
The Gut Microbiota Affects Corticosterone Production in the Murine Small Intestine


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expression of Glucocorticoid-Related Genes in the Intestine of Anti-CD3 Antibody-Treated Mice
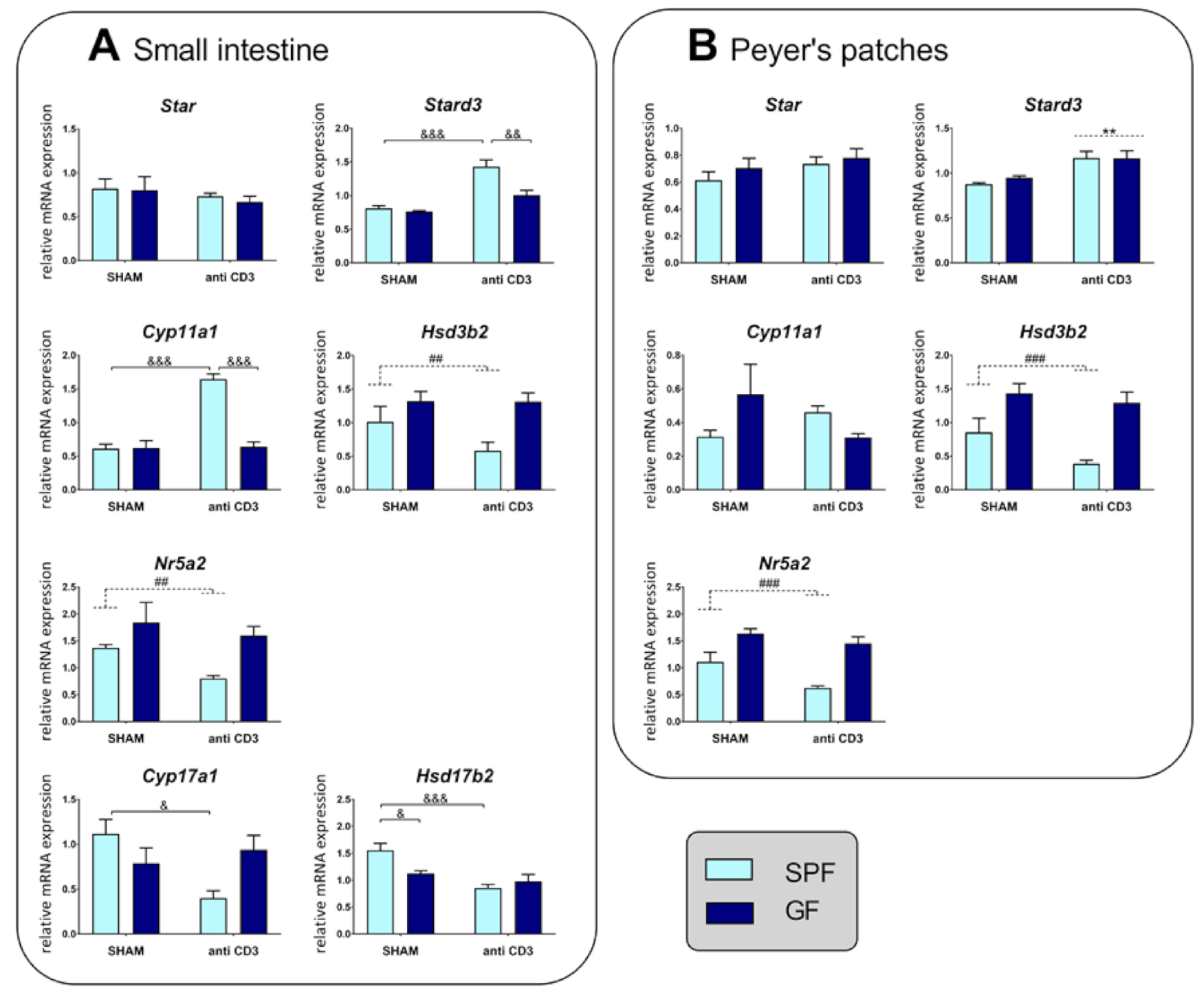
2.2. Effect of the Microbiota and Acute Immune Stress on the Expression of Genes Associated with Steroidogenesis in the Small Intestine and Peyer’s Patches
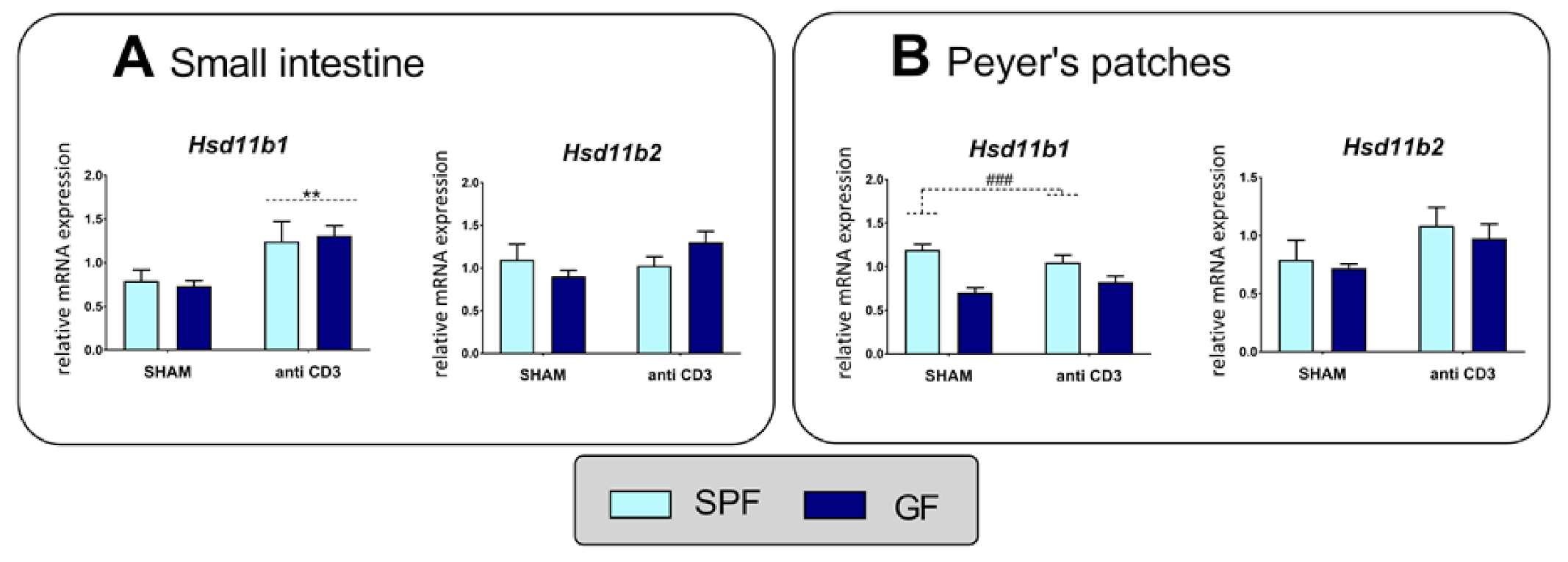
2.3. Expression Levels of 11β-Hydroxysteroid Dehydrogenase Type 1 and Type 2 in the Small Intestine and Peyer’s Patches
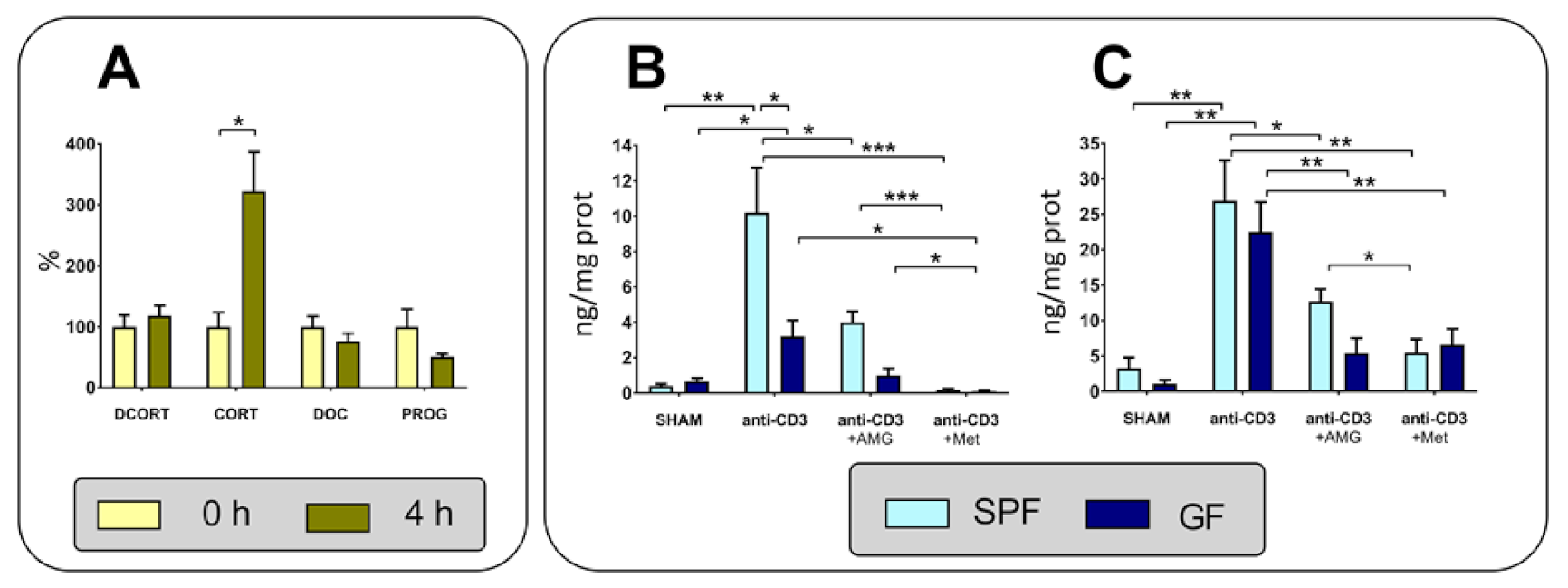
2.4. Production of Steroids in the Intestine
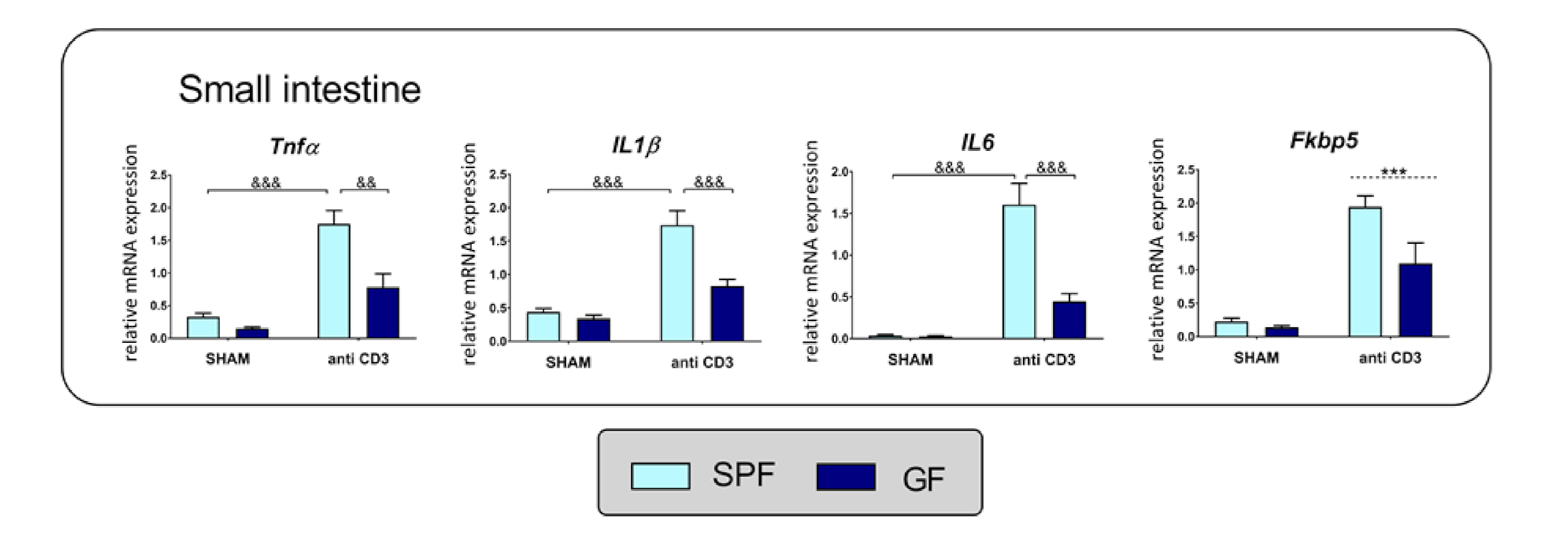
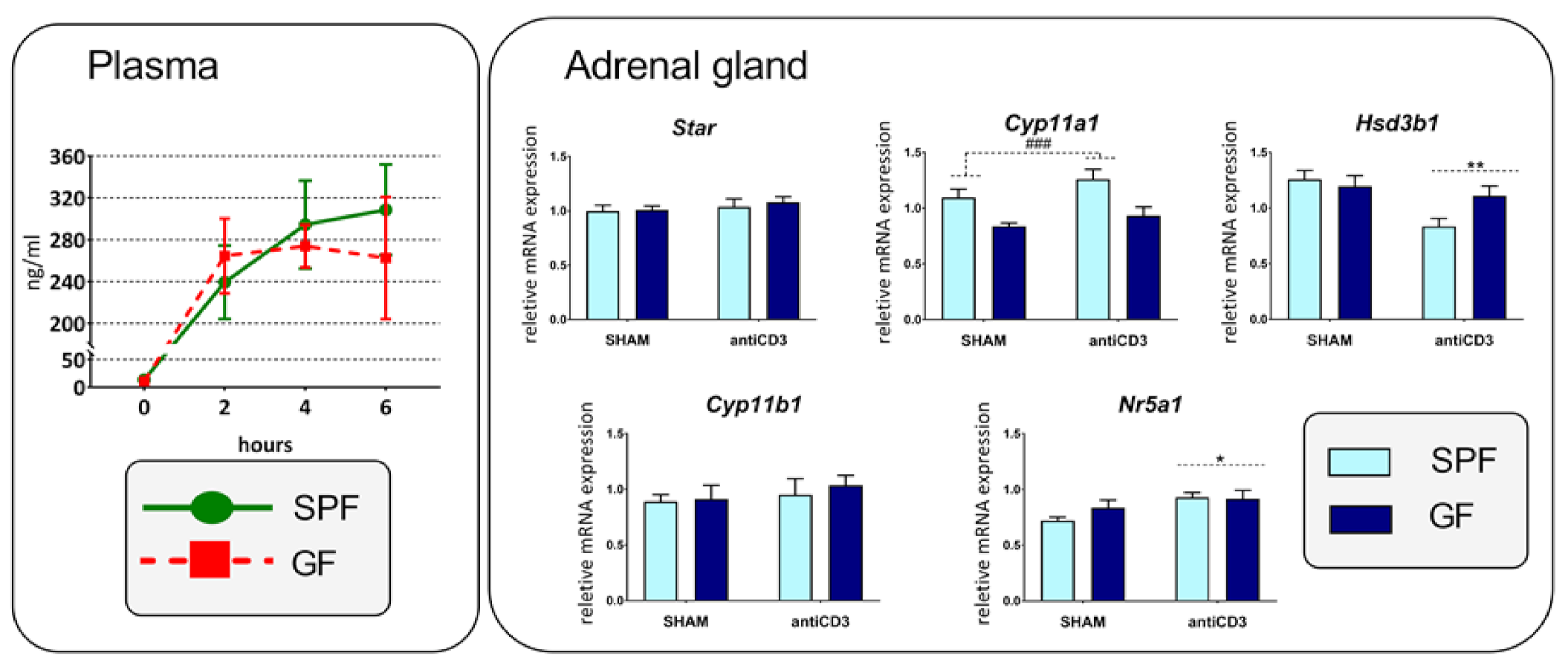
2.5. Response of Cytokines and HPA Axis to Acute Anti-CD3 Challenge
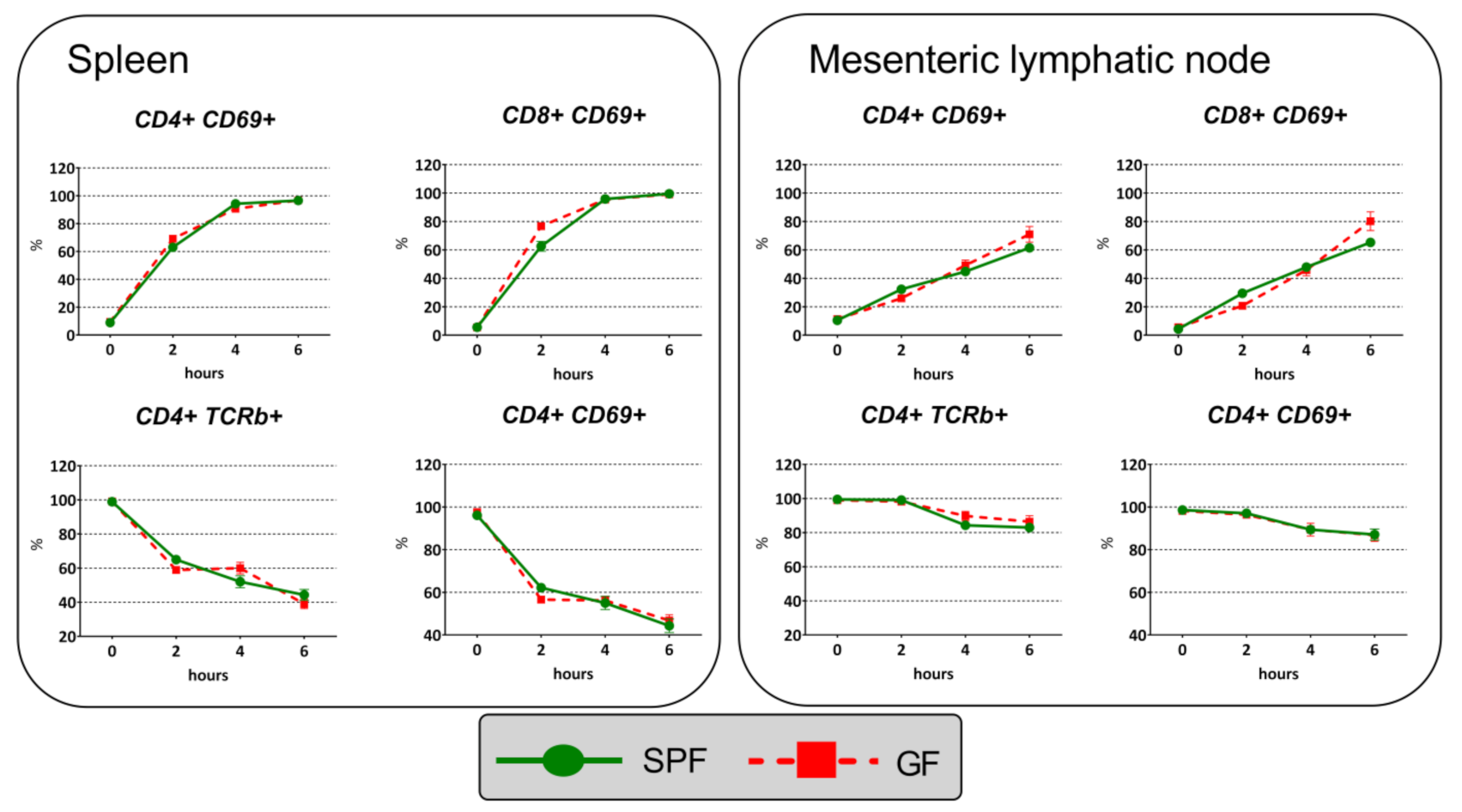
2.6. Immune Response to Anti-CD3 Antibody Injection
3. Discussion
4. Materials and Methods
4.1. Animals, Treatment and Sample Collection
4.2. Sample Preparation and Real-Time PCR
4.3. Steroidogenesis in Intestinal Explants
4.4. Quantification of Steroids Using MS Analysis
4.5. Corticosterone Assay
4.6. Flow Cytometric Analysis of Splenocytes and MLN Cells
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Giuffrè, M.; Moretti, R.; Campisciano, G.; da Silveira, A.B.M.; Monda, V.M.; Comar, M.; Di Bella, S.; Antonello, R.M.; Luzzati, R.; Crocè, L.S. You talking to me? Says the enteric nervous system (ENS) to the microbe. How intestinal microbes interact with the ENS. J. Clin. Med. 2020, 9, 3705. [Google Scholar] [CrossRef]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the microbiota-gut-brain axis: Prebiotics have anxiolytic and antidepressant-like effects and reverse the impact of chronic stress in mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Ait-Belgnaoui, A.; Durand, H.; Cartier, C.; Chaumaz, G.; Eutamene, H.; Ferrier, L.; Houdeau, E.; Fioramonti, J.; Bueno, L.; Theodorou, V. Prevention of gut leakiness by a probiotic treatment leads to attenuated HPA response to an acute psychological stress in rats. Psychoneuroendocrinology 2012, 37, 1885–1895. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cain, D.W.; Cidlowski, J.A. Immune regulation by glucocorticoids. Nat. Rev. Immunol. 2017, 17, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Schmidt, C.; Brunner, T. Extra-adrenal glucocorticoid synthesis in the intestinal mucosa: Between immune homeostasis and immune escape. Front. Immunol. 2019, 10, 1438. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Nikolakis, G.; Manna, P.R.; Skobowiat, C.; Zmijewski, M.; Li, W.; Janjetovic, Z.; Postlethwaite, A.; Zouboulis, C.C.; et al. Steroidogenesis in the skin: Implications for local immune functions. J. Steroid Biochem. Mol. Biol. 2013, 137, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Tiganescu, A.; Walker, E.A.; Hardy, R.S.; Mayes, A.E.; Stewart, P.M. Localization, age- and site-dependent expression, and regulation of 11β-hydroxysteroid dehydrogenase type 1 in skin. J. Investig. Dermatol. 2011, 131, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Cima, I.; Corazza, N.; Dick, B.; Fuhrer, A.; Herren, S.; Jakob, S.; Ayuni, E.; Mueller, C.; Brunner, T. Intestinal epithelial cells synthesize glucocorticoids and regulate T cell activation. J. Exp. Med. 2004, 200, 1635–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noti, M.; Corazza, N.; Mueller, C.; Berger, B.; Brunner, T. TNF suppresses acute intestinal inflammation by inducing local glucocorticoid synthesis. J. Exp. Med. 2010, 207, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Whorwood, C.B.; Ricketts, M.L.; Stewart, P.M. Epithelial cell localization of type 2 11β-hydroxysteroid dehydrogenase in rat and human colon. Endocrinology 1994, 135, 2533–2541. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Domenico, J.; Takeda, K.; Han, J.; Wang, M.; Armstrong, M.; Reisdorph, N.; O’Connor, B.P.; Lucas, J.J.; Gelfand, E.W. Steroidogenic enzyme Cyp11a1 regulates Type 2 CD8+ T cell skewing in allergic lung disease. Proc. Natl. Acad. Sci. USA 2013, 110, 8152–8157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahata, B.; Pramanik, J.; van der Weyden, L.; Polanski, K.; Kar, G.; Riedel, A.; Chen, X.; Fonseca, N.A.; Kundu, K.; Campos, L.S.; et al. Tumors induce de novo steroid biosynthesis in T cells to evade immunity. Nat. Commun. 2020, 11, 3588. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Ding, X.; Daynes, R.A. The expression of 11β-hydroxysteroid dehydrogenase type I by lymphocytes provides a novel means for intracrine regulation of glucocorticoid activities. J. Immunol. 2005, 174, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Ergang, P.; Vytáčková, K.; Švec, J.; Bryndová, J.; Mikšík, I.; Pácha, J. Upregulation of 11β-hydroxysteroid dehydrogenase 1 in lymphoid organs during inflammation in the rat. J. Steroid Biochem. Mol. Biol. 2011, 126, 19–25. [Google Scholar] [CrossRef]
- Noti, M.; Corazza, N.; Tuffin, G.; Schoonjans, K.; Brunner, T. Lipopolysaccharide induces intestinal glucocorticoid synthesis in a TNF-dependent manner. FASEB J. 2010, 24, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Bryndová, J.; Žbánková, S.; Kment, M.; Pácha, J. Colitis up-regulates local glucocorticoid activation and down-regulates inactivation in colonic tissue. Scand. J. Gastroenterol. 2004, 39, 549–553. [Google Scholar] [CrossRef]
- Huang, S.C.; Lee, C.T.; Chung, B.C. Tumor necrosis factor suppresses NR5A2 activity and intestinal glucocorticoid synthesis to sustain chronic colitis. Sci. Signal 2014, 7, ra20. [Google Scholar] [CrossRef]
- Ballegeer, M.; Van Looveren, K.; Timmermans, S.; Eggermont, M.; Vandevyver, S.; Thery, F.; Dendoncker, K.; Souffriau, J.; Vandewalle, J.; Van Wyngene, L.; et al. Glucocorticoid receptor dimers control intestinal STAT1 and TNF-induced inflammation in mice. J. Clin. Investig. 2018, 128, 3265–3279. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Jia, R.; Brunner, T. Local synthesis of immunosuppressive glucocorticoids in the intestinal epithelium regulates anti-viral immune responses. Cell. Immunol. 2018, 334, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, A.E.; Kipari, T.M.; Zhang, Z.; Esteves, C.L.; Lucas, C.D.; Gilmour, J.S.; Webster, S.P.; Walker, B.R.; Hughes, J.; Savill, J.S.; et al. 11β-Hydroxysteroid dehydrogenase type 1 is expressed in neutrophils and restrains an inflammatory response in male mice. Endocrinology 2016, 157, 2928–2936. [Google Scholar] [CrossRef] [PubMed]
- Vodička, M.; Ergang, P.; Hrnčíř, T.; Mikulecká, A.; Kvapilová, P.; Vagnerová, K.; Šestáková, B.; Fajstová, A.; Hermanová, P.; Hudcovic, T.; et al. Microbiota affects the expression of genes involved in HPA axis regulation and local metabolism of glucocorticoids in chronic psychosocial stress. Brain Behav. Immun. 2018, 73, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Vagnerová, K.; Vodička, M.; Hermanová, P.; Ergang, P.; Šrůtková, D.; Klusoňová, P.; Balounová, K.; Hudcovic, T.; Pácha, J. Interactions between gut microbiota and acute restraint stress in peripheral structures of the hypothalamic-pituitary-adrenal axis and the intestine of male mice. Front. Immunol. 2019, 10, 2655. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Opiyo, M.N.; Thomson, M.; Gharbi, K.; Seckl, J.R.; Heger, A.; Chapman, K.E. 11β-hydroxysteroid dehydrogenase-1 deficiency alters the gut microbiome response to Western diet. J. Endocrinol. 2017, 232, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Mukherji, A.; Kobiita, A.; Ye, T.; Chambon, P. Homeostasis in intestinal epithelium is orchestrated by the circadian clock and microbiota cues transduced by TLRs. Cell 2013, 153, 812–827. [Google Scholar] [CrossRef] [Green Version]
- Mueller, M.; Cima, I.; Noti, M.; Fuhrer, A.; Jakob, S.; Dubuquoy, L.; Schoonjans, K.; Brunner, T. The nuclear receptor LRH-1 critically regulates extra-adrenal glucocorticoid synthesis in the intestine. J. Exp. Med. 2006, 203, 2057–2062. [Google Scholar] [CrossRef]
- Valle, L.D.; Couët, J.; Labrie, Y.; Simard, J.; Belvedere, P.; Simontacchi, C.; Labrie, F.; Colombo, L. Occurrence of cytochrome P450c17 mRNA and dehydroepiandrosterone biosynthesis in the rat gastrointestinal tract. Mol. Cell. Endocrinol. 1995, 111, 83–92. [Google Scholar] [CrossRef]
- Reboldi, A.; Cyster, J.G. Peyer’s patches: Organizing B-cell responses at the intestinal frontier. Immunol. Rev. 2016, 271, 230–245. [Google Scholar] [CrossRef]
- Igaz, P.; Tömböl, Z.; Szabó, P.M.; Likó, I.; Rácz, K. Steroid biosynthesis inhibitors in the therapy of hypercortisolism: Theory and practice. Curr. Med. Chem. 2008, 15, 2734–2747. [Google Scholar] [CrossRef]
- Sampath-Kumar, R.; Yu, M.; Khalil, M.W.; Yang, K. Metyrapone is a competitive inhibitor of 11β-hydroxysteroid dehydrogenase type 1 reductase. J. Steroid Biochem. Mol. Biol. 1997, 62, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Marcolongo, P.; Senesi, S.; Gava, B.; Fulceri, R.; Sorrentino, V.; Margittai, E.; Lizák, B.; Csala, M.; Bánhegyi, G.; Benedetti, A. Metyrapone prevents cortisone-induced preadipocyte differentiation by depleting luminal NADPH of the endoplasmic reticulum. Biochem. Pharmacol. 2008, 76, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Ergang, P.; Vodička, M.; Vagnerová, K.; Moravec, M.; Kvapilová, P.; Kment, M.; Pácha, J. Inflammation regulates 11β-hydroxysteroid dehydrogenase type 1 differentially in specific compartments of the gut mucosal immune system. Steroids 2017, 126, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Vermeer, H.; Hendriks-Stegeman, B.I.; van der Burg, B.; van Buul-Offers, S.C.; Jansen, M. Glucocorticoid-induced increase in lymphocytic FKBP51 messenger ribonucleic acid expression: A potential marker for glucocorticoid sensitivity, potency, and bioavailability. J. Clin. Endocrinol. Metab. 2003, 88, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [Green Version]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 2013, 18, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Crumeyrolle-Arias, M.; Jaglin, M.; Bruneau, A.; Vancassel, S.; Cardona, A.; Daugé, V.; Naudon, L.; Rabot, S. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology 2014, 42, 207–217. [Google Scholar] [CrossRef]
- Menezes-Garcia, Z.; Arifa, R.D.D.N.; Acúrcio, L.; Brito, C.B.; Gouvea, J.O.; Lima, R.L.; Bastos, R.W.; Dias, A.C.F.; Antunes Dourado, L.P.; Bastos, L.F.S.; et al. Colonization by Enterobacteriaceae is crucial for acute inflammatory responses in murine small intestine via regulation of corticosterone production. Gut Microbes 2020, 11, 1531–1546. [Google Scholar] [CrossRef]
- Bouguen, G.; Langlois, A.; Djouina, M.; Branche, J.; Koriche, D.; Dewaeles, E.; Mongy, A.; Auwerx, J.; Colombel, J.F.; Desreumaux, P.; et al. Intestinal steroidogenesis controls PPARγ expression in the colon and is impaired during ulcerative colitis. Gut 2015, 64, 901–910. [Google Scholar] [CrossRef]
- Mueller, M.; Atanasov, A.; Cima, I.; Corazza, N.; Schoonjans, K.; Brunner, T. Differential regulation of glucocorticoid synthesis in murine intestinal epithelial versus adrenocortical cell lines. Endocrinology 2007, 148, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Vagnerová, K.; Kverka, M.; Klusoňová, P.; Ergang, P.; Mikšík, I.; Tlaskalová-Hogenová, H.; Pácha, J. Intestinal inflammation modulates expression of 11β-hydroxysteroid dehydrogenase in murine gut. J. Endocrinol. 2006, 191, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Žbánková, S.; Bryndová, J.; Leden, P.; Kment, M.; Švec, A.; Pácha, J. 11β-hydroxysteroid dehydrogenase 1 and 2 expression in colon from patients with ulcerative colitis. J. Gastroenterol. Hepatol. 2007, 22, 1019–1023. [Google Scholar] [CrossRef]
- Jun, Y.J.; Park, S.J.; Kim, T.H.; Lee, S.H.; Lee, K.J.; Hwang, S.M.; Lee, S.H. Expression of 11β-hydroxysteroid dehydrogenase 1 and 2 in patients with chronic rhinosinusitis and their possible contribution to local glucocorticoid activation in sinus mucosa. J. Allergy Clin. Immunol. 2014, 134, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Elustondo, P.; Martin, L.A.; Karten, B. Mitochondrial cholesterol import. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2017, 1862, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Ueno, N.; Hasebe, T.; Kaneko, A.; Yamamoto, M.; Fujiya, M.; Kohgo, Y.; Kono, T.; Wang, C.Z.; Yuan, C.S.; Bissonnette, M.; et al. TU-100 (Daikenchuto) and ginger ameliorate anti-CD3 antibody induced T cell-mediated murine enteritis: Microbe-independent effects involving Akt and NF-κB suppression. PLoS ONE 2014, 9, e97456. [Google Scholar] [CrossRef] [Green Version]
- Matysik, S.; Liebisch, G. Quantification of steroid hormones in human serum by liquid chromatography-high resolution tandem mass spectrometry. J. Chromatogr. A 2017, 1526, 112–118. [Google Scholar] [CrossRef]
- Olisov, D.; Lee, K.; Jun, S.H.; Song, S.H.; Kim, J.H.; Lee, Y.A.; Shin, C.H.; Song, J. Measurement of serum steroid profiles by HPLC-tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2019, 1117, 1–9. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ergang, P.; Vagnerová, K.; Hermanová, P.; Vodička, M.; Jágr, M.; Šrůtková, D.; Dvořáček, V.; Hudcovic, T.; Pácha, J. The Gut Microbiota Affects Corticosterone Production in the Murine Small Intestine. Int. J. Mol. Sci. 2021, 22, 4229. https://doi.org/10.3390/ijms22084229
Ergang P, Vagnerová K, Hermanová P, Vodička M, Jágr M, Šrůtková D, Dvořáček V, Hudcovic T, Pácha J. The Gut Microbiota Affects Corticosterone Production in the Murine Small Intestine. International Journal of Molecular Sciences. 2021; 22(8):4229. https://doi.org/10.3390/ijms22084229
Chicago/Turabian StyleErgang, Peter, Karla Vagnerová, Petra Hermanová, Martin Vodička, Michal Jágr, Dagmar Šrůtková, Václav Dvořáček, Tomáš Hudcovic, and Jiří Pácha. 2021. "The Gut Microbiota Affects Corticosterone Production in the Murine Small Intestine" International Journal of Molecular Sciences 22, no. 8: 4229. https://doi.org/10.3390/ijms22084229