
Nitro-Oleic Acid Inhibits Stemness Maintenance and Enhances Neural Differentiation of Mouse Embryonic Stem Cells via STAT3 Signaling

 , and
, and
Abstract
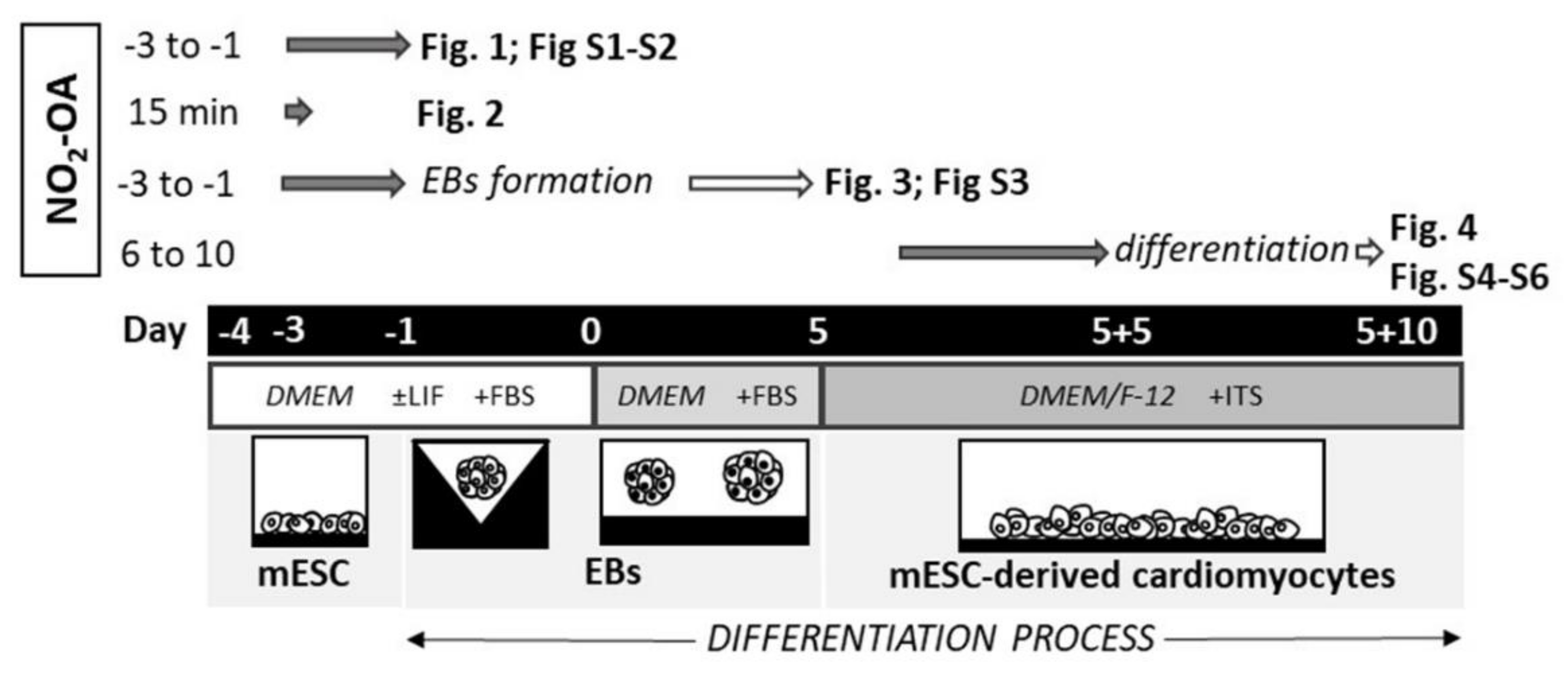
:1. Introduction
2. Results
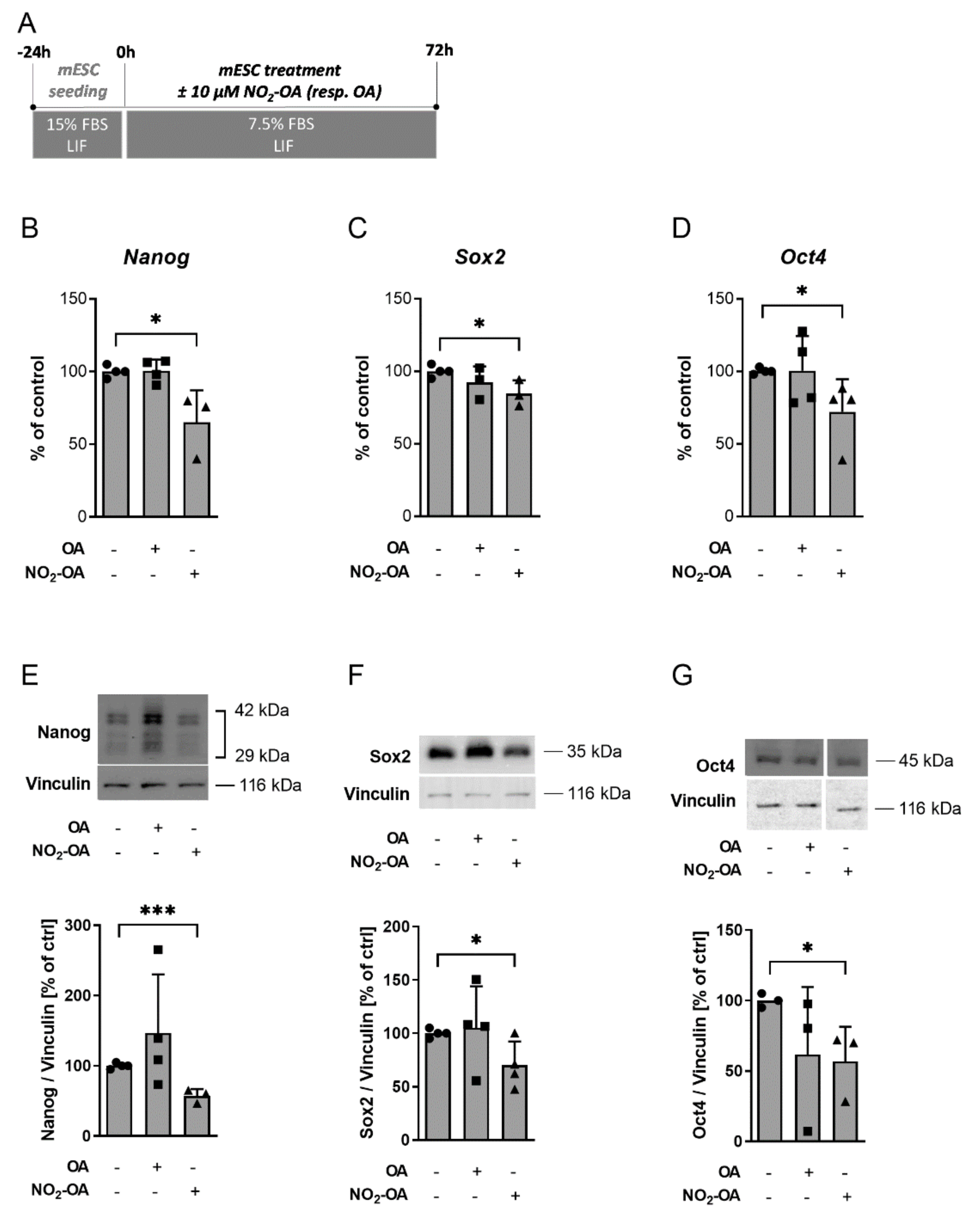
2.1. NO2-OA Significantly Inhibits Expression of Pluripotency Markers in mESC
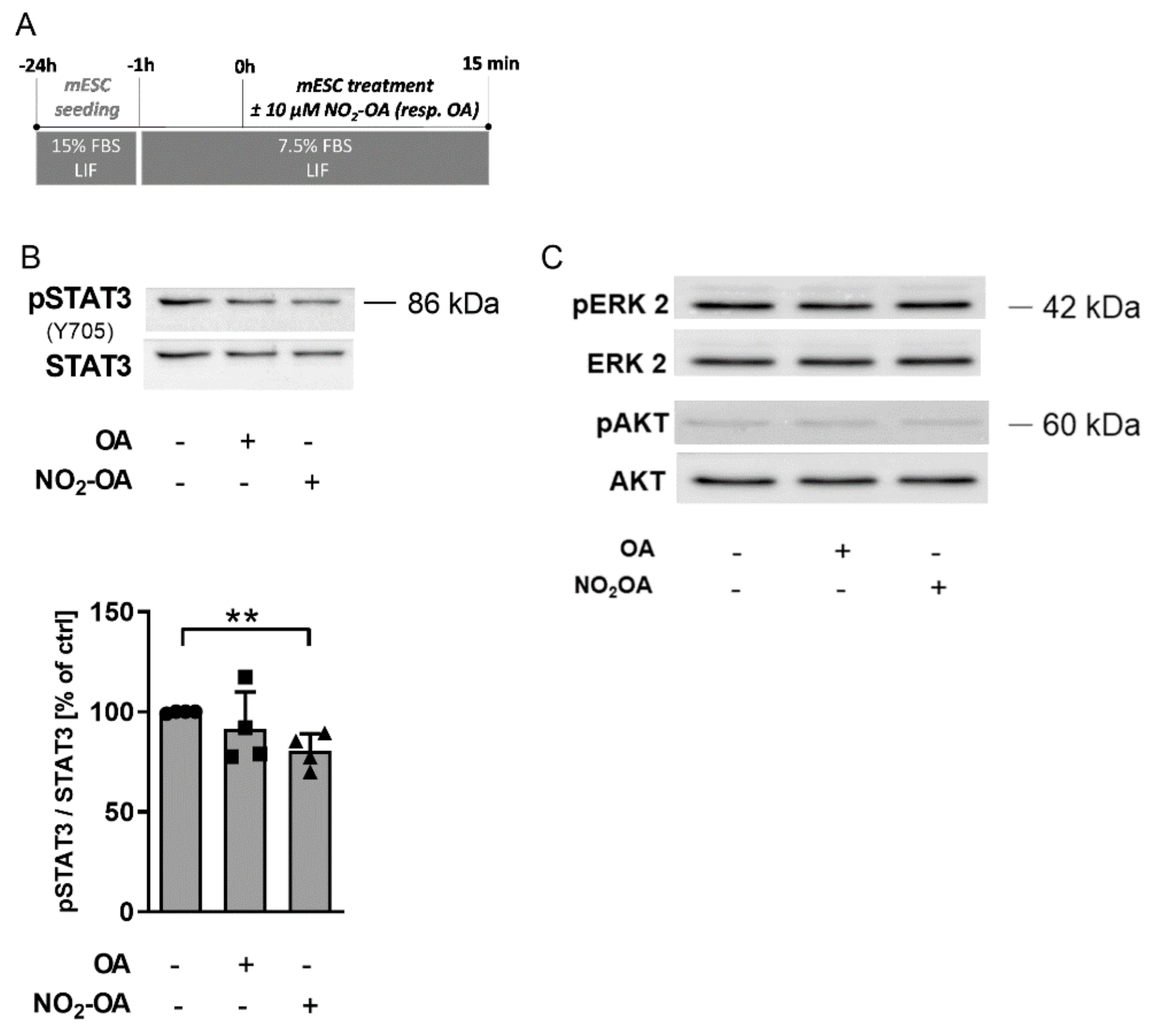
2.2. NO2-OA-Dependent STAT3 Signaling Regulation Is Responsible for the Reduction of mESC Stemness
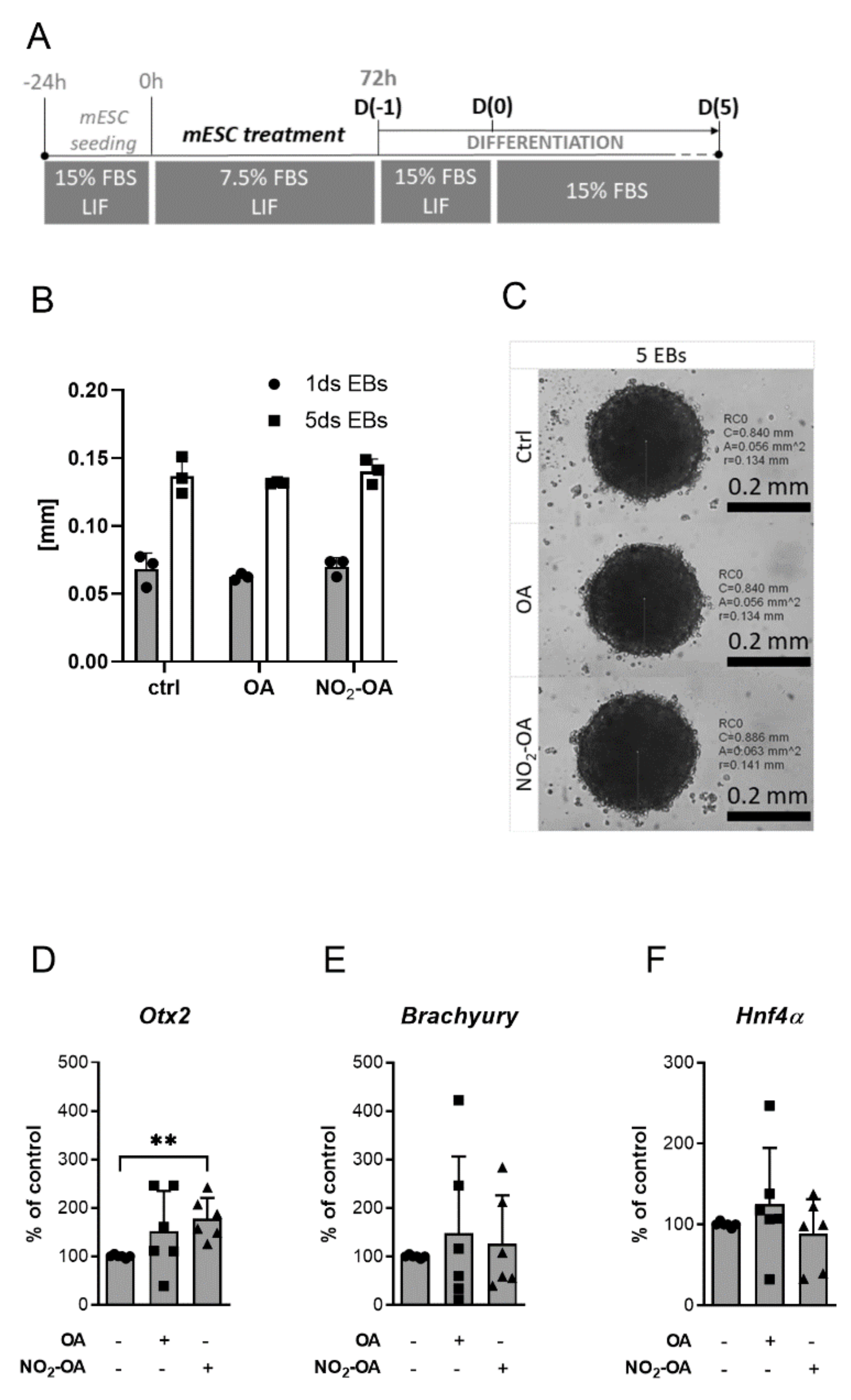
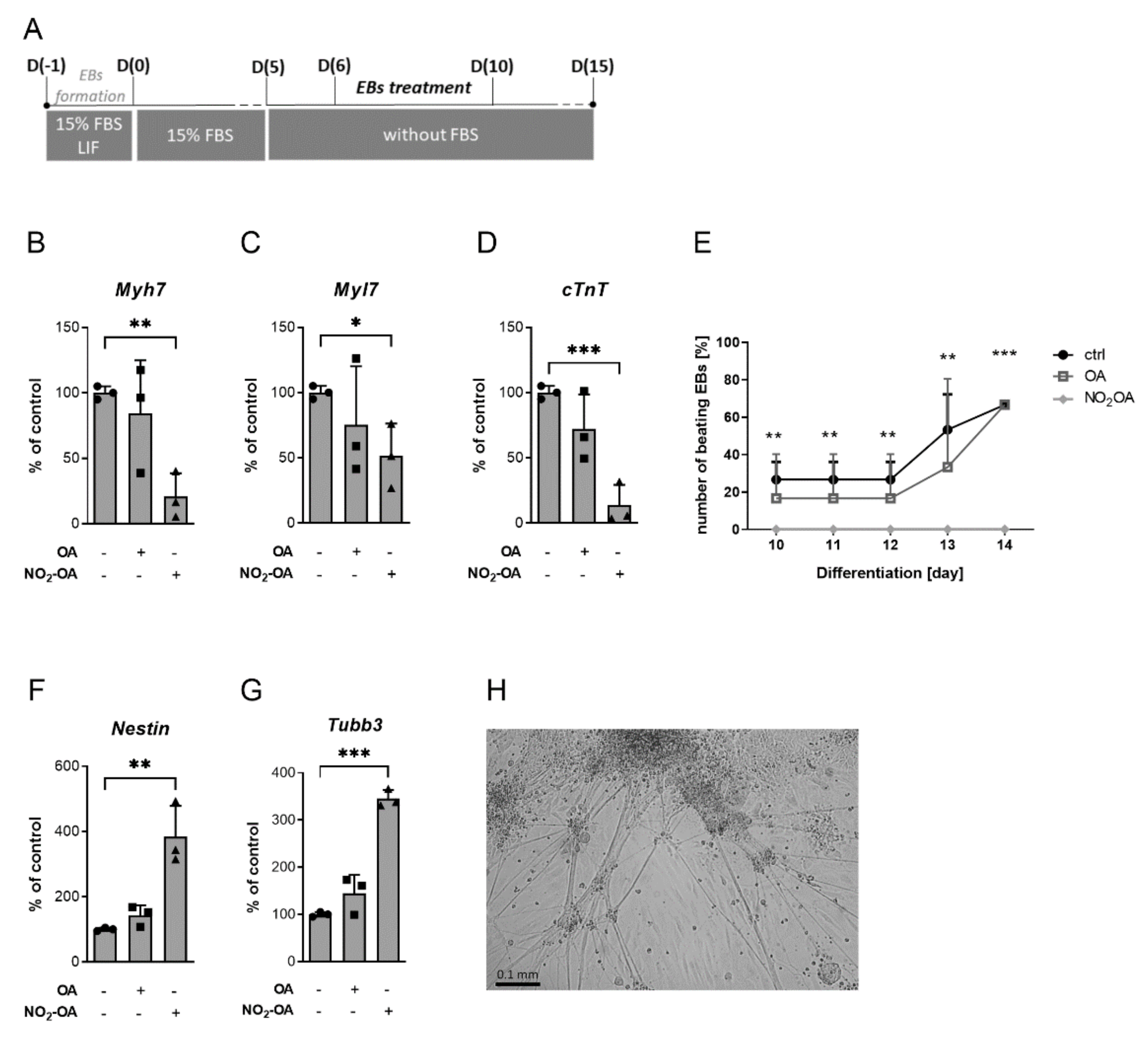
2.3. The Differentiation Process Is Changed by NO2-OA Treatment
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cultivation and Treatment of mESC
4.3. Differentiation of mESC
4.4. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
4.5. Protein Expression Analysis by Western Blot Technique
4.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Dedication
Acknowledgments
Conflicts of Interest
References
- He, R.; Xhabija, B.; Al-Qanber, B.; Kidder, B.L. OCT4 supports extended LIF-independent self-renewal and maintenance of transcriptional and epigenetic networks in embryonic stem cells. Sci. Rep. 2017, 7, 16360. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, T.; Wang, L.; Cai, Y.; Zhong, X.; He, X.; Hu, L.; Tian, S.; Wu, M.; Hui, L.; et al. Fatty acid synthesis is critical for stem cell pluripotency via promoting mitochondrial fission. EMBO J. 2017, 36, 1330–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucera, J.; Netusilova, J.; Sladecek, S.; Lanova, M.; Vasicek, O.; Stefkova, K.; Navratilova, J.; Kubala, L.; Pachernik, J. Hypoxia Downregulates MAPK/ERK but Not STAT3 Signaling in ROS-Dependent and HIF-1-Independent Manners in Mouse Embryonic Stem Cells. Oxid. Med. Cell. Longev. 2017, 2017, 4386947. [Google Scholar] [CrossRef] [PubMed]
- Storm, M.P.; Bone, H.K.; Beck, C.G.; Bourillot, P.Y.; Schreiber, V.; Damiano, T.; Nelson, A.; Savatier, P.; Welham, M.J. Regulation of Nanog expression by phosphoinositide 3-kinase-dependent signaling in murine embryonic stem cells. J. Biol. Chem. 2007, 282, 6265–6273. [Google Scholar] [CrossRef] [Green Version]
- Niwa, H.; Burdon, T.; Chambers, I.; Smith, A. Self-renewal of pluripotent embryonic stem cells is mediated via activation of STAT3. Genes Dev. 1998, 12, 2048–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, H.; Ohno, Y.; Toyoshima, Y.; Ohtake, J.; Homma, S.; Kawamura, H.; Takahashi, N.; Taketomi, A. Interleukin-6/STAT3 signaling as a promising target to improve the efficacy of cancer immunotherapy. Cancer Sci. 2017, 108, 1947–1952. [Google Scholar] [CrossRef]
- Chakraborty, D.; Sumova, B.; Mallano, T.; Chen, C.W.; Distler, A.; Bergmann, C.; Ludolph, I.; Horch, R.E.; Gelse, K.; Ramming, A.; et al. Activation of STAT3 integrates common profibrotic pathways to promote fibroblast activation and tissue fibrosis. Nat. Commun. 2017, 8, 1130. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Ye, S.; Zhou, X.; Liu, D.; Ying, Q.L. Molecular basis of embryonic stem cell self-renewal: From signaling pathways to pluripotency network. Cell. Mol. Life Sci. 2015, 72, 1741–1757. [Google Scholar] [CrossRef] [Green Version]
- Bourillot, P.Y.; Aksoy, I.; Schreiber, V.; Wianny, F.; Schulz, H.; Hummel, O.; Hubner, N.; Savatier, P. Novel STAT3 target genes exert distinct roles in the inhibition of mesoderm and endoderm differentiation in cooperation with Nanog. Stem Cells 2009, 27, 1760–1771. [Google Scholar] [CrossRef]
- Foshay, K.; Rodriguez, G.; Hoel, B.; Narayan, J.; Gallicano, G.I. JAK2/STAT3 directs cardiomyogenesis within murine embryonic stem cells in vitro. Stem Cells 2005, 23, 530–543. [Google Scholar] [CrossRef]
- Foshay, K.M.; Gallicano, G.I. Regulation of Sox2 by STAT3 initiates commitment to the neural precursor cell fate. Stem Cells Dev. 2008, 17, 269–278. [Google Scholar] [CrossRef]
- Huang, G.; Yan, H.; Ye, S.; Tong, C.; Ying, Q.L. STAT3 phosphorylation at tyrosine 705 and serine 727 differentially regulates mouse ESC fates. Stem Cells 2014, 32, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ji, L.; Ten, Y.; Wang, Y.; Pei, X. The molecular mechanism of embryonic stem cell pluripotency and self-renewal. Sci. China C Life Sci. 2007, 50, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Groenendyk, J.; Michalak, M. Disrupted WNT signaling in mouse embryonic stem cells in the absence of calreticulin. Stem Cell Rev. Rep. 2014, 10, 191–206. [Google Scholar] [CrossRef]
- Sinova, R.; Kudova, J.; Nesporova, K.; Karel, S.; Sulakova, R.; Velebny, V.; Kubala, L. Opioid receptors and opioid peptides in the cardiomyogenesis of mouse embryonic stem cells. J. Cell Physiol. 2019, 234, 13209–13219. [Google Scholar] [CrossRef] [PubMed]
- Ambrozova, G.; Fidlerova, T.; Verescakova, H.; Koudelka, A.; Rudolph, T.K.; Woodcock, S.R.; Freeman, B.A.; Kubala, L.; Pekarova, M. Nitro-oleic acid inhibits vascular endothelial inflammatory responses and the endothelial-mesenchymal transition. Biochim. Biophys. Acta 2016, 1860, 2428–2437. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, V.; Rudolph, T.K.; Schopfer, F.J.; Bonacci, G.; Woodcock, S.R.; Cole, M.P.; Baker, P.R.; Ramani, R.; Freeman, B.A. Endogenous generation and protective effects of nitro-fatty acids in a murine model of focal cardiac ischaemia and reperfusion. Cardiovasc. Res. 2010, 85, 155–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopfer, F.J.; Vitturi, D.A.; Jorkasky, D.K.; Freeman, B.A. Nitro-fatty acids: New drug candidates for chronic inflammatory and fibrotic diseases. Nitric Oxide 2018, 79, 31–37. [Google Scholar] [CrossRef]
- Zhang, J.; Villacorta, L.; Chang, L.; Fan, Z.; Hamblin, M.; Zhu, T.; Chen, C.S.; Cole, M.P.; Schopfer, F.J.; Deng, C.X.; et al. Nitro-oleic acid inhibits angiotensin II-induced hypertension. Circ. Res. 2010, 107, 540–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, B.A.; Baker, P.R.; Schopfer, F.J.; Woodcock, S.R.; Napolitano, A.; d’Ischia, M. Nitro-fatty acid formation and signaling. J. Biol. Chem. 2008, 283, 15515–15519. [Google Scholar] [CrossRef] [Green Version]
- Delmastro-Greenwood, M.; Freeman, B.A.; Wendell, S.G. Redox-dependent anti-inflammatory signaling actions of unsaturated fatty acids. Annu. Rev. Physiol. 2014, 76, 79–105. [Google Scholar] [CrossRef] [Green Version]
- Freeman, B.A.; O’Donnell, V.B.; Schopfer, F.J. The discovery of nitro-fatty acids as products of metabolic and inflammatory reactions and mediators of adaptive cell signaling. Nitric. Oxide 2018, 77, 106–111. [Google Scholar] [CrossRef]
- Yang, X.; Rodriguez, M.L.; Leonard, A.; Sun, L.; Fischer, K.A.; Wang, Y.; Ritterhoff, J.; Zhao, L.; Kolwicz, S.C., Jr.; Pabon, L.; et al. Fatty Acids Enhance the Maturation of Cardiomyocytes Derived from Human Pluripotent Stem Cells. Stem Cell Rep. 2019, 13, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Vecera, J.; Kudova, J.; Kucera, J.; Kubala, L.; Pachernik, J. Neural Differentiation Is Inhibited through HIF1alpha/beta-Catenin Signaling in Embryoid Bodies. Stem Cells Int. 2017, 2017, 8715798. [Google Scholar] [CrossRef]
- Saxena, S.; Choudhury, S.; Mohan, K.N. Reproducible differentiation and characterization of neurons from mouse embryonic stem cells. MethodsX 2020, 7, 101073. [Google Scholar] [CrossRef]
- Mukherjee, A.; Kenny, H.A.; Lengyel, E. Unsaturated Fatty Acids Maintain Cancer Cell Stemness. Cell Stem Cell 2017, 20, 291–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasumoto, Y.; Miyazaki, H.; Vaidyan, L.K.; Kagawa, Y.; Ebrahimi, M.; Yamamoto, Y.; Ogata, M.; Katsuyama, Y.; Sadahiro, H.; Suzuki, M.; et al. Inhibition of Fatty Acid Synthase Decreases Expression of Stemness Markers in Glioma Stem Cells. PLoS ONE 2016, 11, e0147717. [Google Scholar] [CrossRef]
- Tai, C.I.; Schulze, E.N.; Ying, Q.L. Stat3 signaling regulates embryonic stem cell fate in a dose-dependent manner. Biol. Open 2014, 3, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulrich, H.; do Nascimento, I.C.; Bocsi, J.; Tarnok, A. Immunomodulation in stem cell differentiation into neurons and brain repair. Stem Cell Rev. Rep. 2015, 11, 474–486. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene of Interest | Forward Primer 5′→3′ | Reverse Primer 5′→3′ | UPL Probe No. |
|---|---|---|---|
| Rpl13a | CATGAGGTCGGGTGGAAGTA | GCCTGTTTCCGTAACCTCAA | # 25 |
| Nanog | GCCTCCAGCAGATGCAAG | GGTTTTGAAACCAGGTCTTAACC | # 25 |
| Sox2 | ACGGCAGCTACAGCATGA | GACGTCGTAGCGGTGCAT | # 68 |
| Oct4 | GTTGGAGAAGGTGGAACCAA | CTCCTTCTGCAGGGCTTTC | # 95 |
| Otx2 | GGTATGGACTTGCTGCATCC | CGAGCTGTGCCCTAGTAAATG | # 55 |
| Brachyury | ACTGGTCTAGCCTCGGAGTG | TTGCTCACAGACCAGAGACTG | # 27 |
| Hnf4a | CAGCAATGGACAGATGTGTGA | TGGTGATGGCTGTGGAGTC | # 27 |
| Myh7 | AGATCCGAAAGCAACTGGAG | GGATCTTGCCCTCCTCGT | # 25 |
| Myl7 | CCCATCAACTTCACCGTCTT | AGGCACTCAGGATGGCTTC | # 7 |
| cTnT | GAGACAGACAGAGAGAGAGAAGAAGA | CACAGCTCCTTGGCCTTC | # 53 |
| Nestin | TCCCTTAGTCTGGAAGTGGCTA | GGTGTCTGCAAGCGAGAGTT | # 67 |
| Tubb3 | GCGCATCAGCGTATACTACAA | CATGGTTCCAGGTTCCAAGT | # 81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereckova, J.; Pekarova, M.; Szamecova, N.; Hoferova, Z.; Kamarytova, K.; Falk, M.; Perecko, T. Nitro-Oleic Acid Inhibits Stemness Maintenance and Enhances Neural Differentiation of Mouse Embryonic Stem Cells via STAT3 Signaling. Int. J. Mol. Sci. 2021, 22, 9981. https://doi.org/10.3390/ijms22189981
Pereckova J, Pekarova M, Szamecova N, Hoferova Z, Kamarytova K, Falk M, Perecko T. Nitro-Oleic Acid Inhibits Stemness Maintenance and Enhances Neural Differentiation of Mouse Embryonic Stem Cells via STAT3 Signaling. International Journal of Molecular Sciences. 2021; 22(18):9981. https://doi.org/10.3390/ijms22189981
Chicago/Turabian StylePereckova, Jana, Michaela Pekarova, Nikoletta Szamecova, Zuzana Hoferova, Kristyna Kamarytova, Martin Falk, and Tomas Perecko. 2021. "Nitro-Oleic Acid Inhibits Stemness Maintenance and Enhances Neural Differentiation of Mouse Embryonic Stem Cells via STAT3 Signaling" International Journal of Molecular Sciences 22, no. 18: 9981. https://doi.org/10.3390/ijms22189981