
31P-NMR Metabolomics Revealed Species-Specific Use of Phosphorous in Trees of a French Guiana Rainforest

 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Plots
2.3. Sample Collection
2.4. Nutrient Pools
2.5. One Dimensional 31P-NMR Analysis
2.6. Statistical Analyses
3. Results
3.1. Phosphorus Compounds Identification
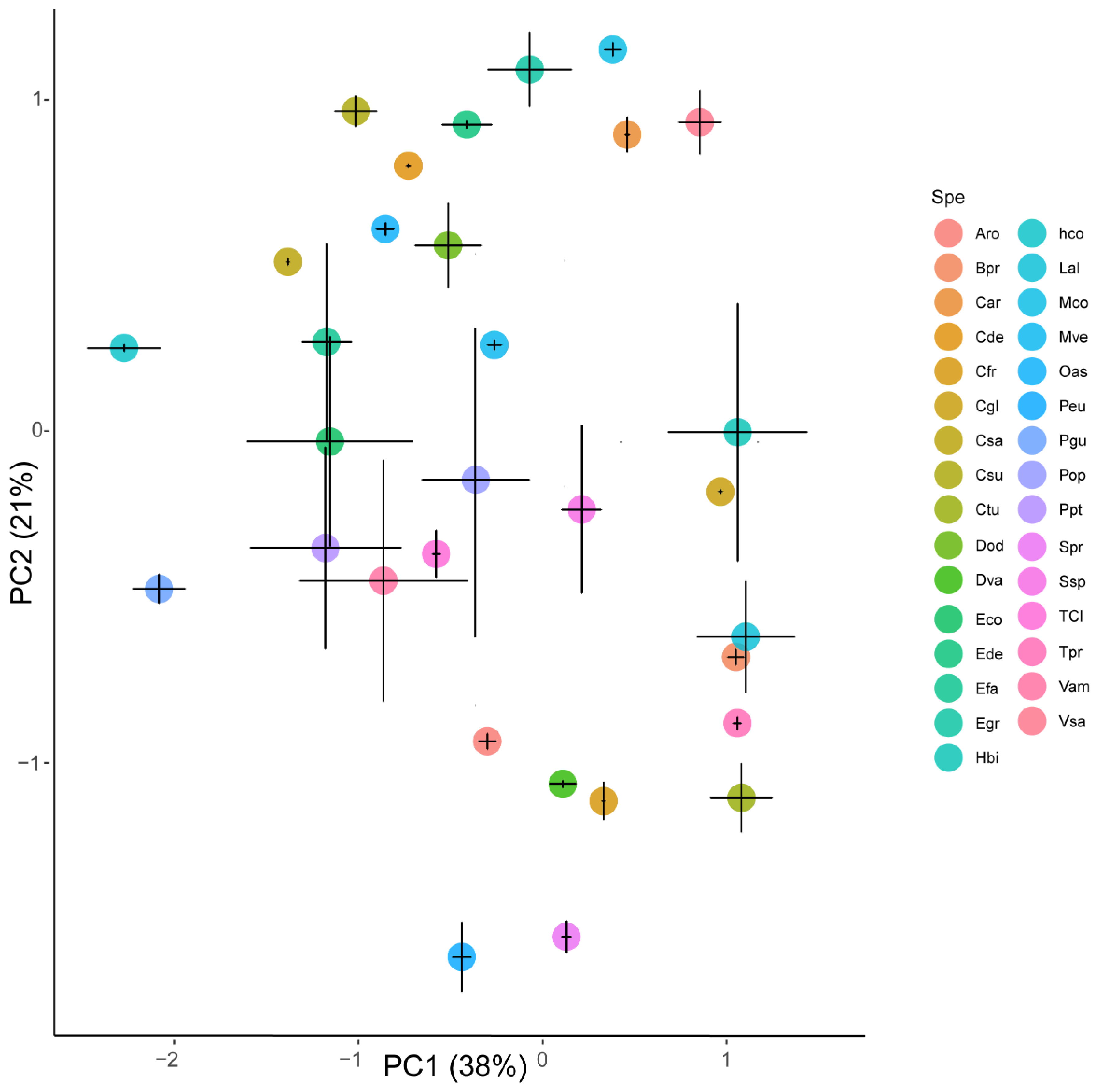
3.2. Phosphorus Compounds and Elements Among Species
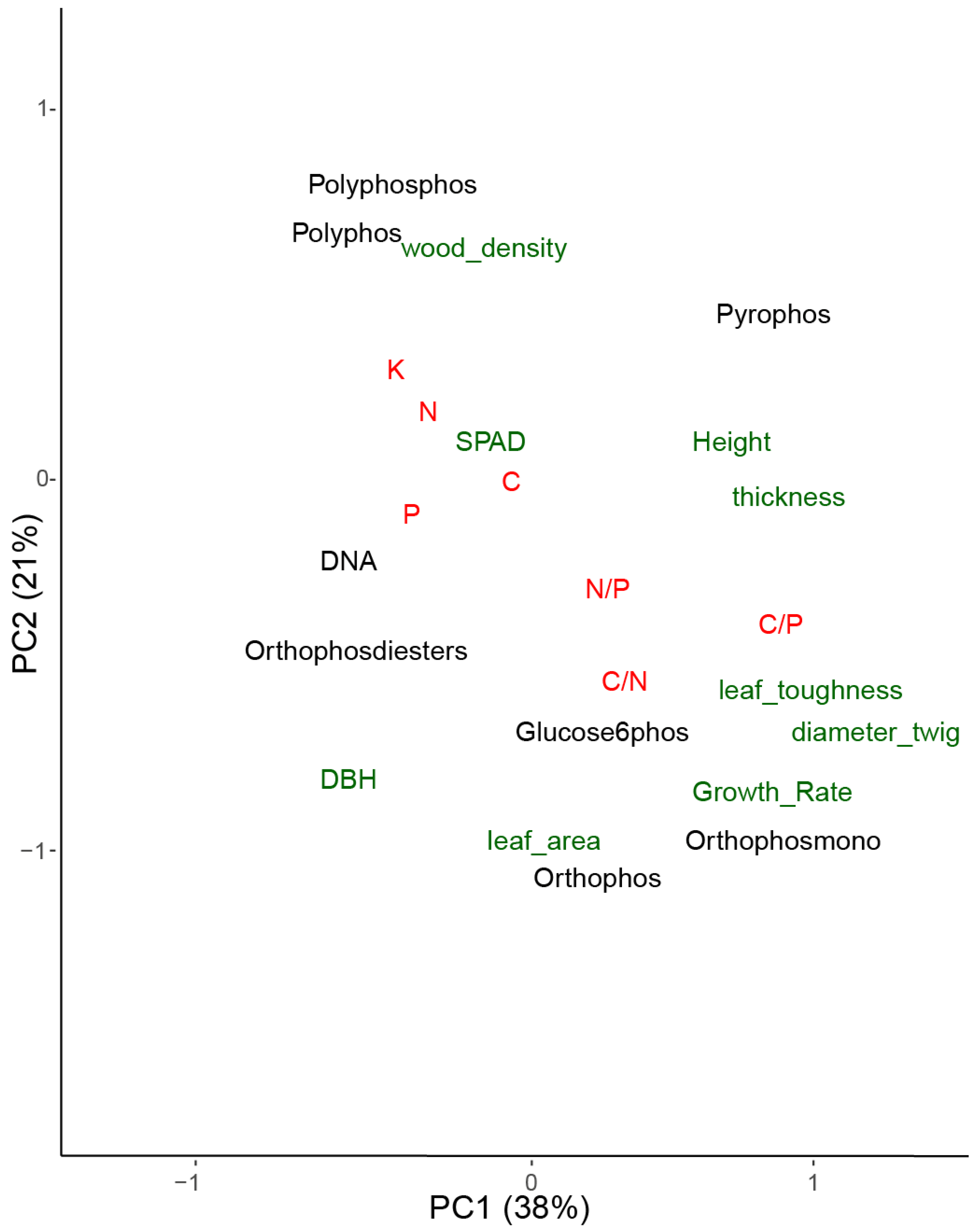
3.3. Phosphorus Compounds and Elements
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vos, J.; Biemond, H. Effects of nitrogen on the development and growth of the potato plant. 1. Leaf appearance, expansion growth, life spans of leaves and stem branching. Ann. Bot. 1992, 70, 27–35. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, E.V.J.; Vitousek, P.M.; Cuevas, E. experimental investigation of nutrient limitation of forest growth on wet tropical mountains. Ecology 1998, 79, 10–22. [Google Scholar] [CrossRef]
- Walker, T.W.; Syers, J.K. The fate of phosphorus during pedogenesis. Geoderma 1976, 15, 1–19. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Rapid nutrient cycling in leaf litter from invasive plants in Hawai. Oecologia 2004, 141, 612–619. [Google Scholar] [CrossRef]
- Wardle, D.A.; Walker, L.R.; Bardgett, R.D. Ecosystem properties and forest decline in contrasting long-term. Science 2004, 305, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Kichenin, E.; Wardle, D.A.; Peltzer, D.A.; Morse, C.W.; Freschet, G.T. Contrasting effects of plant inter- and intraspecific variation on community-level trait measures along an environmental gradient. Funct. Ecol. 2013, 27, 1254–1261. [Google Scholar] [CrossRef]
- Turner, B.L.; Brenes-Arguedas, T.; Condit, R. Pervasive phosphorus limitation of tree species but not communities in tropical forests. Nature 2018, 555, 367–370. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Sanford, R.L., Jr. Nutrient cycling in moist tropical forest. Ann. Rev. Ecol. Syst. 1986, 17, 137–167. [Google Scholar] [CrossRef]
- Vitousek, P.M. Nutrient Cycling and Limitation: Hawai‘i as a Model System; Princeton University Press: Princeton, NJ, USA, 2004. [Google Scholar]
- Corlett, R.T.; Primack, R.B. Tropical rainforests and the need for cross-continental comparisons. Trends Ecol. Evol. 2006, 21, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Aiba, S.I.; Sawada, Y.; Takyu, M.; Eino, T.; Kitayama, K.; Repin, R. Structure, floristics and diversity of tropical montane rain forests over ultramafic soils on Mount Kinabalu (Borneo) compared with those on non-ultramafic soils. Aust. J. Bot. 2015, 63, 191–203. [Google Scholar] [CrossRef]
- John, R.; Dalling, J.W.; Harms, K.E.; Yavitt, J.B.; Stallard, R.F.; Mirabello, M.; Hubbell, S.P.; Valencia, R.; Navarrete, H.; Vallejo, M.; et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. USA 2007, 104, 864–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennington, R.T.; Lavin, M.; Oliveira-Filho, A. woody plant diversity, evolution, and ecology in the tropics: Perspectives from seasonally dry tropical forests. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 437–457. [Google Scholar] [CrossRef] [Green Version]
- Matos, F.A.R.; Magnago, L.F.S.; Gastauer, M.; Carreiras, J.M.B.; Simonelli, M.; Meira-Neto, J.A.A.; Edwards, D.P. Effects of landscape configuration and composition on phylogenetic diversity of trees in a highly fragmented tropical forest. J. Ecol. 2017, 105, 265–276. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Shibata, M.; Kitajima, K.; Ichie, T.; Kitayama, K.; Turner, B.L. Plant–soil interactions maintain biodiversity and functions of tropical forest ecosystems. Ecol. Res. 2017, 33, 1–12. [Google Scholar] [CrossRef]
- Xu, W.; Ci, X.; Song, C.; He, T.; Zhang, W.; Li, Q.; Li, J. Soil phosphorus heterogeneity promotes tree species diversity and phylogenetic clustering in a tropical seasonal rainforest. Ecol. Evol. 2016, 6, 8719–8726. [Google Scholar] [CrossRef]
- Martins, K.G.; Marques, M.C.M.; dos Santos, E.; Marques, R. Effects of soil conditions on the diversity of tropical forests across a successional gradient. For. Ecol. Manage. 2015, 349, 4–11. [Google Scholar] [CrossRef]
- Mandl, N.A.; Kessler, M.; Robbert Gradstein, S. Effects of environmental heterogeneity on species diversity and composition of terrestrial bryophyte assemblages in tropical montane forests of southern Ecuador. Plant. Ecol. Divers. 2009, 2, 313–321. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A.; Read, J.M. Edaphic variation and the mesoscale distribution of tree species in a neotropical rain forest. J. Ecol. 1998, 86, 101–112. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, J.X. On the factors that promote the diversity of herbivorous insects and plants in tropical forests. Proc. Natl. Acad. Sci. USA 2015, 112, 6098–6103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidaka, A.; Kitayama, K. Allocation of foliar phosphorus fractions and leaf traits of tropical tree species in response to decreased soil phosphorus availability on Mount Kinabalu, Borneo. J. Ecol. 2011, 99, 849–857. [Google Scholar] [CrossRef]
- Lambers, H.; Finnegan, P.M.; Laliberté, E.; Pearse, S.J.; Ryan, M.H.; Shane, M.W.; Veneklaas, E.J. Phosphorus nutrition of Proteaceae in severely phosphorus-impoverished soils: Are there lessons to be learned for future crops? Plant. Physiol. 2011, 156, 1058–1066. [Google Scholar] [CrossRef] [Green Version]
- Chapin III, F.S.; Kedrowski, R.A. Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees. Ecology 1983, 64, 376–391. [Google Scholar] [CrossRef]
- Ågren, G.I.; Wetterstedt, J.; Billberger, M.F.K. Nutrient limitation on terrestrial plant growth--modeling the interaction between nitrogen and phosphorus. New Phytol. 2012, 194, 953–960. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.-R.; Shane, M.W.; White, P.J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef]
- Elser, J.J.; Brien, W.J.O.; Dobberfuhl, D.R.; Dowling, T.E. The evolution of ecosystem processes: Growth rate and elemental stoichiometry of a key herbivore in temperate and arctic habitats. J. Evol. Biol. 2000, 13, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Schlüter, U.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Weber, A.P.M.; Zellerhoff, N.; Bucher, M.; Fahnenstich, H.; Sonnewald, U. Adaptation of maize source leaf metabolism to stress related disturbances in carbon, nitrogen and phosphorus balance. BMC Genom. 2013, 14, 442. [Google Scholar] [CrossRef] [Green Version]
- Turner, B.L.; Mahieu, N.; Condron, L.M. Phosphorus-31 nuclear magnetic resonance spectral assignments of phosphorus compounds in soil NaOH–EDTA extracts. Soil Sci. Soc. Am. J. 2003, 67, 497–510. [Google Scholar] [CrossRef]
- Cade-Menun, B.J.; Liu, C.W.; Nunlist, R.; McColl, J.G. Soil and litter Phosphorus-31 nuclear magnetic resonance spectroscopy. J. Environ. Qual. 2002, 31, 457–465. [Google Scholar] [PubMed]
- Vincent, A.G.; Turner, B.L.; Tanner, E.V.J. Soil organic phosphorus dynamics following perturbation of litter cycling in a tropical moist forest. Eur. J. Soil Sci. 2010, 61, 48–57. [Google Scholar] [CrossRef]
- Vestergren, J.; Vincent, A.G.; Jansson, M.; Persson, P.; Ilstedt, U.; Giesler, R. High-resolution characterization of organic phosphorus in soil extracts using 2D 1 H−31 P NMR correlation spectroscopy. Environ. Sci. Technol. 2012, 46, 3950–3956. [Google Scholar] [CrossRef]
- Me Chave, J.R.; Ra, B.R.; Dubois, M.-A. Estimation of biomass in a neotropical forest of French Guiana: Spatial and temporal variability. J. Trop. Ecol. 2001, 17, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Gourlet-Fleury, S.; Guehl, J.-M.; Laroussinie, O. Ecology and Management of a Neotropical Rainforest: Lessons Drawn from Paracou, a Long-Term Experimental Research Site in French Guiana; Elseiver: Paris, France, 2004. [Google Scholar]
- Courtois, E.A.; Stahl, C.; Van den Berge, J.; Bréchet, L.; Van Langenhove, L.; Richter, A.; Urbina, I.; Soong, J.L.; Peñuelas, J.; Janssens, I.A. Spatial variation of soil CO2, CH4 and N2O fluxes across topographical positions in tropical forests of the guiana shield. Ecosystems 2018, 21, 1445–1458. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Peñuelas, J.; Rivas-Ubach, A. Ecological metabolomics: Overview of current developments and future challenges. Chemoecology 2011, 21, 191–225. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Pérez-Trujillo, M.; Sardans, J.; Gargallo-Garriga, A.; Parella, T.; Peñuelas, J. Ecometabolomics: Optimized NMR-based method. Methods Ecol. Evol. 2013, 4, 464–473. [Google Scholar] [CrossRef]
- Deborde, C.; Moing, A.; Roch, L.; Jacob, D.; Rolin, D.; Giraudeau, P. Plant metabolism as studied by NMR Spectrosc. Prog. Nucl. Magn. Reson. Spectrosc. 2017, 102–103, 61–97. [Google Scholar] [CrossRef]
- Wilson, I.D.; Plumb, R.; Granger, J.; Major, H.; Williams, R.; Lenz, E.M. HPLC-MS-based methods for the study of metabonomics. J. Chromatogr. B 2005, 817, 67–76. [Google Scholar] [CrossRef]
- Cade-Menun, B.J.; Preston, C.M. A comparison of soil extraction procedures for 31P NMR spectroscopy. Soil Sci. 1996, 161, 770–785. [Google Scholar] [CrossRef]
- Makarov, M.I.; Haumaier, L.; Zech, W.; Marfenina, O.E.; Lysak, L.V. Can 31P NMR spectroscopy be used to indicate the origins of soil organic phosphates? Soil Biol. Biochem. 2005, 37, 15–25. [Google Scholar] [CrossRef]
- Turner, B.L. Soil organic phosphorus in tropical forests: An assessment of the NaOH-EDTA extraction procedure for quantitative analysis by solution 31P NMR spectroscopy. Eur. J. Soil Sci. 2008, 59, 453–466. [Google Scholar] [CrossRef]
- Smernik, R.J.; Doolette, A.L.; Marschner, P.; Bu, E.K.; Stonor, R.; Wakelin, S.A.; Mcneill, A.M. Soil biology & biochemistry forms of phosphorus in bacteria and fungi isolated from two Australian Soils. Soil Biol. Biochem. 2008, 40, 1908–1915. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA + for PRI-MER: Guide to Software and Statistical Methods; Primer-E: Plymouth, UK, 2008. [Google Scholar]
- Wright, S.J. Plant diversity in tropical forests: A review of mechanisms of species coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Williams, W.T.; Lance, G.N.; Webb, L.J.; Tracey, J.G.; Connell, J.H. Studies in the numerical analysis of complex rain-forest communities: IV. A method for the elucidation of small-scale forest pattern. J. Ecol. 1969, 57, 635–654. [Google Scholar] [CrossRef]
- Whitmore, T.C. Tropical Rain Forests of the Far East; Clarendon: Oxford, UK, 1975. [Google Scholar]
- Richards, P.W. The Tropical Rain Forest; An Ecological Study; Cambridge Univ. Press: New York, NY, USA, 1952. [Google Scholar]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Ogaya, R.; Estiarte, M. Nutrient stoichiometric relations and biogeochemical niche in coexisting plant species: Effect of simulated climate change. Pol. J. Ecol. 2008, 56, 613–622. [Google Scholar]
- Gargallo-Garriga, A.; Sardans, J.; Granda, V.; Llusià, J.; Peguero, G.; Asensio, D.; Ogaya, R.; Urbina, I.; Van Langenhove, L.; Verryckt, L.T.; et al. Different “metabolomic niches” of the highly diverse tree species of the French Guiana rainforests. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Penuelas, J.; Sardans, J.; Llusià, J.; Owen, S.M.; Carnicer, J.; Giambelluca, T.W.; Rezende, E.L.; Waite, M.; Niinemets, Ü. Faster returns on ‘leaf economics’ and different biogeochemical niche in invasive compared with native plant species. Glob. Chang. Biol. 2010, 16, 2171–2185. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Fernández-Martinez, M.; Ciais, P.; Jou, D.; Piao, S.; Obersteiner, M.; Vicca, S.; Janssens, I.A.; Sardans, J. The bioelements, the elementome, and the biogeochemical niche. Ecology 2019, 100, e02652. [Google Scholar] [CrossRef]
- Sardans, J.; Janssens, I.A.; Alonso, R.; Veresoglou, S.D.; Rillig, M.C.; Sanders, T.G.; Carnicer, J.; Filella, I.; Farré-Armengol, G.; Peñuelas, J. Foliar elemental composition of European forest tree species associated with evolutionary traits and present environmental and competitive conditions. Glob. Ecol. Biogeogr. 2015, 24, 240–255. [Google Scholar] [CrossRef] [Green Version]
- Urbina, I.; Sardans, J.; Grau, O.; Beierkuhnlein, C.; Jentsch, A.; Kreyling, J.; Peñuelas, J. Plant community composition affects the species biogeochemical niche. Ecosphere 2017, 8, e01801. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E. Environmental heterogeneity and plant species diversity: A hypothesis. Am. Nat. 1977, 111, 376–381. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds http://globalecology.creaf.cat/ are available from the authors. |



{kind=link}
{kind=link}
| Source | df | SS | MS | F | R2 | p |
|---|---|---|---|---|---|---|
| Species | 33 | 10206396 | 309,285 | 3.08 | 0.67 | 0.001 |
| Residuals | 49 | 4914344 | 100,293 | 0.33 | ||
| Total | 82 | 15120740 | 1 |
| Species | Total P (%, D.W.) | Total C (%, D.W.) | Total N (%, D.W.) | C:N | C:P | N:P | K (%, D.W.) | Phosphorus Species Distribution, % of Total P in Extract | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Polyphos | Orthophos | Glucose-6phos | Orthophos-mono | Orthophos-diesters | DNA | Pyrophos | Poly-P | ||||||||
| Licania alba | 0.04 | 48.59 | 1.44 | 34.7 | 1151.91 | 33.17 | 0.35 | 3.55 | 7.11 | 0 | 14.12 | 28.57 | 16.57 | 17.30 | 12.79 |
| Vouacapoua americana | 0.07 | 50.2 | 1.99 | 25.6 | 687.46 | 26.84 | 0.67 | 1.67 | 8.57 | 24.17 | 22.41 | 15.52 | 6.75 | 13.92 | 6.99 |
| Chrysophyllum argenteum | 0.03 | 49.7 | 1.34 | 37.2 | 1435.60 | 38.59 | 0.40 | 40.25 | 0 | 0 | 11.77 | 11.57 | 16.82 | 0 | 19.60 |
| Oxandra asbeckii | 0.05 | 48.1 | 1.94 | 24.8 | 1042.83 | 42.13 | 1.69 | 24.25 | 63.24 | 0 | 0.65 | 1.30 | 10.04 | 0 | 0.52 |
| Hirtella bicornis | 0.04 | 47.1 | 1.32 | 35.9 | 1215.23 | 33.94 | 0.75 | 0 | 0 | 0 | 26.38 | 13.88 | 9.03 | 25.13 | 25.58 |
| Tovomita sp. | 0.07 | 49.3 | 1.61 | 30.6 | 670.28 | 21.91 | 0.72 | 0 | 6.65 | 16.55 | 6.99 | 16.12 | 22.00 | 0 | 31.70 |
| Moronobea coccinea | 0.03 | 48.8 | 1.28 | 38.4 | 1469.22 | 38.49 | 0.27 | 5.88 | 0 | 0 | 1.82 | 46.14 | 23.60 | 0 | 22.55 |
| Eschweilera coriacea | 0.07 | 50.0 | 1.71 | 30.7 | 975.27 | 30.72 | 0.50 | 1.60 | 16.35 | 10.46 | 17.44 | 22.40 | 16.29 | 8.91 | 6.56 |
| Hymanea courbaril | 0.12 | 48.5 | 1.91 | 25.3 | 412.80 | 16.29 | 0.87 | 42.19 | 38.05 | 3.09 | 0 | 0 | 11.35 | 5.31 | 0 |
| Eschweilera decolorans | 0.05 | 50.0 | 1.77 | 28.2 | 1103.39 | 39.14 | 0.58 | 32.27 | 0 | 0 | 9.36 | 27.51 | 6.00 | 0 | 24.85 |
| Capirona decorticans | 0.05 | 48.7 | 2.08 | 23.4 | 1077.96 | 46.08 | 0.41 | 0 | 0 | 0 | 11.37 | 53.25 | 32.62 | 0 | 2.77 |
| Pouteria eugeniifolia | 0.08 | 48.6 | 2.89 | 16.9 | 650.54 | 38.58 | 0.69 | 0 | 35.00 | 0 | 15.72 | 19.61 | 19.61 | 0 | 10.06 |
| Eperua falcata | 0.08 | 49.6 | 1.70 | 30.1 | 687.85 | 23.49 | 0.63 | 7.37 | 0 | 6.41 | 25.85 | 43.73 | 10.22 | 0 | 6.41 |
| Catostemma fragrans | 0.03 | 25.2 | 0.90 | 14.0 | 455.01 | 16.29 | 0.28 | 0 | 49.45 | 2.50 | 28.06 | 0 | 20 | 0 | 0 |
| Caryocar glabrum | 0.05 | 52.4 | 1.34 | 39.2 | 1009.50 | 25.75 | 0.36 | 0 | 22.91 | 0 | 20.06 | 20.06 | 32.87 | 3.59 | 0.51 |
| Eperua grandiflora | 0.04 | 50.5 | 1.39 | 37.8 | 1289.63 | 34.47 | 0.34 | 6.39 | 7.22 | 6.46 | 25.13 | 19.73 | 12.13 | 18.70 | 4.25 |
| Paloue guianensis | 0.05 | 52.0 | 1.76 | 29.5 | 1086.24 | 36.77 | 0.22 | 2.21 | 0 | 19.81 | 7.14 | 21.05 | 21.63 | 28.16 | 0 |
| Dipteryx odorata | 0.06 | 49.4 | 1.38 | 35.9 | 891.48 | 24.81 | 0.89 | 0.31 | 12.79 | 8.35 | 4.57 | 16.65 | 3.13 | 0 | 54.19 |
| Protium opacum | 0.06 | 47.7 | 1.25 | 38.5 | 856.32 | 22.39 | 0.67 | 14.49 | 32.93 | 0 | 40.44 | 0.80 | 7.12 | 0 | 4.21 |
| Talisia praealta | 0.04 | 48.9 | 1.23 | 39.9 | 1203.11 | 30.12 | 0.21 | 0 | 0 | 0 | 10.14 | 18.78 | 18.78 | 23.31 | 28.98 |
| Bocoa prouacensis | 0.04 | 48.8 | 1.66 | 31.0 | 1471.96 | 47.02 | 0.35 | 0 | 27.66 | 14.21 | 14.21 | 4.61 | 2.66 | 0.46 | 36.19 |
| Sterculia pruriens | 0.07 | 46.9 | 1.42 | 33.2 | 686.14 | 20.66 | 1.14 | 0 | 19.99 | 22.74 | 44.54 | 0 | 12.73 | 0 | 0 |
| Pradosia ptychandra | 0.08 | 46.7 | 1.79 | 27.1 | 727.94 | 25.16 | 1.57 | 10.66 | 0 | 0 | 28.35 | 17.79 | 10.19 | 0 | 33.01 |
| Aniba rosaeodora | 0.05 | 48.2 | 1.44 | 33.5 | 1019.02 | 30.46 | 0.97 | 0 | 0 | 0 | 50 | 30 | 20 | 0 | 0 |
| Vochysia sabatieri | 0.04 | 47.31 | 1.22 | 39.03 | 1357.56 | 34.89 | 0.23 | 0 | 2.20 | 0 | 30.08 | 41.23 | 14.48 | 2.68 | 9.33 |
| Chrysophyllum sanguinolentum | 0.06 | 48.02 | 2.42 | 19.85 | 816.72 | 41.14 | 1.47 | 0 | 0 | 0 | 18.18 | 59.02 | 13.77 | 0 | 9.03 |
| Sterculia speciosa | 0.05 | 48.63 | 1.35 | 36.10 | 1020.57 | 28.27 | 0.86 | 1.07 | 12.07 | 7.77 | 59.09 | 10 | 10 | 0 | 0 |
| Carapa surinamensis | 0.06 | 48.49 | 1.41 | 34.61 | 816.65 | 23.59 | 0.50 | 30.42 | 0 | 13.52 | 0 | 21.87 | 22.02 | 0 | 12.17 |
| Chimarrhis turbinata | 0.05 | 47.42 | 1.99 | 23.87 | 941.32 | 39.24 | 0.23 | 0 | 15.06 | 22.14 | 15.74 | 17.90 | 4.48 | 24.69 | 0 |
| Drypetes variabilis | 0.05 | 45.13 | 1.45 | 31.15 | 847.58 | 27.14 | 1.03 | 22.84 | 15.94 | 3.22 | 37.99 | 16.00 | 4.00 | 0 | 0 |
| Micropholis venulosa | 0.04 | 47.48 | 1.90 | 25.04 | 1149.58 | 45.91 | 0.65 | 0 | 0 | 0 | 27.64 | 23.10 | 28.69 | 0 | 20.57 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gargallo-Garriga, A.; Sardans, J.; Llusià, J.; Peguero, G.; Asensio, D.; Ogaya, R.; Urbina, I.; Langenhove, L.V.; Verryckt, L.T.; Courtois, E.A.; et al. 31P-NMR Metabolomics Revealed Species-Specific Use of Phosphorous in Trees of a French Guiana Rainforest. Molecules 2020, 25, 3960. https://doi.org/10.3390/molecules25173960
Gargallo-Garriga A, Sardans J, Llusià J, Peguero G, Asensio D, Ogaya R, Urbina I, Langenhove LV, Verryckt LT, Courtois EA, et al. 31P-NMR Metabolomics Revealed Species-Specific Use of Phosphorous in Trees of a French Guiana Rainforest. Molecules. 2020; 25(17):3960. https://doi.org/10.3390/molecules25173960
Chicago/Turabian StyleGargallo-Garriga, Albert, Jordi Sardans, Joan Llusià, Guille Peguero, Dolores Asensio, Romà Ogaya, Ifigenia Urbina, Leandro Van Langenhove, Lore T. Verryckt, Elodie A. Courtois, and et al. 2020. "31P-NMR Metabolomics Revealed Species-Specific Use of Phosphorous in Trees of a French Guiana Rainforest" Molecules 25, no. 17: 3960. https://doi.org/10.3390/molecules25173960