Biogeographical Importance of the Livingstone Mountains in Southern Tanzania: Comparative Genetic Structure of Small Non-volant Mammals
 Laura N. Cuypers1
Laura N. Cuypers1  Christopher Sabuni2
Christopher Sabuni2  Radim Šumbera3 Tatiana Aghová4 Edita Lišková3
Radim Šumbera3 Tatiana Aghová4 Edita Lišková3  Herwig Leirs1
Herwig Leirs1  Stuart J. E. Baird4 Joëlle Goüy de Bellocq4,5
Stuart J. E. Baird4 Joëlle Goüy de Bellocq4,5  Josef Bryja4,6*
Josef Bryja4,6*
- 1Evolutionary Ecology Group, Department of Biology, University of Antwerp, Antwerp, Belgium
- 2Pest Management Center, Sokoine University of Agriculture, Morogoro, Tanzania
- 3Department of Zoology, Faculty of Science, University of South Bohemia, České Budějovice, Czechia
- 4Institute of Vertebrate Biology of the Czech Academy of Sciences, Brno, Czechia
- 5Department of Zoology and Fisheries, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
- 6Department of Botany and Zoology, Faculty of Science, Masaryk University, Brno, Czechia
The Livingstone Mountains (LM; also known as the Kipengere Range) found in south-western Tanzania at the northern end of Lake Nyasa are an important region for understanding the biogeography of Eastern Africa. The two branches of the East African Rift Valley meet here and the mountains might represent stepping stones for colonization and migration between different parts of the Eastern Afromontane Biodiversity Hotspot (especially the link between the Eastern Arc Mountains, EAM, and the Southern Rift Mountains, SRM), as well as an efficient barrier to gene flow for taxa living in drier savannahs in lower elevations. Here we combine new mitochondrial sequence data from 610 recently sampled rodents and shrews with available georeferenced genetic data (3538 specimens) from southern Tanzania, northern Malawi/Zambia and northern Mozambique and compare the spatial genetic structure among different taxa. There is no universal phylogeographic pattern in taxa preferring humid montane habitats. For some of them, the Makambako Gap acts as a barrier between the SRM and the EAM, but other taxa can bridge this gap. Barriers within the EAM (frequently) and within the SRM (sometimes) appear more important. The Rukwa rift between the SRM and the ARM is an important barrier that perhaps can only be crossed by taxa that are not that strictly tied to humid montane environments. For mammals living in lower-elevation savannah-like habitats, the LM can act as a strict barrier to gene flow, and together with the Ufipa Plateau, Lake Nyasa and the EAM create a very similar phylogeographic pattern with three recognizable genetic groups in most savannah-dwellers. The Livingstone Mountains thus appear to be one of the most important biogeographic crossroads in Eastern Africa.
Introduction
The Eastern Afromontane Biodiversity Hotspot (EABH) is the biodiversity hotspot with the second highest number of endemic higher vertebrate genera, after Madagascar and the Indian Ocean islands (Mittermeier et al., 2011). It is estimated to be home to at least 7,598 plant species (2,356 endemic), 1,325 bird species (157 endemic), 490 mammal species (104 endemic), 347 reptile species (93 endemic) and 323 amphibian species (100 endemic) (Mittermeier et al., 2004; Birdlife International, 2012). These numbers are underestimates as new surveys continue to discover new species (Rovero et al., 2014) and genotyping efforts continue to uncover cryptic diversity (Demos et al., 2014; Greenbaum et al., 2015; Pozzi et al., 2020). These mountains are often thought of as islands of humid forest surrounded by drier lowland savannahs and woodlands (White, 1983). However, the montane forest islands were not always as isolated from each other. It is generally accepted that in the early Miocene forests likely spanned from the west to the east coast of Africa (Coetzee, 1993). Later on, the climate became drier in eastern Africa due to substantial tectonic uplift. The forests started to fragment and the EABH forests were separated from the Guineo-Congolian forests (Sepulchre et al., 2006). Subsequent climatic oscillations in the Plio-Pleistocene were crucial for forming contemporary species/genetic diversity in the EABH. Dry and humid climatic phases alternated, contracting and expanding forests (Williamson, 1985; Trauth et al., 2007). During humid cycles, forest expansion may have connected humid forests of different mountains ranges or has at least facilitated dispersal between them, e.g., through the riverine forests (see Couvreur et al., 2021 for more details).
The EABH corresponds to the elevated regions of the East African Rift: from the Asir Mountains of southwest Saudi Arabia and the highlands of Yemen in the north; along the Eastern Rift branch including the Ethiopian Highlands, the Kenyan and Tanzanian Highlands and the Eastern Arc Mountains (EAM); and along the Western Rift branch including the Albertine Rift Mountains (ARM), the Southern Rift Mountains (SRM) and distant outliers in the Chimanimani Highlands of eastern Zimbabwe and Gorongosa of western Mozambique (Mittermeier et al., 2004; Figure 1A). The Eastern and Western Rift branches meet at the northern end of the Southern Rift Mountains, the so-called Livingstone Mountains (LM; also known as the Kipengere Range; Figure 1B). The LM are comprised of the Mbeya Range north of Mbeya, the Uporoto Mts. (including Mt. Ngozi) in the west, Mt. Rungwe, the Kitulo Plateau (formerly known as the Elton Plateau), the Kipengere Range (in the stricter sense) in the east and the Livingstone Mountains (in the stricter sense), including the Livingstone Escarpment along the north-eastern coast of Lake Nyasa, in the southwest (Figure 2).
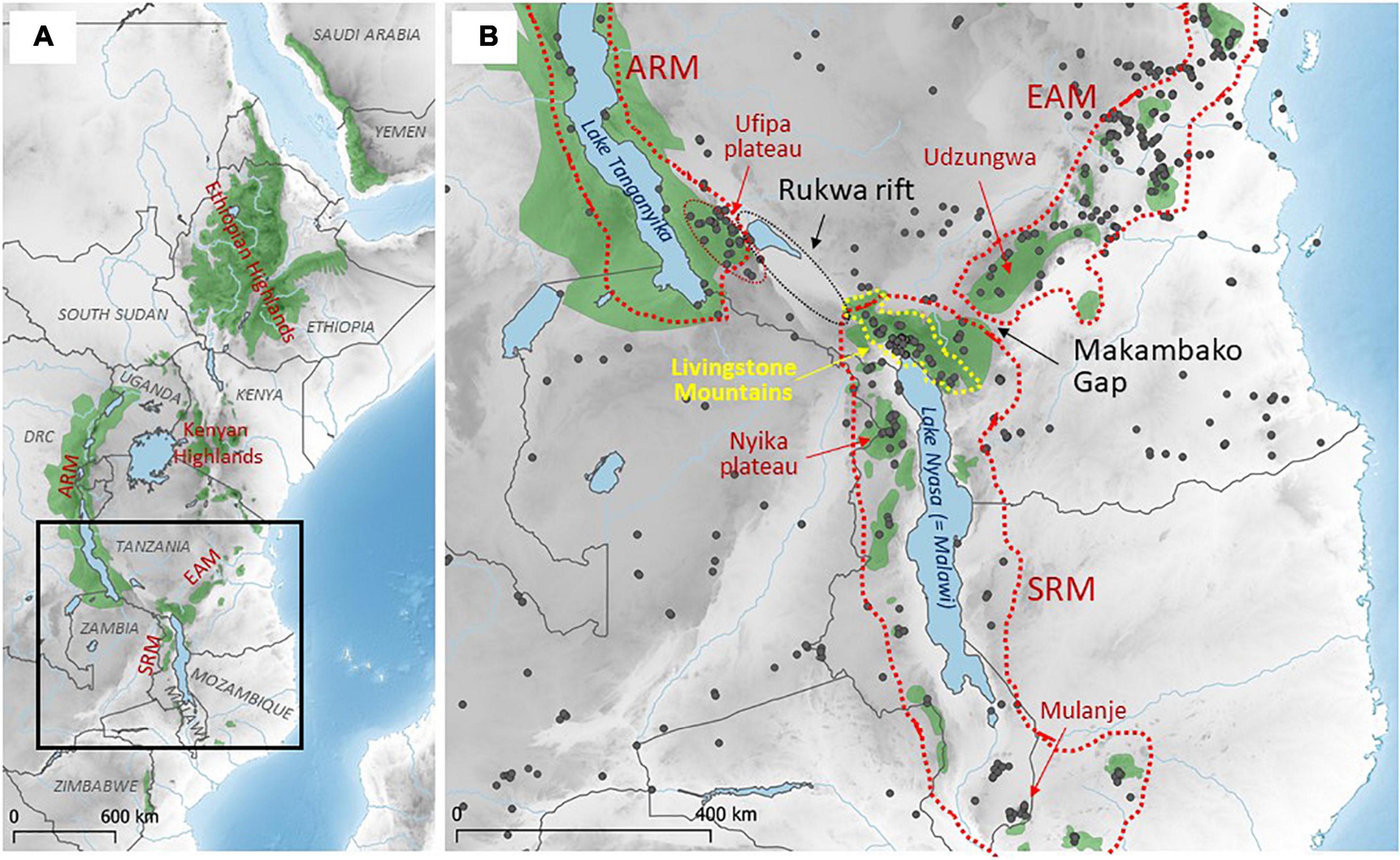
Figure 1. (A) Map of the Eastern Afromontane Biodiversity Hotspot (in green; following Birdlife International, 2012). Its most important larger blocks are the Ethiopian Highlands, Albertine Rift Mountains (ARM), Kenyan Highlands, Eastern Arc Mountains (EAM) and Southern Rift Mountains (SRM). (B) Detailed map of the study area, i.e., the Livingstone Mountains (LM) (yellow dotted line) and neighboring highland ecosystems (in green) and lowland savannah-like habitats. Gray dots are positions of genotyped small mammals relevant to this study. For a comparative analysis of highland taxa, we included samples from the SRM, the EAM and the southernmost part of the ARM. Two important gaps in the current distribution of montane ecosystems are marked by black arrows (i.e., areas of lowland savannahs, called the Rukwa rift and the Makambako Gap). For a comparative analysis of savannah/woodland taxa, we used data collected from these habitats in Tanzania, Zambia, Malawi and Mozambique.
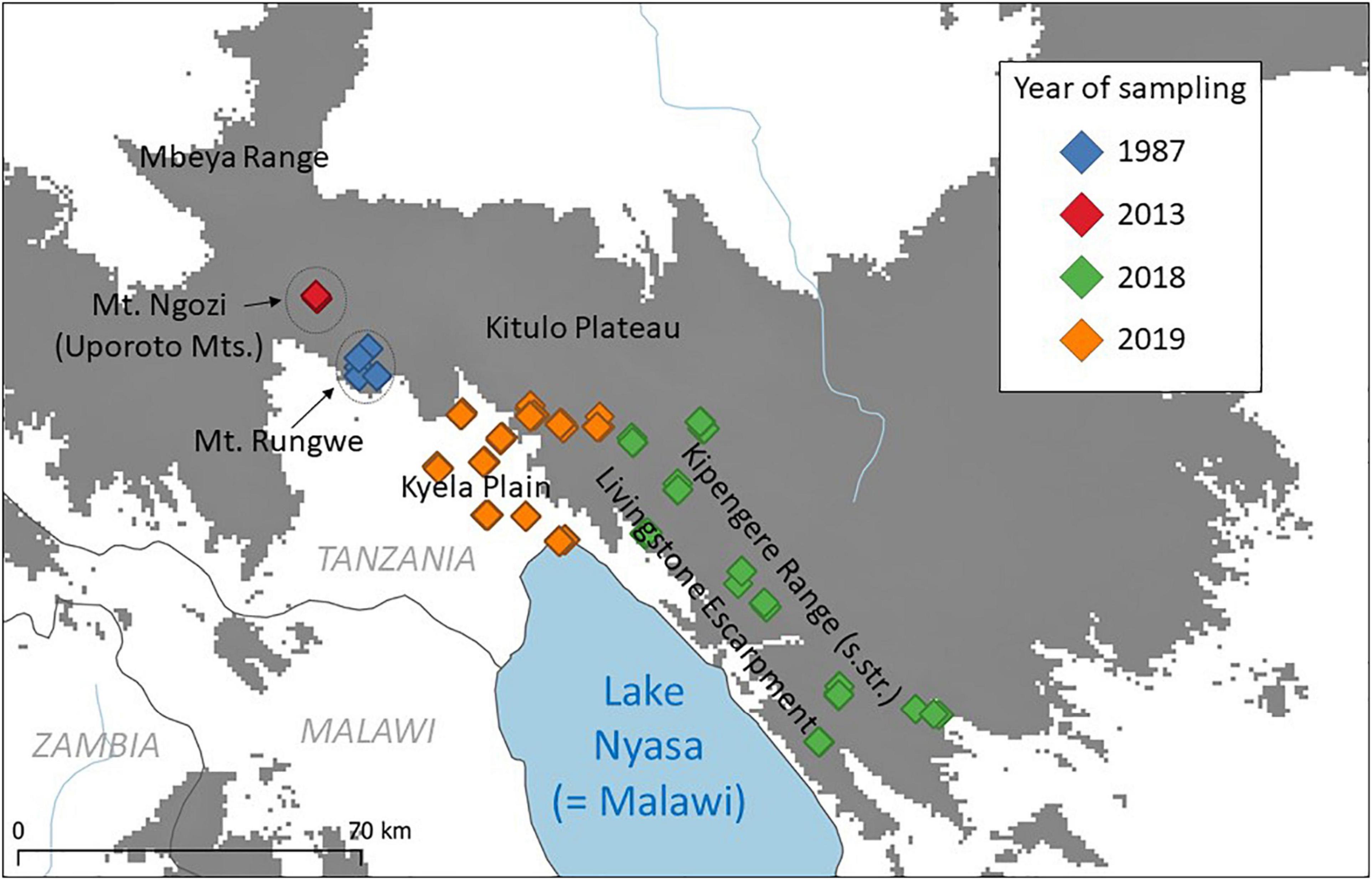
Figure 2. New localities sampled within this study in and around the Livingstone Mountains. Most samples were collected on Mt. Ngozi (in the Uporoto Mountains) in 2013 (red), and in the Livingstone Escarpment and the Kyela Plain at its base in 2018 (green) and 2019 (orange). We also present unpublished data from Mt. Rungwe (blue; collected in 1987 by W. Verheyen, J. Stuyck and M. Colyn) that is available online on the African Mammalia database (Van de Perre et al., 2019). Elevation above 1,500 m a.s.l. is colored in gray.
Small non-volant mammals (rodents and shrews) are very suitable models for reconstructions of historical biogeography, because they are relatively abundant and easy to capture, their dispersal capacity is limited and their affinity to particular types of habitats is very high. The unique location of the LM at the crossroads of the SRM and the intensively studied and species-rich ARM and EAM makes this understudied region a crucial area to improve our understanding of the biogeographic history of eastern Africa. Recently we performed multiple field expeditions and collected small mammals both in and around the LM. The focus of this paper is to compare phylogeographic patterns of small mammals living both in moist highland ecosystems and in drier savannahs and woodlands that surround them. Comparison of multiple mammalian taxa rather than focusing on one or two allows a more general picture on past habitat connectivity to be formed, as particular taxa might show different patterns due to variability in their habitat specificity, diet, dispersal ability etc. Specifically, we assess: (1) how unique the LM small mammals are in comparison with those on other montane islands of the SRM, the EAM and the ARM; and (2) the role of these mountains in forming a barrier to gene flow in savannah-dwelling taxa. The aim of this study is not to perform any taxonomic analysis or to test the role of any mechanisms behind the evolution of current biodiversity, but to compare general patterns in the distribution of genetic variability that have likely been created by changes in connectivity of montane forests and/or savannahs due to past climatic changes from the Pliocene onward.
Materials and Methods
Sampling
In order to assess the role of the LM in connecting different parts of the EABH and as a barrier to gene flow for savannah/woodland taxa, we used numerous georeferenced and genotyped specimens, 3538 published and 610 newly sampled or sequenced (Supplementary Material 1). The distribution of genetic diversity was analyzed for 17 taxa (at the level of closely related species and/or intraspecific lineages), which represent a significant part of assemblages of rodents and shrews that can be trapped by the use of standard live- and snap-traps (only few rarely sampled taxa were omitted because of the lack of material). In 2013 we collected small mammals in a montane forest and its margins on Mt. Ngozi using snap traps and Sherman live traps (H.B. Sherman Traps Inc., Tallahassee, United States). In 2018 and 2019 we captured small mammals in the Livingstone Escarpment and in the neighboring Kyela Plain (lowland at the north-western tip of Lake Nyasa at the foot of the Livingstone scarp) by Sherman live traps baited with a mixture of peanut butter and maize flour. Traps were usually set in and on the edge of agricultural fields in lines of 10 traps with equal distance in between or in half-grids of 5 by 10 traps. Live-caught small mammals were euthanized by cervical dislocation or an overdose of Isoflurane prior to dissection. Samples are stored in 96% ethanol at the Institute of Vertebrate Biology (IVB) of the Czech Academy of Sciences and at the University of Antwerp (UA). The field work was approved by the UA Ethical Committee for Animal Experimentation (2014-98 and 2017-75) and complied with regulations of the Research Policy of Sokoine University of Agriculture as stipulated in the Code of Conduct for Research Ethics. Because of geographical proximity, we also summarized unpublished data available in the African Mammalia database (Van de Perre et al., 2019) on animals collected on Mt. Rungwe in 1987 by W. Verheyen, J. Stuyck, and M. Colyn (Figure 2).
Genotyping and Phylogenetic Analysis
The analysis of genetic diversity is primarily based on the comparison of mitochondrial (mt) DNA sequences. There is a growing database of rodent and shrew mtDNA sequences for the cytochrome b gene (CYTB), enabling comparative analyses of genetic structure. Even though CYTB is only one non-recombining locus, which might introgress into other evolutionary lineages (e.g., between rodent taxa in geomorphologically complex Ethiopian Highlands; Bryja et al., 2018b; Komarova et al., 2021), it is an informative marker to explore genetic structure in the studied area, as demonstrated by comparison of mtDNA structure with that obtained by nuclear markers (Bryja et al., 2014b,2017; Krásová et al., 2019; Onditi et al., 2021) or even genomic approaches (our unpublished data). Therefore, we performed comparative analysis of phylogeographic structure using CYTB as a main marker. DNA was extracted using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) for the 610 newly sampled small mammals and the complete CYTB gene (1,140 bp) was amplified using L14723 and H15915 primers (Lecompte et al., 2002) according to the protocols described in Bryja et al. (2014b). The sequences were aligned with all relevant published data (including those in GenBank and the African Mammalia database; Van de Perre et al., 2019). The alignments are available upon request, new sequences were submitted to GenBank (accession numbers OK093418-OK094020). For a list of all included georeferenced and genotyped samples (see Supplementary Material 1).
Species and intraspecific clades in particular groups of small mammals were identified by ML analysis in RAxML 8.2.8 (Stamatakis, 2014), using the GTRCAT substitution model as suggested by the authors of the program. The robustness of the nodes was evaluated by the default bootstrap procedure with 1,000 replicates and new sequences were considered as part of previously defined taxa/clades if the bootstrap support was > 95%. Specifically, we compared the new sequences with relevant data in recent phylogenetic studies (Castiglia et al., 2012; Colangelo et al., 2013; Bryja et al., 2014a,b; Stanley et al., 2015; Bryja et al., 2017, 2018a; Mazoch et al., 2018; Petružela et al., 2018; Sabuni et al., 2018; Krásová et al., 2019, 2021; Hánová et al., 2021a,b; Onditi et al., 2021; Dianat et al., unpublished data) and/or with unpublished sequences obtained by our teams in East Africa in the last decade. The distribution maps of genetic clades were created using the Free and Open Source QGIS v. 3.16 (QGIS.org, 2021).
Results
Small Mammals of the Livingstone Mountains
The field collections during four expeditions in the LM comprised 992 small mammals, classified into 4 shrew and 20 rodent species (Table 1). They represent taxa living in a wide spectrum of habitats. Besides the taxa typical for montane forests (Montemys, Hylomyscus), forest margins (Mus triton, Lophuromys, Grammomys) and humid grasslands (Rhabdomys, Dendromus, Crocidura montis group), we also collected taxa living in savannahs (Mus minutoides, Saccostomus, Acomys, Crocidura hirta/flavescens). The combination of newly collected genetic data with recently published phylogeographical studies thus allows assessment of the role of the LM in creating patterns of genetic diversity in both highland and lowland types of ecosystems (summarized in Figures 3, 4). We did not include taxa with very limited data or taxa without georeferenced and genotyped samples in the LM area (with the exception of Heliophobius and two species of Aethomys that were not captured during our survey, but are very likely to live in the area). Two species, Lemniscomys striatus and Crocidura hildegardeae, are reported for the first time in this area and these records push the boundary of their known distribution range more southwards.
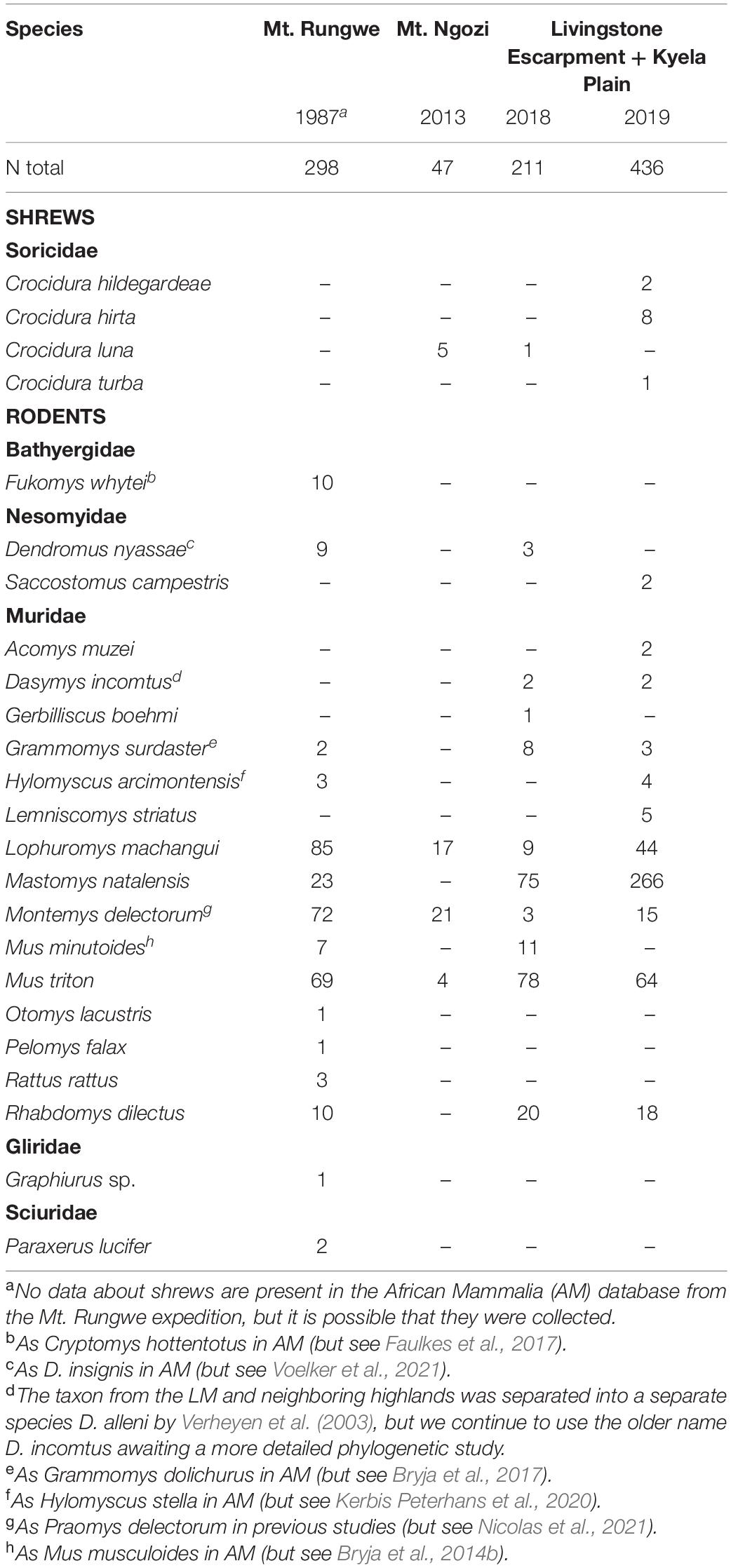
Table 1. Small mammals collected in the Livingstone mountains and the Kyela plain (see Figure 2 for sampled localities).
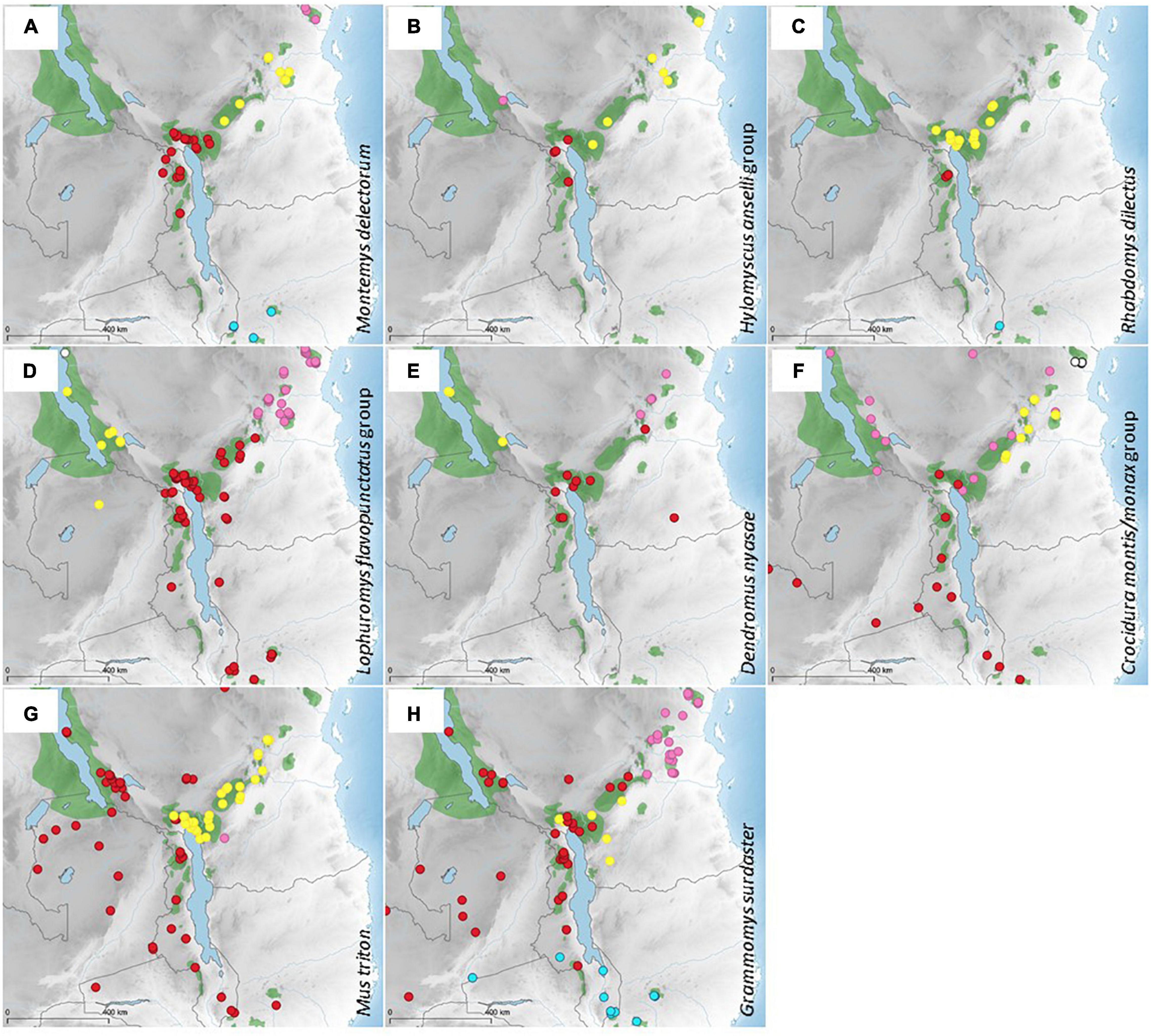
Figure 3. Distribution of genetic clades of eight taxa of small mammals living in humid montane forests and their margins in the Livingstone Mountains. Published data were complemented by new records from the studied area and reanalyzed. For more details (data source, GPS coordinates, GenBank accession numbers, etc.) see Supplementary Material 1. Different symbols/colors represent genetic clades delimited as follows: (A) Montemys delectorum (based on Bryja et al., 2014a and Sabuni et al., 2018: blue—M. delectorum clade A, red—M. delectorum clade B1, yellow—M. delectorum clade B2, pink—M. delectorum clade C). (B) Hylomyscus anselli group (based on Kerbis Peterhans et al., 2020; Nicolas et al., 2020: pink—H. stanleyi, yellow—H. arcimontensis clade 1, red—H. arcimontensis clade 2). (C) Rhabdomys dilectus dilectus (based on Castiglia et al., 2012; Sabuni et al., 2018: yellow—R. d. dilectus clade A1, blue—R. d. dilectus clade A2, red—R. d. dilectus clade A3). (D) Lophuromys flavopunctatus group (based on Sabuni et al., 2018; Onditi et al., 2021: red—L. machangui, pink—L. kilonzoi, yellow—L. sabunii, white—L. laticeps). (E) Dendromus nyasae (based on Voelker et al., 2021: yellow—D. nyasae clade 1, red—D. nyasae clade 2, pink—D. nyasae clade 3). (F) Crocidura montis/monax group (based on Stanley et al., 2015; Sabuni et al., 2018: pink—C. montis clade 1, red—C. montis clade 3, i.e., C. luna, yellow—C. munissii, white—C. tansaniana, blue—C. usambarae). (G) Mus triton (based on Krásová et al., 2019: yellow—M. triton clade C, red—M. triton clade D, pink—M. triton clade E). (H) Grammomys surdaster (based on Bryja et al., 2017 and modified on the basis of unpublished genomic ddRAD analysis: red—G. surdaster clades su3, su4, su7, su8-North grouped together based on ddRAD, yellow—G. surdaster clade su9, blue—G. surdaster clade su8-South, pink—G. surdaster clade su10, except for the southernmost locality that has mtDNA su9, but clearly groups with the pink clade at nuclear genomic data).
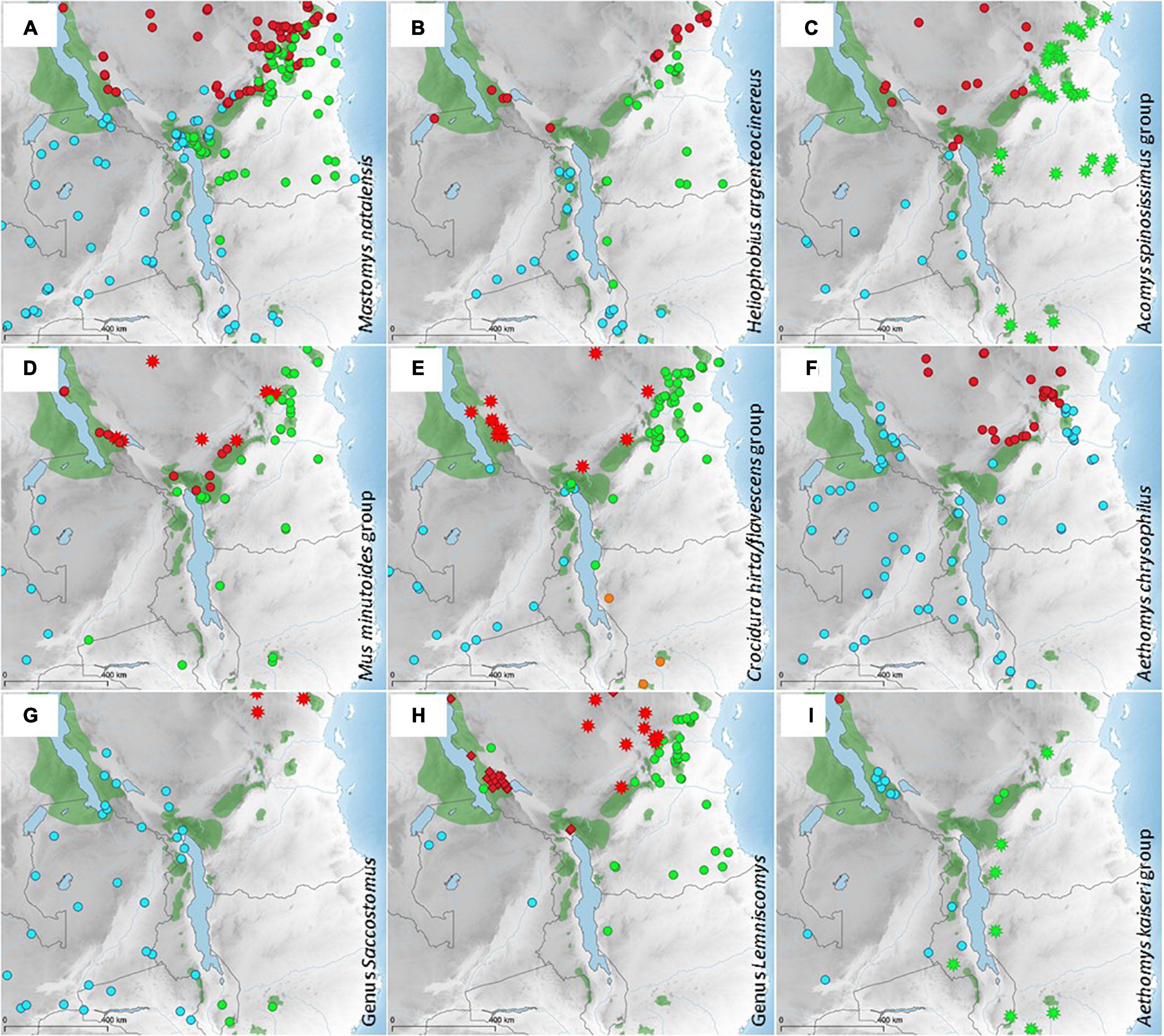
Figure 4. Distribution of genetic clades of nine taxa of small mammals living in savannahs and woodlands (e.g., miombo or mopane woodlands) around the Livingstone Mountains (LM). Published data were complemented by new records from the studied area and reanalyzed. For more details (data source, GPS coordinates, GenBank accession numbers, etc.) (see Supplementary Material 1). Different symbols/colors represent the following genetic clades: (A) Mastomys natalensis (based on Colangelo et al., 2013; Hánová et al., 2021a): red—M. natalensis clade B-IV, green—M. natalensis clade B-V, blue—M. natalensis clade B-VI). (B) Heliophobius argenteocinereus (from Bryja et al., 2018a; Uhrová et al., 2021: red—H. argenteocinereus clade N, green—H. argenteocinereus clade SE, blue—H. argenteocinereus clade SW). (C) Acomys spinosissimus group (based on Petružela et al., 2018: red—A. muzei clade m4, blue—A. muzei other clades, green stars—A. ngurui). (D) Mus minutoides group (based on Bryja et al., 2014b: red—M. minutoides clade TZw, green—M. minutoides clade SE, blue—M. minutoides clade ZA, red stars—M. cf. gerbillus). (E) Crocidura hirta/flavescens complex (based on Dianat et al., unpublished data: red—C. cf. flavescens, green—C. hirta clade I, blue—C. hirta clade II, orange—C. hirta clade III). (F) Aethomys chrysophilus (based on Mazoch et al., 2018: red—A. chrysophilus clade E, blue—A. chrysophilus clade G). (G) Saccostomus (based on Mikula et al., 2016: blue—S. campestris campestris clade B, green S. campestris campestris clade C, red stars—S. umbriventer. (H) Lemniscomys (based on Hánová et al., 2021b: blue—L. cf. roseveari, green—L. rosalia clade A, red rhombuses—L. striatus clade F, red stars—L. zebra clade B). (I) Aethomys kaiseri group (based on Krásová et al., 2021: blue—A. kaiseri clade I, green—A. kaiseri clade II, red—A. kaiseri clade III, green stars—A. silindensis.
Comparative Genetic Structure of Taxa Living in Humid Highland Habitats of the Livingstone Mountains
The distribution of evolutionary clades of taxa living in relatively humid highland habitats (montane forest and their margins) is summarized in Figure 3. There is no universal pattern across the eight analyzed taxa. The Montemys delectorum clade occurring in the LM is also found in SRM in northern Malawi. A very distinct clade occurs in the southern outliers of the SRM and two more clades are found in the EAM, one from the Udzungwa Mts. up to the Uluguru and Ukaguru Mts. and a second in the Usambara Mts. and northern Tanzanian highlands (Figure 3A). Another montane forest specialist, Hylomyscus arcimontensis, probably has a more restricted distribution (i.e., it is not recorded in the northern Tanzanian highlands and the southernmost part of the SRM) and the break from one clade to another occurs within the LM rather than between the LM and EAM. The Ufipa plateau (Mbizi Mts., already part of the ARM) is inhabited by another species from the same species group, H. stanleyi (Figure 3B).
The following species are not strictly associated with forests, but they live in the mosaic of highland forests and open moist grassy habitats. The SRM Rhabdomys dilectus dilectus are divided in three clades, one in the LM, which also occurs in the Udzungwa Mts. (EAM), one in the Nyika Plateau and one in the Mulanje Mts. (Figure 3C). The Lophuromys flavopunctatus group (sometimes also called L. aquilus group) has been split into several species with a parapatric distribution in the EABH. Lophuromys machangui lives in the SRM (including the LM) and the Udzungwa Mts, while L. kilonzoi lives in the rest of the EAM, L. sabuni in the southern part of the ARM including the Ufipa Plateau, and L. laticeps more north in the ARM (Figure 3D). Three clades of Dendromus nyasae, living in highland grasslands, have a very similar distribution to three species of Lophuromys (Figure 3E). Crocidura montis clade 3 (i.e., C. luna) was captured in the SRM, including the west of the LM. In the east of the LM, the ARM and in the Udzungwa Mts., C. montis clade 1 was captured. A further three species from the Crocidura montis/monax group from the EAM were delimited by integrative taxonomic analysis, C. munissi, C. tansaniana, and C. usambarae (Figure 3F).
The last two species are more opportunistic, and besides montane forests they can also occupy drier habitats, such as miombo woodland. Most Mus triton caught in the LM belong to the same clade as those found in the EAM, but two individuals caught in 1987 around Mt. Rungwe belong to the same clade as those found in the other SRM and the ARM (Figure 3G). An individual captured east of the LM forms a distinct genetic clade. For Grammomys surdaster, we clustered the mtDNA clades described in Bryja et al. (2017) into four groups based on unpublished ddRAD analyses (Bryja et al., unpublished data). The widespread genomic multilocus clade found in the ARM and Zambia occurs also in the SRM, including the LM, and Udzungwa. A separate group was found in lower elevations of the western LM and east of the LM and Udzungwa. Two additional genetic clades inhabit the southernmost part of the SRM and the central and northern EAM (Figure 3H).
Comparative Genetic Structure of Taxa Living in Savannahs and Woodlands Around the Livingstone Mountains
For savannah and woodland taxa around the LM, the patterns are more uniform. In most taxa, there are three genetic groups, even though their genetic distances are not comparable across taxa (as suggested in the original phylogenetic-taxonomic publications, for more details see Supplementary Material 2): (1) a group in north-western/central Tanzania; (2) a group in south-eastern Tanzania and north-eastern Mozambique; and (3) a group in Zambia, Malawi, the rest of Mozambique and part of south-western Tanzania (Figure 4). The groups (1) vs. (2) often appear separated by the EAM, (1) vs. (3) by the LM and/or the Ufipa Plateau, and (2) vs. (3) by Lake Nyasa and the LM. For example, in the Crocidura hirta/flavescens complex, C. cf. flavescens occurs north of the EAM, the LM and on the Ufipa Plateau, C. hirta clade I occurs around the EAM and east of the LM and Lake Nyasa, and C. hirta clade II occurs southwest of the LM and south of the Ufipa Plateau (Figure 4E). Another clear example is Heliophobius argenteocinereus where the N clade is found north of the EAM, northwest of the LM and in the ARM including the Ufipa Plateau, the SE clade is found south east of the EAM, the LM and Lake Tanganyika; and the SW clade is found southwest of the LM, west of Lake Nyasa and more to the south (Figure 4B).
There are some variations on this general pattern. For example, in several taxa, the distribution of the south-western (M. natalensis, A. chrysophilus, S. campestris, A. kaiseri) or south-eastern (L. rosalia) group extends up until the Ufipa Plateau (Figures 4A,F–I, respectively), where it can meet the northern clade (in M. natalensis, Figure 4A). For some taxa, only two groups can be distinguished with current data. For example, the same clade of Aethomys chrysophilus is found in southern Tanzania, Zambia, Malawi and Mozambique, as opposed to northern and central Tanzania (Figure 4F). For the Acomys spinosissimus group, A. muzei is found in north-western Tanzania, Zambia and Malawi, as opposed to A. ngurui in south-eastern Tanzania and Mozambique (Figure 4C). However, if we split the internal Tanzanian crown clade m4 of A. muzei from the other A. muzei clades, we do see the typical geographical pattern again. Two generalist species were also caught in agricultural fields in the LM. For M. natalensis a different clade was captured on the Livingstone Escarpment compared to the area around Mt. Rungwe and on the Kyela Plain. For M. minutoides one clade is observed rather central on the Livingstone Escarpment, while another occurs more south and at a locality on the Kyela Plain (Figure 4D).
Discussion
The Livingstone Mountains: Diversity Hotspots Within the Eastern Afromontane Biodiversity Hotspot
Compared to the extremely rich biodiversity of ARM and EAM, the SRM are generally considered less diverse (Mittermeier et al., 2004). However, the LM as the northernmost tip of the SRM are uniquely positioned at the crossroads of all three mountain blocks, which may substantially increase diversity of its biota. While the humid Afromontane forests of the EABH are currently isolated in an archipelago-like fashion, faunal exchanges likely took place during more humid periods of the Pleistocene, allowing range expansion and gene flow in taxa adapted to moist habitats currently restricted to patches at higher elevations (e.g., Bryja et al., 2014a). The LM could therefore have acted as stepping stones for the colonization/migration between different parts of the EABH and they still harbor a mosaic of (intra) specific clades from the neighboring EABH complexes (Figure 3). There is no simple pattern of today’s genetic structure across different taxa. As small mammals vary in many different traits, such as level of habitat specialization and dispersal ability, they have reacted differently to the same climate-driven habitat shifts and certain geographic features might have been a barrier to some, but not to others. It should be noted that several small mammals living in the mosaic of humid highland habitats (i.e., those shown in Figure 3) were also caught in the edges of agricultural fields. The genetic structure of more opportunistic species that also might live outside of montane forests such as M. triton or G. surdaster, already seems less spatially segregated compared to taxa with more specific habitat requirements such as M. delectorum, H. arcimontensis or the L. flavopunctatus group.
Despite differences among individual taxa, the comparison of genetic structure of multiple species living in the same sky islands in the EABH allows identification of geomorphological features that have been important as barriers to gene flow in the past. Here we assume a barrier if a given taxon only occurs in the LM, but not in the neighboring mountain ranges. Conversely, a taxon that occurs in the LM and in the ARM and/or EAM shows connectivity between those mountain ranges in the past.
The Makambako Gap is an unforested, dry, low-lying region covered by miombo woodland separating the LM (i.e., the SRM) and the EAM (Figure 1B). It has been shown to be an important barrier for a range of taxa, such as Tornier’s cat snake (Gravlund, 2002), some bush vipers (Menegon et al., 2014), double-bearded chameleons (Ceccarelli et al., 2014), mountain greenbuls (Roy et al., 1998, reviewed in Kahindo et al., 2007; Fjeldså and Bowie, 2008), olive woodpeckers (Fuchs et al., 2021), and Tanzanian vlei rats (Taylor et al., 2009), but not for others, e.g., Uzungwe Mountain Bush Vipers (Menegon et al., 2014), forest batis (Fjeldså et al., 2006, reviewed in Kahindo et al., 2007; Fjeldså and Bowie, 2008), green barbets (Fjeldså and Bowie, 2008; Fjeldså et al., 2010) and Kihaule’s mouse shrews (Stanley and Esselstyn, 2010). Similar conclusions can be drawn from our review of small mammals. The Makambako Gap has probably been (and currently clearly is) a Quaternary barrier to M. delectorum and some, but not all clades of the C. montis/monax group. On the other hand, H. arcimontensis, R. dilectus dilectus, L. machangui, and M. triton clade C do bridge the gap. Furthermore, some of our clades (R. dilectus dilectus clade A1, M. triton clade C and G. surdaster clade su9) only occur in the LM and the Udzungwa Mts., but were not trapped further south. This is also the case for certain bush vipers (Menegon et al., 2014), kipunjis (Davenport et al., 2006), and Kinyongia msuyae chameleons (Menegon et al., 2015). It should also be noted that the savannahs separating the central and northern EAM are often more important in forming the genetic structure of highland taxa than the Makambako Gap (Figure 3; see Fjeldså and Bowie, 2008 for a review of bird taxa).
The Rukwa rift is a grassland and woodland basin, separating montane habitats of the ARM from the SRM (Figure 1B). For small terrestrial mammals, the fauna of southern part of the ARM is very distinct from eastern mountains (i.e., the EAM and the SRM). They share the same mitochondrial lineages only in relatively opportunistic taxa that can live also in higher-elevation miombo woodlands, e.g., M. triton, G. surdaster and the C. montis/monax group. The situation is different in more specialized taxa living in high-elevation humid habitats. The genera Rhabdomys and Montemys do not occur in the ARM at all (the latter is replaced there by its ecological equivalent Praomys jacksoni; Mizerovská et al., 2019). The SRM species in the genera Hylomyscus and Lophuromys are replaced in the ARM by phylogenetically distant congeneric species (Kerbis Peterhans et al., 2020; Onditi et al., 2021). In other taxa, the phylogenetic affinities probably also depend on the level of specialization to montane forests. The Rukwa rift appears to have been a barrier for some mountain greenbuls (Roy et al., 1998; reviewed in Kahindo et al., 2007), the Cinnyris afer complex (Bowie et al., 2016), some bush vipers (Menegon et al., 2014) and the epiphytic plant Canarina eminii (Mairal et al., 2017), but not to Tanzanian vlei rats (Taylor et al., 2009) and Mt. Rungwe bush vipers (Menegon et al., 2014).
The Livingstone Mountains: Drivers of Diversification in Low Elevation Savannahs and Woodlands
While an expansion of moist montane forests during more humid periods of the Pleistocene likely connected taxa living in these forests and their edges, at the same time it effectively isolated the taxa living in the surrounding savannahs and woodlands (Colangelo et al., 2013; Mazoch et al., 2018; Petružela et al., 2018). These allopatric taxa could then diverge from each other in their respective “savannah refugia” and meet again during drier periods, such as the present. This seems to have occurred for a wide range of taxa living in open savannah-like habitats, e.g., ungulates (Lorenzen et al., 2012), baboons (Dunn et al., 2013), and fiscal shrikes (Fuchs et al., 2011). Moreover, the geographical areas where the clades that diverged in allopatry come into secondary contact, are roughly the same for many taxa and may therefore represent suture zones. In eastern Africa these zones are mostly located perpendicular to the main mountain ranges in the north-south direction (Figure 4). However, despite geographical coincidence across multiple taxa, there is variability in the level (i.e., time) of divergence, and the taxa have not diverged to the same extent. As a result, their secondary contact ranges from free admixture to limited hybridization to reproductive isolation (Lorenzen et al., 2012).
In our data we indeed observe that the humid Afromontane mountains are/were a barrier to gene flow for taxa living in drier, savannah-like ecosystems. Even though some clades are currently in contact, the EAM, the LM, Lake Nyasa and the Ufipa Plateau appear to have separated many taxa into three genetic groups (Figure 4). The different clades often represent intraspecific variation (indicated by dot symbols in Figure 4) driven by relatively recent Pleistocene climate oscillations, but in some cases, they represent well-distinguished species (indicated by stars or rhombuses). In two groups, the Acomys spinosissimus group and the Aethomys kaiseri group, the most distinct taxa (A. ngurui and A. silindensis, respectively) are limited to south-eastern savannahs/miombo woodlands (Petružela et al., 2018; Krásová et al., 2021; green stars in Figures 4C,I). However, this pattern is not the most prevalent. Environmentally speaking the ecological conditions are most distinct in the so-called Masai steppe in central Tanzania, and accordingly, the northern (red in Figure 4) groups are phylogenetically more distinct in many taxa. This is the case for Mus cf. gerbillus (vs. three intraspecific clades of M. minutoides; Bryja et al., 2014b; Figure 4D), Crocidura cf. flavescens (vs. three intraspecific clades of C. hirta; Dianat et al., unpublished data; Figure 4E), Saccostomus umbriventer (vs. two intraspecific clades of S. campestris campestris; Mikula et al., 2016; Figure 4G), Lemniscomys striatus and L. zebra (very distinct from two lineages of the L. griselda group, i.e., L. cf. roseveari and L. rosalia; Hánová et al., 2021b; Figure 4H). In two additional taxa (Heliophobius and Aethomys chrysophilus), the northern group is not yet taxonomically separated into a different species, but the northern populations are genetically and morphologically the most distinct and deserve a separate species status (Mazoch et al., 2018; Uhrová et al., 2021). The most striking exception from this pattern is Acomys muzei, which has a very high genetic diversity in Malawi and Zambia (Petružela et al., 2018). One internal clade of this species relatively recently colonized western and central Tanzania (Figure 4C). No other Acomys sp. are present there, so it may have filled an empty niche. Whether or not this colonization went hand in hand with adaptations to more arid environments is a hypothesis worth testing.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: GenBank, OK093418-OK094020.
Ethics Statement
The animal study was reviewed and approved by the UA Ethical Committee for Animal Experimentation (2014-98 and 2017-75).
Author Contributions
LC, RŠ, SJEB, JGB, and JB conceived the study. HL, SJEB, JGB, and JB provided funding. LC, CS, RŠ, TA, EL, JGB, and JB collected the material. LC and TA performed genetic analyses. LC and JB analyzed the data and wrote the first draft of the manuscript that was complemented by all authors. All authors also approved the final version of the manuscript.
Funding
This study was supported by the Czech Science Foundation (18-19629S, 20-07091J) and the Research Foundation—Flanders (FWO; G0A4815N). LC was a Ph.D. fellow with the FWO (1171921N). This collaborative work was also part of the Czech Republic—Flanders, Belgium Mobility projects 2020 (FWO number VS07521N and CAS number FWO-21-02).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank S. Gryseels, A. Hánová, Z. Hiadlovská, A. S. Katakweba, K. Kibwana, J. Komárková, H. Konvičková, A. Konečný, J. Krásová, J. Kreisinger, M. Lövy, V. Mazoch, O. Mikula, G. Phamphi, F. Sedláček J. Šklíba, R. Smolinský, S. R. M. Van den Burg, and J. Zima Jr. for their help in the field, and J. Ademola, A. Saanya, W. T. Stanley (†) and the FMNH (Field Museum of Natural History in Chicago) for providing additional samples.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.742851/full#supplementary-material
Abbreviations
AM, African Mammalia (database); ARM, Albertine Rift Mountains; CYTB, cytochrome b; EABH, Eastern Afromontane Biodiversity Hotspot; EAM, Eastern Arc Mountains; LM, Livingstone Mountains; SRM, Southern Rift Mountains.
References
Birdlife International (2012). Eastern Afromontane Biodiversity Hotspot. Cambridge: Birdlife International.
Bowie, R. C. K., Fjeldså, J., Kiure, J., and Kristensen, J. B. (2016). A new member of the greater double-collared sunbird complex (Passeriformes: Nectariniidae) from the Eastern Arc Mountains of Africa. Zootaxa 4175, 023–042. doi: 10.11646/zootaxa.4175.1.3
Bryja, J., Konvičková, H., Bryjová, A., Mikula, O., Makundi, R., Chitaukalie, W. N., et al. (2018a). Differentiation underground: range-wide multilocus genetic structure of the silvery mole-rat does not support current taxonomy based on mitochondrial sequences. Mamm. Biol. 93, 82–92. doi: 10.1016/j.mambio.2018.08.006
Bryja, J., Kostin, D., Meheretu, Y., Šumbera, R., Bryjová, A., Kasso, M., et al. (2018b). Reticulate pleistocene evolution of Ethiopian rodent genus along remarkable altitudinal gradient. Mol. Phylogenet. Evol. 118, 75–87. doi: 10.1016/j.ympev.2017.09.020
Bryja, J., Mikula, O., Patzenhauerová, H., Oguge, N. O., Šumbera, R., and Verheyen, E. (2014a). The role of dispersal and vicariance in the Pleistocene history of an East African mountain rodent, Praomys delectorum. J. Biogeogr. 41, 196–208. doi: 10.1111/JBI.12195
Bryja, J., Mikula, O., Šumbera, R., Meheretu, Y., Aghová, T., Lavrenchenko, L. A., et al. (2014b). Pan-African phylogeny of Mus (subgenus Nannomys) reveals one of the most successful mammal radiations in Africa. BMC Evol. Biol. 14:256. doi: 10.1186/s12862-014-0256-2
Bryja, J., Šumbera, R., Kerbis Peterhans, J. C., Aghová, T., Bryjová, A., Mikula, O., et al. (2017). Evolutionary history of the thicket rats (genus Grammomys) mirrors the evolution of African forests since late Miocene. J. Biogeogr. 44, 182–194. doi: 10.1111/jbi.12890
Castiglia, R., Solano, E., Makundi, R. H., Hulselmans, J., Verheyen, E., and Colangelo, P. (2012). Rapid chromosomal evolution in the mesic four-striped grass rat Rhabdomys dilectus (Rodentia, Muridae) revealed by mtDNA phylogeographic analysis. J. Zool. Syst. Evol. Res. 50, 165–172. doi: 10.1111/J.1439-0469.2011.00627.X
Ceccarelli, F. S., Menegon, M., Tolley, K. A., Tilbury, C. R., Gower, D. J., Laserna, M. H., et al. (2014). Evolutionary relationships, species delimitation and biogeography of Eastern Afromontane horned chameleons (Chamaeleonidae: Trioceros). Mol. Phylogenet. Evol. 80, 125–136. doi: 10.1016/j.ympev.2014.07.023
Coetzee, J. A. (1993). “African flora since the terminal Jurassic,” in Biological Relationships between Africa and South America, ed. P. Goldblatt (New Haven, CT: Yale University Press), 37–61. doi: 10.2307/j.ctt22726mc
Colangelo, P., Verheyen, E., Leirs, H., Tatard, C., Denys, C., Dobigny, G., et al. (2013). A mitochondrial phylogeographic scenario for the most widespread African rodent, Mastomys natalensis. Biol. J. Linn. Soc. 108, 901–916. doi: 10.1111/bij.12013
Couvreur, T. L. P., Dauby, G., Blach-Overgaard, A., Deblauwe, V., Dessein, S., Droissart, V., et al. (2021). Tectonics, climate and the diversification of the tropical African terrestrial flora and fauna. Biol. Rev. 96, 16–51. doi: 10.1111/brv.12644
Davenport, T. R., Stanley, W. T., Sargis, E. J., De Luca, D. W., Mpunga, N. E., Machaga, S. J., et al. (2006). A new genus of African monkey, Rungwecebus: morphology, ecology, and molecular phylogenetics. Science 312, 1378–1381. doi: 10.1126/science.1125631
Demos, T. C., Kerbis Peterhans, J. C., Agwanda, B., and Hickerson, M. J. (2014). Uncovering cryptic diversity and refugial persistence among small mammal lineages across the Eastern Afromontane biodiversity hotspot. Mol. Phylogenet. Evol. 71, 41–54. doi: 10.1016/J.YMPEV.2013.10.014
Dunn, J., Cardini, A., and Elton, S. (2013). Biogeographic variation in the baboon: dissecting the cline. J. Anat. 223, 337–352. doi: 10.1111/joa.12085
Faulkes, C. G., Mgode, G. F., Archer, E. K., and Bennett, N. C. (2017). Relic populations of Fukomys mole-rats in Tanzania: description of two new species F. livingstoni sp. nov. and F. hanangensis sp. nov. PeerJ 5:e3214. doi: 10.7717/peerj.3214
Fjeldså, J., and Bowie, R. C. K. (2008). New perspectives on the origin and diversification of Africa’s forest avifauna. Afr. J. Ecol. 46, 235–247. doi: 10.1111/j.1365-2028.2008.00992.x
Fjeldså, J., Bowie, R. C. K., and Kiure, J. (2006). The forest batis, Batis mixta, is two species: description of a new, narrowly distributed Batis species in the Eastern Arc biodiversity hotspot. J. Ornithol. 147, 578–590. doi: 10.1007/s10336-006-0082-4
Fjeldså, J., Kiure, J., Doggart, N., Hansen, L. A., and Perkin, A. (2010). Distribution of highland forest birds across a potential dispersal barrier in the Eastern Arc Mountains of Tanzania. Steenstrupia 32, 1–43.
Fuchs, J., Bowie, R. C. K., Melo, M., Boano, G., Pavia, M., and Fjeldså, J. (2021). Phylogeographical history of the Olive Woodpecker Dendropicos griseocephalus, a species widely distributed across Africa. Ibis 163, 417–428. doi: 10.1111/ibi.12875
Fuchs, J., Crowe, T. M., and Bowie, R. C. (2011). Phylogeography of the fiscal shrike (Lanius collaris): a novel pattern of genetic structure across the arid zones and savannas of Africa. J. Biogeogr. 38, 2210–2222. doi: 10.1111/j.1365-2699.2011.02545.x
Gravlund, P. (2002). Molecular phylogeny of Tornier’s cat snake (Crotaphopeltis tornieri), endemic to East African mountain forests: biogeography, vicariance events and problematic species boundaries. J. Zool. Syst. Evol. Res. 40, 46–56. doi: 10.1046/j.1439-0469.2002.00175.x
Greenbaum, E., Portillo, F., Jackson, K., and Kusamba, C. (2015). A phylogeny of Central African Boaedon (Serpentes: Lamprophiidae), with the description of a new cryptic species from the Albertine rift. Afr. J. Herpetol. 64, 18–38. doi: 10.1080/21564574.2014.996189
Hánová, A., Konečný, A., Mikula, O., Bryjová, A., Šumbera, R., and Bryja, J. (2021a). Diversity, distribution and evolutionary history of the most studied African rodents, multimammate mice of the genus Mastomys: a review after quarter of century of using DNA sequencing. J. Zool. Syst. Evol. Res. 1–19. doi: 10.1111/jzs.12569
Hánová, A., Konečný, A., Nicolas, V., Denys, C., Granjon, L., Lavrenchenko, L. A., et al. (2021b). Multilocus phylogeny of African striped grass mice (Lemniscomys): stripe pattern only partly reflects evolutionary relationships. Mol. Phylogenet. Evol. 155:107007. doi: 10.1016/J.YMPEV.2020.107007
Kahindo, C., Bowie, R. C. K., and Bates, J. M. (2007). The relevance of data on genetic diversity for the conservation of Afro-montane regions. Biol. Conserv. 134, 262–270. doi: 10.1016/j.biocon.2006.08.019
Kerbis Peterhans, J. C., Hutterer, R., Doty, J. B., Malekani, J. M., Moyer, D. C., Krásová, J., et al. (2020). Four new species of the Hylomyscus anselli group (Mammalia: Rodentia: Muridae) from the Democratic Republic of Congo and Tanzania. Bonn Zool. Bull. 69, 55–83. doi: 10.20363/BZB-2020.69.1.055
Komarova, V. A., Kostin, D. S., Bryja, J., Mikula, O., Bryjová, A., Čížková, D., et al. (2021). Complex reticulate evolution of speckled brush-furred rats (Lophuromys) in the Ethiopian centre of endemism. Mol. Ecol. 30, 2349–2365. doi: 10.1111/mec.15891
Krásová, J., Mikula, O., Bryja, J., Baptista, N. L., António, T., Aghová, T., et al. (2021). Biogeography of Angolan rodents: the first glimpse based on phylogenetic evidence. Divers. Distrib. 27, 2571–2583. doi: 10.1111/ddi.13435
Krásová, J., Mikula, O., Mazoch, V., Bryja, J., Říčan, O., and Šumbera, R. (2019). Evolution of the Grey-bellied pygmy mouse group: highly structured molecular diversity with predictable geographic ranges but morphological crypsis. Mol. Phylogenet. Evol. 130, 143–155. doi: 10.1016/J.YMPEV.2018.10.016
Lecompte, E., Granjon, L., Kerbis Peterhans, J., and Denys, C. (2002). Cytochrome b-based phylogeny of the Praomys group (Rodentia, Murinae): a new African radiation? C. R. Biol. 325, 827–840. doi: 10.1016/S1631-0691(02)01488-9
Lorenzen, E. D., Heller, R., and Siegismund, H. R. (2012). Comparative phylogeography of African savannah ungulates. Mol. Ecol. 21, 3656–3670. doi: 10.1111/j.1365-294X.2012.05650.x
Mairal, M., Sanmartín, I., Herrero, A., Pokorny, L., Vargas, P., Aldasoro, J. J., et al. (2017). Geographic barriers and Pleistocene climate change shaped patterns of genetic variation in the Eastern Afromontane biodiversity hotspot. Sci. Rep. 7:45749. doi: 10.1038/srep45749
Mazoch, V., Mikula, O., Bryja, J., Konvičková, H., Russo, I.-R., Verheyen, E., et al. (2018). Phylogeography of a widespread sub-Saharan murid rodent Aethomys chrysophilus: the role of geographic barriers and paleoclimate in the Zambezian bioregion. Mammalia 82, 373–387. doi: 10.1515/mammalia-2017-0001
Menegon, M., Loader, S. P., Davenport, T. R., Howell, K. M., Tilbury, C. R., and Machago, S. (2015). A new species of chameleon (Sauria: Chamaeleonidae: Kinyongia) highlights the biological affinities between the Southern Highlands and Eastern Arc Mountains of Tanzania. Acta Herpetol. 10, 111–120. doi: 10.13128/Acta_Herpetol-17171
Menegon, M., Loader, S. P., Marsden, S. J., Branch, W. R., Davenport, T. R. B., and Ursenbacher, S. (2014). The genus Atheris (Serpentes: Viperidae) in East Africa: phylogeny and the role of rifting and climate in shaping the current pattern of species diversity. Mol. Phylogenet. Evol. 79, 12–22. doi: 10.1016/j.ympev.2014.06.007
Mikula, O., Šumbera, R., Aghová, T., Mbau, J. S., Katakweba, A. S., Sabuni, C. A., et al. (2016). Evolutionary history and species diversity of African pouched mice (Rodentia: Nesomyidae: Saccostomus). Zool. Scr. 45, 595–617. doi: 10.1111/zsc.12179
Mittermeier, R., Gil, P. R., Hoffmann, M., Pilgrim, J., Brooks, T., Mittermeier, C. G., et al. (2004). Hotspots Revisited. Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions. Mexico: Cemex.
Mittermeier, R. A., Turner, W. R., Larsen, F. W., Brooks, T. M., and Gascon, C. (2011). “Global biodiversity conservation: the critical role of hotspots,” in Biodiversity Hotspots: Distribution and Protection of Conservation Priority Areas, eds F. E. Zachos and J. C. Habel (Berlin: Springer), 3–22. doi: 10.1007/978-3-642-20992-5.
Mizerovská, D., Nicolas, V., Demos, T. C., Akaibe, D., Colyn, M., Denys, C., et al. (2019). Genetic variation of the most abundant forest-dwelling rodents in Central Africa (Praomys jacksoni complex): Evidence for Pleistocene refugia in both montane and lowland forests. J. Biogeogr. 46, 1466–1478. doi: 10.1111/JBI.13604
Nicolas, V., Fabre, P. H., Bryja, J., Denys, C., Verheyen, E., Missoup, A. D., et al. (2020). The phylogeny of the African wood mice (Muridae, Hylomyscus) based on complete mitochondrial genomes and five nuclear genes reveals their evolutionary history and undescribed diversity. Mol. Phylogenet. Evol. 144:106703. doi: 10.1016/J.YMPEV.2019.106703
Nicolas, V., Mikula, O., Lavrenchenko, L. A., Šumbera, R., Bartáková, V., Bryjová, A., et al. (2021). Phylogenomics of African radiation of Praomyini (Muridae: Murinae) rodents: first fully resolved phylogeny, evolutionary history and delimitation of extant genera. Mol. Phylogenet. Evol. 163:107263. doi: 10.1016/j.ympev.2021.107263
Onditi, K. O., Demos, T. C., Kerbis Peterhans, J., Chen, Z.-Z., Bryja, J., Lavrenchenko, L. A., et al. (2021). Historical biogeography, systematics, and integrative taxonomy of the non-Ethiopian speckled pelage brush-furred rats (Lophuromys flavopunctatus group). BMC Ecol. Evol. 21:89. doi: 10.1186/s12862-021-01813-w
Petružela, J., Šumbera, R., Aghová, T., Bryjová, A., Katakweba, A. S., Sabuni, C. A., et al. (2018). Spiny mice of the Zambezian bioregion-phylogeny, biogeography and ecological differentiation within the Acomys spinosissimus complex. Mamm. Biol. 91, 79–90. doi: 10.1016/j.mambio.2018.03.012
Pozzi, L., Penna, A., Bearder, S. K., Karlsson, J., Perkin, A., and Disotell, T. R. (2020). Cryptic diversity and species boundaries within the Paragalago zanzibaricus species complex. Mol. Phylogenet. Evol. 150:106887. doi: 10.1016/j.ympev.2020.106887
Rovero, F., Menegon, M., Fjeldså, J., Collett, L., Doggart, N., Leonard, C., et al. (2014). Targeted vertebrate surveys enhance the faunal importance and improve explanatory models within the Eastern Arc Mountains of Kenya and Tanzania. Divers. Distrib. 20, 1438–1449. doi: 10.1111/ddi.12246
Roy, M. S., Arctander, P., and Fjeldså, J. (1998). Speciation and taxonomy of montane greenbuls of the genus Andropadus (Aves: Pycnonotidae). Steenstrupia 24, 51–66.
Sabuni, C., Aghová, T., Bryjová, A., Šumbera, R., and Bryja, J. (2018). Biogeographic implications of small mammals from Northern Highlands in Tanzania with first data from the volcanic Mount Kitumbeine. Mammalia 82, 360–372. doi: 10.1515/mammalia-2017-0069
Sepulchre, P., Ramstein, G., Fluteau, F., Schuster, M., Tiercelin, J.-J., and Brunet, M. (2006). Tectonic uplift and Eastern Africa aridification. Science 313, 1419–1423. doi: 10.1126/science.1129158
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Stanley, W. T., and Esselstyn, J. A. (2010). Biogeography and diversity among montane populations of mouse shrew (Soricidae: Myosorex) in Tanzania. Biol. J. Linn. Soc. 100, 669–680. doi: 10.1111/j.1095-8312.2010.01448.x
Stanley, W. T., Hutterer, R., Giarla, T. C., and Esselstyn, J. A. (2015). Phylogeny, phylogeography and geographical variation in the Crocidura monax (Soricidae) species complex from the montane islands of Tanzania, with descriptions of three new species. Zool. J. Linn. Soc. 174, 185–215. doi: 10.1111/ZOJ.12230
Taylor, P. J., Maree, S., Van Sandwyk, J., Kerbis Peterhans, J. C., Stanley, W. T., Verheyen, E., et al. (2009). Speciation mirrors geomorphology and palaeoclimatic history in African laminate-toothed rats (Muridae: Otomyini) of the Otomys denti and Otomys lacustris species-complexes in the ‘Montane Circle’ of East Africa. Biol. J. Linn. Soc. 96, 913–941. doi: 10.1111/j.1095-8312.2008.01153.x
Trauth, M. H., Maslin, M. A., Deino, A. L., Strecker, M. R., Bergner, A. G. N., and Dühnforth, M. (2007). High- and low-latitude forcing of Plio-Pleistocene East African climate and human evolution. J. Hum. Evol. 53, 475–486. doi: 10.1016/j.jhevol.2006.12.009
Uhrová, M., Mikula, O., Bennett, N. C., Van Daele, P., Piálek, L., Bryja, J., et al. (2021). Species limits and phylogeographic structure in two genera of solitary African mole-rats Georychus and Heliophobius. Mol. Phylogenet. Evol. 167:107337. doi: 10.1016/j.ympev.2021.107337
Van de Perre, F., Adriaensen, F., Terryn, L., Pauwels, O., Leirs, H., Gilissen, E., et al. (2019). African Mammalia. Available online at: http://projects.biodiversity.be/africanmammalia (accessed July 7, 2021).
Verheyen, W. N., Hulselmans, J. L. J., Dierckx, T., Colyn, M., Leirs, H., and Verheyen, E. (2003). A craniometric and genetic approach to the systematics of the genus Dasymys Peters, 1875, selection of a neotype and description of three new taxa (Rodentia, Muridae, Africa). Bull. Koninklijkk Belg. Inst. Nat. Wet. 73, 27–71.
Voelker, G., Huntley, J. W., Bryja, J., Denys, C., Šumbera, R., Demos, T. C., et al. (2021). Molecular systematics and biogeographic history of the African climbing-mouse complex (Dendromus). Mol. Phylogenet. Evol. 161:107166. doi: 10.1016/j.ympev.2021.107166
White, F. (1983). The Vegetation of Africa: A Descriptive Memoir to Accompany the UNESCO/AETFAT/UNSO Vegetation Map of Africa. Paris: UNESCO, 1–356.
Keywords: comparative phylogeography, rodents, shrews, Eastern Arc Mountains, Southern Rift Mountains, Eastern Afromontane Biodiversity Hotspot, savannah
Citation: Cuypers LN, Sabuni C, Šumbera R, Aghová T, Lišková E, Leirs H, Baird SJE, Goüy de Bellocq J and Bryja J (2022) Biogeographical Importance of the Livingstone Mountains in Southern Tanzania: Comparative Genetic Structure of Small Non-volant Mammals. Front. Ecol. Evol. 9:742851. doi: 10.3389/fevo.2021.742851
Received: 16 July 2021; Accepted: 07 December 2021;
Published: 18 January 2022.
Edited by:
Jon Fjeldså, University of Copenhagen, DenmarkReviewed by:
Adriaan Engelbrecht, University of the Western Cape, South AfricaCássia Bitencourt, Royal Botanic Gardens, Kew, United Kingdom
Copyright © 2022 Cuypers, Sabuni, Šumbera, Aghová, Lišková, Leirs, Baird, Goüy de Bellocq and Bryja. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Josef Bryja, bryja@ivb.cz