
Abstract
In their recent article in Alpine Botany (133:63-67, 2023a), Körner et al. revisit the outcome of an interesting experiment from 2009 (Lenz et al., Plant Ecolog Divers 6:365–375, 2013). Although I appreciate the new focus on cell wall lignification, I disagree with their main conclusion. Rather than questioning the role cold temperatures play in cell wall lignification, the authors provide experimental evidence for a thermal threshold under which the secondary cell walls of mountain pines (Pinus uncinata) at the upper treeline in the Swiss Alps exhibit a reduced lignin content.
Similar content being viewed by others


The thermally induced and conceptually defined potential cold distribution limit of upright plant growth fascinates scholars since at least two centuries (von Humboldt and Bonpland 1807; Troll 1973; Körner 2012). While the highest alpine treelines reach almost 5000 m asl in eastern Tibet and the Bolivian Andes (Hoch and Körner 2005; Miehe et al. 2007), the northernmost arctic treeline approaches sea level at around 73° North on the Russian Taimyr Peninsula in central Siberia (Naurzbaev et al. 2002). Although reasonably well explained by a universal cold temperature isotherm, the mechanistic controls on the global treeline position are still not fully understood (see synthesis Figure 2 in a recent perspective by Körner and Hoch 2023). This lack of understanding has motivated a recent debate about physiological and biochemical factors that may, or may not, contribute to the thermal limit of upright plant growth (Büntgen et al. 2023; Körner et al. 2023b).
The hypothesis of a possible inability of plants to lignify their secondary cell walls under too cold temperatures has been introduced as an additional rather than an alternative concept for better understanding the global treeline position (Crivellaro and Büntgen 2020; Crivellaro et al. 2022). This large-scale biogeographic theory has been corroborated by a simple time-for-space surrogate, the formation of so-called Blue Rings during summers with unusually cold temperatures that often follow large volcanic eruptions (Piermattei et al. 2020; Büntgen et al. 2022a). ‘Blue Rings’ are wood anatomical anomalies characterised by a lack of cell wall lignification across entire (very rare) or partial (more frequent) tree rings.
With just a few local studies and yet not enough time-for-space surrogates (Fig. 1), we are still far away from having a comprehensive picture of the biotic and abiotic drivers of ‘Blue Ring’ formation. However, and contrary to the motivation by Körner et al. (2023a), there is no evidence that constant cooling of 3 K over much of the growing season at the upper treeline in the Swiss Alps would limit cell wall lignification. In fact, any long-term treatment should be meaningless because it does not matter if very warm summers and extremely cold winters are chronically cooler or warmer by 3 K. What matters is the frequency, intensity, and duration of critical cold spells during the growing season. Moreover, the in situ tissue cooling of the 2009 experiment was conducted at the Stillberg research site between 2140 and 2170 m asl in the eastern Swiss Alps near Davos (46°46’ N, 9°52’ E; Lenz et al. 2013), which is approximately 250–300 m below the potential treeline (Büntgen et al. 2022b). Since the realised and potential treeline positions are almost never in equilibrium, most treeline studies of the past decades describe too warm in situ conditions as the ecotone (realised treeline) is lacking the rate of recent anthropogenic warming (potential treeline).

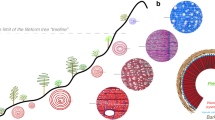
a Since the first wood anatomical descriptions of a lack of cell wall lignification in herbs and dwarf shrubs from coastal East Greenland (L1; Büntgen et al. 2015) and the formation of ‘Blue Rings’ (BRs) in Pinus nigra from Italy (H1; Piermattei et al. 2015), further studies revealed non-lignified stems of Ladakiella klimesii from Eastern Ladakh (L2; Dolezal et al. 2016), BRs in Pinus sylvestris from Romania (H2; Semeniuc et al. 2016), a lack of cell wall lignification in herbs and dwarf shrubs from coastal East Greenland (L3; Büntgen et al. 2018), BRs in Pinus contorta from Western Canada and the USA (H3; Montwé et al. 2018), BRs in Pinus nigra from Italy (H4; Crivellaro et al. 2018), BRs in Larix decidua from the Russian Altai (H5; Büntgen et al. 2020), BRs in Pinus sylvestris from Latvia (H6; Matisons et al. 2020), BRs in Pinus uncinata from the Spanish central Pyrenees (H7; Piermattei et al. 2020), BRs in Pinus longaeva from Nevada, USA (H8; Tardif et al. 2020), BRs in Pinus sylvestris from Finland (H9; Björklund et al. 2021), BRs in 12 species from 14 sites in the Northern Hemisphere extra-tropics (H10; Büntgen et al. 2022a); BRs in Pinus sylvestris from Poland (H11; Matulewski et al. 2022), a tendency towards less lignified cell walls in small plants at cold sites (G1; Crivellaro et al. 2022), BRs in Pinus sylvestris from Estonia (H12; Greaves et al. 2023), and an experimentally forced lack of cell wall lignification in Pinus uncinata from the eastern Swiss Alps (E1; Körner et al. 2023a). b Schematic overview of local, historical, global, and experimental evidence for temperature controls on biotic and abiotic factors and processes relevant for plant cell wall lignification obtained from the interrelated fields of botany, paleoclimatology, biogeography, and plant physiology
Importantly, Körner et al. (2023a) provide the first experimental in situ treeline evidence for a temperature-induced disruption of the biochemical process of cell wall lignification in the stems of upright growing conifer trees. Although the authors write that their experiment ‘does not suggest that lignification is low-temperature limited in treeline trees’, their study shows the opposite and confirms the importance of ephemeral cold spells during the growing season. In contrast to their interpretation and conclusion, Körner et al. (2023a) corroborate previously published local, global, and historical indication for a low-temperature threshold under which cell wall lignification ceases (see Fig. 1 for a topical review).
Moreover, Körner et al. (2023a) raise at least five scientific issues that merit further discussion (1–5): (1) a constant 3 K cooling with computer-controlled Peltier collars from early June until the end of September is arguably a too short seasonal window to represent an elevational upward shift of 500–600 m. A constant summer cooling also neglects the possibility of an increasing range of temperature extremes that may occur on daily to annual scales at higher elevations where wind speed and cloud cover change as well. (2) So-called Frost Rings are characterised by cell deformation and/or collapse but not necessarily by a lack of cell wall lignification. (3) It has never been speculated that alpine herbs are small because they cannot produce lignin, but it has been observed that small plants above the upper or northern treelines may grow and survive even with less lignified cell walls. (4) Neither has a gradual decline in latewood lignification been reported nor should it be related to wood density measurements that are traditionally based on the two-dimensional ratio between tracheid cell wall thickness and transverse cell lumen area (Björklund et al. 2019). (5) ‘Blue Rings’ have not been reported to follow the exceptionally warm summer of 1976 in western and central Europe. However, a remarkably high occurrence of ‘Blue Rings’ in Estonian Scots pines in 1976 was likely triggered by exceptionally cold September and October temperatures over eastern Europe (Greaves et al. 2023), where the summer months were also slightly cooler than the 1961–1990 average.
Despite the above inconsistencies, I agree with Körner et al. (2023a) that species-specific cold distribution limits of trees, shrubs, and herbs are likely not carbon (photosynthesis)-limited, and that it is still poorly understood what permits perennial plants from growing and ageing under critically low (mean and/or extreme)-temperature conditions. I also concur that smaller stature plants above the treeline can experience warmer microclimatic conditions during favourable weather conditions (Körner et al. 2023b). This observation, however, does not reject the hypothesis that a thermally induced lack of cell wall lignification can play a role in the global treeline position (Crivellaro et al. 2022; Büntgen et al. 2023). Notwithstanding recent advancements at the interface of dendrochronology and wood anatomy, it remains unclear what the double staining of anatomical cross sections with Astra Blue and Safranin actually reveals, how cold it needs to be for how long to trigger entire or partial ‘Blue Rings’, and if different tree, shrub and herb species exhibit different temperature thresholds under which the secondary cell walls of their stems lack the necessary degree of lignin that is needed to grow upright and conduct water over large distance.
Future research may therefore use controlled laboratory or open field experiments to explore the thermal limits of biosynthesis within different cell wall components to better understand xylogenesis and lignin formation. Such studies should include plant stem anatomical analyses of different cell functional types to quantify the climatic drivers of ‘Blue Rings’ within and between species and lifeforms. Mutants of Arabidopsis thaliana (L.) Heynh and other model plants with and without the expression of lignin-relevant key enzymes could be used to investigate the biochemical basis for a putative temperature threshold for lignin formation and to investigate the ability of lignin to penetrate cell walls. Genetic control of different types of lignin in plant cell walls is expected to provide new insights into the potential and limitations of Safranin and Astra Blue double staining. Confocal fluorescence imaging may possibly help to quantify the degree of cell wall lignification in ‘Blue Rings’, which would offer more nuanced climatic information beyond a simple binary classification. Moreover, combined analyses of dendrochronological and wood anatomical data from different species should be conducted along undisturbed elevational gradients in various high-mountain regions.
In synthesis, I consider the ongoing debate a refreshing impetus for critical thinking and scientific exchange within and beyond biologists, ecologists, and geographers interested in, and fascinated by, the global treeline phenomenon.
Data availability
No unpublished data have been used in this work.
References
Björklund J, von Arx G, Nievergelt D, Wilson R, Van den Bulcke J, Günther B, Loader N, Rydval M, Fonti P, Scharnweber T, Andreu-Hayles L, Büntgen U, D’Arrigo R, Davi N, De Mil T, Esper J, Gärtner H, Geary J, Gunnarson B, Hartl C, Hevia A, Huiming S, Janecka K, Kaczka R, Kirdyanov AV, Kochbeck M, Liu Y, Meko M, Mundo I, Nicolussi K, Oelkers R, Pichler T, Sánchez Salguero R, Schneider L, Schweingruber F, Timonen M, Trouet V, Van Acker J, Verstege A, Villalba R, Wilmking M, Frank D (2019) Scientific merits and analytical challenges of tree-ring densitometry. Rev Geophys 57:1224–1264
Björklund J, Fonti M, Fonti P, Van den Bulcke J, von Arx G (2021) Cell wall dimensions reign supreme: cell wall composition is irrelevant for the temperature signal of latewood density/blue intensity in Scots pine. Dendrochronologia 65:125785
Büntgen U, Hellmann L, Tegel W, Normand S, Myer-Smith I, Kirdyanov A, Nievergelt D, Schweingruber FH (2015) Temperature-induced recruitment pulses of Arctic dwarf shrub communities. J Ecol 103:489–501
Büntgen U, Bolze N, Hellmann L, Sittler B, Frauenberger B, Piermattei A, Kirdyanov A, Schweingruber FH, Ludemann T, Krusic PJ (2018) Long-term recruitment dynamics of Arctic dwarf shrub communities in coastal east Greenland. Dendrochronologia 50:70–80
Büntgen U, Arseneault D, Boucher É, Churakova Sidorova OV, Gennaretti F, Crivellaro A, Hughes MK, Kirdyanov A, Klippel L, Krusic PJ, Linderholm HW, Ljungqvist FC, Ludescher J, McCormick M, Myglan VS, Nicolussi K, Piermattei A, Oppenheimer C, Reinig F, Sigl M, Vaganov EA, Esper J (2020) Prominent role of volcanism in Common Era climate variability and human history. Dendrochronologia 64:125757
Büntgen U, Crivellaro A, Arseneault D, Baillie M, Barclay D, Bernabei M, Bontadi J, Boswijk G, Brown D, Christie DA, Churakova Sidorova OV, Cook E, D’Arrigo R, Davi N, Esper J, Fonti P, Greaves C, Hantemirov RM, Hughes MK, Kirdyanov AV, Krusic PJ, Le Quesne C, Ljungqvist FC, Myglan VS, Nicolussi K, Oppenheimer C, Palmer J, Qin C, Reinig F, Salzer M, Stoffel M, Torbenson M, Villalba R, Wiesenberg N, Wiles G, Yang B, Piermattei A (2022a) Global wood anatomical network reveals repeated summer cooling at the onset of the late antique little ice age. Sci Bull 67:2336–2344
Büntgen U, Piermattei A, Crivellaro A, Reinig F, Krusic PJ, Trnka M, Torbenson M, Esper J (2022b) Common Era treeline fluctuations and their implications for climate reconstructions. Global Planet Change 219:103979
Büntgen U, Piermattei A, Dolezal J, Dupree P, Crivellaro A (2023) Reply to: Biogeographic implications of plant stature and microclimate in cold regions. Commun Biol 6:662
Crivellaro A, Büntgen U (2020) New evidence of thermally-constraint plant cell wall lignification. Trends Plant Sci 24:322–324
Crivellaro A, Reverenna M, Ruffinatto F, Urbinati C, Piermattei A (2018) The anatomy of “blue ring” in the wood of Pinus nigra. Les Wood 67:21–28
Crivellaro A, Piermattei A, Dolezal J, Dupree P, Büntgen U (2022) Biogeographic implication of temperature-induced plant cell wall lignification. Commun Biol 5:767
Dolezal J, Dvorsky M, Kopecky M, Liancourt P, Hiiesalu I, Macek M, Altman J, Chlumska Z, Rehakova K, Capkova K, Borovec J, Mudrak O, Wild J, Schweingruber F (2016) Vegetation dynamics at the upper elevational limit of vascular plants in Himalaya. Sci Rep 6:24881
Greaves C, Crivellaro A, Piermattei A, Krusic PJ, Oppenheimer C, Potapov A, Hordo M, Metslaid S, Kask R, Kangur A, Büntgen U (2023) Remarkably high blue ring occurrence in Estonian Scots pines in 1976 reveals wood anatomical evidence of extreme autumnal cooling. Trees Struct Funct 37:511–522
Hoch G, Körner C (2005) Growth, demography and carbon relations of Polylepis trees at the world’s highest treeline. Funct Ecol 19:941–951
Körner C (2012) Alpine treelines. Springer, Cham
Körner C, Hoch G (2023) Not every high-latitude or high-elevation forest edge is a treeline. J Biogeogr 50:838–845
Körner C, Lenz A, Hoch G (2023a) Chronic in situ tissue cooling does not reduce lignification at the Swiss treeline but enhances the risk of ‘blue’ frost rings. Alp Bot 133:63–67
Körner C, Fajardo A, Hiltbrunner E (2023b) Biogeographic implications of plant stature and microclimate in cold regions. Commun Biol 6:663
Lenz A, Hoch G, Körner C (2013) Early season temperature controls cambial activity and total tree ring width at the alpine treeline. Plant Ecolog Divers 6:365–375
Matisons R, Gärtner H, Elferts D, Kārkliņa A, Adamovičs A, Jansons Ā (2020) Occurrence of ‘blue’ and ‘frost’ rings reveal frost sensitivity of eastern Baltic provenances of Scots pine. For Ecol Manage 457:117729
Matulewski P, Buchwal A, Gärtner H, Jagodziński A, Čufar K (2022) Altered growth with blue rings: comparison of radial growth and wood anatomy between trampled and non-trampled Scots pine roots. Dendrochronologia 72:125922
Miehe G, Miehe S, Vogel J, Co S, Duo L (2007) Highest treeline in the Northern Hemisphere found in Southern Tibet. Mt Res Dev 27:169–173
Montwé D, Isaac-Renton M, Hamann A, Spiecker H (2018) Cold adaptation recorded in tree rings highlights risks associated with climate change and assisted migration. Nat Commun 9:1–7
Naurzbaev MM, Vaganov EA, Sidorova OV, Schweingruber FH (2002) Summer temperatures in eastern Taimyr inferred from a 2427-year late-Holocene tree-ring chronology and earlier floating series. Holocene 12:727–736
Piermattei A, Crivellaro A, Carrer M, Urbinati C (2015) The “blue ring”: anatomy and formation hypothesis of a new tree-ring anomaly in conifers. Trees Struct Funct 29:613–620
Piermattei A, Crivellaro A, Krusic PJ, Esper J, Vítek P, Oppenheimer C, Felhofer M, Gierlinger N, Reinig F, Urban O, Verstege A, Lobo H, Büntgen U (2020) A millennium-long ‘Blue-Ring’ chronology from the Spanish Pyrenees reveals sever ephemeral summer cooling after volcanic eruptions. Environ Res Lett 15:124016
Semeniuc A, Sidor C, Popa I (2016) Scots pine tree ring structure modifications and relation with climate. Eurasian J Forest Sci 4:1–7
Tardif J, Salzer M, Conciatori F, Bunn A, Hughes M (2020) Formation, structure and climatic significance of blue rings and frost rings in high elevation bristlecone pine (Pinus longaeva D.K. Bailey). Quat Sci Rev 244:106516
Troll C (1973) The upper timberlines in different climatic zones. Arct Alp Res 5:A3–A18
von Humboldt A, Bonpland A (1807) Ideen zu einer Geographie der Pflanzen nebst einem Naturgemälde der Tropenländer: auf Beobachtungen und Messungen gegründet, welche vom 10ten Grade nördlicher bis zum 10ten Grade südlicher Breite, in den Jahren 1799, 1800, 1801, 1802 und 1803 angestellt worden sind. Tübingen, Bey F.G. Cotta
Acknowledgements
I received generous supported from the Czech Science Foundation grant HYDRO8 (23-08049S) and the ERC Advanced grant MONOSTAR (AdG 882727). Alma Piermattei, Alan Crivellaro, Jiri Dolezal, and many more colleagues contributed to relevant scientific discussion.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
I declare no conflict of interest besides scientific debate, and I confirm that my research did not involve human participants and/or animals.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Büntgen, U. Experimental evidence for a thermal limitation of plant cell wall lignification at the alpine treeline. Alp Botany 133, 179–182 (2023). https://doi.org/10.1007/s00035-023-00299-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00035-023-00299-0