.M. Understanding lipotoxicity in NAFLD pathogenesis: Is CD36 a key driver? Cell Death Dis. 2020, 11, 802. [Google Scholar] [CrossRef]
- Pei, K.; Gui, T.; Kan, D.; Feng, H.; Jin, Y.; Yang, Y.; Zhang, Q.; Du, Z.; Gai, Z.; Wu, J.; et al. An Overview of Lipid Metabolism and Nonalcoholic Fatty Liver Disease. Biomed. Res. Int. 2020, 2020, 4020249. [Google Scholar] [CrossRef]
- Okada, L.S.; Oliveira, C.P.; Stefano, J.T.; Nogueira, M.A.; Silva, I.; Cordeiro, F.B.; Alves, V.A.F.; Torrinhas, R.S.; Carrilho, F.J.; Puri, P.; et al. Omega-3 PUFA modulate lipogenesis, ER stress, and mitochondrial dysfunction markers in NASH—Proteomic and lipidomic insight. Clin. Nutr. 2018, 37, 1474–1484. [Google Scholar] [CrossRef]
- Yonezawa, Y.; Hada, T.; Uryu, K.; Tsuzuki, T.; Eitsuka, T.; Miyazawa, T.; Murakami-Nakai, C.; Yoshida, H.; Mizushina, Y. Inhibitory effect of conjugated eicosapentaenoic acid on mammalian DNA polymerase and topoisomerase activities and human cancer cell proliferation. Biochem. Pharmacol. 2005, 70, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Chapkin, R.S.; Davidson, L.A.; Ly, L.; Weeks, B.R.; Lupton, J.R.; McMurray, D.N. Immunomodulatory effects of (n-3) fatty acids: Putative link to inflammation and colon cancer. J. Nutr. 2007, 137, 200S–204S. [Google Scholar] [CrossRef] [PubMed]
- Kalas, M.A.; Chavez, L.; Leon, M.; Taweesedt, P.T.; Surani, S. Abnormal liver enzymes: A review for clinicians. World J. Hepatol. 2021, 13, 1688–1698. [Google Scholar] [CrossRef] [PubMed]
- Videla, L.A.; Rodrigo, R.; Araya, J.; Poniachik, J. Insulin resistance and oxidative stress interdependency in non-alcoholic fatty liver disease. Trends Mol. Med. 2006, 12, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, R.; Espinosa, A.; Gonzalez-Manan, D.; D’Espessailles, A.; Fernandez, V.; Videla, L.A.; Tapia, G. N-3 long-chain polyunsaturated fatty acid supplementation significantly reduces liver oxidative stress in high fat induced steatosis. PLoS ONE 2012, 7, e46400. [Google Scholar] [CrossRef] [PubMed]
- Aronis, A.; Madar, Z.; Tirosh, O. Mechanism underlying oxidative stress-mediated lipotoxicity: Exposure of J774.2 macrophages to triacylglycerols facilitates mitochondrial reactive oxygen species production and cellular necrosis. Free Radic. Biol Med 2005, 38, 1221–1230. [Google Scholar] [CrossRef]
- Zhou, Y.E.; Kubow, S.; Dewailly, E.; Julien, P.; Egeland, G.M. Decreased activity of desaturase 5 in association with obesity and insulin resistance aggravates declining long-chain n-3 fatty acid status in Cree undergoing dietary transition. Br. J. Nutr. 2009, 102, 888–894. [Google Scholar] [CrossRef]
- Pettinelli, P.; Del Pozo, T.; Araya, J.; Rodrigo, R.; Araya, A.V.; Smok, G.; Csendes, A.; Gutierrez, L.; Rojas, J.; Korn, O.; et al. Enhancement in liver SREBP-1c/PPAR-alpha ratio and steatosis in obese patients: Correlations with insulin resistance and n-3 long-chain polyunsaturated fatty acid depletion. Biochim. Biophys. Acta 2009, 1792, 1080–1086. [Google Scholar] [CrossRef]
- Elizondo, A.; Araya, J.; Rodrigo, R.; Signorini, C.; Sgherri, C.; Comporti, M.; Poniachik, J.; Videla, L.A. Effects of weight loss on liver and erythrocyte polyunsaturated fatty acid pattern and oxidative stress status in obese patients with non-alcoholic fatty liver disease. Biol. Res. 2008, 41, 59–68. [Google Scholar] [CrossRef]
- Duan, Y.; Li, F.; Li, L.; Fan, J.; Sun, X.; Yin, Y. n-6:n-3 PUFA ratio is involved in regulating lipid metabolism and inflammation in pigs. Br. J. Nutr. 2014, 111, 445–451. [Google Scholar] [CrossRef]
- Dasilva, G.; Pazos, M.; Garcia-Egido, E.; Gallardo, J.M.; Rodriguez, I.; Cela, R.; Medina, I. Healthy effect of different proportions of marine omega-3 PUFAs EPA and DHA supplementation in Wistar rats: Lipidomic biomarkers of oxidative stress and inflammation. J. Nutr. Biochem. 2015, 26, 1385–1392. [Google Scholar] [CrossRef]
- Ferramosca, A.; Conte, A.; Burri, L.; Berge, K.; De Nuccio, F.; Giudetti, A.M.; Zara, V. A krill oil supplemented diet suppresses hepatic steatosis in high-fat fed rats. PLoS ONE 2012, 7, e38797. [Google Scholar] [CrossRef] [PubMed]
- Depner, C.M.; Philbrick, K.A.; Jump, D.B. Docosahexaenoic acid attenuates hepatic inflammation, oxidative stress, and fibrosis without decreasing hepatosteatosis in a Ldlr(-/-) mouse model of western diet-induced nonalcoholic steatohepatitis. J. Nutr. 2013, 143, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Gorden, D.L.; Myers, D.S.; Ivanova, P.T.; Fahy, E.; Maurya, M.R.; Gupta, S.; Min, J.; Spann, N.J.; McDonald, J.G.; Kelly, S.L.; et al. Biomarkers of NAFLD progression: A lipidomics approach to an epidemic. J. Lipid Res. 2015, 56, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Birerdinc, A.; Younossi, Z. Can NASH lipidome provide insight into the pathogenesis of obesity-related non-alcoholic fatty liver disease? J. Hepatol. 2015, 62, 761–762. [Google Scholar] [CrossRef]
- Mayo, R.; Crespo, J.; Martinez-Arranz, I.; Banales, J.M.; Arias, M.; Minchole, I.; Aller de la Fuente, R.; Jimenez-Aguero, R.; Alonso, C.; de Luis, D.A.; et al. Metabolomic-based noninvasive serum test to diagnose nonalcoholic steatohepatitis: Results from discovery and validation cohorts. Hepatol. Commun. 2018, 2, 807–820. [Google Scholar] [CrossRef]
- Duan, J.; Song, Y.; Zhang, X.; Wang, C. Effect of omega-3 Polyunsaturated Fatty Acids-Derived Bioactive Lipids on Metabolic Disorders. Front. Physiol. 2021, 12, 646491. [Google Scholar] [CrossRef]
- Lamaziere, A.; Wolf, C.; Barbe, U.; Bausero, P.; Visioli, F. Lipidomics of hepatic lipogenesis inhibition by omega 3 fatty acids. Prostaglandins Leukot. Essent. Fatty Acids 2013, 88, 149–154. [Google Scholar] [CrossRef]
- Spahis, S.; Alvarez, F.; Dubois, J.; Ahmed, N.; Peretti, N.; Levy, E. Plasma fatty acid composition in French-Canadian children with non-alcoholic fatty liver disease: Effect of n-3 PUFA supplementation. Prostaglandins Leukot. Essent. Fatty Acids 2015, 99, 25–34. [Google Scholar] [CrossRef]
- Yea, K.; Kim, J.; Yoon, J.H.; Kwon, T.; Kim, J.H.; Lee, B.D.; Lee, H.J.; Lee, S.J.; Kim, J.I.; Lee, T.G.; et al. Lysophosphatidylcholine activates adipocyte glucose uptake and lowers blood glucose levels in murine models of diabetes. J. Biol. Chem. 2009, 284, 33833–33840. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Ohishi, T.; Matsui, T.; Saito, T.; Matsumoto, M.; Takasaki, J.; Matsumoto, S.; Kamohara, M.; Hiyama, H.; Yoshida, S.; et al. Lysophosphatidylcholine enhances glucose-dependent insulin secretion via an orphan G-protein-coupled receptor. Biochem. Biophys. Res. Commun. 2005, 326, 744–751. [Google Scholar] [CrossRef]
- Grzelczyk, A.; Gendaszewska-Darmach, E. Novel bioactive glycerol-based lysophospholipids: New data—New insight into their function. Biochimie 2013, 95, 667–679. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, J.C.; Massey, K.; Nicolaou, A. Fish oil supplementation alters levels of lipid mediators of inflammation in microenvironment of acute human wounds. Wound Repair. Regen. 2011, 19, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Brahmbhatt, V.; Oliveira, M.; Briand, M.; Perrisseau, G.; Bastic Schmid, V.; Destaillats, F.; Pace-Asciak, C.; Benyacoub, J.; Bosco, N. Protective effects of dietary EPA and DHA on ischemia-reperfusion-induced intestinal stress. J. Nutr. Biochem. 2013, 24, 104–111. [Google Scholar] [CrossRef] [PubMed]
- de Castro, G.S.; Calder, P.C. Non-alcoholic fatty liver disease and its treatment with n-3 polyunsaturated fatty acids. Clin. Nutr. 2018, 37, 37–55. [Google Scholar] [CrossRef]
- Shang, T.; Liu, L.; Zhou, J.; Zhang, M.; Hu, Q.; Fang, M.; Wu, Y.; Yao, P.; Gong, Z. Protective effects of various ratios of DHA/EPA supplementation on high-fat diet-induced liver damage in mice. Lipids Health Dis. 2017, 16, 65. [Google Scholar] [CrossRef]
- Anstee, Q.M.; Goldin, R.D. Mouse models in non-alcoholic fatty liver disease and steatohepatitis research. Int. J. Exp. Pathol. 2006, 87, 1–16. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Carlson, L.A. Extraction of lipids from human whole serum and lipoproteins and from rat liver tissue with methylene chloride-methanol: A comparison with extraction with chloroform-methanol. Clin. Chim. Acta 1985, 149, 89–93. [Google Scholar] [CrossRef]
- Rubert, J.; Monforte, A.; Hurkova, K.; Perez-Martinez, G.; Blesa, J.; Navarro, J.L.; Stranka, M.; Soriano, J.M.; Hajslova, J. Untargeted metabolomics of fresh and heat treatment Tiger nut (Cyperus esculentus L.) milks reveals further insight into food quality and nutrition. J. Chromatogr. A 2017, 1514, 80–87. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | CP | M | MP | |
|---|---|---|---|---|
| body weight [g] | 31.6 ± 1.2 | 30.7 ± 2.4 | 34.9 ± 2.6 ** | 31.2 ± 1.2 ++ |
| liver weight [g] | 1.38 ± 0.14 | 1.45 ± 0.24 | 1.75 ± 0.14 ** | 1.39 ± 0.13 ++ |
| bilirubin [μmol/L] | 1.85 ± 0.65 | 1.30 ± 0.28 | 1.72 ± 0.5 | 1.79 ± 0.65 |
| ALT [μkat/L] | 0.64 ± 0.2 | 0.52 ± 0.12 | 1.29 ± 0.35 ** | 0.83 ± 0.15 + |
| AST [μkat/L] | 1.65 ± 0.37 | 1.08 ± 0.19 ** | 1.43 ± 0.33 | 1.38 ± 0.5 |
| ALP [μkat/L] | 0.73 ± 0.07 | 0.78 ± 0.13 | 0.95 ± 0.15 ** | 0.83 ± 0.15 |
| cholesterol [mmol/L] | 2.02 ± 0.12 | 1.49 ± 0.15 ** | 3.28 ± 0.62 ** | 2.36 ± 0.29 ++ |
| TAG [mmol/L] | 1.06 ± 0.06 | 0.86 ± 0.16 * | 0.85 ± 0.23 | 0.59 ± 0.16 + |
| total protein [g/L] | 44.6 ± 2.0 | 44.1 ± 1.9 | 46.5 ± 3.2 | 46.1 ± 2.5 |
| C | CP | M | MP | |
|---|---|---|---|---|
| sum of FAs [μmol/mL] | 7.26 ± 1.0 | 5.97 ± 0.4 * | 9.08 ± 1.4 * | 6.56 ± 1.4 ++ |
| saturated FAs [μmol/mL] | 2.36 ± 0.3 | 1.91 ± 0.1 ** | 2.47 ± 0.4 | 1.8 ± 0.3 ++ |
| monosaturated [μmol/mL] | 1.76 ± 0.3 | 1.51 ± 0.1 | 1.82 ± 0.3 | 1.07 ± 0.4 ++ |
| sum of n-6 [μmol/mL] | 2.63 ± 0.4 | 1.88 ± 0.1 ** | 4.13 ± 0.7 ** | 2.69 ± 0.6 ++ |
| sum of n-3 [μmol/mL] | 0.48 ± 0.1 | 0.65 ± 0.1 * | 0.28 ± 0.1 ** | 0.72 ± 0.1 ++ |
| n-6/n-3 ratio | 5.5 | 2.9 | 14.7 | 3.7 |
| total liver lipid [g] | 3.08 ± 0.36 | 3.08 ± 0.56 | 13.97 ± 3.36 ** | 8.29 ± 3.57 ++ |
| [μmol/mL] | C | CP | M | MP |
|---|---|---|---|---|
| 12:0 | 0.0062 ± 0.003 | 0.0075 ± 0.003 | 0.0062 ± 0.001 | 0.0067 ± 0.003 |
| 14:0 | 0.0283 ± 0.005 | 0.0273 ± 0.004 | 0.0205 ± 0.002 ** | 0.0216 ± 0.009 |
| 14:1n-5 | 0.0016 ± 0.001 | 0.0017 ± 0.001 | 0.0008 ± 0.0006 | 0.0011 ± 0.0005 |
| 16:0 | 1.681 ± 0.23 | 1.395 ± 0.09 * | 1.514 ± 0.23 | 1.089 ± 0.21 ++ |
| 16:1n-9 | 0.043 ± 0.006 | 0.036 ± 0.003 * | 0.029 ± 0.005 ** | 0.033 ± 0.032 |
| 16:1n-7 | 0.272 ± 0.048 | 0.243 ± 0.022 | 0.116 ± 0.029 ** | 0.074 ± 0.028 + |
| 18:0 | 0.625 ± 0.05 | 0.464 ± 0.03 ** | 0.911 ± 0.14 ** | 0.664 ± 0.1 ++ |
| 18:1trans | 0.022 ± 0.006 | 0.016 ± 0.005 | 0.374 ± 0.13 ** | 0.290 ± 0.12 |
| 18:1n-9c | 1.237 ± 0.22 | 1.052 ± 0.11 | 1.49 ± 0.21 | 0.869 ± 0.29 ++ |
| 18:1n-7c | 0.172 ± 0.03 | 0.147 ± 0.01 | 0.165 ± 0.03 | 0.08 ± 0.03 ++ |
| 18:2n6cc | 1.793 ± 0.26 | 1.474 ± 0.14 * | 3.0 ± 0.47 ** | 2.29 ± 0.59 + |
| 18:3n-6alc | 0.025 ± 0.004 | 0.014 ± 0.003 ** | 0.015 ± 0.002 ** | 0.009 ± 0.001 ++ |
| 18:3n-3alc | 0.039 ± 0.008 | 0.0317 ± 0.007 | 0.011 ± 0.002 ** | 0.008 ± 0.002 + |
| 20:0 | 0.023 ± 0.003 | 0.016 ± 0.003 * | 0.021 ± 0.004 | 0.016 ± 0.003 + |
| 20:1n-9c | 0.0375 ± 0.01 | 0.032 ± 0.007 | 0.021 ± 0.004 ** | 0.014 ± 0.005 + |
| 20:2n-6cc | 0.027 ± 0.005 | 0.023 ± 0.002 | 0.043 ± 0.008 ** | 0.012 ± 0.003 ++ |
| 20:3n-6alc | 0.095 ± 0.016 | 0.076 ± 0.01 * | 0.169 ± 0.037 ** | 0.056 ± 0.011 ++ |
| 20:4n-6alc | 0.668 ± 0.115 | 0.288 ± 0.046 ** | 0.869 ± 0.233 | 0.313 ± 0.131 ++ |
| 20:5n-3alc | 0.043 ± 0.008 | 0.117 ± 0.037 ** | 0.016 ± 0.006 ** | 0.195 ± 0.063 ++ |
| 22:4n-6alc | 0.011 ± 0.002 | 0.004 ± 0.001 ** | 0.012 ± 0.002 | 0.002 ± 0.002 ++ |
| 22:5n-6 | 0.012 ± 0.003 | 0.003 ± 0.001 ** | 0.022 ± 0.005 ** | 0.004 ± 0.002 ++ |
| 22:5n-3alc | 0.027 ± 0.004 | 0.05 ± 0.01 ** | 0.012 ± 0.007 ** | 0.038 ± 0.006 ++ |
| 22:6n-3alc | 0.37 ± 0.07 | 0.452 ± 0.05 * | 0.243 ± 0.09 ** | 0.478 ± 0.07 ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šmíd, V.; Dvořák, K.; Stehnová, K.; Strnad, H.; Rubert, J.; Stříteský, J.; Staňková, B.; Stránská, M.; Hajšlová, J.; Brůha, R.; et al. The Ameliorating Effects of n-3 Polyunsaturated Fatty Acids on Liver Steatosis Induced by a High-Fat Methionine Choline-Deficient Diet in Mice. Int. J. Mol. Sci. 2023, 24, 17226. https://doi.org/10.3390/ijms242417226
Šmíd V, Dvořák K, Stehnová K, Strnad H, Rubert J, Stříteský J, Staňková B, Stránská M, Hajšlová J, Brůha R, et al. The Ameliorating Effects of n-3 Polyunsaturated Fatty Acids on Liver Steatosis Induced by a High-Fat Methionine Choline-Deficient Diet in Mice. International Journal of Molecular Sciences. 2023; 24(24):17226. https://doi.org/10.3390/ijms242417226
Chicago/Turabian StyleŠmíd, Václav, Karel Dvořák, Kamila Stehnová, Hynek Strnad, Josep Rubert, Jan Stříteský, Barbora Staňková, Milena Stránská, Jana Hajšlová, Radan Brůha, and et al. 2023. "The Ameliorating Effects of n-3 Polyunsaturated Fatty Acids on Liver Steatosis Induced by a High-Fat Methionine Choline-Deficient Diet in Mice" International Journal of Molecular Sciences 24, no. 24: 17226. https://doi.org/10.3390/ijms242417226