Are Temperate Alpine Plants With Distinct Phenology More Vulnerable to Extraordinary Climate Events Than Their Continuously Flowering Relatives in Tropical Mountains?
 Zdenka Křenová1,2*
Zdenka Křenová1,2*  Pavel Kindlmann1,2
Pavel Kindlmann1,2  J. Stephen Shelly3 Petr Sklenář4 Susanne Sivila1
J. Stephen Shelly3 Petr Sklenář4 Susanne Sivila1  Karolína Bílá1
Karolína Bílá1  Ricardo Jaramillo5
Ricardo Jaramillo5- 1Department of Biodiversity Research, Global Change Research Institute, Brno, Czechia
- 2Faculty of Science, Institute for Environmental Studies, Charles University, Prague, Czechia
- 3US Forest Service, Missoula, MT, United States
- 4Department of Botany, Faculty of Science, Charles University, Prague, Czechia
- 5Escuela de Ciencias Biológicas, Pontificia Universidad Católica del Ecuador, Quito, Ecuador
Alpine plants are perceived as some of the most vulnerable to extinction due to the global climate change. We expected that their life history strategies depend, among others, on the latitude they live in: those growing in temperate regions are likely to have a distinct phenology with short seasonal peaks, while tropical alpine plants can potentially exploit favorable year-round growing conditions and different individuals within a population may flower at different times of the year. In species, whose flowering is synchronized into short seasonal peaks, extraordinary climate events, which may become stronger and more frequent with climate change, can potentially destroy reproductive organs of all synchronized individuals. This may result in reducing fitness or even extinction of such species. We studied field populations of five groups of closely related Andean alpine plant species to test our expectations on their latitude-dependent synchronization of flowering. Our results confirmed these expectations: (i) Tropical alpine species were least synchronized and flowering peaks of different individuals in their populations were distributed across many months. Thus, in tropical alpine species, if an extraordinary event happens, only some individuals are affected and other members of the population successfully reproduce in other parts of the long season. (ii) Higher synchronicity in flowering of temperate and subtropical alpine plants resulted even in some of these species using only a part of the short growing season to reproduce, which increases their vulnerability to extraordinary climatic events. However, we did not find any unique pattern valid for all species, groups and regions. The diversity in flowering phenology (i.e., different levels of seasonality and synchronicity) that we found increases the likelihood of plants successfully coping with climate change.
Introduction
A major concern in contemporary conservation biology is the drastic rate of species extinction causing a decline of biodiversity (IPBES, 2019). One of the main factors causing this rapid extinction is global climate change. With increases in temperature, organisms may be forced to move toward the poles and toward higher elevations (Lenoir and Svenning, 2013). Plants are in an especially difficult position with respect to this because they cannot move actively, which can result in their extinctions (Scheffers et al., 2016). This especially holds for high-mountain plants (Dullinger et al., 2012). Microclimatic heterogeneity present in mountain ecosystems may partly buffer against poleward and upslope migration of alpine plant species. Therefore, at least the species depending on the very cold micro-habitats are in serious troubles (Scherrer and Körner, 2011). Of course, we are limited in our ability to predict climate-change-induced extinctions of many plants caused by climate change, because of our inadequate understanding of how extinctions are affected by non-climatic factors, such as plant phenology (Brown and Vellend, 2014).
Temperature change due to global warming has affected high elevations more than the lowlands (Verrall and Pickering, 2020). One of the obvious signs is the ongoing deglaciation of the tropical high Andes (e.g., Ceballos et al., 2006). This tropical alpine area, with exceptional biodiversity and endemism, is highly sensitive to climate change (Herzog et al., 2011). Tropical alpine species, which are mostly isolated on the highest peaks with specific abiotic conditions and therefore have very limited opportunities to avoid climate change by migrating to higher elevations or latitudes (Jake et al., 2018), are considered more likely to be affected by climate change than temperate alpine species. This is because elevational migration of tropical alpine plants is often impossible due to lack of soil at higher elevations (Quinn, 2008) and latitudinal shift of species range is impossible due to lack of connectivity between the highest peaks (Buytaert et al., 2011). These limitations can have fatal consequences for many tropical alpine species.
Although many alpine plants are perennials, successful generative reproduction is important for survival of their populations (Eriksson, 1997). This is especially important during climate change, as generative reproduction (1) enables plants to escape worsening conditions by migration to new suitable sites and (2) enables population genetic variability and potentially adaptive evolutionary changes that may mitigate the negative effects of climate change (Körner, 2021; Till-Bottraud et al., 2021).
During their evolution, alpine plant species growing in different latitudinal regions have adopted different strategies to optimize their growth and reproductive success (Parmesan, 2006). Alpine plant communities in temperate regions have a distinct phenology with seasonal peaks in flower abundance, and the length of flowering of individual plants varies mostly between 2 and 8 weeks (Arroyo et al., 1981; Körner, 2021). In alpine areas in the tropics, the year-round growing conditions enable plants to use most of the available resources for continuous growth (Diemer, 1996, 1998). Therefore, flowering there is more evenly spread throughout the year, although the patterns may range from almost continuous production of flowers in some species to distinct temporal peaks in others (e.g., Kudo and Hirao, 2006; Fagua and Gonzalez, 2007; Inouye, 2008; Kudo, 2014).
Year-round flower production can offer an additional evolutionary advantage during the changing climate, when – among others – extraordinary climate fluctuations and events (e.g., late frost, changes in patterns and form of precipitation and cloud cover, extremely high precipitation, drought etc.) occur more often (IPCC, 2014; Kohler et al., 2014). They affect mountain vegetation (Foster, 2001; Ruiz et al., 2008; Vuille et al., 2008; Henry et al., 2014) and might reduce success in flowering and seed production (Alatalo et al., 2016). These unusual events typically occur during short discrete instants, rather than continuously throughout the whole season (Jentsch and Beierkuhnlein, 2008). In plants with year-round production of flowers, such events will likely affect only part of the population that is currently in flower, unlike plants with a short reproduction period and synchronized phenology. Thus, one would expect that plant species with continuous production of flowers in their populations have a higher probability of successfully reproducing while populations with synchronized growth and flowering can experience significantly decreased seed production if an extraordinary climatic event affects all individuals simultaneously in a sensitive phenological period. Critical analysis of such expectations is essential for understanding why some alpine species are more likely to go extinct with climate change than others are.
To test the expectation above, it is necessary to compare phenologies of related alpine species growing in different latitudes. However, little is known about the flowering phenology of tropical alpine plants or whether phylogenetically related species growing in alpine regions of different latitudes have adopted similar reproductive strategies, i.e., whether they can similarly react to extraordinary events and thus mitigate climate change consequences (Herzog et al., 2011). The complexity of mountains is tightly associated with high biodiversity, but the processes underlying this association are poorly known (Perrigo et al., 2020). More detailed knowledge of alpine plant phenology in different latitudinal regions can improve our understanding of vulnerability of alpine plants to climate change. We suppose that the less constrained phenologies and unsynchronized flowering in tropical alpine plants may improve their survival rate in the face of climate change. Thus, we test here the hypothesis that the flowering phenology of temperate alpine species, which is more synchronized than that of their relatives growing in tropical alpine regions, causes the former to be more stressed by extraordinary climatic events. We test the same for vegetative growth. To do this, we studied groups of closely related species (with known phylogenies to infer species relationships) in the tropics, subtropics, and temperate regions that have the same growth form and occur in equivalent habitats. We tested, whether the vegetative growth traits (VGTs henceforth) and the flowering phenology traits (FPTs) recorded in the populations of closely related species growing in tropical, subtropical and temperate alpine regions are synchronized during the season or not, i.e., whether an extraordinary event can significantly reduce the total number of seeds production of the species.
Materials and Methods
Study Sites
The continuous cordilleras in North and South America provide unique settings for broad-scale comparative ecological studies. We conducted parallel research in three alpine regions, i.e., the Andes of Ecuador and Bolivia, representing tropical and subtropical regions, respectively, and the Rocky Mts. in the United States, representing the north temperate region (Appendix 1).
The study in Ecuador was carried out on the western side of the Antisana volcano, which was thoroughly surveyed (Sklenář et al., 2016). Vegetation is composed of tussocks of Calamagrostis intermedia along with sclerophyllous shrubs (Chuquiraga jussieui), rosulate herbs (Valeriana, Werneria), and prostrate subshrubs (Baccharis caespitosa). This vegetation gives way to patchy upper superpáramo vegetation of small herbs and grasses at higher elevation. Mean air temperature is 3.7°C with a mean annual sum of 1,098.4 mm of precipitation at 4,280 m (Sklenář et al., 2015). Valeriana plantaginea was studied in the Páramo de Guamaní, located ca 18 km to the north of Antisana. Vegetation is composed of sclerophyllous shrubs (Loricaria and Diplostephium) that share dominance with cushion plants (Azorella, Plantago, and Werneria), and locally also with bamboo and tussock grasses (Neurolepis, Calamagrostis, and Festuca) at lower elevation. At higher elevations, prostrate subshrubs (Disterigma and Pernettya) along with herbs and small grasses (e.g., Lachemilla, Oritrophium, and Agrostis) are common.
In Bolivia, the study was conducted in the southwest edge of the Cotapata National Park and Integrated Management Natural Area, La Paz department. Study sites were located in the Yungas páramo, a typical local habitat with strips and patches of almost constantly humid grasslands and shrublands. The most common genera of the Yungas páramo are Stipa, Festuca, Brachyotum, Satureja, Mutisia, Chuquiraga, Baccharis, Calceolaria, and Gnaphalium (Bach et al., 2003). Gentiana sedifolia is growing in fens and spring areas with high cover of mosses. Other study species prefer mountain grasslands and often grow on steep slopes. Mean annual temperature is about 6°C and mean annual sum of precipitation is more than 3,000 mm at 3,100 m (Körner, 2021). The rainy season is December–March, while June and July are the driest months.
The Niwot Ridge Long Term Ecological Research site in the Rocky Mountains, Colorado, United States (Bowman and Seasted, 2001) was the study site for six species (Appendix 1). Mean annual temperature is −4.1°C and mean annual precipitation is 993 mm at 3,750 m. Alpine tundra is dominated by Geum rossii, Deschampsia caespitosa, and Kobresia myosuroides. Species were studied in the vicinity of the Saddle climatic station. Castilleja, Valeriana, and Cerastium inhabit dry meadow stands, while Caltha and both species of Gentiana inhabit wet meadow stands. There can be annual fluctuations, but the vegetation season usually runs from June 10 to the end of September when the new snow cover begins to accumulate (Spasojevic et al., 2013).
Populations of three target species were studied in Line Creek Plateau Research Natural Area, located near the Montana/Wyoming border, United States (Appendix 1). Line Creek Plateau is the easternmost alpine area in the Beartooth Mountains, where alpine plant phenology has been already studied (Křenová et al., 2019). Part of the study site occurs in the community types with Geum rossii (Castilleja pulchella), while the other part (with Caltha leptosepala and Gentiana algida) occurs in the Salix glauca/Deschampsia cespitosa c.t. The average temperature on Line Creek Plateau is 9–11°C, with an average low temperature of −2°C and an average high temperature of 21–23°C. The plateau receives an average of 1,016 mm of precipitation annually, predominantly in the form of snow (USDA Forest Service, 2001). The vegetation season is about 10 days shorter than in Niwot, usually beginning around June 15 and ending in the middle of September (Lesica, 1993).
Study Species
We studied five species groups (with known phylogenies to infer species relationships) that occur in all three regions (Appendix 1). These plant species groups meet the following criteria: (i) they have the same growth form and (ii) occur in equivalent habitats in the compared regions. The following perennial species were studied:
1. North American Caltha leptosepala (Lesica, 2012), a white flowering herb with a developmental preformation (Aydelotte and Diggle, 1997) and its sister species, C. sagittata, the Andean-Patagonian species with solitary yellowish actinomorphic flowers (Schuettpelz and Hoot, 2004).
2. Cerastium arvense, a highly polymorphic perennial herb growing up to 30 cm, South American C. arvensiforme, species derived from a North American clade (Scheen et al., 2004).
3. South American Castilleja nubigena (Jørgensen and León-Yánez, 1999), an erect subshrub with diffuse stems, along with North American C. puberula (Ackerfield, 2015) and C. pulchella (Lesica, 2012).
4. Gentianella foliosa, an ascending polycarpic herb that is an endemic species from Ecuador (Jørgensen and León-Yánez, 1999), Gentiana sedifolia and Gentinella punicea both growing in Bolivia (Jørgensen et al., 2014), and Gentiana calycosa and Gentiana algida growing in the temperate zone (Lesica, 2012).
5. Valeriana edulis, a dioecious perennial plant native to western and central North America (Ackerfield, 2015), V. plantaginea, an erect plant growing in humid páramo (Jørgensen and León-Yánez, 1999) and Valeriana micropterina in Bolivia (Jørgensen et al., 2014).
Plant Traits and Synchronization of Population Members
Depending on the population size of target species, we randomly selected 15 – 40 individuals and recorded the plant traits summarized in Table 1 to describe their habit and phenology status. Flowering phenology traits (FPTs – see Table 1) were recorded for each plant if flowers were available and all marked plants were characterized by at least three vegetative growth traits (VGTs – see Table 1).

Table 1. Plant traits recorded in populations of studied species.
The data were collected in 2019. In tropical and subtropical regions, we started with monitoring in February and recorded the traits monthly during the whole year. In the temperate zone, we started monitoring immediately with melting of snow and repeatedly recorded the traits monthly in Montana and each 2 weeks in Colorado throughout the whole season, defined as a time between the last and the first continuous snow cover in our study sites, i.e., June 10 – September 25 in Colorado and June 15 – September 20 in Montana.
Statistical Analyses
The values of traits were repeatedly recorded during the data collection on the same individuals. All FPT and VGT data were log (x+1) transformed prior the analyses to ensure normality. Therefore, to test for seasonality (the combined effect of time on the values of measured traits), we have performed the GLM Repeated Measures analysis of variance separately for all recorded FPT and VGT traits. This was done for each region and species group, if data were available. FPTs (i.e., buds, open flowers, old flowers and ripen fruits) describe whether these reproductive phases overlapped or are consecutive in the population. VGTs were studied to see, whether vegetative growth is continuous throughout the season or occurs only in its part and whether beginning of flowering is delayed after beginning of vegetative growth. We test here the zero hypotheses that the VGTs and FPTs are constant functions of time. For each species and trait, significance of seasonality is then expressed by the value of p and shown together with mean values (points) and SDs (bars) for recorded traits (Figures 1–5). Significant p-values are marked by * and – biologically – indicate seasonality of the measured traits. We feel that publicizing this data with only this meagre analysis is still useful – at least because it provides knowledge about flowering phenology of these species, and some additional evidence for our claims.

Figure 1. Caltha group. (A) Vegetative growth traits and (B) flowering phenology traits of Caltha species monitored in three latitudinal regions: tropical – Ecuador, subtropical – Bolivia, and temperate – Colorado, Montana, United States. Studied species are listed for each alpine region. Mean values (points) and SDs (bars) are displayed. Significant p-values are marked by ∗ and – biologically – indicate seasonality of the measured traits. X-axes indicate time in local units of time, specific for each region, which standardized the data, so that the season duration was always 365 local units.
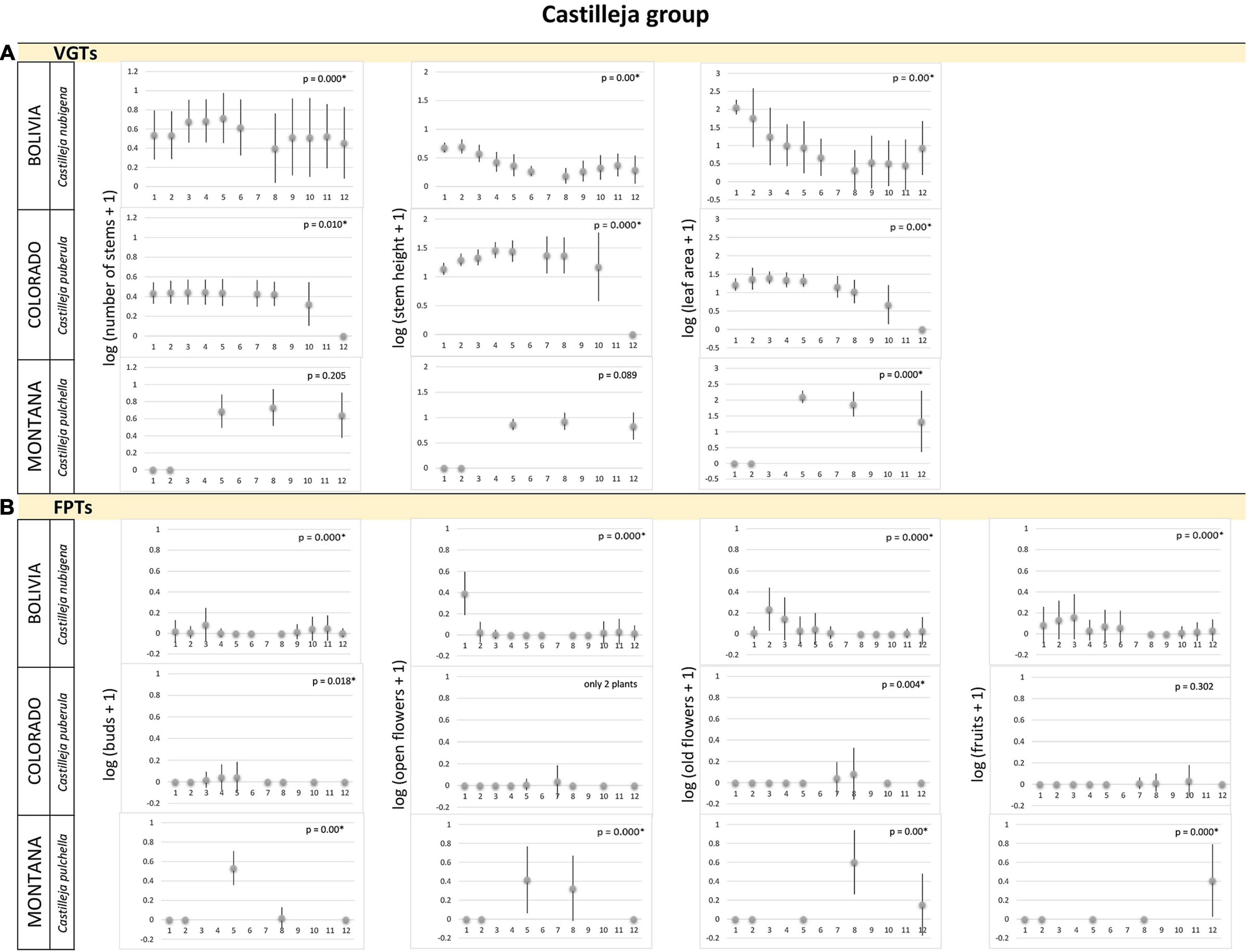
Figure 2. Castilleja group. (A) Vegetative growth traits and (B) flowering phenology traits of Castilleja species monitored in latitudinal regions: subtropical – Bolivia, temperate – Colorado, Montana, United States. Studied species are listed for each alpine region. Mean values (points) and SDs (bars) are displayed. Significant p-values are marked by ∗ and – biologically – indicate seasonality of the measured traits. X-axes indicate time in local units of time, specific for each region, which standardized the data, so that the season duration was always 365 local units.
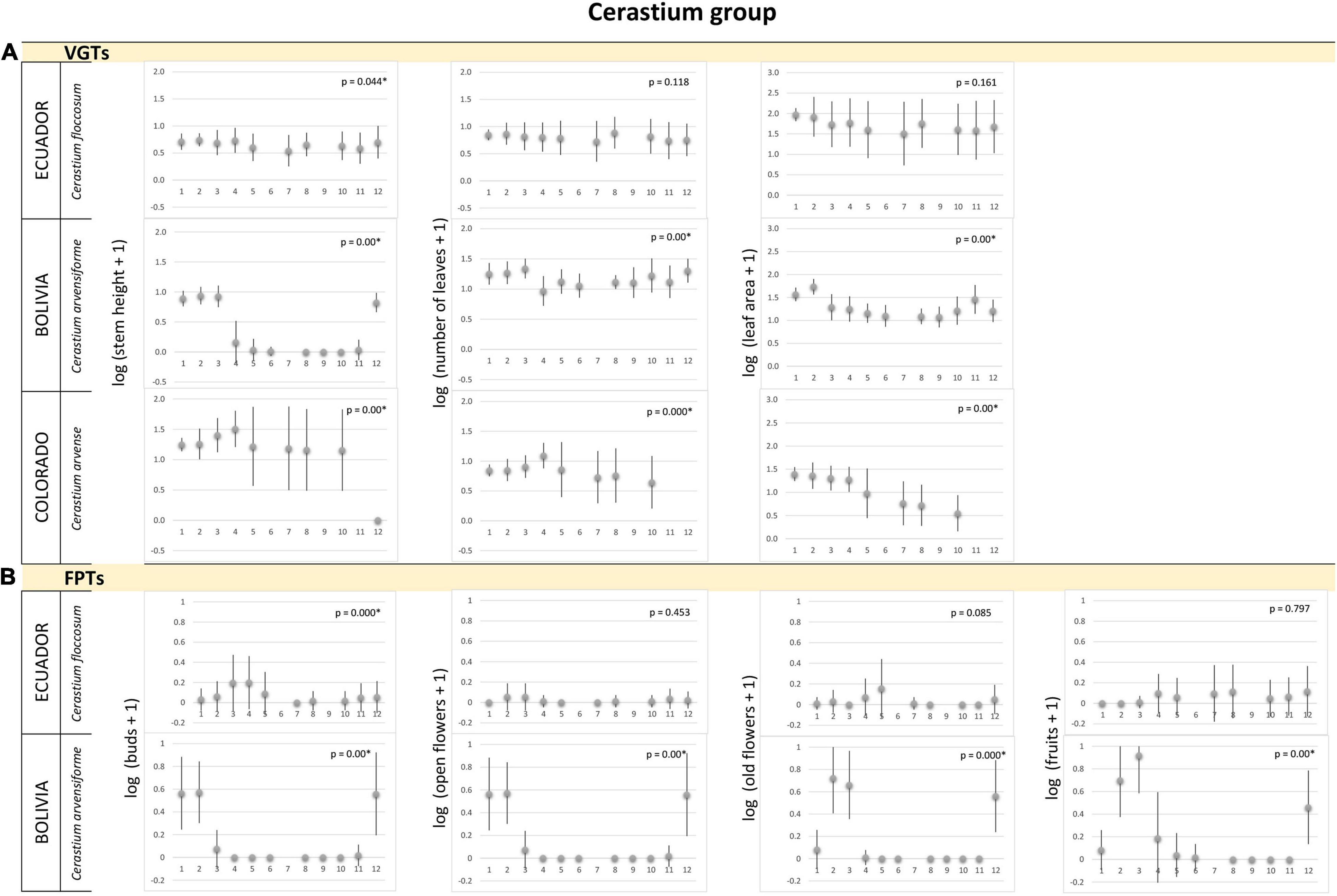
Figure 3. Cerastium group. (A) Vegetative growth traits and (B) flowering phenology traits of Cerastium species monitored in three latitudinal regions: tropical – Ecuador, subtropical – Bolivia, and temperate – Colorado, United States. Studied species are listed for each alpine region. Mean values (points) and SDs (bars) are displayed. Significant p-values are marked by ∗ and – biologically – indicate seasonality of the measured traits. X-axes indicate time in local units of time, specific for each region, which standardized the data, so that the season duration was always 365 local units.
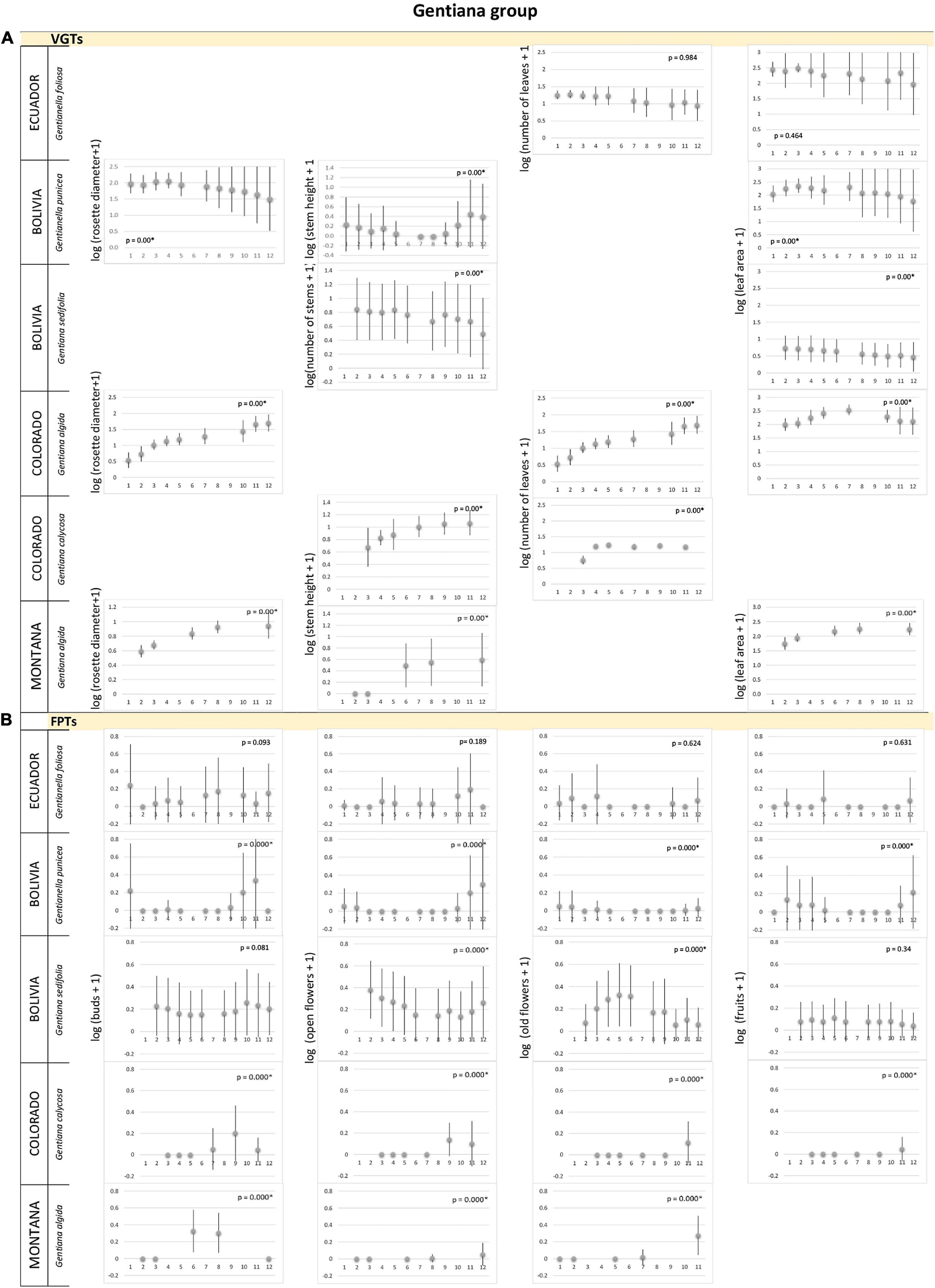
Figure 4. Gentiana group. (A) Vegetative growth traits and (B) flowering phenology traits of Gentiana species monitored in three latitudinal regions: tropical – Ecuador, subtropical – Bolivia, and temperate – Colorado, Montana, United States. Studied species are listed for each alpine region. Mean values (points) and SDs (bars) are displayed. Significant p-values are marked by ∗ and – biologically – indicate seasonality of the measured traits. X-axes indicate time in local units of time, specific for each region, which standardized the data, so that the season duration was always 365 local units.
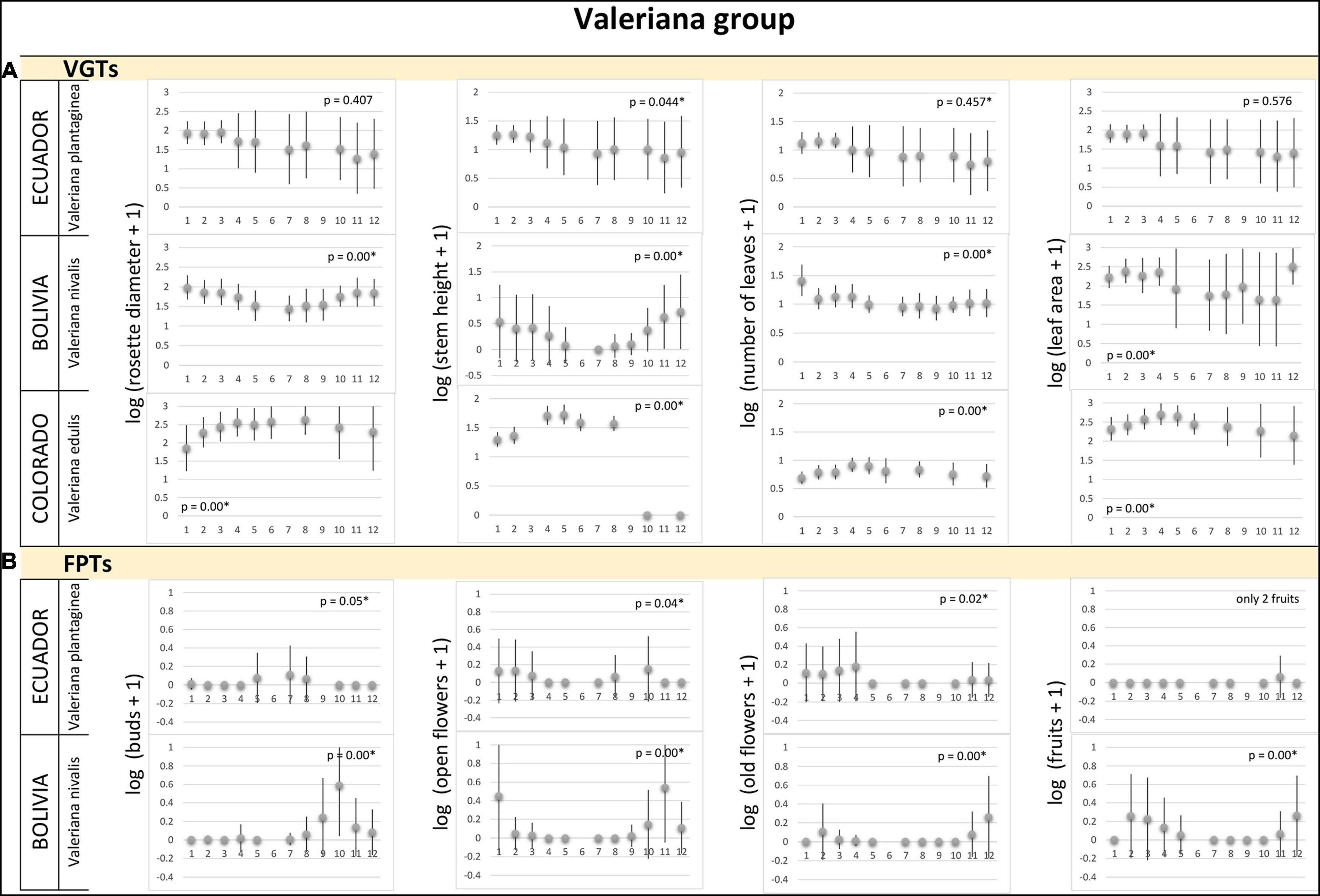
Figure 5. Valeriana group. (A) Vegetative growth traits and (B) flowering phenology traits of Valeriana species monitored in three latitudinal regions: tropical – Ecuador, subtropical – Bolivia, and temperate – Colorado, United States. Studied species are listed for each alpine region. Mean values (points) and SDs (bars) are displayed. Significant p-values are marked by ∗ and – biologically – indicate seasonality of the measured traits. X-axes indicate time in local units of time, specific for each region, which standardized the data, so that the season duration was always 365 local units.
When testing for synchronicity in the population, we first had to standardize the data with respect to interregional differences in season duration: in the temperate alpine region, the season is shorter, compared with the tropics and subtropics. Therefore, we introduced new local units of time, specific for each region, which standardized the data, so that the season duration was always 365 local units, independently of the region, and used these units as measures of time in all subsequent analyses, unless otherwise indicated. This was done as follows:
In the tropics and subtropics, we set the unit duration equal to 1 day, so the season duration was 365 units (=365 days).
In Colorado, 1 day was set equal to 3.38 local units, as the season lasts here 108 days, which is then 365 local units (as 108 × 3.38 = 365).
Similarly, in Montana, where the season is only 98 days, 1 day was set equal to 3.72 local units, as 98 × 3.72 = 365.
We calculated synchronicity with respect to trait r within a population in several steps:
We defined as Xr,i,t the value of trait r on plant i at time t (expressed in the local time units).
We defined the individual peak for trait r and plant i as IPr,i = maxt(Xr,i,t) and the population peak for trait r as PPr = maxt[averagei(Xr,i,t)].
For trait r and plant, i, we defined distance, Dr,i, between PPr and IPr,i as: Dr,i = | PPr – IPr,i|.
Synchronicity, Sr, of a population in terms of trait r was then defined as Sr = averagei(Dr,i).
Biologically, the larger the Sr, the lower the degree of synchronization of the plants in the population with respect to trait r. On the dataset consisting of all Dr calculated for all plants from each population from each region and species group, we then performed factorial ANOVA with factors “region” and “species group”; Duncan’s test was then applied for post-hoc comparison, where appropriate. We performed this only on traits, which were available for all species groups (thus only on leaf area and on numbers of open flowers) and only on species groups, for which data were available from all three latitudinal regions (thus, Castilleja species were excluded).
Statistical analyses were performed in STATISTICA 12 (Anonymous, 2012).
Results
Seasonality
We found no significant differences, i.e., no seasonality, in most of VGTs collected monthly for Caltha, Cerastium, Gentiana and Valeriana plants (and no data for Castilleja) in Ecuador (Figures 1A, 3A, 4A, 5A). Thus, vegetative growth in the tropical alpine region is non-seasonal. With respect to FPTs, the situation was different. Caltha sagittata, Cerastium floccosum and Gentianella foliosa produced flowers and fruits almost continuously, no significant differences were found (Figures 1B, 3B, 4B). However, Cerastium floccosum produced majority of buds in March–May, while ripened fruits were recorded mainly during the second half of the year (Figure 3B). Valeriana plantaginea showed significant differences in FPTs during the year (Figure 5B) and some seasonal trends can be recognized, too. To summarize, flowers and fruits occurred almost at any time of the year in each of the tropical alpine populations despite certain fluctuations.
The species studied in the subtropical alpine region showed a seasonal pattern in their VGTs (Figures 1A–5A). Their maximum VGT values were recorded in November–March, FPTs of most of studied species studied in the subtropical alpine region had a seasonal pattern, i.e., their monthly recorded values significantly differed (Figures 2B–5B). No seasonality was recorded in population of C. sagitatta, in which only less than 10% of marked individuals flowered. Gentiana sedifolia did not show seasonality in production of buds and had different peaks of flowering and fruit production compared to Gentianella punicea growing in the same region (Figure 4B). Gentiana sedifolia created buds primarily in August and September, most of the flowers were recorded in September–November, and fruits were observed only in November and December (Figure 4B). In some species, flowers and fruits were missing in the first half of the year. Flowering seasons of Castilleja nubigena (Figure 2B) and Cerastium arvensiforme (Figure 3B) lasted only half of the year, but they started later than in G. sedifolia. The buds and flowers appeared in December–March, and most of the fruits were recorded in January–May (June). However, there was an obvious pattern of seasonal proportionality of different phenological phases. Numbers of buds and opened flowers in the population increased in September–December and decreased in February–June, when numbers of old flowers increased. Slightly different flowering phenology was recorded for Valeriana micropterina (Figure 5B), which reproduced during the whole year, but each phenological phase lasted only several months. This species had most of the buds in August–December, flowers in October–February, and fruits in December–April (May). To summarize, most of the studied species in the subtropical alpine region showed at least partial seasonal pattern in their VGTs, as well as in production of flowers and fruits. Some species showed a pattern of seasonal proportionality of different phenological phases.
The vegetation season in the temperate alpine zone is much shorter than that in tropical and subtropical alpine areas. However, even in this significantly shorter period, we observed that some species used only a part of the season. Both flowering and vegetative growth were limited for only several weeks. All species showed significant differences in measured traits, both VGTs and PFTs, except of several cases when only few flowering plants were recorded in the studied populations. Caltha leptosepala (Figure 1) was the earliest species. The buds and the first opened flowers were recorded on plants growing out of the just-melting snow. Old flowers were present in July and the fruits were ripe before the middle of August. The values of VGTs culminated in the middle of the season and then they declined. In September, many of the marked plants were senescent and their leaves and flowering stems were withered. In Colorado, Castilleja puberula (Figure 2A) showed higher values of VGTs during the first two thirds of the season and declined from the second week of September. At that time, the ripe fruits were recorded. Its sister species, C. pulchella, monitored in Montana, showed a different strategy: no individuals were recorded at the study site before July 10, values of VGTs were very similar from July to August, and slightly declined in the middle of September (Figure 2A). Flowers appeared at the end of July and in August (Figure 2B). The first ripe fruits were recorded in the middle of September; however, many individuals were already senescent with broken stems at that time and their fruits had not reached maturity. Cerastium arvense (Figure 3A), monitored only in Colorado, showed peak VGT values in the middle of July and the plants were already senescent during the second week of September. Gentiana species, G. algida and especially G. calycosa, are late-summer species. VGTs of G. calycosa were first recorded at the beginning of July, i.e., 3 weeks after the season began (Figure 4A). Buds appeared during the second week of August, and most opened flowers were recorded at the end of August and beginning of September. The first ripe fruits were found in the middle of September (Figure 4B). VGTs of G. algida, monitored both in Colorado and Montana (Figure 4), showed a slight increase in values throughout the whole season. In Montana, the first buds appeared at the beginning of August and flowering continued until the last week of September, when new snow covered the study site again. No ripe fruits were recorded (Figure 4B). Unfortunately, we do not have data for FPTs from Colorado for G. algida. Valeriana edulis was studied only in Colorado. Values of VGTs slightly increased from the beginning of season to the end of August and then declined (Figure 5A). To summarize, the alpine plants growing in the temperate region are the most synchronized ones. Some of them used only a part of the short growing season to flower and produce fruits.
Synchronicity
We found significant differences among regions in synchronicity of the number of open flowers and maximum leaf area (Table 2). The synchronicity significantly increased (and Sr got smaller – see end of section “Statistical Analyses”) from the tropical to the temperate regions (Figure 6). Thus, in all tested species groups, the temperate alpine species were the most synchronized, while the tropical alpine species were the least synchronized.
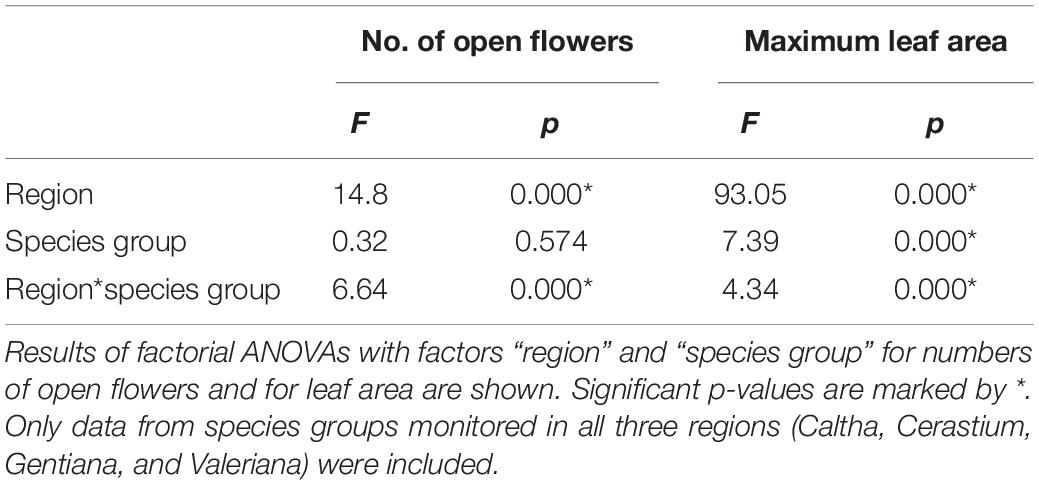
Table 2. Synchronicity of flowering (i.e., the number of open flowers) and maximum leaf area.
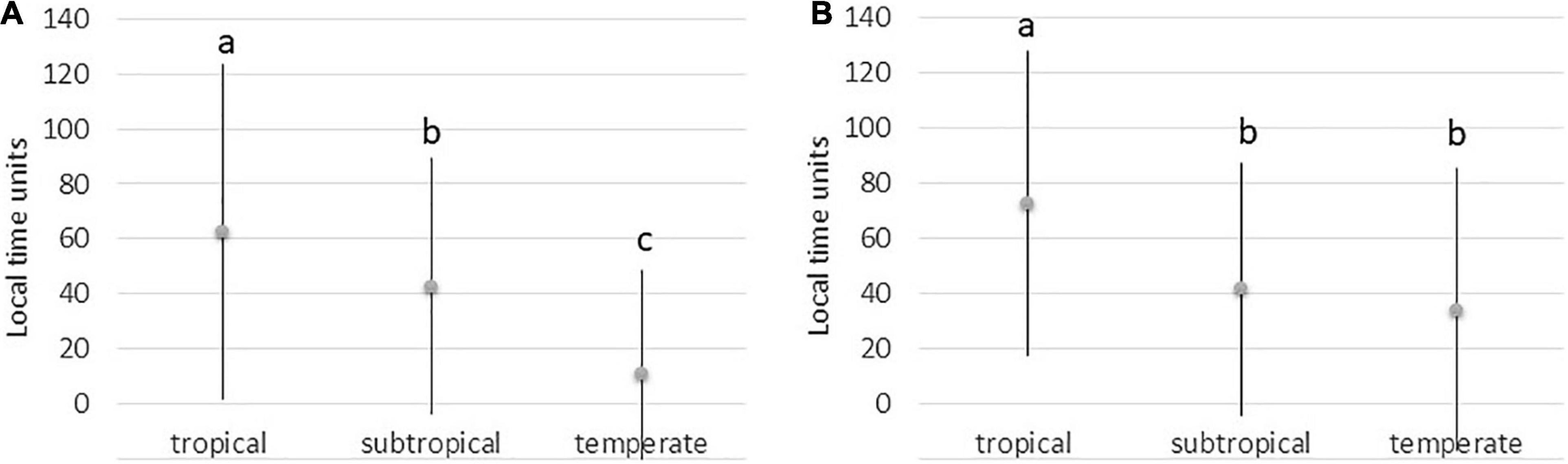
Figure 6. Synchronicity of the numbers of open flowers (A) and maximum leaf area (B) in tropical, subtropical, and temperate alpine regions. Means and SDs of the differences between individual and population peaks of flowering and maximum leaf area expressed in local time units are shown. Local time units were used to standardize interregional differences in season duration. Letters above the bars indicate results of post-hoc comparisons; i.e., different letters mark statistically different values.
The average distance between the individual and population peaks of flowering was 65 local time units in the tropical alpine region vs. 10.65 local time units in the temperate alpine region (Figure 6A). For leaf area, it was 73 local time units in the tropical alpine region vs. 34 local time units in the temperate alpine region (Figure 6B). Local time units were used to standardize interregional differences in season duration.
Discussion
Climate change scenarios predict an increased frequency of extreme climatic events. In Arctic and alpine regions, one of the most profound of these are extreme and sudden winter warming events in which temperatures increase rapidly to above freezing, often causing snow melt across whole landscapes and exposure of ecosystems to warm temperatures. Vegetation and soils no longer insulated below snow are then exposed to rapidly returning extreme cold. These extreme climatic events can negatively affect reproduction success of plants growing in these biotopes. For example, Bokhorst et al. (2008) recorded reduced flowering and fruit production in populations of Empetrum hermaphroditum and Vaccinium vitis-idaea exposed to a single, short, extreme winter warming event. They conclude that, although climate change may lengthen the growing season by earlier spring snowmelt, there is a profound danger for these high-latitude ecosystems if extreme, short-lived warming in winter exposes plants to initial warm temperatures, but then extreme cold for the rest of the winter.
In our study, we have emphasized other types of extreme climatic events (e.g., heavy rain, heel storms, or flooding). Our analyses show that alpine plants growing in the temperate region are the most synchronized ones. This increases their vulnerability to extraordinary climatic events, which could destroy reproductive organs of all synchronized individuals or disrupt plant-pollinator interactions (Memmott et al., 2007). We found that some of the temperate alpine plants therefore used only a part of the short growing season to reproduce. The way, how we calculated synchronicity, proves that delayed or replacement (second) flowering does not help due to the short growing season. Even an earlier start of the growing season, i.e., early snow melting, could probably not solve this. No climate-induced advances in the timing of first flowering, but the date of last flowering emerged were recognized as being a more important determinant of flowering duration and the maximum number of flowers in alpine vegetation (Dorji et al., 2020).
On the contrary to temperate region, we found that alpine plants growing in the tropical region are not very synchronized and the growth and flowering peaks of different members of each population are distributed across many months. If an extraordinary event happens, low synchronicity means in biological terms that only some individuals are affected, while other members of the population can successfully reproduce in other parts of the long season. Even the individuals with damaged flowers can regenerate and repeat their flowering several weeks later, because conditions are suitable year-round. In the subtropical region, synchronicity is not as distinct. Growth and flowering peaks of individuals are less synchronized than in the temperate region; however, some extraordinary events could block successful regeneration, too. Further, replacement flowering could be shifted to months with a lack of precipitation, which can cause the plants to dry before the seeds ripen (Körner, 2021).
Regardless of their evolutionary relationships and similar life forms, species in the same species group growing in alpine regions of different latitudes have adopted various strategies to optimize their reproduction and vegetative growth with respect to the local conditions. The species expressed continuous vegetative growth in the tropical alpine region and seasonal patterns in subtropical and temperate regions. In the subtropical region, values of VGTs often followed patterns of local rainfall (Körner, 2021). E.g., in Bolivia, they declined during the first half of the year and increased from July or August. Gentiana sedifolia was the only subtropical species deviating from this pattern, growing and flowering continuously. This can be explained by the fact that bogs and fens are typical habitats of G. sedifolia and the species’ phenology is not determined by soil humidity, as we could have observed for other species in this region.
The tropical alpine Caltha sagittata was the only species in this study with almost continuous production of buds, flowers and ripe fruits. However, the C. sagittata population monitored in the subtropical region indicated possible seasonality and two temperate populations of C. leptosepala showed very clear seasonality in FPTs. Flowering phenology of all other studied tropical alpine species was also more or less seasonal. However as mentioned above, some species in our study produced buds, flowers and ripe fruits continuously during the whole season. Some species showed a clear pattern between the individual phenological phases, which were mostly separate and arranged one after the other. Other species reproduced only during part of the season, either early or late. Their flowering phases overlapped, i.e., some individuals of the same population were early while others formed new buds when their neighbors were already bearing fruits. We presume that not all of these adaptations may be the most advantageous in a time of climate change when extraordinary events, local climate fluctuations and various disturbances occur more often. Some alpine species may respond to these new challenges better than others may. For example, the Caltha species in both the tropical alpine region (where continuous flowering occurs) and in the subtropical and temperate regions (where the flowering period is short) seem to be more resistant to climatic stresses. The species’ short growth periods together with a developmental preformation, well documented for C. leptosepala (Aydelotte and Diggle, 1997) and presumed for others, ensure successful reproduction even in the years when the snow melts late or the season is unusually short due to early cooling and new snow. On the contrary, the phenologically later species, e.g., some Castilleja or Gentiana species, are not able to produce seeds if the season in a temperate alpine region is too short. In Montana in 2019, neither Castilleja nor Gentiana were able to produce seeds because of the short season, while both species successfully reproduced in the longer 2018 season (Křenová et al., 2019). In the subtropical alpine region, delayed flowering can cause fruit ripening in times of low soil moisture, when plants are withering before delivering the ripe seeds. We observed this with Cerastium in Bolivia.
Our data have partially confirmed our prediction that flowering as well as vegetative growth of tropical alpine plants are less synchronized, and their flowers occur at any time of the year. This means that generative reproduction of tropical alpine plants may be less stressed by extraordinary climatic events, but the situation is more complicated. Different tropical alpine species showed different levels of synchronicity. For example, Caltha sagittata and Gentiana foliosa populations showed even production of buds, flowers and fruits during the whole year, while the Cerastium flucosum population showed some seasonal fluctuations.
Clearly, there is no single pattern or strategy shared among all species groups or regions. Therefore, the diversity of patterns observed suggests that plant populations may be able to persist in the face of unpredicted events brought on by climate change.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
ZK and PK conceived the idea, collected and analyzed the data, developed and designed methods, wrote and edited the original draft, and prepared the final version. JS collected the data and edited the English and original draft. PS collected the data and edited the original draft. SS and RJ collected the data. KB prepared the data for analysis. All authors contributed to the article and approved the submitted version.
Funding
This study was funded by the Czech Science Foundation (project no. 17-12420S) and Ministry of Education, Youth and Sports of CR within the CzeCOS program, grant number LM2018123, partly supported the manuscript preparation.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank M. Mantas, for helping with field data collection in Montana.
References
Ackerfield, J. (2015). Flora of Colorado. Fort Worth, TX: BRIT Press, Botanical Research Institute of Texas.
Alatalo, J. M., Jagerbrand, A. K., and Molau, U. (2016). Impacts of different climate change regimes and extreme climatic events on an alpine meadow community. Sci. Rep. 6:21720. doi: 10.1038/srep21720
Arroyo, M. T. K., Armesto, J. J., and Villagrán, C. (1981). Plant phenological patterns in the high Andean cordillera of Central Chile. J. Ecol. 69, 205–223. doi: 10.2307/2259826
Aydelotte, A. R., and Diggle, P. K. (1997). Analysis of developmental preformation in the alpine herb Caltha leptosepala (Ranunculaceae). Am. J. Bot. 84, 1646–1657. doi: 10.2307/2446462
Bach, K., Schawe, M., Beck, S., Gerold, G., Gradstein, S. R., and Mónica Moraes, R. (2003). Vegetación, suelos y clima en los diferentes pisos altitudinales de bosques montañosos de Yungas (Bolivia). Ecol. Boliv. 38, 3–14.
Bokhorst, S., Bjerke, J. W., Bowles, F. W., Melillo, J., Callaghan, T. V., and Phoenix, G. K. (2008). Impacts of extreme winter warming in the sub-Arctic: growing season responses of dwarf shrub heathland. Glob. Change Biol. 14, 2603–2612. doi: 10.1111/j.1365-2486.2008.01689.x
Bowman, W. D., and Seasted, T. R. (2001). Structure And Function Of An Alpine Ecosystem Niwot Ridge, Colorado. Oxford: Oxford University Press.
Brown, C. D., and Vellend, M. (2014). Non-climatic constraints on upper elevational plant range expansion under climate change. Proc. Roy. Soc. B Biol. Sci. 281:20141779. doi: 10.1098/rspb.2014.1779
Buytaert, W., Cuesta-Camacho, F., and Tobon, C. (2011). Potential impacts of climate change on the environmental services of humid tropical alpine regions. Glob. Ecol. Biogeogr. 20, 19–33.
Ceballos, J. L., Euscategui, C., Ramirez, J., Cañon, M., Huggel, C., Haeberli, W., et al. (2006). Fast shrinkage of tropical glaciers in Colombia. Ann. Glaciol. 43, 194–201. doi: 10.3189/172756406781812429
Diemer, M. (1996). Microclimatic convergence of high-elevation tropical páramo and temperate-zone alpine environments. J. Veg. Sci. 7, 821–830. doi: 10.2307/3236460
Diemer, M. (1998). Leaf lifespan of high-elevation, aseasonal Andean shrub species in relation to leaf trait and leaf habit. Glob. Ecol. Biogeogr. Lett. 7, 457–465. doi: 10.1046/j.1466-822x.1998.00318.x
Dorji, T., Hopping, K. A., Meng, F., Wang, S., Jiang, L., and Klein, J. A. (2020). Impacts of climate change on flowering phenology and production in alpine plants: the importance of end of flowering. Agric. Ecosyst. Environ. 291:106795. doi: 10.1016/j.agee.2019.106795
Dullinger, S., Gattringer, A., Thuiller, W., Moser, D., Zimmermann, N. E., Guisan, A., et al. (2012). Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Change 2, 619–622. doi: 10.1038/nclimate1514
Eriksson, O. (1997). “Clonal life history and the evolution of seed recruitment,” in The Ecology and Evolution of Clonal Plants, eds H. de Kroon and J. M. van Groenendael (Leiden: Backhuys Publishers), 211–226.
Fagua, J., and Gonzalez, V. (2007). Growth rates, reproductive phenology, and pollination ecology of espeletia grandiflora (asteraceae), a giant andean caulescent rosette. Plant Biol. 9, 127–135. doi: 10.1055/s-2006-924544
Foster, P. (2001). The potential negative impacts of global climate change on tropical montane cloud forests. Earth Sci. Rev. 55, 73–106. doi: 10.1016/s0012-8252(01)00056-3
Henry, F. D., Raymond, S. B., and Liang, N. (2014). Climatic changes in mountain regions of the American Cordillera and the tropics: historical changes and future outlook. Arct. Antarct. Alp. Res. 46, 735–743. doi: 10.1657/1938-4246-46.4.735
Herzog, S., Martinez, R., Jørgensen, P., and Tiessen, H. (2011). Climate Change And Biodiversity In The Tropical Andes. Paris: Inter-American Institute for Global Change Research (IAI) and Scientific Committee on Problems of the Environment (SCOPE).
Inouye, D. W. (2008). Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology 89, 353–362. doi: 10.1890/06-2128.1
IPBES (2019). Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Bonn: IPBES Secretariat.
IPCC (2014). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva: IPCC Secretariat.
Jake, A., Chalmandrier, L., Lenoir, J., Burgess, T., Essl, F., Haider, S., et al. (2018). Lags in the response of mountain plant communities to climate change. Glob. Change Biol. 24, 563–579. doi: 10.1111/gcb.13976
Jentsch, A., and Beierkuhnlein, C. (2008). Research frontiers in climate change: effects of extreme meteorological events on ecosystems. C. R. Geosci. 340, 621–628. doi: 10.1016/j.crte.2008.07.002
Jørgensen, P. M., and León-Yánez, S. (1999). Catalogue Of The Vascular Plants Of Ecuador: Monographs in Systematic Botany from the Missouri Botanical Garden. St. Louis, MO: Missouri Botanical Garden Press.
Jørgensen, P. M., Beck, S. G., and Nee, M. (2014). Catálogo De Las Plantas Vasculares De Bolivia: Monographs in Systematic Botany from the Missouri Botanical Garden. St. Louis, MO: Missouri Botanical Garden Press.
Kohler, T., Wehrli, A., and Jurek, M. (eds) (2014). Mountains And Climate Change: A Global Concern. Bern: Centre for Development and Environment (CDE).
Körner, C. (2021). Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlin: Springer.
Křenová, Z., Shelly, J. S., and Chocholoušková, Z. (2019). Methods for monitoring alpine plant phenology: a pilot study in the Line Creek Plateau Research Natural Area, USA. Eur. J. Environ. Sci. 9, 77–86.
Kudo, G. (2014). Vulnerability of phenological synchrony between plants and pollinators in an alpine ecosystem. Ecol. Res. 29, 571–581. doi: 10.1007/s11284-013-1108-z
Kudo, G., and Hirao, A. (2006). Habitat-specific responses in the flowering phenology and seed set of alpine plants to climate variation: Implications for global-change impacts. Popul. Ecol. 48, 49–58.
Lenoir, J., and Svenning, J. C. (2013). “Latitudinal and elevational range shifts under contemporary climate change,” in Encyclopedia of Biodiversity, 2nd Edn, ed. S. Levin (Waltham, MA: Academic Press), 599–611. doi: 10.1002/ecy.3525
Lesica, P. (1993). Vegetation And Flora Of The Line Creek Plateau Area, Carbon County, Montana. Helena, MT: Montana Natural Heritage Program.
Memmott, J., Craze, P. G., Waser, N. M., and Price, M. V. (2007). Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 10, 710–717. doi: 10.1111/j.1461-0248.2007.01061.x
Parmesan, C. (2006). Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 37, 637–669. doi: 10.1146/annurev.ecolsys.37.091305.110100
Perrigo, A., Hoorn, C., and Antonelli, A. (2020). Why mountains matter for biodiversity. J. Biogeogr. 47, 315–325. doi: 10.1111/gcb.15951
Quinn, J. A. (2008). Arctic and Alpine Biomes Greenwood Guides to Biomes of the World. Westport, CT: Greenwood Press.
Ruiz, D., Moreno, H. A., Gutierrez, M. E., and Zapata, P. A. (2008). Changing climate and endangered high mountain ecosystems in Colombia. Sci. Total Environ. 398, 122–132. doi: 10.1016/j.scitotenv.2008.02.038
Scheen, A. C., Brochmann, C., Brysting, A. K., Elven, R., Morris, A., Soltis, D. E., et al. (2004). Northern hemisphere biogeography of Cerastium (Caryophyllaceae): insights from phylogenetic analyses of noncoding plastid nucleotide sequences. Am. J. Bot. 91, 943–952. doi: 10.3732/ajb.91.6.943
Scheffers, B., De Meester, L., Bridge, T., Hoffmann, A., Pandolfi, J. M., Corlett, R. T., et al. (2016). The broad footprint of climate change from genes to biomes to people. Science 354:7671. doi: 10.1126/science.aaf7671
Scherrer, D., and Körner, C. (2011). Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J. Biogeogr. 38, 406–416.
Schuettpelz, E., and Hoot, S. B. (2004). Phylogeny and biogeography of Caltha (Ranunculaceae) based on chloroplast and nuclear DNA sequences. Am. J. Bot. 91, 247–253. doi: 10.3732/ajb.91.2.247
Sklenář, P., Kučerová, A., Macková, J., and Macek, P. (2015). Temporal variation of microclimate in high-altitude páramo of Antisana. Ecuador. Geogr. Fis. Din. Quarter. 38, 67–78.
Sklenář, P., Kuèerová, A., Macková, J., and Romoleroux, K. (2016). Temperature microclimates of plants in a tropical alpine environment: how much does growth form matter? Arct. Antarct. Alp. Res. 48, 61–78. doi: 10.1657/aaar0014-084
Spasojevic, M. J., Bowman, W. D., Humphries, H. C., Seastedt, T. R., and Siding, K. N. (2013). Changes in alpine vegetation over 21 years: are patterns across a heterogeneous landscape consistent with predictions? Ecosphere 4:117.
Till-Bottraud, I., Aubert, S., and Douzet, R. (2021). Reproductive Strategies Of Alpine Plants, Encyclopedia Of The Environment, [online ISSN 2555-0950]. Available online at: https://www.encyclopedie-environnement.org/en/life/reproductive-strategies-of-alpine-plants/ (accessed December 17, 2021).
USDA Forest Service (2001). Establishment Record for Line Creek Plateau Research Natural Area. Missoula, MT: USDA Forest Service, Northern and Rocky Mountain Regions.
Verrall, B., and Pickering, C. M. (2020). Alpine vegetation in the context of climate change: a global review of past research and future directions. Sci. Total Environ. 748:141344. doi: 10.1016/j.scitotenv.2020.141344
Vuille, M., Francou, B., Wagnon, P., Juen, I., Kaser, G., Mark, B. G., et al. (2008). Climate change and tropical Andean glaciers: past, present and future. Earth Sci. Rev. 89, 79–96. doi: 10.1016/j.earscirev.2008.04.002
Appendix
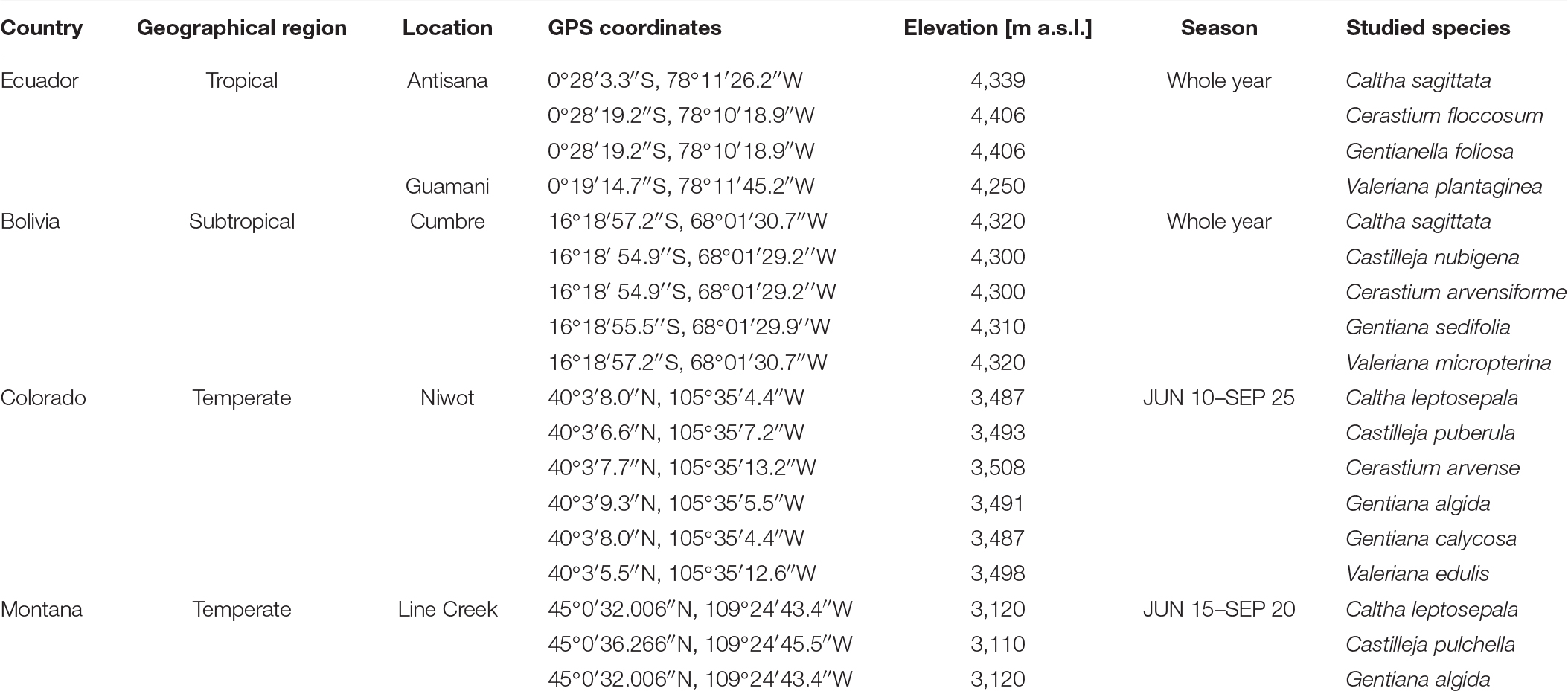
Appendix A1. Study sites, length of season, and studied species.
Keywords: alpine plants, climate change, latitudinal trends, phenology, reproduction success
Citation: Křenová Z, Kindlmann P, Shelly JS, Sklenář P, Sivila S, Bílá K and Jaramillo R (2022) Are Temperate Alpine Plants With Distinct Phenology More Vulnerable to Extraordinary Climate Events Than Their Continuously Flowering Relatives in Tropical Mountains? Front. Ecol. Evol. 9:804102. doi: 10.3389/fevo.2021.804102
Received: 28 October 2021; Accepted: 09 December 2021;
Published: 05 January 2022.
Edited by:
Xiangjin Shen, Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Dongsheng Zhao, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences (CAS), ChinaJianjun Zhao, Northeast Normal University, China
Copyright © 2022 Křenová, Kindlmann, Shelly, Sklenář, Sivila, Bílá and Jaramillo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zdenka Křenová, krenova.z@czechglobe.cz