
Anticonvulsive Effects and Pharmacokinetic Profile of Cannabidiol (CBD) in the Pentylenetetrazol (PTZ) or N-Methyl-D-Aspartate (NMDA) Models of Seizures in Infantile Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
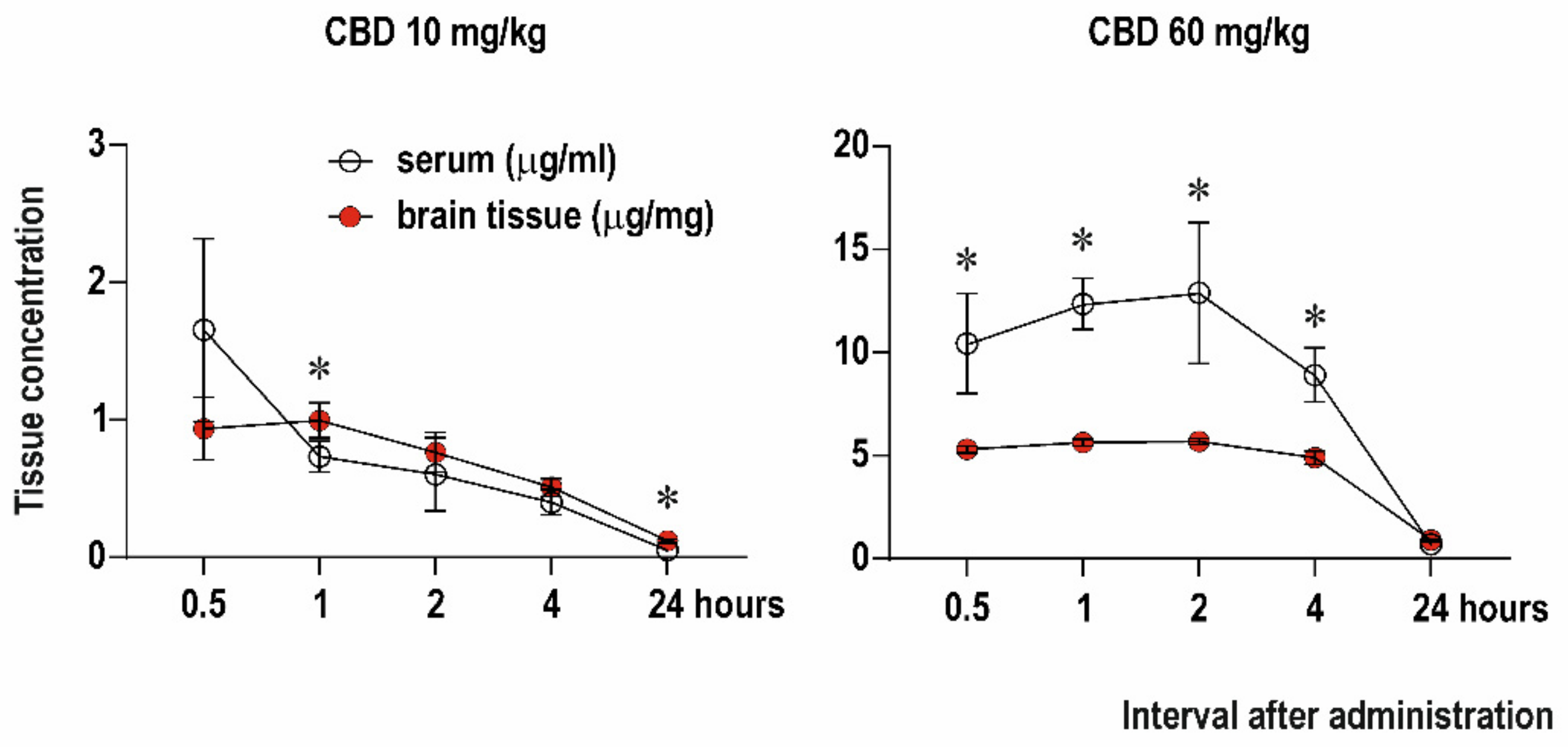
2.1. Serum and Brain Levels of CBD
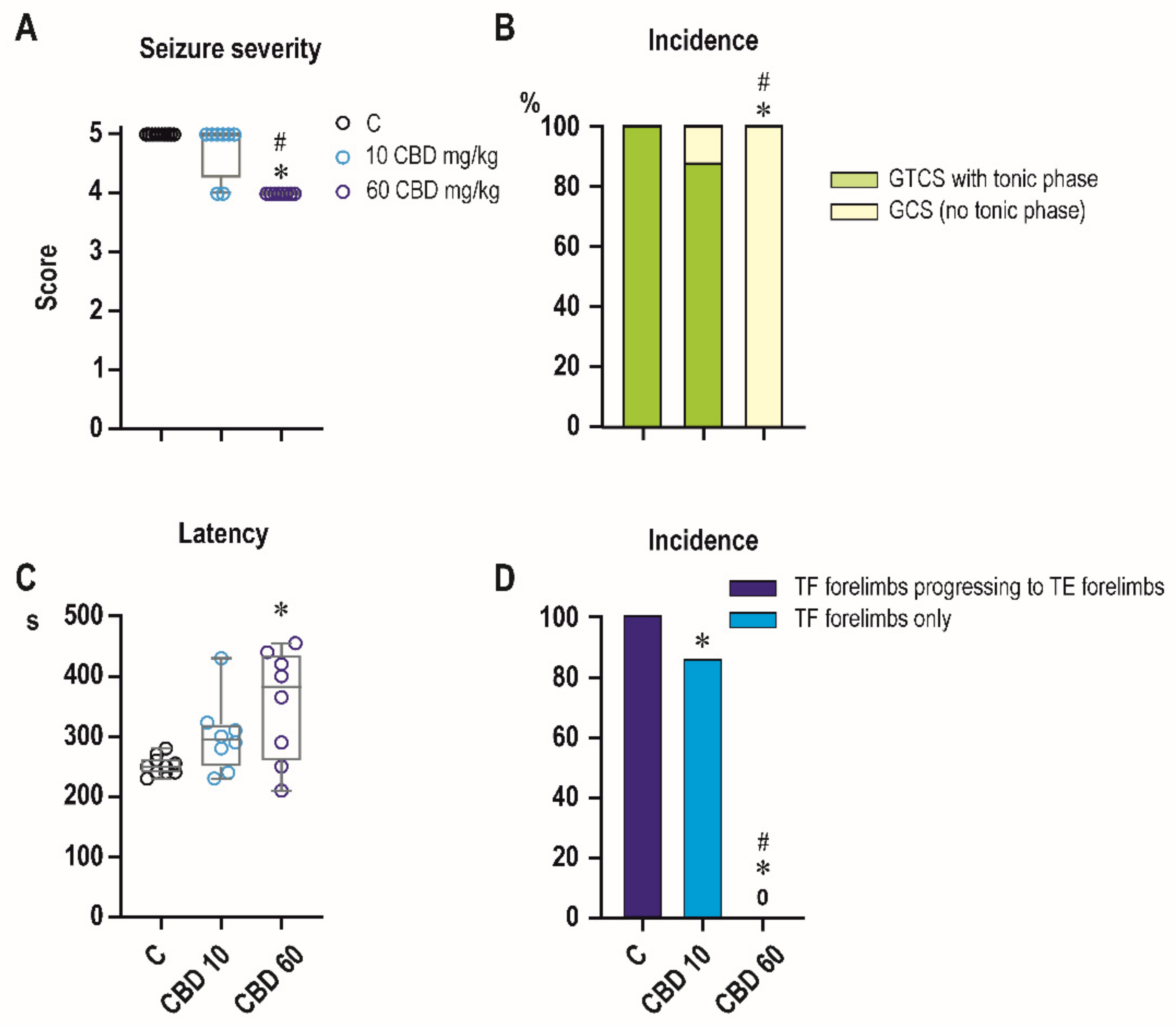
2.2. Effects of CBD on PTZ-Induced Seizures
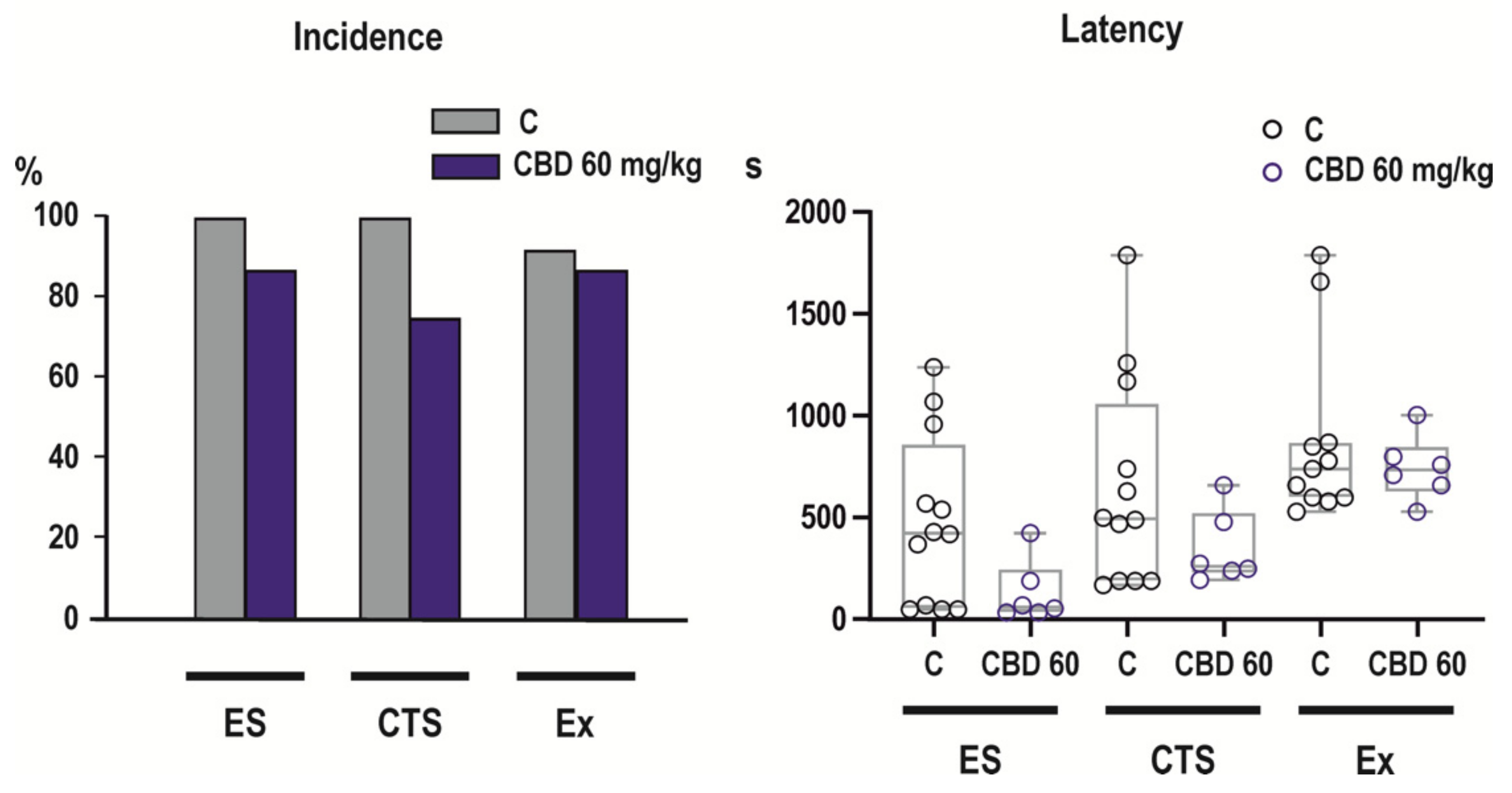
2.3. Effects of CBD on NMDA-Induced Seizures
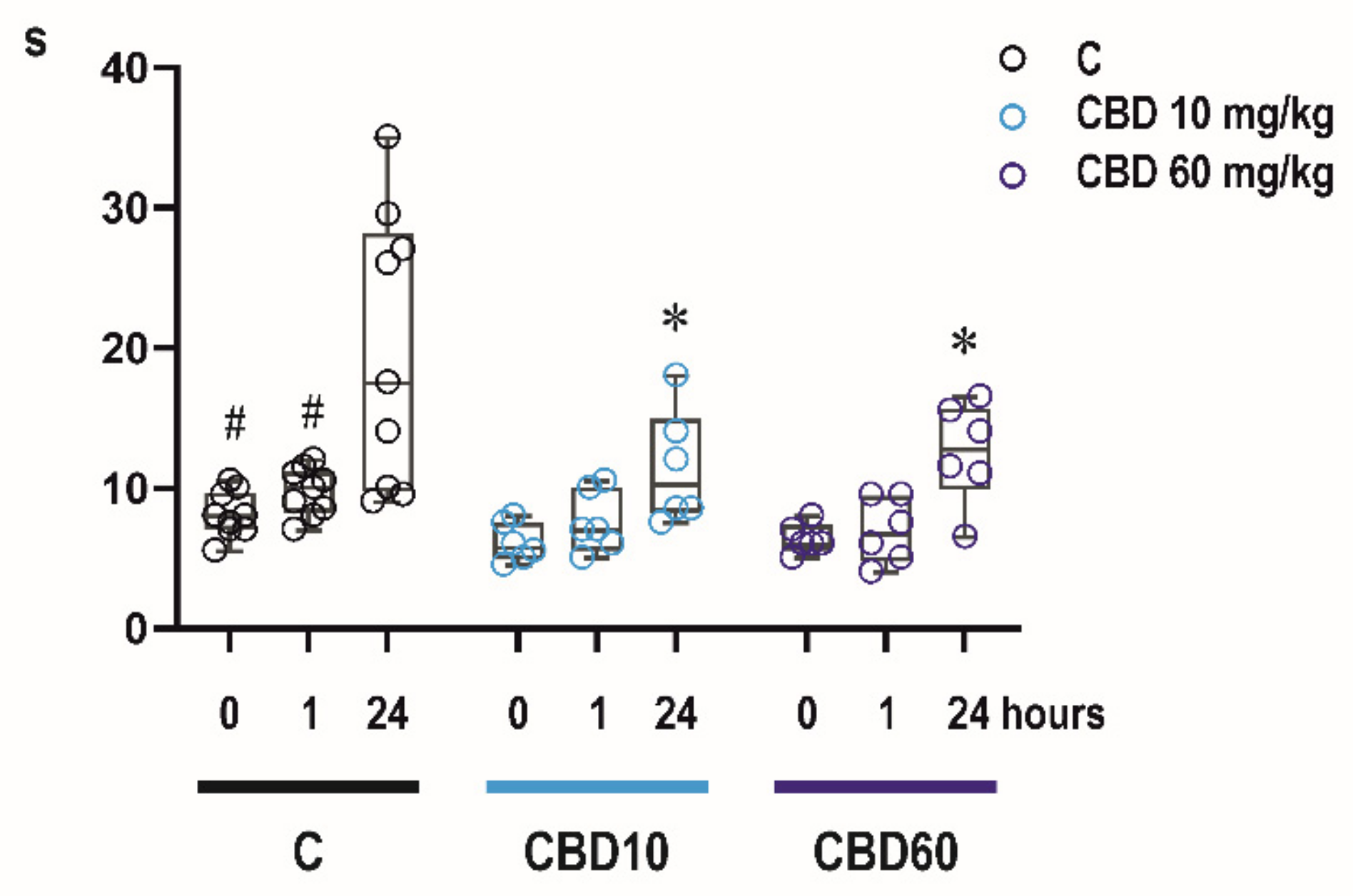
2.4. Effects of CBD on Sensorimotor Abilities of Infant Rats
2.4.1. Bar Holding Test
2.4.2. Righting Reflexes
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drug Administration and Samples Collection
4.3. Assessment of Plasma and Brain Levels of CBD
4.4. Anticonvulsant Effects of CBD
4.4.1. PTZ Model
4.4.2. NMDA Model
- Clonic-tonic seizures (CTS) appeared after flexion seizures. The pattern of generalized clonic-tonic seizures was further analyzed as concerns the presence and distribution of the tonic phase.
4.5. Effects of CBD on Motor Performance
4.5.1. Righting Reflexes
4.5.2. Bar holding
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alexander, S.P.H. Therapeutic potential of cannabis-related drugs. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2016, 64, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Morrison, P.D.; Fusar-Poli, P.; Martin-Santos, R.; Borgwardt, S.; Wintonbrown, T.T.; Nosarti, C.; Carroll, C.M.O.; Seal, M.L.; Allen, P.; et al. Opposite effects of δ-9-tetrahydrocannabinol and cannabidiol on human brain function and psychopathology. Neuropsychopharmacology 2010, 35, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Englund, A.; Morrison, P.D.; Nottage, J.F.; Hague, D.; Kane, F.; Bonaccorso, S.; Stone, J.M.; Reichenberg, A.; Brenneisen, R.; Holt, D.; et al. Cannabidiol inhibits THC-elicited paranoid symptoms and hippocampal-dependent memory impairment. J. Psychopharmacol. 2013, 27, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Mishima, K.; Fujiwara, M. Therapeutic Potential of Non-Psychotropic Cannabidiol in Ischemic Stroke. Pharmaceuticals 2010, 3, 2197–2212. [Google Scholar] [CrossRef]
- McGuire, P.; Robson, P.; Cubała, W.; Vasile, D.; Morrison, P.D.; Barron, R.; Taylor, A.; Wright, S. Cannabidiol (CBD) as an Adjunctive Therapy in Schizophrenia: A Multicenter Randomized Controlled Trial. Am. J. Psychiatry 2018, 175, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Resstel, L.B.M.; Joca, S.R.L.; Moreira, F.A.; Correa, F.M.A.; Guimaraes, F.S. Effects of cannabidiol and diazepam on behavioral and cardiovascular responses induced by contextual conditioned fear in rats. Behav. Brain Res. 2006, 172, 294–298. [Google Scholar] [CrossRef]
- Schiavon, A.P.; Bonato, J.M.; Milani, H.; Guimaraes, F.S.; de Oliveira, R.M.W. Influence of single and repeated cannabidiol administration on emotional behavior and markers of cell proliferation and neurogenesis in non-stressed mice. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2016, 64, 27–34. [Google Scholar] [CrossRef]
- Devinsky, O.; Cross, H.; Laux, L.; Marsh, E.; Miller, I.; Nabbout, R.; Scheffer, I.; Thiele, E.A.; Wright, S. Trial of Cannabidiol for Drug-Resistant Seizures in the Dravet Syndrome. N. Engl. J. Med. 2017, 376, 2011–2020. [Google Scholar] [CrossRef] [Green Version]
- Devinsky, O.; Patel, A.D.; Cross, H.; Villanueva, V.; Wirrell, E.C.; Privitera, M.; Greenwood, S.M.; Roberts, C.; Checketts, D.; VanLandingham, K.E.; et al. Effect of Cannabidiol on Drop Seizures in the Lennox–Gastaut Syndrome. N. Engl. J. Med. 2018, 378, 1888–1897. [Google Scholar] [CrossRef] [Green Version]
- Corroon, J.; Kight, R. Regulatory Status of Cannabidiol in the United States: A Perspective. Cannabis Cannabinoid Res. 2018, 3, 190–194. [Google Scholar] [CrossRef]
- Izquierdo, I.; Tannhauser, M. Letter: The effect of cannabidiol on maximal electroshock seizures in rats. J. Pharm. Pharmacol. 1973, 25, 916–917. [Google Scholar] [CrossRef]
- Chesher, G.B.; Dahl, C.J.; Everingham, M.; Jackson, D.M.; Marchant-Williams, H.; Starmer, G.A. The effect of cannabinoids on intestinal motility and their antinociceptive effect in mice. Br. J. Pharmacol. 1973, 49, 588–594. [Google Scholar] [CrossRef] [Green Version]
- Franco, V.; Perucca, E. Pharmacological and Therapeutic Properties of Cannabidiol for Epilepsy. Drugs 2019, 79, 1435–1454. [Google Scholar] [CrossRef]
- Franco, V.; Bialer, M.; Perucca, E. Cannabidiol in the treatment of epilepsy: Current evidence and perspectives for further research. Neuropharmacology 2021, 185, 108442. [Google Scholar] [CrossRef]
- Rosenberg, E.C.; Tsien, R.W.; Whalley, B.J.; Devinsky, O. Cannabinoids and Epilepsy. Neurotherapeutics 2015, 12, 747–768. [Google Scholar] [CrossRef] [Green Version]
- Senn, L.; Cannazza, G.; Biagini, G. Receptors and channels possibly mediating the effects of phytocannabinoids on seizures and epilepsy. Pharmaceuticals 2020, 13, 174. [Google Scholar] [CrossRef]
- Benn, E.K.; Hauser, W.A.; Shih, T.; Leary, L.; Bagiella, E.; Dayan, P.; Green, R.; Andrews, H.; Thurman, D.J.; Hesdorffer, D.C. Estimating the incidence of first unprovoked seizure and newly diagnosed epilepsy in the low-income urban community of Northern Manhattan, New York City. Epilepsia 2008, 49, 1431–1439. [Google Scholar] [CrossRef]
- Cowan, L.D. The epidemiology of the epilepsies in children. Ment. Retard. Dev. Disabil. Res. Rev. 2002, 8, 171–181. [Google Scholar] [CrossRef]
- Clancy, B.; Darlington, R.B.; Finlay, B.L. Translating developmental time across mammalian species. Neuroscience 2001, 105, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Workman, A.D.; Charvet, C.J.; Clancy, B.; Darlington, R.B.; Finlay, B.L. Modeling transformations of neurodevelopmental sequences across mammalian species. J. Neurosci. 2013, 33, 7368–7383. [Google Scholar] [CrossRef]
- Ellingson, R.J. Studies of the Electrical Activity of the Developing Human Brain. Prog. Brain Res. 1964, 9, 26–53. [Google Scholar] [CrossRef]
- Mares, P.; Velisek, L. N-methyl-D-aspartate (NMDA)-induced seizures in developing rats. Dev. Brain Res. 1992, 65, 185–189. [Google Scholar] [CrossRef]
- Klein, B.D.; Jacobson, C.A.; Metcalf, C.S.; Smith, M.D.; Wilcox, K.S.; Hampson, A.J.; Kehne, J.H. Evaluation of Cannabidiol in Animal Seizure Models by the Epilepsy Therapy Screening Program (ETSP). Neurochem. Res. 2017, 42, 1939–1948. [Google Scholar] [CrossRef]
- Huizenga, M.N.; Sepulveda-Rodriguez, A.; Forcelli, P.A. Preclinical safety and efficacy of cannabidivarin for early life seizures. Neuropharmacology 2019, 148, 189–198. [Google Scholar] [CrossRef]
- Staňková, L.; Kubová, H.; Mareš, P. Anticonvulsant action of lamotrigine during ontogenesis in rats. Epilepsy Res. 1992, 13, 17–22. [Google Scholar] [CrossRef]
- Kubová, H.; Maresš, P. Anticonvulsant Action of Oxcarbazepine, Hydroxycarbamazepine, and Carbamazepine Against Metrazol-Induced Motor Seizures in Developing Rats. Epilepsia 1993, 34, 188–192. [Google Scholar] [CrossRef]
- Haugvicová, R.; Kubová, H.; Mareš, P. Qualitative changes of anticonvulsant action of felbamate during development in rats. Brain Dev. 1998, 20, 222–226. [Google Scholar] [CrossRef]
- Löscher, W. Critical review of current animal models of seizures and epilepsy used in the discovery and development of new antiepileptic drugs. Seizure 2011, 20, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Kabova, R.; Liptakova, S.; Slamberova, R.; Pometlova, M.; Velisek, L. Age-specific N-methyl-D-aspartate-induced seizures: Perspectives for the West syndrome model. Epilepsia 1999, 40, 1357–1369. [Google Scholar] [CrossRef]
- Herlopian, A.; Hess, E.J.; Barnett, J.; Geffrey, A.L.; Pollack, S.F.; Skirvin, L.; Bruno, P.; Sourbron, J.; Thiele, E.A. Cannabidiol in treatment of refractory epileptic spasms: An open-label study. Epilepsy Behav. 2020, 106, 106988. [Google Scholar] [CrossRef]
- Rodríguez-Muñoz, M.; Onetti, Y.; Cortés-Montero, E.; Garzón, J.; Sánchez-Blázquez, P. Cannabidiol enhances morphine antinociception, diminishes NMDA-mediated seizures and reduces stroke damage via the sigma 1 receptor 11 Medical and Health Sciences 1109 Neurosciences 11 Medical and Health Sciences 1115 Pharmacology and Pharmaceutical Scien. Mol. Brain 2018, 11, 1–12. [Google Scholar] [CrossRef]
- Macdonald, R.L.; Barker, J.L. Specific antagonism of GABA-mediated postsynaptic inhibition in cultured mammalian spinal cord neurons: A common mode of convulsant action. Neurology 1978, 28, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Kubová, H.; Folbergrová, J.; Mares, P. Seizures induced by homocysteine in rats during ontogenesis. Epilepsia 1995, 36, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Mares, P.; Folbergrová, J.; Langmeier, M.; Haugvicová, R.; Kubová, H. Convulsant action of D,L-homocysteic acid and its stereoisomers in immature rats. Epilepsia 1997, 38, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Insel, T.R.; Miller, L.P.; Gelhard, R.E. The ontogeny of excitatory amino acid receptors in rat forebrain—I. N-methyl-D-aspartate and quisqualate receptors. Neuroscience 1990, 35, 31–43. [Google Scholar] [CrossRef]
- Tsumoto, T.; Hagihara, K.; Sato, H.; Hata, Y. NMDA receptors in the visual cortex of young kittens are more effective than those of adult cats. Nature 1987, 327, 513–514. [Google Scholar] [CrossRef]
- Hamon, B.; Heinemann, U. Developmental changes in neuronal sensitivity to excitatory amino acids in area CA1 of the rat hippocampus. Brain Res. 1988, 466, 286–290. [Google Scholar] [CrossRef]
- Kubová, H. Ontogenesis and treatment efficacy: Prevention of seizures in the immature brain. Adv. Neurol. 1999, 81, 357–361. [Google Scholar]
- Kubová, H.; Mares, P. Vigabatrin but not valproate prevents development of age-specific flexion seizures induced by N-methyl-D-aspartate (NMDA) in immature rats. Epilepsia 2010, 51, 469–472. [Google Scholar] [CrossRef]
- Bittigau, P.; Sifringer, M.; Genz, K.; Reith, E.; Pospischil, D.; Govindarajalu, S.; Dzietko, M.; Pesditschek, S.; Mai, I.; Dikranian, K.; et al. Antiepileptic drugs and apoptotic neurodegeneration in the developing brain. Proc. Natl. Acad. Sci. USA 2002, 99, 15089–15094. [Google Scholar] [CrossRef] [Green Version]
- Ikonomidou, C.; Turski, L. Antiepileptic drugs and brain development. Epilepsy Res. 2010, 88, 11–22. [Google Scholar] [CrossRef]
- Altman, J.; Sudarshan, K. Postnatal development of locomotion in the laboratory rat. Anim. Behav. 1975, 23, 896–920. [Google Scholar] [CrossRef]
- Jänicke, B.; Schulze, G.; Coper, H. Motor performance achievements in rats of different ages. Exp. Gerontol. 1983, 18, 393–407. [Google Scholar] [CrossRef]
- Deiana, S.; Watanabe, A.; Yamasaki, Y.; Amada, N.; Arthur, M.; Fleming, S.; Woodcock, H.; Dorward, P.; Pigliacampo, B.; Close, S.; et al. Plasma and brain pharmacokinetic profile of cannabidiol (CBD), cannabidivarine (CBDV), Δ 9-tetrahydrocannabivarin (THCV) and cannabigerol (CBG) in rats and mice following oral and intraperitoneal administration and CBD action on obsessive-compulsive behav. Psychopharmacology 2012, 219, 859–873. [Google Scholar] [CrossRef]
- McPhail, B.T.; White, C.A.; Cummings, B.S.; Muralidhara, S.; Wilson, J.T.; Bruckner, J.V. The immature rat as a potential model for chemical risks to children: Ontogeny of selected hepatic P450s. Chem. Biol. Interact. 2016, 256, 167–177. [Google Scholar] [CrossRef]
- Hložek, T.; Uttl, L.; Kadeřábek, L.; Balíková, M.; Lhotková, E.; Horsley, R.R.; Nováková, P.; Šíchová, K.; Štefková, K.; Tylš, F.; et al. Pharmacokinetic and behavioural profile of THC, CBD, and THC + CBD combination after pulmonary, oral, and subcutaneous administration in rats and confirmation of conversion in vivo of CBD to THC. Eur. Neuropsychopharmacol. 2017, 27, 1223–1237. [Google Scholar] [CrossRef]
- Vacek, J.; Papouskova, B.; Polanska, H.; Hönigova, K.; Storch, J.; Babula, P.; Masarik, M. CBD is not converted to THC in rats: A framework interpretation and discussion. Eur. Neuropsychopharmacol. 2021, 50, 135–136. [Google Scholar] [CrossRef]
- Velisek, L.; Kubova, H.; Pohl, M.; Stankova, L.; Mareš, P.; Schickerova, R. Pentylenetetrazol-induced seizures in rats: An ontogenetic study. Naunyn Schmiedeberg’s Arch. Pharmacol. 1992, 346, 588–591. [Google Scholar] [CrossRef]
- Pohl, M.; Mares, P. Effects of flunarizine on Metrazol-induced seizures in developing rats. Epilepsy Res. 1987, 1, 302–305. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uttl, L.; Hložek, T.; Mareš, P.; Páleníček, T.; Kubová, H. Anticonvulsive Effects and Pharmacokinetic Profile of Cannabidiol (CBD) in the Pentylenetetrazol (PTZ) or N-Methyl-D-Aspartate (NMDA) Models of Seizures in Infantile Rats. Int. J. Mol. Sci. 2022, 23, 94. https://doi.org/10.3390/ijms23010094
Uttl L, Hložek T, Mareš P, Páleníček T, Kubová H. Anticonvulsive Effects and Pharmacokinetic Profile of Cannabidiol (CBD) in the Pentylenetetrazol (PTZ) or N-Methyl-D-Aspartate (NMDA) Models of Seizures in Infantile Rats. International Journal of Molecular Sciences. 2022; 23(1):94. https://doi.org/10.3390/ijms23010094
Chicago/Turabian StyleUttl, Libor, Tomáš Hložek, Pavel Mareš, Tomáš Páleníček, and Hana Kubová. 2022. "Anticonvulsive Effects and Pharmacokinetic Profile of Cannabidiol (CBD) in the Pentylenetetrazol (PTZ) or N-Methyl-D-Aspartate (NMDA) Models of Seizures in Infantile Rats" International Journal of Molecular Sciences 23, no. 1: 94. https://doi.org/10.3390/ijms23010094