
Filamentous Hemagglutinin of Bordetella pertussis Does Not Interact with the β2 Integrin CD11b/CD18

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
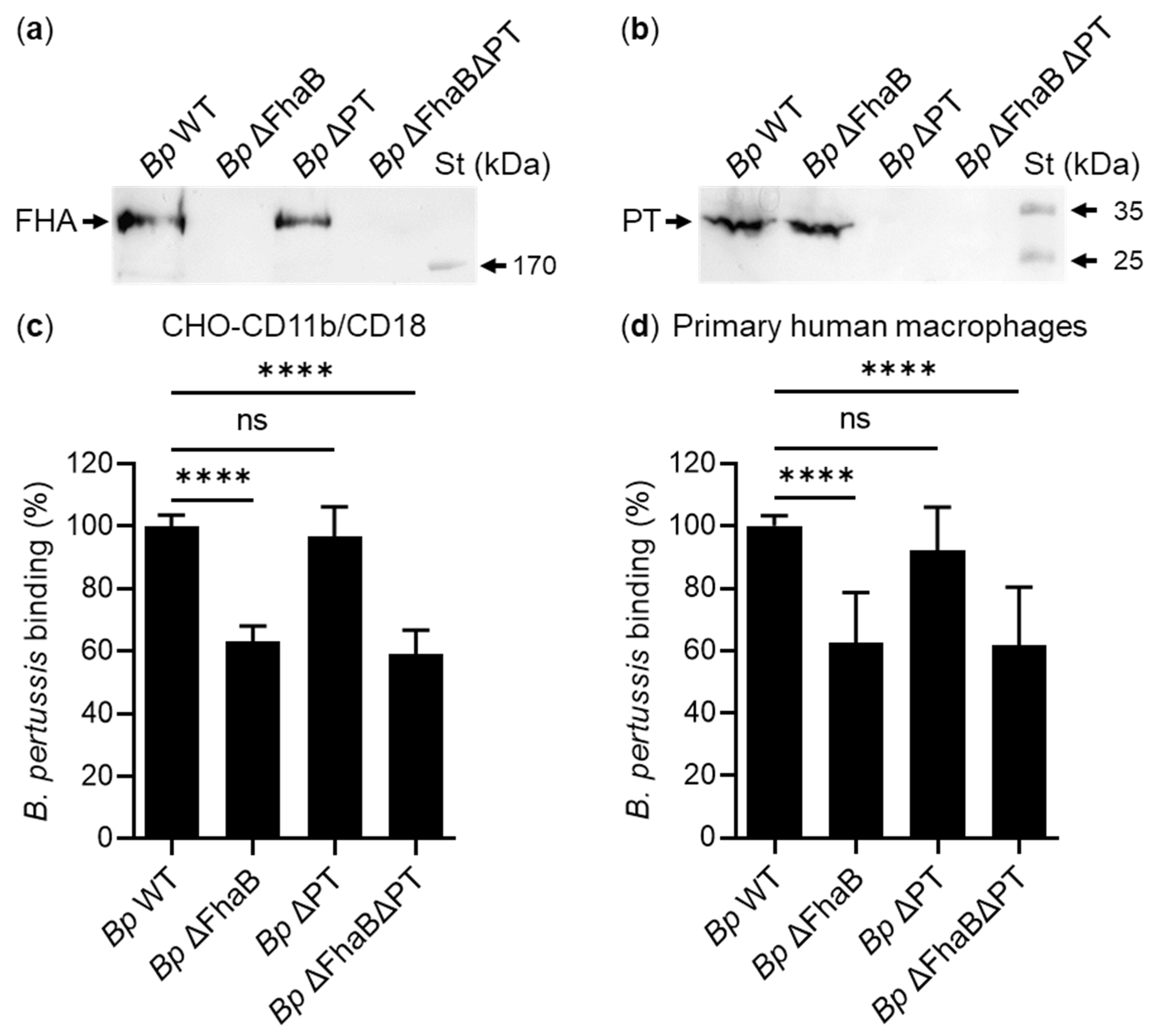
2. Results
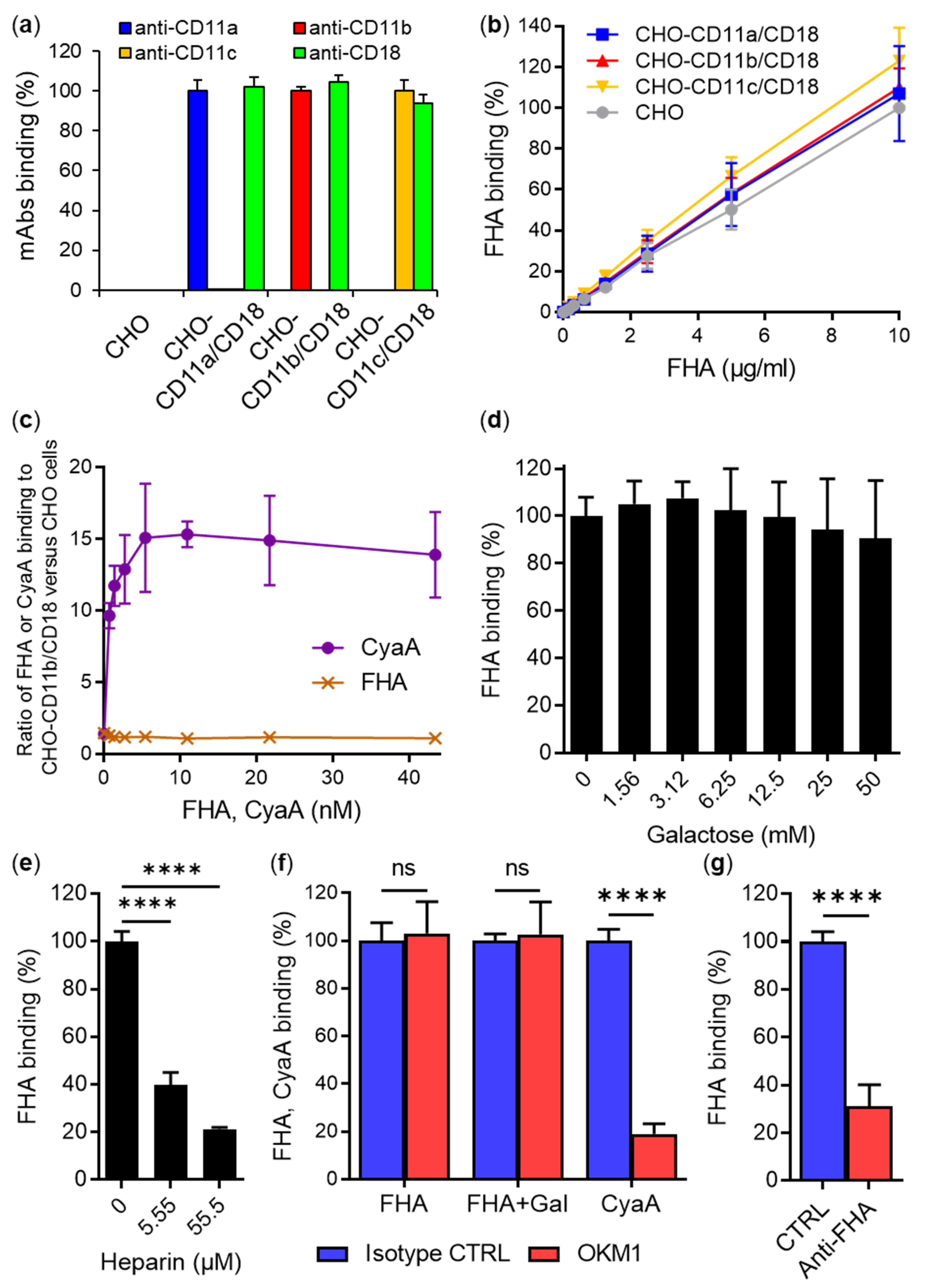
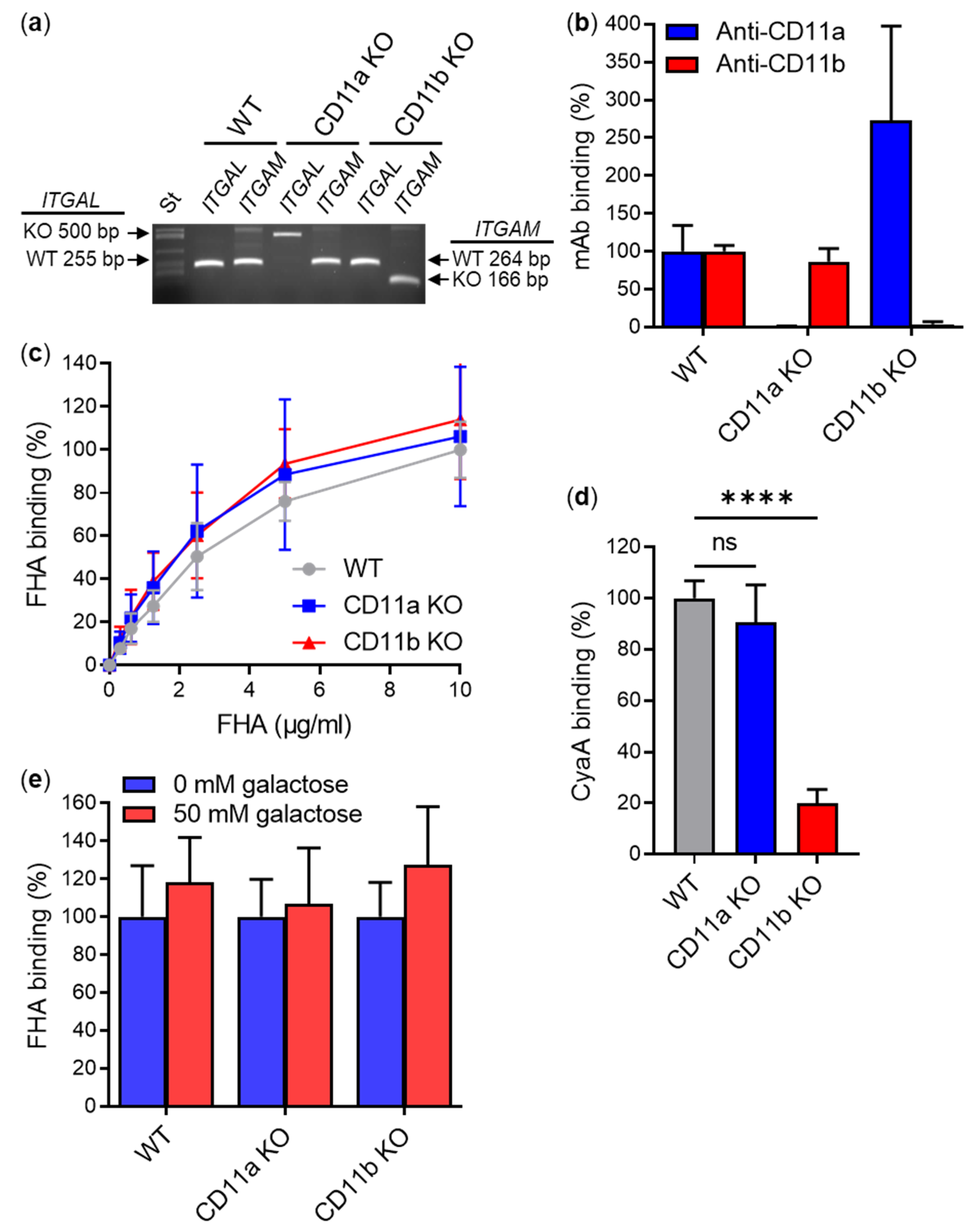
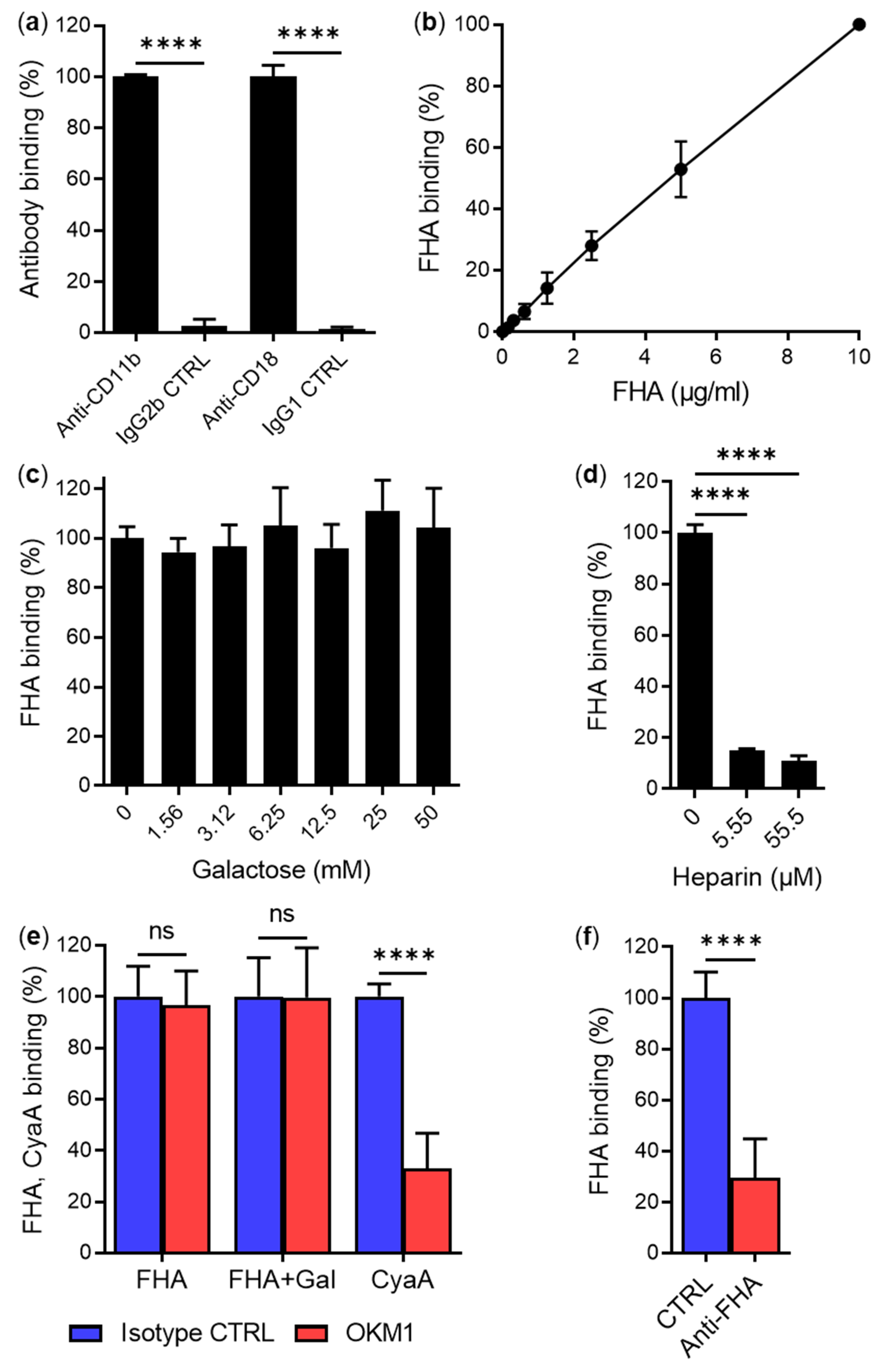
2.1. Secreted Mature FHA Does Not Bind the CD11b/CD18 Integrin
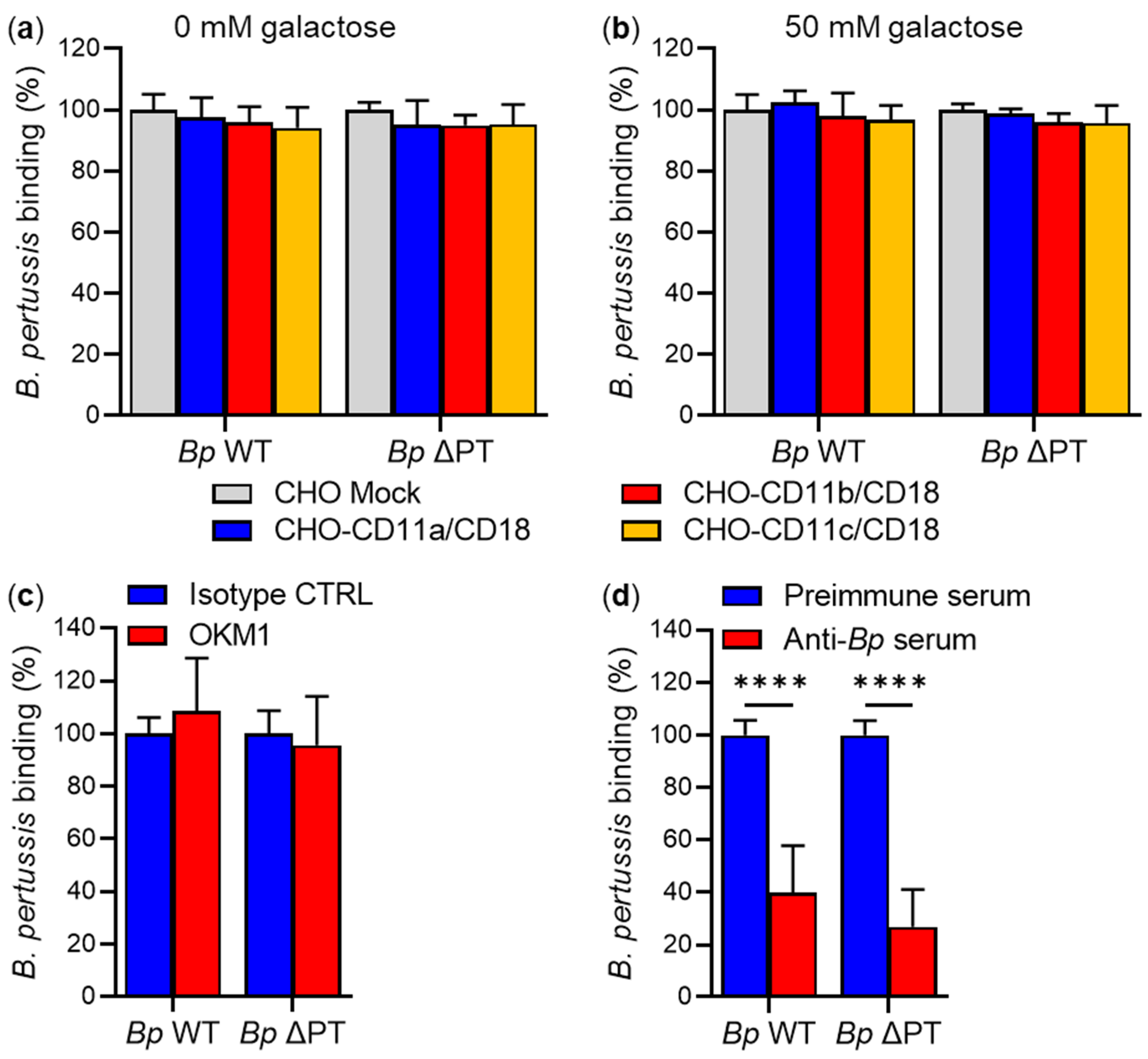
2.2. FhaB/FHA Associated with the Surface of B. pertussis Cells Does Not Recognize the CD11b/CD18 Integrin
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. Bacterial Strains and Growth Conditions
4.3. Cell Lines and Growth Conditions
4.4. Primary Mouse Macrophages
4.5. Primary Human Macrophages
4.6. Construction of B. pertussis Mutant Strains
4.7. Detection of FHA and PT in B. pertussis Strains by Western Blot
4.8. Production, Purification and Labeling of Mature FHA
4.9. Production, Purification and Labeling of CyaA
4.10. Binding of FHA, CyaA, and mAbs to Cells
4.11. Binding of FHA to Cells in the Presence of D-Galactose, Heparin, OKM1 mAb, and Anti-FHA Serum
4.12. Binding of B. pertussis Strains to Cells
4.13. Binding of B. pertussis Strains to Cells in the Presence of D-Galactose, OKM1 mAb, and Anti-B. pertussis Serum
4.14. Ethical Statement
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mattoo, S.; Cherry, J.D. Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin. Microbiol. Rev. 2005, 18, 326–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melvin, J.A.; Scheller, E.V.; Miller, J.F.; Cotter, P.A. Bordetella pertussis pathogenesis: Current and future challenges. Nat. Rev. Microbiol. 2014, 12, 274–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, K.H.T.; Duclos, P.; Nelson, E.A.S.; Hutubessy, R.C.W. An update of the global burden of pertussis in children younger than 5 years: A modelling study. Lancet Infect. Dis. 2017, 17, 974–980. [Google Scholar] [CrossRef]
- Malandra, A.; Rahman, W.U.; Klimova, N.; Streparola, G.; Holubova, J.; Osickova, A.; Bariselli, S.; Sebo, P.; Osicka, R. Bordetella Adenylate Cyclase Toxin Elicits Airway Mucin Secretion through Activation of the cAMP Response Element Binding Protein. Int. J. Mol. Sci. 2021, 22, 9064. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.; Cerny, O.; Osickova, A.; Linhartova, I.; Masin, J.; Bumba, L.; Sebo, P.; Osicka, R. Structure-Function Relationships Underlying the Capacity of Bordetella Adenylate Cyclase Toxin to Disarm Host Phagocytes. Toxins 2017, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Masin, J.; Osicka, R.; Bumba, L.; Sebo, P. Bordetella adenylate cyclase toxin: A unique combination of a pore-forming moiety with a cell-invading adenylate cyclase enzyme. Pathog. Dis. 2015, 73, ftv075. [Google Scholar] [CrossRef] [Green Version]
- Tuomanen, E.; Towbin, H.; Rosenfelder, G.; Braun, D.; Larson, G.; Hansson, G.C.; Hill, R. Receptor analogs and monoclonal antibodies that inhibit adherence of Bordetella pertussis to human ciliated respiratory epithelial cells. J. Exp. Med. 1988, 168, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Ewanowich, C.A.; Melton, A.R.; Weiss, A.A.; Sherburne, R.K.; Peppler, M.S. Invasion of HeLa 229 cells by virulent Bordetella pertussis. Infect. Immun. 1989, 57, 2698–2704. [Google Scholar] [CrossRef] [Green Version]
- Relman, D.; Tuomanen, E.; Falkow, S.; Golenbock, D.T.; Saukkonen, K.; Wright, S.D. Recognition of a bacterial adhesion by an integrin: Macrophage CR3 (alpha M beta 2, CD11b/CD18) binds filamentous hemagglutinin of Bordetella pertussis. Cell 1990, 61, 1375–1382. [Google Scholar] [CrossRef]
- Saukkonen, K.; Cabellos, C.; Burroughs, M.; Prasad, S.; Tuomanen, E. Integrin-mediated localization of Bordetella pertussis within macrophages: Role in pulmonary colonization. J. Exp. Med. 1991, 173, 1143–1149. [Google Scholar] [CrossRef]
- Friedman, R.L.; Nordensson, K.; Wilson, L.; Akporiaye, E.T.; Yocum, D.E. Uptake and intracellular survival of Bordetella pertussis in human macrophages. Infect. Immun. 1992, 60, 4578–4585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, S.M.; Yin, Y.; Rodzinski, E.; Tuomanen, E.I.; Masure, H.R. Identification of a carbohydrate recognition domain in filamentous hemagglutinin from Bordetella pertussis. Infect. Immun. 1993, 61, 2780–2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazenbos, W.L.; van den Berg, B.M.; van′t Wout, J.W.; Mooi, F.R.; van Furth, R. Virulence factors determine attachment and ingestion of nonopsonized and opsonized Bordetella pertussis by human monocytes. Infect. Immun. 1994, 62, 4818–4824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, Y.; Claus, S.; Relman, D.A. Bordetella pertussis filamentous hemagglutinin interacts with a leukocyte signal transduction complex and stimulates bacterial adherence to monocyte CR3 (CD11b/CD18). J. Exp. Med. 1994, 180, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Bassinet, L.; Gueirard, P.; Maitre, B.; Housset, B.; Gounon, P.; Guiso, N. Role of adhesins and toxins in invasion of human tracheal epithelial cells by Bordetella pertussis. Infect. Immun. 2000, 68, 1934–1941. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, Y.; Relman, D.A.; Nishikawa, A. Invasion of human respiratory epithelial cells by Bordetella pertussis: Possible role for a filamentous hemagglutinin Arg-Gly-Asp sequence and alpha5beta1 integrin. Microb. Pathog. 2001, 30, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Villarino Romero, R.; Osicka, R.; Sebo, P. Filamentous hemagglutinin of Bordetella pertussis: A key adhesin with immunomodulatory properties? Future Microbiol. 2014, 9, 1339–1360. [Google Scholar] [CrossRef]
- Abramson, T.; Kedem, H.; Relman, D.A. Proinflammatory and proapoptotic activities associated with Bordetella pertussis filamentous hemagglutinin. Infect. Immun. 2001, 69, 2650–2658. [Google Scholar] [CrossRef] [Green Version]
- Abramson, T.; Kedem, H.; Relman, D.A. Modulation of the NF-kappaB pathway by Bordetella pertussis filamentous hemagglutinin. PLoS ONE 2008, 3, e3825. [Google Scholar] [CrossRef] [Green Version]
- Dieterich, C.; Relman, D.A. Modulation of the host interferon response and ISGylation pathway by B. pertussis filamentous hemagglutinin. PLoS ONE 2011, 6, e27535. [Google Scholar] [CrossRef]
- Henderson, M.W.; Inatsuka, C.S.; Sheets, A.J.; Williams, C.L.; Benaron, D.J.; Donato, G.M.; Gray, M.C.; Hewlett, E.L.; Cotter, P.A. Contribution of Bordetella filamentous hemagglutinin and adenylate cyclase toxin to suppression and evasion of interleukin-17-mediated inflammation. Infect. Immun. 2012, 80, 2061–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuirk, P.; Mills, K.H. Direct anti-inflammatory effect of a bacterial virulence factor: IL-10-dependent suppression of IL-12 production by filamentous hemagglutinin from Bordetella pertussis. Eur. J. Immunol. 2000, 30, 415–422. [Google Scholar] [CrossRef]
- McGuirk, P.; McCann, C.; Mills, K.H. Pathogen-specific T regulatory 1 cells induced in the respiratory tract by a bacterial molecule that stimulates interleukin 10 production by dendritic cells: A novel strategy for evasion of protective T helper type 1 responses by Bordetella pertussis. J. Exp. Med. 2002, 195, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J.J.; Arai, H.; Cole, R.L. Mouse-protecting and histamine-sensitizing activities of pertussigen and fimbrial hemagglutinin from Bordetella pertussis. Infect. Immun. 1981, 32, 243–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherry, J.D.; Gornbein, J.; Heininger, U.; Stehr, K. A search for serologic correlates of immunity to Bordetella pertussis cough illnesses. Vaccine 1998, 16, 1901–1906. [Google Scholar] [CrossRef]
- Watanabe, M.; Nakase, Y. Mutant of Bordetella pertussis which lacks ability to produce filamentous hemagglutinin. Infect. Immun. 1982, 35, 1018–1023. [Google Scholar] [CrossRef] [Green Version]
- Poolman, J.T. Shortcomings of pertussis vaccines: Why we need a third generation vaccine. Expert Rev. Vaccines 2014, 13, 1159–1162. [Google Scholar] [CrossRef] [Green Version]
- Delisse-Gathoye, A.M.; Locht, C.; Jacob, F.; Raaschou-Nielsen, M.; Heron, I.; Ruelle, J.L.; de Wilde, M.; Cabezon, T. Cloning, partial sequence, expression, and antigenic analysis of the filamentous hemagglutinin gene of Bordetella pertussis. Infect. Immun. 1990, 58, 2895–2905. [Google Scholar] [CrossRef] [Green Version]
- Domenighini, M.; Relman, D.; Capiau, C.; Falkow, S.; Prugnola, A.; Scarlato, V.; Rappuoli, R. Genetic characterization of Bordetella pertussis filamentous haemagglutinin: A protein processed from an unusually large precursor. Mol. Microbiol. 1990, 4, 787–800. [Google Scholar] [CrossRef]
- Lambert-Buisine, C.; Willery, E.; Locht, C.; Jacob-Dubuisson, F. N-terminal characterization of the Bordetella pertussis filamentous haemagglutinin. Mol. Microbiol. 1998, 28, 1283–1293. [Google Scholar] [CrossRef]
- Mazar, J.; Cotter, P.A. Topology and maturation of filamentous haemagglutinin suggest a new model for two-partner secretion. Mol. Microbiol. 2006, 62, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Clantin, B.; Gruss, F.; Dewitte, F.; Delattre, A.S.; Jacob-Dubuisson, F.; Hiller, S.; Villeret, V. Conserved Omp85 lid-lock structure and substrate recognition in FhaC. Nat. Commun. 2015, 6, 7452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerin, J.; Bigot, S.; Schneider, R.; Buchanan, S.K.; Jacob-Dubuisson, F. Two-Partner Secretion: Combining Efficiency and Simplicity in the Secretion of Large Proteins for Bacteria-Host and Bacteria-Bacteria Interactions. Front. Cell. Infect. Microbiol. 2017, 7, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutte, L.; Antoine, R.; Drobecq, H.; Locht, C.; Jacob-Dubuisson, F. Subtilisin-like autotransporter serves as maturation protease in a bacterial secretion pathway. EMBO J. 2001, 20, 5040–5048. [Google Scholar] [CrossRef] [Green Version]
- Jurnecka, D.; Man, P.; Sebo, P.; Bumba, L. Bordetella pertussis and Bordetella bronchiseptica filamentous hemagglutinins are processed at different sites. FEBS Open Bio. 2018, 8, 1256–1266. [Google Scholar] [CrossRef]
- Noel, C.R.; Mazar, J.; Melvin, J.A.; Sexton, J.A.; Cotter, P.A. The prodomain of the Bordetella two-partner secretion pathway protein FhaB remains intracellular yet affects the conformation of the mature C-terminal domain. Mol. Microbiol. 2012, 86, 988–1006. [Google Scholar] [CrossRef] [Green Version]
- Nash, Z.M.; Cotter, P.A. Regulated, sequential processing by multiple proteases is required for proper maturation and release of Bordetella filamentous hemagglutinin. Mol. Microbiol. 2019, 112, 820–836. [Google Scholar] [CrossRef]
- Johnson, R.M.; Nash, Z.M.; Dedloff, M.R.; Shook, J.C.; Cotter, P.A. DegP Initiates Regulated Processing of Filamentous Hemagglutinin in Bordetella bronchiseptica. mBio 2021, 12, e0146521. [Google Scholar] [CrossRef]
- Coutte, L.; Alonso, S.; Reveneau, N.; Willery, E.; Quatannens, B.; Locht, C.; Jacob-Dubuisson, F. Role of adhesin release for mucosal colonization by a bacterial pathogen. J. Exp. Med. 2003, 197, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Julio, S.M.; Inatsuka, C.S.; Mazar, J.; Dieterich, C.; Relman, D.A.; Cotter, P.A. Natural-host animal models indicate functional interchangeability between the filamentous haemagglutinins of Bordetella pertussis and Bordetella bronchiseptica and reveal a role for the mature C-terminal domain, but not the RGD motif, during infection. Mol. Microbiol. 2009, 71, 1574–1590. [Google Scholar] [CrossRef]
- Melvin, J.A.; Scheller, E.V.; Noel, C.R.; Cotter, P.A. New Insight into Filamentous Hemagglutinin Secretion Reveals a Role for Full-Length FhaB in Bordetella Virulence. mBio 2015, 6, e01189-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menozzi, F.D.; Gantiez, C.; Locht, C. Interaction of the Bordetella pertussis filamentous hemagglutinin with heparin. FEMS Microbiol. Lett. 1991, 62, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Makhov, A.M.; Hannah, J.H.; Brennan, M.J.; Trus, B.L.; Kocsis, E.; Conway, J.F.; Wingfield, P.T.; Simon, M.N.; Steven, A.C. Filamentous hemagglutinin of Bordetella pertussis. A bacterial adhesin formed as a 50-nm monomeric rigid rod based on a 19-residue repeat motif rich in beta strands and turns. J. Mol. Biol. 1994, 241, 110–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannah, J.H.; Menozzi, F.D.; Renauld, G.; Locht, C.; Brennan, M.J. Sulfated glycoconjugate receptors for the Bordetella pertussis adhesin filamentous hemagglutinin (FHA) and mapping of the heparin-binding domain on FHA. Infect. Immun. 1994, 62, 5010–5019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menozzi, F.D.; Mutombo, R.; Renauld, G.; Gantiez, C.; Hannah, J.H.; Leininger, E.; Brennan, M.J.; Locht, C. Heparin-inhibitable lectin activity of the filamentous hemagglutinin adhesin of Bordetella pertussis. Infect. Immun. 1994, 62, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, Y.; Yoshimura, K.; Nishikawa, A.; Claus, S.; Laudanna, C.; Relman, D.A. Role of phosphatidylinositol 3-kinase in the binding of Bordetella pertussis to human monocytes. Cell. Microbiol. 2002, 4, 825–833. [Google Scholar] [CrossRef]
- Osicka, R.; Osickova, A.; Hasan, S.; Bumba, L.; Cerny, J.; Sebo, P. Bordetella adenylate cyclase toxin is a unique ligand of the integrin complement receptor 3. eLife 2015, 4, e10766. [Google Scholar] [CrossRef]
- Arnaout, M.A.; Mahalingam, B.; Xiong, J.P. Integrin structure, allostery, and bidirectional signaling. Annu. Rev. Cell Dev. Biol. 2005, 21, 381–410. [Google Scholar] [CrossRef] [Green Version]
- Ulanova, M.; Gravelle, S.; Barnes, R. The role of epithelial integrin receptors in recognition of pulmonary pathogens. J. Innate Immun. 2009, 1, 4–17. [Google Scholar] [CrossRef]
- Ruoslahti, E. RGD and other recognition sequences for integrins. Annu. Rev. Cell Dev. Biol. 1996, 12, 697–715. [Google Scholar] [CrossRef]
- Hazenbos, W.L.; van den Berg, B.M.; Geuijen, C.W.; Mooi, F.R.; van Furth, R. Binding of FimD on Bordetella pertussis to very late antigen-5 on monocytes activates complement receptor type 3 via protein tyrosine kinases. J. Immunol. 1995, 155, 3972–3978. [Google Scholar] [PubMed]
- Guermonprez, P.; Khelef, N.; Blouin, E.; Rieu, P.; Ricciardi-Castagnoli, P.; Guiso, N.; Ladant, D.; Leclerc, C. The adenylate cyclase toxin of Bordetella pertussis binds to target cells via the alpha(M)beta(2) integrin (CD11b/CD18). J. Exp. Med. 2001, 193, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Leininger, E.; Ewanowich, C.A.; Bhargava, A.; Peppler, M.S.; Kenimer, J.G.; Brennan, M.J. Comparative roles of the Arg-Gly-Asp sequence present in the Bordetella pertussis adhesins pertactin and filamentous hemagglutinin. Infect. Immun. 1992, 60, 2380–2385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relman, D.A.; Domenighini, M.; Tuomanen, E.; Rappuoli, R.; Falkow, S. Filamentous hemagglutinin of Bordetella pertussis: Nucleotide sequence and crucial role in adherence. Proc. Natl. Acad. Sci. USA 1989, 86, 2637–2641. [Google Scholar] [CrossRef] [Green Version]
- Leininger, E.; Kenimer, J.G.; Brennan, M.J. Surface Proteins of Bordetella pertussis: Role in Adherence. In Proceedings of the Sixth International Symposium on Pertussis; Manclark, C.R., Ed.; Department of Health and Human Services, United States Public Health Service: Bethesda, MD, USA, 1990; pp. 100–105. [Google Scholar]
- Irons, L.I.; Ashworth, L.A.; Wilton-Smith, P. Heterogeneity of the filamentous haemagglutinin of Bordetella pertussis studied with monoclonal antibodies. J. Gen. Microbiol. 1983, 129, 2769–2778. [Google Scholar] [CrossRef] [Green Version]
- Stainer, D.W.; Scholte, M.J. A simple chemically defined medium for the production of phase I Bordetella pertussis. J. Gen. Microbiol. 1970, 63, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Rahman, W.U.; Osickova, A.; Klimova, N.; Lora, J.; Balashova, N.; Osicka, R. Binding of Kingella kingae RtxA Toxin Depends on Cell Surface Oligosaccharides, but Not on beta2 Integrins. Int. J. Mol. Sci. 2020, 21, 9092. [Google Scholar] [CrossRef]
- Ding, Z.M.; Babensee, J.E.; Simon, S.I.; Lu, H.; Perrard, J.L.; Bullard, D.C.; Dai, X.Y.; Bromley, S.K.; Dustin, M.L.; Entman, M.L.; et al. Relative contribution of LFA-1 and Mac-1 to neutrophil adhesion and migration. J. Immunol. 1999, 163, 5029–5038. [Google Scholar]
- Coxon, A.; Rieu, P.; Barkalow, F.J.; Askari, S.; Sharpe, A.H.; von Andrian, U.H.; Arnaout, M.A.; Mayadas, T.N. A novel role for the beta 2 integrin CD11b/CD18 in neutrophil apoptosis: A homeostatic mechanism in inflammation. Immunity 1996, 5, 653–666. [Google Scholar] [CrossRef] [Green Version]
- Menck, K.; Behme, D.; Pantke, M.; Reiling, N.; Binder, C.; Pukrop, T.; Klemm, F. Isolation of human monocytes by double gradient centrifugation and their differentiation to macrophages in teflon-coated cell culture bags. J. Vis. Exp. JoVE 2014, 91, e51554. [Google Scholar] [CrossRef] [Green Version]
- Inatsuka, C.S.; Xu, Q.; Vujkovic-Cvijin, I.; Wong, S.; Stibitz, S.; Miller, J.F.; Cotter, P.A. Pertactin is required for Bordetella species to resist neutrophil-mediated clearance. Infect. Immun. 2010, 78, 2901–2909. [Google Scholar] [CrossRef] [PubMed]
- Klimova, N.; Holubova, J.; Streparola, G.; Tomala, J.; Brazdilova, L.; Stanek, O.; Bumba, L.; Sebo, P. Pertussis toxin suppresses dendritic cell-mediated delivery of B. pertussis into lung-draining lymph nodes. PLoS Pathog. 2022, 18, e1010577. [Google Scholar] [CrossRef] [PubMed]
- Osicka, R.; Osickova, A.; Basar, T.; Guermonprez, P.; Rojas, M.; Leclerc, C.; Sebo, P. Delivery of CD8(+) T-cell epitopes into major histocompatibility complex class I antigen presentation pathway by Bordetella pertussis adenylate cyclase: Delineation of cell invasive structures and permissive insertion sites. Infect. Immun. 2000, 68, 247–256. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Primer Name | Primer Sequence 1 |
|---|---|---|
| Bp ΔPT | ΔPtx SpeI for | CTACTAGTGCGGTGCTGGAACATATCC |
| ΔPtx SacI rev | CTGAGCTCCATCCCGTCTTCCCCTCTG | |
| ΔPtx SacI for | CTGAGCTCTGAGCCGCCGGCTCGGATC | |
| ΔPtx BamHI rev | CTGGATCCCAGCGGCGCATAGACGGTAC | |
| Bp ΔFHA | ΔFhaB NotI for | CTGCGGCCGCGGCATTGATGACCTCGTGCAG |
| ΔFhaB SpeI rev | GAACTAGTCGTGTTCATATTCCGACCAGC | |
| ΔFhaB SpeI for | CTACTAGTAACAAATAGGTAGTCGCGGCCTG | |
| ΔFhaB BamHIrev | GAGGATCCCATGCCGCCTTGCCGCTTTAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golshani, M.; Rahman, W.U.; Osickova, A.; Holubova, J.; Lora, J.; Balashova, N.; Sebo, P.; Osicka, R. Filamentous Hemagglutinin of Bordetella pertussis Does Not Interact with the β2 Integrin CD11b/CD18. Int. J. Mol. Sci. 2022, 23, 12598. https://doi.org/10.3390/ijms232012598
Golshani M, Rahman WU, Osickova A, Holubova J, Lora J, Balashova N, Sebo P, Osicka R. Filamentous Hemagglutinin of Bordetella pertussis Does Not Interact with the β2 Integrin CD11b/CD18. International Journal of Molecular Sciences. 2022; 23(20):12598. https://doi.org/10.3390/ijms232012598
Chicago/Turabian StyleGolshani, Maryam, Waheed Ur Rahman, Adriana Osickova, Jana Holubova, Jinery Lora, Nataliya Balashova, Peter Sebo, and Radim Osicka. 2022. "Filamentous Hemagglutinin of Bordetella pertussis Does Not Interact with the β2 Integrin CD11b/CD18" International Journal of Molecular Sciences 23, no. 20: 12598. https://doi.org/10.3390/ijms232012598