
Silver Nanoparticles Alter Microtubule Arrangement, Dynamics and Stress Phytohormone Levels

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Direct Visualization of AgNPs Clusters and Specific Changes in GFP-TUA6 Arabidopsis
2.2. Hormonal Profiles in Whole Arabidopsis Plants after AgNPs and Ag+ Treatment
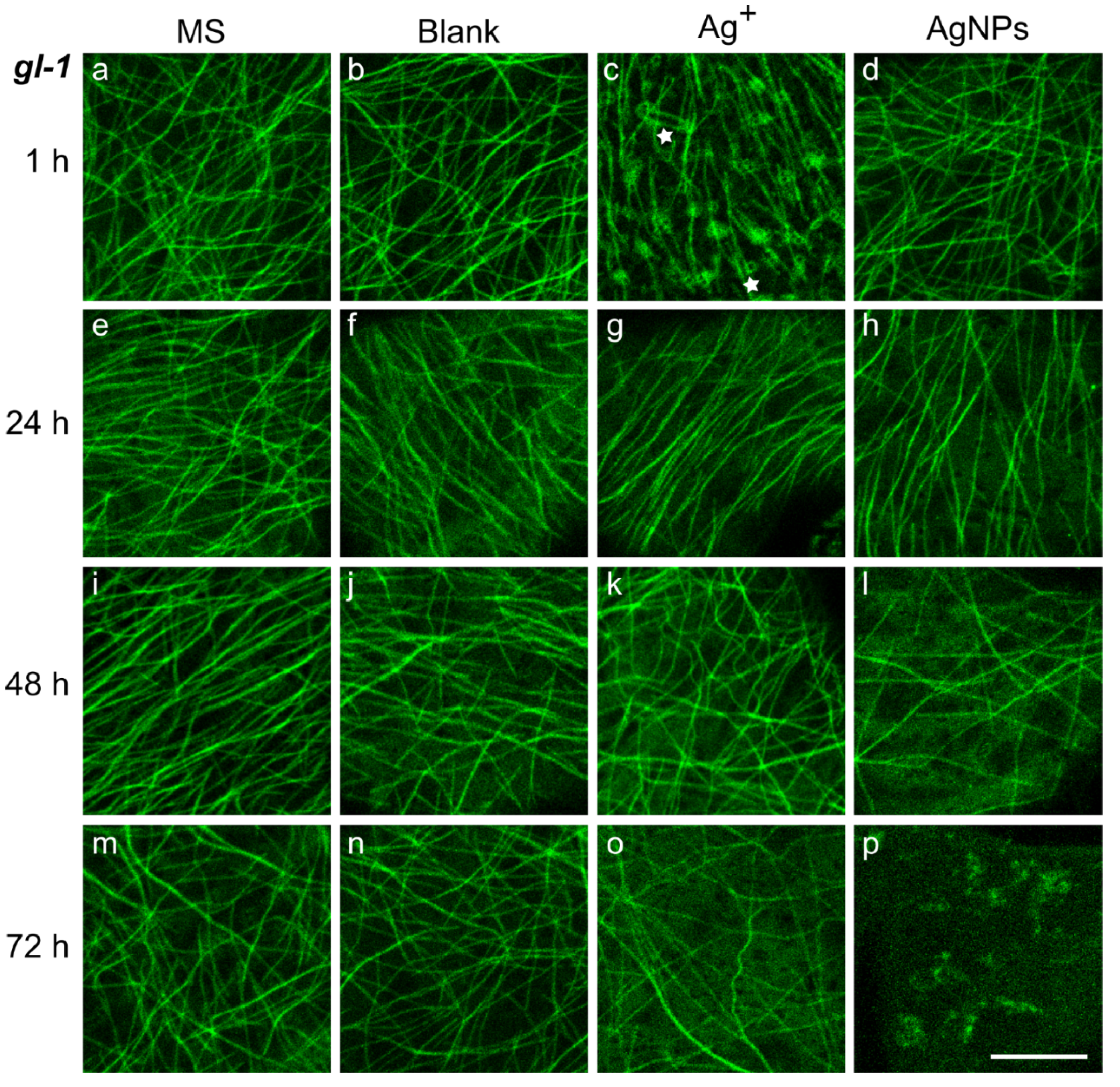
2.3. Changes in Microtubule Pattern after Treatment with Ag+ or AgNPs
2.4. The Number of MTs Appearing De Novo in Cotyledon Cells
2.5. AgNPs Differently Decelerate Microtubular Dynamics in WT and Mutant Plants
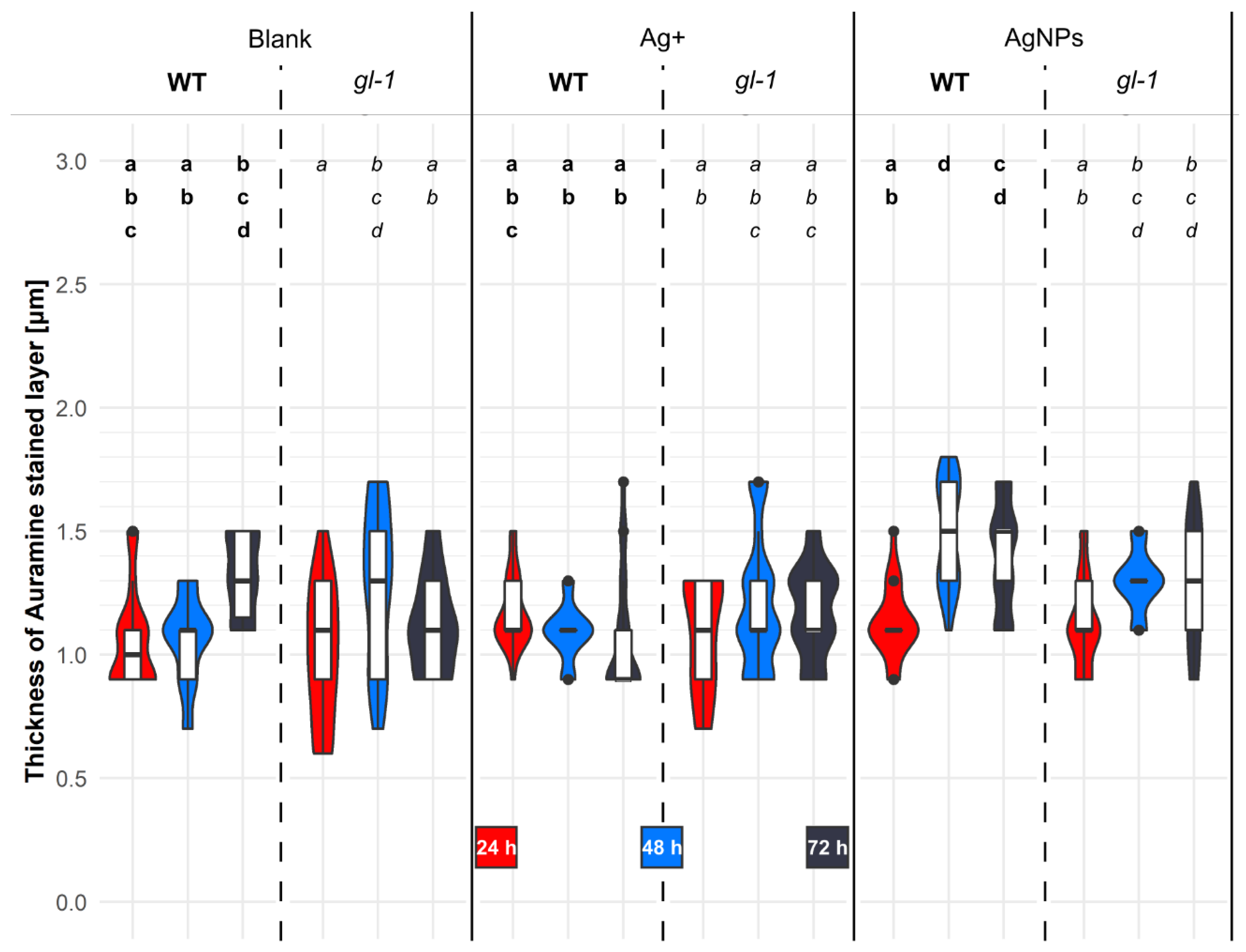
2.6. The Thickness of the Auramine O-Stained Layer in Cotyledons after Treatment with AgNPs
3. Discussion
3.1. Nonstressed Plant Lines Differ in Microtubular Dynamics and Hormonal Profiles
3.2. Ag+ Treatment Led to the Organelle Appearing in the Cortical Cell Layer and a Quick Rearrangement of the MTs to Parallel Arrangement
3.3. AgNPs Gradually Destroyed the Microtubule Dynamics and Pattern, and Substantially Altered the Hormonal Profile and Layer Thickness above the Plasma Membrane
4. Materials and Methods
4.1. Synthesis and Characterization of Nanoparticles
4.2. Preparation of Silver Nanoparticles for Experiments
4.3. Plant Material
4.3.1. Plant Cultivation for Microtubular Dynamics Experiments
4.3.2. Modified Cultivation for Auramine O-Stained Layer Observation and Phytohormone Analysis
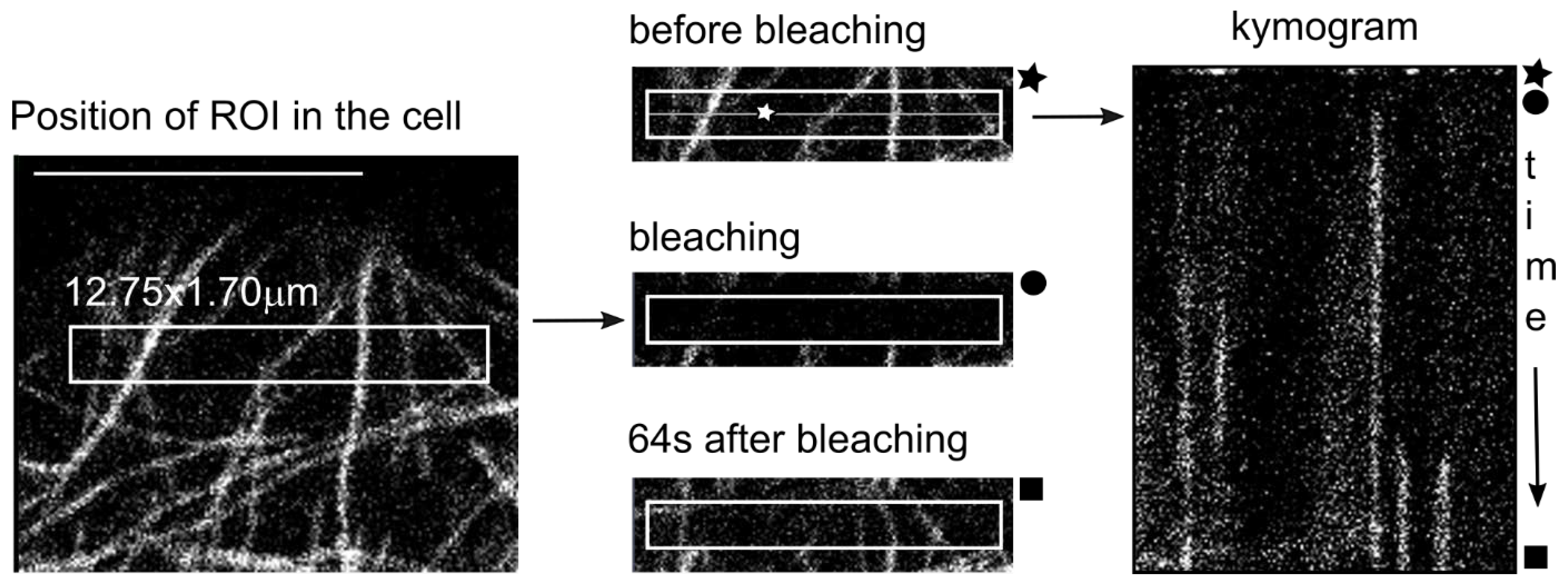
4.4. Modified FRAP Method
4.5. Technical Notes on Confocal Microscopy—Imaging of MT
4.6. Microscopy and Image Processing for Auamine O Stained Layers
4.7. Analysis of Phytohormones
4.8. Statistical Procedures and Plot Design
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falsini, S.; Clemente, I.; Papini, A.; Tani, C.; Schiff, S.; Salvatici, M.C.; Petruccelli, R.; Benelli, C.; Giordano, C.; Gonnelli, C.; et al. When Sustainable Nanochemistry Meets Agriculture: Lignin Nanocapsules for Bioactive Compound Delivery to Plantlets. ACS Sustain. Chem. Eng. 2019, 7, 19935–19942. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.H.; Al-whaibi, M.H.; Mohammad, F. Nanotechnology and Plant Sciences; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 9783319145013. [Google Scholar]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of Metal and Metal Oxide Nanoparticles on Plant: A Critical Review. Front. Chem. 2017, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, A.; Chen, Z. Impacts of Silver Nanoparticles on Plants: A Focus on the Phytotoxicity and Underlying Mechanism. Int. J. Mol. Sci. 2019, 20, 1003. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.; Venkatachalam, P.; Sahi, S.; Sharma, N. Reprint of: Silver and Titanium Dioxide Nanoparticle Toxicity in Plants: A Review of Current Research. Plant Physiol. Biochem. 2017, 110, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Geisler-Lee, J.; Wang, Q.; Yao, Y.; Zhang, W.; Geisler, M.; Li, K.; Huang, Y.; Chen, Y.; Kolmakov, A.; Ma, X. Phytotoxicity, Accumulation and Transport of Silver Nanoparticles by Arabidopsis Thaliana. Nanotoxicology 2012, 7, 323–337. [Google Scholar] [CrossRef]
- Kaegi, R.; Sinnet, B.; Zuleeg, S.; Hagendorfer, H.; Mueller, E.; Vonbank, R.; Boller, M.; Burkhardt, M. Release of Silver Nanoparticles from Outdoor Facades. Environ. Pollut. 2010, 158, 2900–2905. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Tripathi, A.; Shweta; Singh, S.; Singh, Y.; Vishwakarma, K.; Yadav, G.; Sharma, S.; Singh, V.K.; Mishra, R.K.; et al. Uptake, Accumulation and Toxicity of Silver Nanoparticle in Autotrophic Plants, and Heterotrophic Microbes: A Concentric Review. Front. Microbiol. 2017, 8, 7. [Google Scholar] [CrossRef]
- Nawrath, C.; Schreiber, L.; Franke, R.B.; Geldner, N.; Reina-Pinto, J.J.; Kunst, L. Apoplastic Diffusion Barriers in Arabidopsis. Arab. Book 2013, 11, e0167. [Google Scholar] [CrossRef] [Green Version]
- Bao, D.; Oh, Z.G.; Chen, Z. Characterization of Silver Nanoparticles Internalized by Arabidopsis Plants Using Single Particle ICP-MS Analysis. Front. Plant Sci. 2016, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Larue, C.; Castillo-Michel, H.; Sobanska, S.; Cécillon, L.; Bureau, S.; Barthès, V.; Ouerdane, L.; Carrière, M.; Sarret, G. Foliar Exposure of the Crop Lactuca Sativa to Silver Nanoparticles: Evidence for Internalization and Changes in Ag Speciation. J. Hazard. Mater. 2014, 264, 98–106. [Google Scholar] [CrossRef]
- Schwab, F.; Zhai, G.; Kern, M.; Turner, A.; Schnoor, J.L.; Wiesner, M.R. Barriers, Pathways and Processes for Uptake, Translocation and Accumulation of Nanomaterials in Plants—Critical Review. Nanotoxicology 2016, 10, 257–278. [Google Scholar] [CrossRef]
- Berhin, A.; de Belllis, D.; Franke, R.B.; Andrade Buono, R.; Nowack, M.; Nawrath, C. The Root Cap Cuticle: A Cell Wall Structure for Seedling Establishment and Lateral Root Formation. Cell 2019, 176, 1367–1378.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegebarth, D.; Buschhaus, C.; Wu, M.; Bird, D.; Jetter, R. The Composition of Surface Wax on Trichomes of Arabidopsis Thaliana Differs from Wax on Other Epidermal Cells. Plant J. 2016, 88, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Hegebarth, D.; Jetter, R. Cuticular Waxes of Arabidopsis Thaliana Shoots: Cell-Type-Specific Composition and Biosynthesis. Plants 2017, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Yu, K.; Navarre, D.; Seebold, K.; Kachroo, A.; Kachroo, P. The Glabra1 Mutation Affects Cuticle Formation and Plant Responses to Microbes. Plant Physiol. 2010, 154, 833–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattanaik, S.; Patra, B.; Singh, S.K.; Yuan, L. An Overview of the Gene Regulatory Network Controlling Trichome Development in the Model Plant, Arabidopsis. Front. Plant Sci. 2014, 5, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perazza, D.; Herzog, M.; Hülskamp, M.; Brown, S.; Dorne, A.-M.; Bonneville, J.-M. Trichome Cell Growth in Arabidopsis Thaliana Can Be Derepressed by Mutations in at Least Five Genes. Genetics 1999, 152, 461–476. [Google Scholar] [CrossRef]
- Marks, M.D. Molecular Genetic Analysis of Trichome Development in Arabidopsis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 137–163. [Google Scholar] [CrossRef]
- Liang, S.; Yang, X.; Deng, M.; Zhao, J.; Shao, J.; Qi, Y.; Liu, X.; Yu, F.; An, L. A New Allele of the SPIKE1 Locus Reveals Distinct Regulation of Trichome and Pavement Cell Development and Plant Growth. Front. Plant Sci. 2019, 10, 16. [Google Scholar] [CrossRef]
- Kim, G.-T.; Shoda, K.; Tsuge, T.; Cho, K.-H.; Uchimiya, H.; Yokoyama, R.; Nishitani, K.; Tsukaya, H. The ANGUSTIFOLIA Gene of Arabidopsis, a Plant CtBP Gene, Regulates Leaf-Cell Expansion, the Arrangement of Cortical Microtubules in Leaf Cells and Expression of a Gene Involved in Cell-Wall Formation. EMBO J. 2002, 21, 1267–1279. [Google Scholar] [CrossRef] [Green Version]
- Lü, B.; Wang, J.; Zhang, Y.; Wang, H.; Liang, J.; Zhang, J. AT14A Mediates the Cell Wall-Plasma Membrane-Cytoskeleton Continuum in Arabidopsis Thaliana Cells. J. Exp. Bot. 2012, 63, 4061–4069. [Google Scholar] [CrossRef] [PubMed]
- Milewska-Hendel, A.; Zubko, M.; Stróż, D.; Kurczyńska, E. Effect of Nanoparticles Surface Charge on the Arabidopsis Thaliana (L.) Roots Development and Their Movement into the Root Cells and Protoplasts. Int. J. Mol. Sci. 2019, 20, 1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Geiser-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between Engineered Nanoparticles (ENPs) and Plants: Phytotoxicity, Uptake and Accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Bücker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between Plant Hormones and Heavy Metals Responses. Genet. Mol. Biol. 2017, 40, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Ara, T.; Aoki, K.; Suzuki, H.; Shibata, D. Transient Increase in Salicylic Acid and Its Glucose Conjugates after Wounding in Arabidopsis Leaves. Plant Biotechnol. 2010, 27, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Floková, K.; Tarkowská, D.; Miersch, O.; Strnad, M.; Wasternack, C.; Novák, O. UHPLC–MS/MS Based Target Profiling of Stress-Induced Phytohormones. Phytochemistry 2014, 105, 147–157. [Google Scholar] [CrossRef]
- Hu, B.; Deng, F.; Chen, G.; Chen, X.; Gao, W.; Long, L.; Xia, J.; Chen, Z.-H. Evolution of Abscisic Acid Signaling for Stress Responses to Toxic Metals and Metalloids. Front. Plant Sci. 2020, 11, 909. [Google Scholar] [CrossRef]
- Sanzari, I.; Leone, A.; Ambrosone, A. Nanotechnology in Plant Science: To Make a Long Story Short. Front. Bioeng. Biotechnol. 2019, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Karami Mehrian, S.; de Lima, R. Nanoparticles Cyto and Genotoxicity in Plants: Mechanisms and Abnormalities. Environ. Nanotechnol. Monit. Manag. 2016, 6, 184–193. [Google Scholar] [CrossRef]
- Lv, J.; Christie, P.; Zhang, S. Uptake, Translocation, and Transformation of Metal-Based Nanoparticles in Plants: Recent Advances and Methodological Challenges. Environ. Sci. Nano 2019, 6, 41–59. [Google Scholar] [CrossRef]
- Kaveh, R.; Li, Y.S.; Ranjbar, S.; Tehrani, R.; Brueck, C.L.; Van Aken, B. Changes in Arabidopsis Thaliana Gene Expression in Response to Silver Nanoparticles and Silver Ions. Environ. Sci. Technol. 2013, 47, 10637–10644. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.; Eder, M.; Crowell, E.F.; Hampson, J.; Calder, G.; Lloyd, C. Microtubules and CESA Tracks at the Inner Epidermal Wall Align Independently of Those on the Outer Wall of Light-Grown Arabidopsis Hypocotyls. J. Cell Sci. 2011, 124, 1088–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geitmann, A.; Nebenfãhr, A. Navigating the Plant Cell: Intracellular Transport Logistics in the Green Kingdom. Mol. Biol. Cell 2015, 26, 3373–3378. [Google Scholar] [CrossRef] [PubMed]
- Dixit, R.; Cyr, R. The Cortical Microtubule Array: From Dynamics to Organization. Plant Cell 2004, 16, 2546–2552. [Google Scholar] [CrossRef] [Green Version]
- Cai, G. Assembly and Disassembly of Plant Microtubules: Tubulin Modifications and Binding to MAPs. J. Exp. Bot. 2010, 61, 623–626. [Google Scholar] [CrossRef]
- Ma, H.; Liu, M. The Microtubule Cytoskeleton Acts as a Sensor for Stress Response Signaling in Plants. Mol. Biol. Rep. 2019, 46, 5603–5608. [Google Scholar] [CrossRef]
- Nick, P. Microtubules, Signalling and Abiotic Stress. Plant J. 2013, 75, 309–323. [Google Scholar] [CrossRef]
- Hardham, A.R. Microtubules and Biotic Interactions. Plant J. 2013, 75, 278–289. [Google Scholar] [CrossRef]
- Zhou, S.; Chen, Q.; Li, X.; Li, Y. MAP65-1 Is Required for the Depolymerization and Reorganization of Cortical Microtubules in the Response to Salt Stress in Arabidopsis. Plant Sci. 2017, 264, 112–121. [Google Scholar] [CrossRef]
- Thion, L.; Mazars, C.; Nacry, P.; Bouchez, D.; Moreau, M.; Ranjeva, R.; Thuleau, P. Plasma Membrane Depolarization-Activated Calcium Channels, Stimulated by Microtubule-Depolymerizing Drugs in Wild-Type Arabidopsis Thaliana Protoplasts, Display Constitutively Large Activities and a Longer Half-Life in Ton 2 Mutant Cells Affected in the organization of cortical microtubules. Plant J. 1998, 13, 603–610. [Google Scholar] [CrossRef]
- Krasylenko, Y.A.; Yemets, A.I.; Blume, Y.B. Plant Microtubules Reorganization under the Indirect UV-B Exposure and during UV-B-Induced Programmed Cell Death. Plant Signal. Behav. 2013, 8, e24031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacques, E.; Verbelen, J.P.; Vissenberg, K. Mechanical Stress in Arabidopsis Leaves Orients Microtubules in a “continuous” Supracellular Pattern. BMC Plant Biol. 2013, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yang, Q.; Wang, Y.; Wang, L.; Fu, Y.; Wang, X. Brassinosteroids Regulate Pavement Cell Growth by Mediating BIN2-Induced Microtubule Stabilization. J. Exp. Bot. 2018, 69, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Ursache, R.; Andersen, T.G.; Marhavý, P.; Geldner, N. A Protocol for Combining Fluorescent Proteins with Histological Stains for Diverse Cell Wall Components. Plant J. 2018, 93, 399–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhamidi, S.; Shi, L.; Chatterjee, D.; Belisle, J.T.; Crick, D.C.; McNeil, M.R. A Bioanalytical Method to Determine the Cell Wall Composition of Mycobacterium Tuberculosis Grown in Vivo. Anal. Biochem. 2012, 421, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Buda, G.J.; Isaacson, T.; Matas, A.J.; Paolillo, D.J.; Rose, J.K.C. Three-Dimensional Imaging of Plant Cuticle Architecture Using Confocal Scanning Laser Microscopy. Plant J. 2009, 60, 378–385. [Google Scholar] [CrossRef]
- Mayumi, K.; Shibaoka, H. The Cyclic Reorientation of Cortical Microtubules on Walls with a Crossed Polylamellate Structure: Effects of Plant Hormones and an Inhibitor of Protein Kinases on the Progression of the Cycle. Protoplasma 1996, 195, 112–122. [Google Scholar] [CrossRef]
- Marc, J.; Granger, C.L.; Brincat, J.; Fisher, D.D.; Kao, T.H.; McCubbin, A.G.; Cyr, R.J. A GFP-MAP4 Reporter Gene for Visualizing Cortical Microtubule Rearrangements in Living Epidermal Cells. Plant Cell 1998, 10, 1927–1939. [Google Scholar] [CrossRef] [Green Version]
- Sambade, A.; Pratap, A.; Buschmann, H.; Morris, R.J.; Lloyd, C. The Influence of Light on Microtubule Dynamics and Alignment in the Arabidopsis Hypocotyl. Plant Cell 2012, 24, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, D.W.; Shaw, S.L. Microtubule Dynamics and Organization in the Plant Cortical Array. Annu. Rev. Plant Biol. 2006, 57, 859–875. [Google Scholar] [CrossRef] [PubMed]
- Medford, J.I.; Behringer, F.J.; Callos, J.D.; Feldmann, K.A. Normal and Abnormal Development in the Arabidopsis Vegetative Shoot Apex. Plant Cell 1992, 4, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Maes, L.; Inzé, D.; Goossens, A. Functional Specialization of the TRANSPARENT TESTA GLABRA1 Network Allows Differential Hormonal Control of Laminal and Marginal Trichome Initiation in Arabidopsis Rosette Leaves. Plant Physiol. 2008, 148, 1453–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noir, S.; Bömer, M.; Takahashi, N.; Ishida, T.; Tsui, T.L.; Balbi, V.; Shanahan, H.; Sugimoto, K.; Devoto, A. Jasmonate Controls Leaf Growth by Repressing Cell Proliferation and the Onset of Endoreduplication While Maintaining a Potential Stand-by Mode. Plant Physiol. 2013, 161, 1930–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vleesschauwer, D.; Seifi, H.S.; Filipe, O.; Haeck, A.; Huu, S.N.; Demeestere, K.; Höfte, M. The DELLA Protein SLR1 Integrates and Amplifies Salicylic Acid- and Jasmonic Acid-Dependent Innate Immunity in Rice. Plant Physiol. 2016, 170, 1831–1847. [Google Scholar] [CrossRef] [Green Version]
- Denness, L.; McKenna, J.F.; Segonzac, C.; Wormit, A.; Madhou, P.; Bennett, M.; Mansfield, J.; Zipfel, C.; Hamann, T. Cell Wall Damage-Induced Lignin Biosynthesis Is Regulated by a Reactive Oxygen Species- and Jasmonic Acid-Dependent Process in Arabidopsis. Plant Physiol. 2011, 156, 1364–1374. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [Green Version]
- Blume, Y.B.; Krasylenko, Y.A.; Yemets, A.I. The Role of the Plant Cytoskeleton in Phytohormone Signaling under Abiotic and Biotic Stresses. In Mechanism of Plant Hormone Signaling under Stress; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 127–185. [Google Scholar]
- Nakamura, M. Microtubule Nucleating and Severing Enzymes for Modifying Microtubule Array Organization and Cell Morphogenesis in Response to Environmental Cues. New Phytol. 2015, 205, 1022–1027. [Google Scholar] [CrossRef]
- Tulin, A.; McClerklin, S.; Huang, Y.; Dixit, R. Single-Molecule Analysis of the Microtubule Cross-Linking Protein MAP65-1 Reveals a Molecular Mechanism for Contact-Angle-Dependent Microtubule Bundling. Biophys. J. 2012, 102, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Luptovčiak, I.; Komis, G.; Takáč, T.; Ovečka, M.; Šamaj, J. Katanin: A Sword Cutting Microtubules for Cellular, Developmental, and Physiological Purposes. Front. Plant Sci. 2017, 8, 1982. [Google Scholar] [CrossRef] [Green Version]
- McNally, F.J.; Okawa, K.; Iwamatsu, A.; Vale, R.D. Katanin, the Microtubule-Severing ATPase, Is Concentrated at Centrosomes. J. Cell Sci. 1996, 109, 561–567. [Google Scholar] [CrossRef]
- Elliott, A.; Shaw, S.L. Update: Plant Cortical Microtubule Arrays. Plant Physiol. 2018, 176, 94–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, I.; Sassmann, S.; Schmidt, B.; Komis, G. Plasmolysis: Loss of Turgor and Beyond. Plants 2014, 3, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Nakano, R.T.; Yamada, K.; Bednarek, P.; Nishimura, M.; Hara-Nishimura, I. ER Bodies in Plants of the Brassicales Order: Biogenesis and Association with Innate Immunity. Front. Plant Sci. 2014, 5, 73. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Hara-Nishimura, I.; Nishimura, M. Unique Defense Strategy by the Endoplasmic Reticulum Body in Plants. Plant Cell Physiol. 2011, 52, 2039–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazaki, A.; Yamada, K.; Kunieda, T.; Sugiyama, R.; Hirai, M.Y.; Tamura, K.; Hara-Nishimura, I.; Shimada, T. Leaf Endoplasmic Reticulum Bodies Identified in Arabidopsis Rosette Leaves Are Involved in Defense against Herbivory. Plant Physiol. 2019, 179, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Beyer, E.M. A Potent Inhibitor of Ethylene Action in Plants. Plant Physiol. 1976, 58, 268–271. [Google Scholar] [CrossRef] [Green Version]
- Plett, J.M.; Mathur, J.; Regan, S. Ethylene Receptor ETR2 Controls Trichome Branching by Regulating Microtubule Assembly in Arabidopsis Thaliana. J. Exp. Bot. 2009, 60, 3923–3933. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, D.; Xavier, P.L.; Chaudhari, K.; John, R.; Dasgupta, A.K.; Pradeep, T.; Chakrabarti, G. Unprecedented Inhibition of Tubulin Polymerization Directed by Gold Nanoparticles Inducing Cell Cycle Arrest and Apoptosis. Nanoscale 2013, 5, 4476–4489. [Google Scholar] [CrossRef]
- Wen, Y.; Geitner, N.K.; Chen, R.; Ding, F.; Chen, P.; Andorfer, R.E.; Govindan, P.N.; Ke, P.C. Binding of Cytoskeletal Proteins with Silver Nanoparticles. RSC Adv. 2013, 3, 22002. [Google Scholar] [CrossRef]
- Li, S.; Lei, L.; Somerville, C.R.; Gu, Y. Cellulose Synthase Interactive Protein 1 (CSI1) Links Microtubules and Cellulose Synthase Complexes. Proc. Natl. Acad. Sci. USA 2012, 109, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Mirabet, V.; Krupinski, P.; Hamant, O.; Meyerowitz, E.M.; Jönsson, H.; Boudaoud, A. The Self-Organization of Plant Microtubules inside the Cell Volume Yields Their Cortical Localization, Stable Alignment, and Sensitivity to External Cues. PLoS Comput. Biol. 2018, 14, e1006011. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Zhu, C.; Chen, W.; Dixit, R. FRA1 Kinesin Modulates the Lateral Stability of Cortical Microtubules through Cellulose Synthase–Microtubule Uncoupling Proteins. Plant Cell 2020, 32, 2508–2524. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Han, L.; Feng, Z.; Wang, G.; Liu, W.; Ma, Y.; Yu, Y.; Kong, Z. Orchestration of Microtubules and the Actin Cytoskeleton in Trichome Cell Shape Determination by a Plant-Unique Kinesin. eLife 2015, 4, e09351. [Google Scholar] [CrossRef] [PubMed]
- Seung, D.; Webster, M.W.; Wang, R.; Andreeva, Z.; Marc, J. Dissecting the Mechanism of Abscisic Acid-Induced Dynamic Microtubule Reorientation Using Live Cell Imaging. Funct. Plant Biol. 2013, 40, 224. [Google Scholar] [CrossRef]
- Sakiyama-Sogo, M.; Shibaoka, H. Gibberellin A3 and Abscisic Acid Cause the Reorientation of Cortical Microtubules in Epicotyl Cells of the Decapitated Dwarf Pea. Plant Cell Physiol. 1993, 34, 431–437. [Google Scholar] [CrossRef]
- Lü, B.; Chen, F.; Gong, Z.H.; Xie, H.; Zhang, J.H.; Liang, J.S. Intracellular Localization of Integrin-like Protein and Its Roles in Osmotic Stress-Induced Abscisic Acid Biosynthesis in Zea Mays. Protoplasma 2007, 232, 35–43. [Google Scholar] [CrossRef]
- De Torres Zabala, M.; Bennett, M.H.; Truman, W.H.; Grant, M.R. Antagonism between Salicylic and Abscisic Acid Reflects Early Host-Pathogen Conflict and Moulds Plant Defence Responses. Plant J. 2009, 59, 375–386. [Google Scholar] [CrossRef]
- Li, H.; Xia, H.; Ding, W.; Li, Y.; Shi, Q.; Wang, D.; Tao, X. Synthesis of Monodisperse, Quasi-Spherical Silver Nanoparticles with Sizes Defined by the Nature of Silver Precursors. Langmuir 2014, 30, 2498–2504. [Google Scholar] [CrossRef]
- Ambrose, J.C.; Cyr, R. The Kinesin ATK5 Functions in Early Spindle Assembly in Arabidopsis. Plant Cell 2007, 19, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Ueda, K.; Matsuyama, T.; Hashimoto, T. Visualization of Microtubules in Living Cells of TransgenicArabidopsis Thaliana. Protoplasma 1999, 206, 201–206. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Blank | Ag+ | AgNPs | |
|---|---|---|---|---|
| pmol g−1 FW (mean ± StD) | pmol g−1 FW (mean ± StD) | pmol g−1 FW (mean ± StD) | ||
| cis-OPDA | WT | 32,560 ± 8007 b | 153,900 ± 8903 a | 158,100 ± 4047 a |
| gl-1 | 80,580 ± 3976 c | 38,820 ± 10,080 b | 145,600 ± 17,760 a | |
| JA | WT | 191± 24 d | 364 ± 26 ef | 1575 ± 55 g |
| gl-1 | 274 ± 48 e | 482 ± 97 f | 1802 ± 415 g | |
| JA-Ile | WT | 14.9 ± 2.5 h | 18.3 ± 0.6 h | 240 ± 22 j |
| gl-1 | 18.9 ± 1.5 h | 31.3 ± 2.6 i | 458 ± 60 k | |
| 9,10-DHJA | WT | 435 ± 133 lo | 721 ± 172 lm | 766 ± 86 lm |
| gl-1 | 267 ± 79 n | 200 ± 37 n | 1242 ± 416 m | |
| 11,12-OH-JA | WT | 1140 ± 168 q | 2400 ± 381 q | 10,730 ± 1013 r |
| gl-1 | 2653 ± 730 q | 1569 ± 259 qp | 15,230 ± 4360 r |
| Line | Blank | Ag+ | AgNPs | |
|---|---|---|---|---|
| pmol g−1 FW (mean ± StD) | pmol g−1 FW (mean ± StD) | pmol g−1 FW (mean ± StD) | ||
| ABA | WT | 3.88 ± 0.4 a | 6.55 ± 0.6 a | 51.2 ± 2.1 c |
| gl-1 | 7.92 ± 0.7 b | 6.78 ± 1.0 a | 115.0 ± 13.0 d | |
| DPA | WT | ND | ND | 270 ± 18 e |
| gl-1 | ND | ND | 630 ± 136 f | |
| PA | WT | ND | ND | 283 ± 17 g |
| gl-1 | ND | ND | 374 ± 42 g | |
| SA | WT | 1957 ± 293 h | 2124 ± 412 hi | 2857 ± 184 i |
| gl-1 | 1560 ± 192 h | 971 ± 162 j | 5032 ± 535 k |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelini, J.; Klassen, R.; Široká, J.; Novák, O.; Záruba, K.; Siegel, J.; Novotná, Z.; Valentová, O. Silver Nanoparticles Alter Microtubule Arrangement, Dynamics and Stress Phytohormone Levels. Plants 2022, 11, 313. https://doi.org/10.3390/plants11030313
Angelini J, Klassen R, Široká J, Novák O, Záruba K, Siegel J, Novotná Z, Valentová O. Silver Nanoparticles Alter Microtubule Arrangement, Dynamics and Stress Phytohormone Levels. Plants. 2022; 11(3):313. https://doi.org/10.3390/plants11030313
Chicago/Turabian StyleAngelini, Jindřiška, Ruslan Klassen, Jitka Široká, Ondřej Novák, Kamil Záruba, Jakub Siegel, Zuzana Novotná, and Olga Valentová. 2022. "Silver Nanoparticles Alter Microtubule Arrangement, Dynamics and Stress Phytohormone Levels" Plants 11, no. 3: 313. https://doi.org/10.3390/plants11030313