
Lipidized PrRP Analog Exhibits Strong Anti-Obesity and Antidiabetic Properties in Old WKY Rats with Obesity and Glucose Intolerance
 , , ,
, , ,
Abstract
:1. Introduction
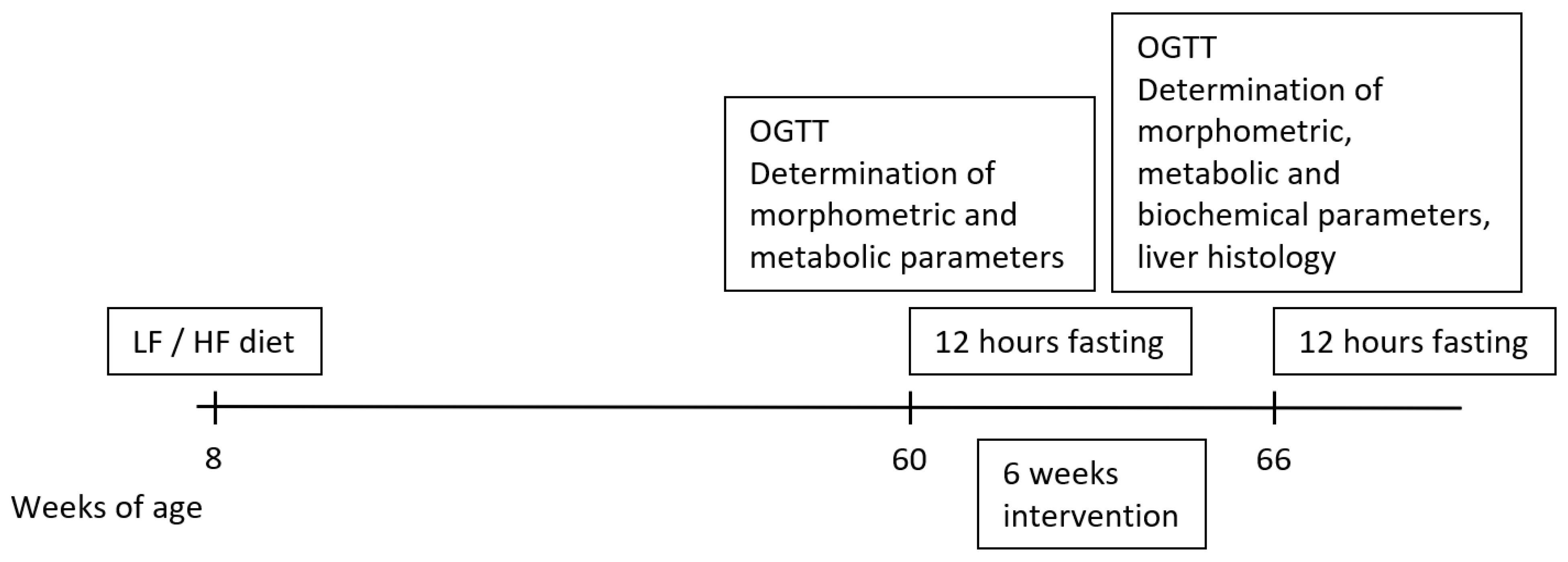
2. Materials and Methods
2.1. Substances
2.2. Experimental Animals
2.3. Oral Glucose Tolerance Test
2.4. Determination of Biochemical Parameters
2.5. Western Blotting
2.6. Liver Histology
2.7. NMR-Based Metabolomics in Urine and Plasma
2.8. MALDI MSI of Lipids in Liver
2.9. LC—MS Lipidomics in Liver
2.10. Determination of mRNA Expression
2.11. Immunohistochemistry
2.12. Statistical Analysis
3. Results
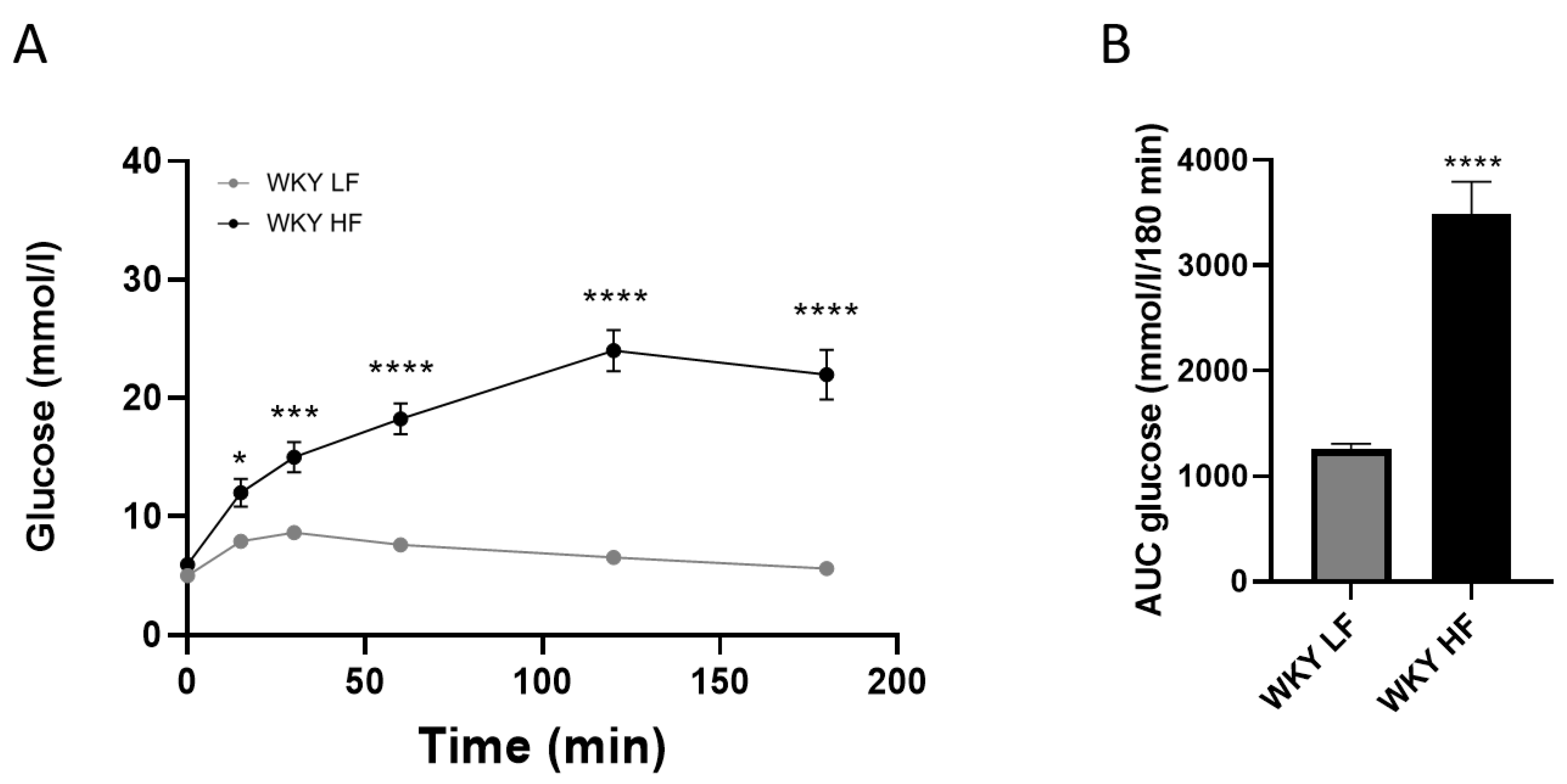
3.1. Metabolic Parameters of 60-Week-Old WKY Rats Fed a HF or LF Diet
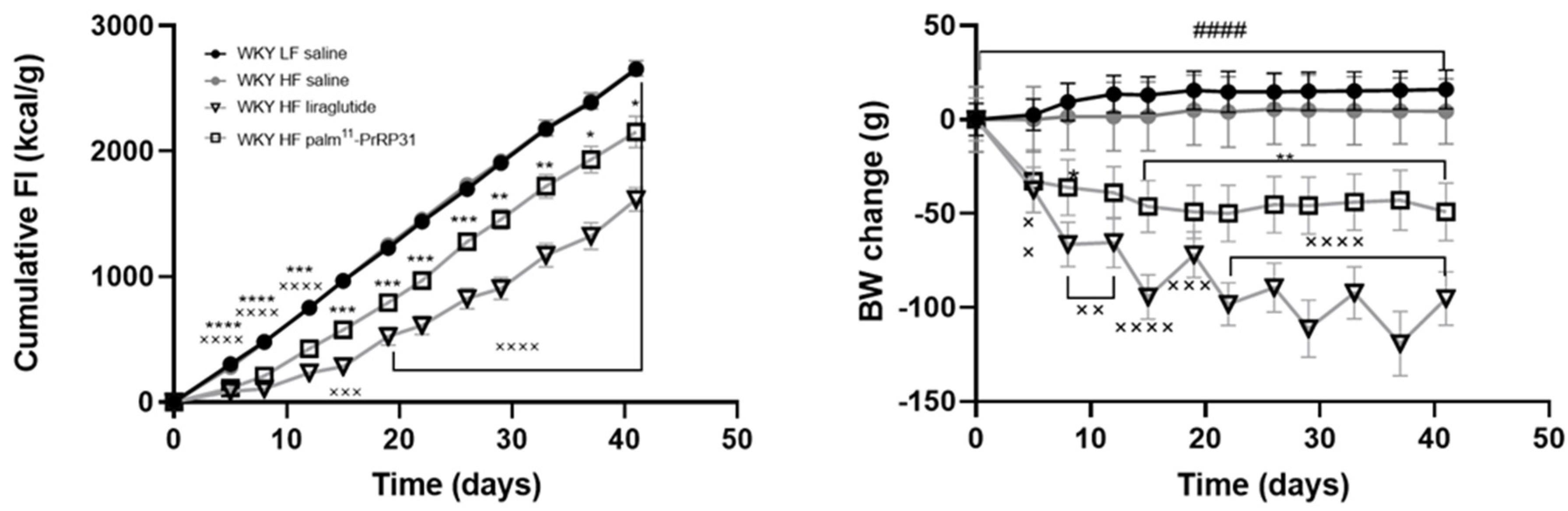
3.2. Palm11-PrRP31 and Liraglutide Treatment Significantly Decreased Body Weight, Food Intake and Plasma Leptin Levels
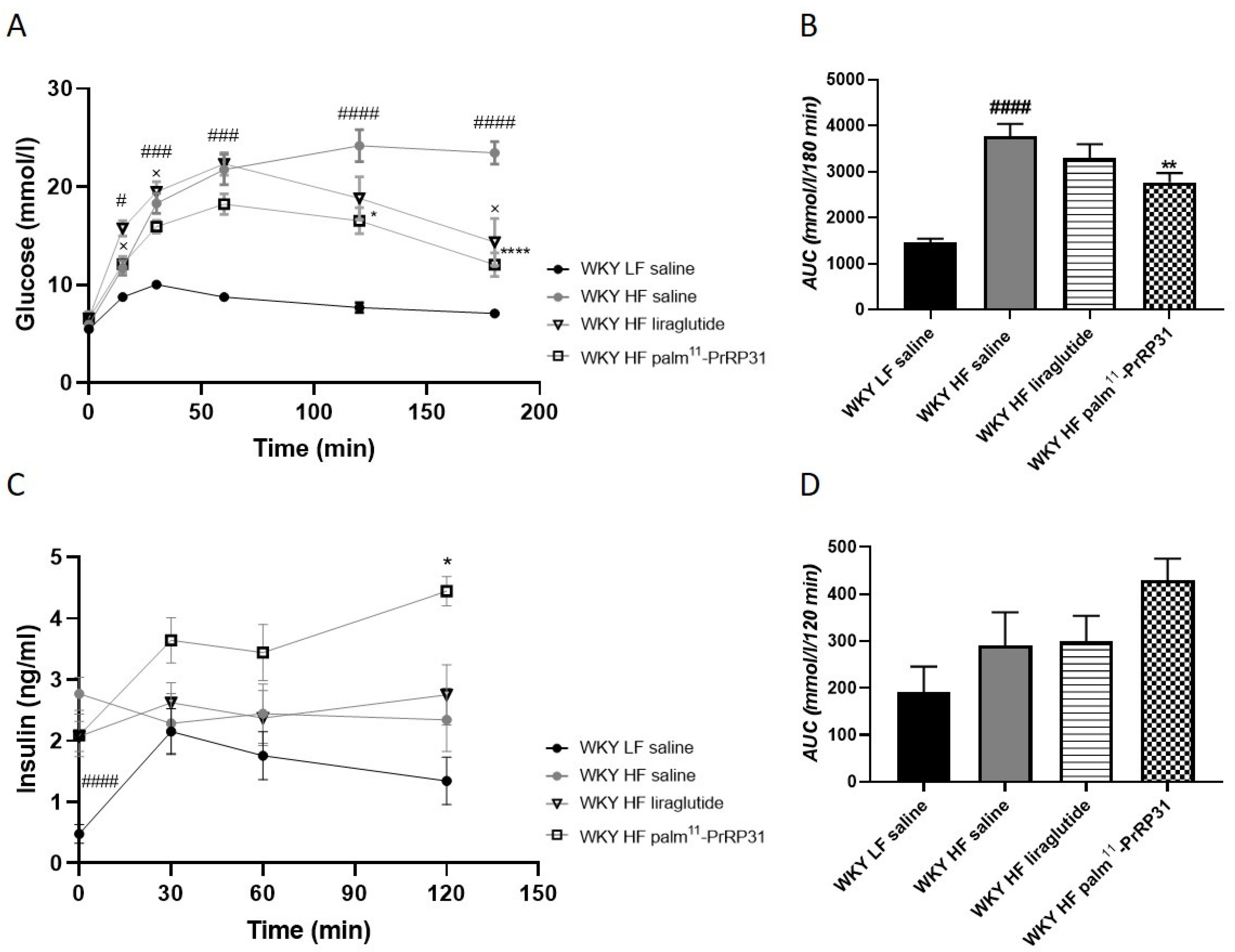
3.3. Palm11-PrRP31 and Liraglutide Treatment Significantly Improved Tolerance to Glucose
3.4. NMR-Based Metabolomics
3.4.1. The Impact of a HF Diet on Urine and Plasma Metabolic Profiles
3.4.2. The Impact of Palm11-PrRP31 and Liraglutide Treatment on Urine and Plasma Metabolic Profiles
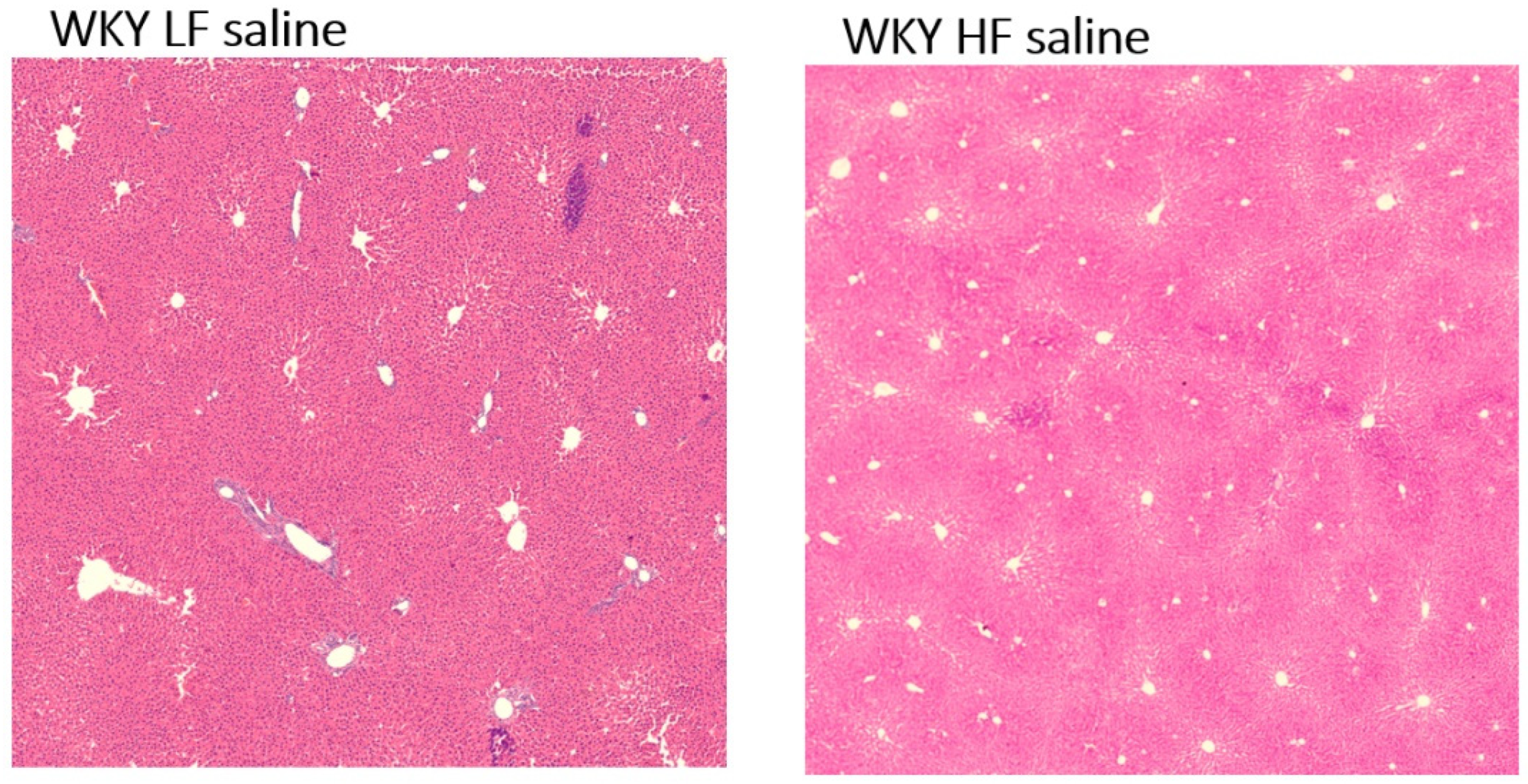
3.5. No Liver Steatosis but Different Lipid Profiles in 66-Week-Old WKY Rats Fed a HF Diet
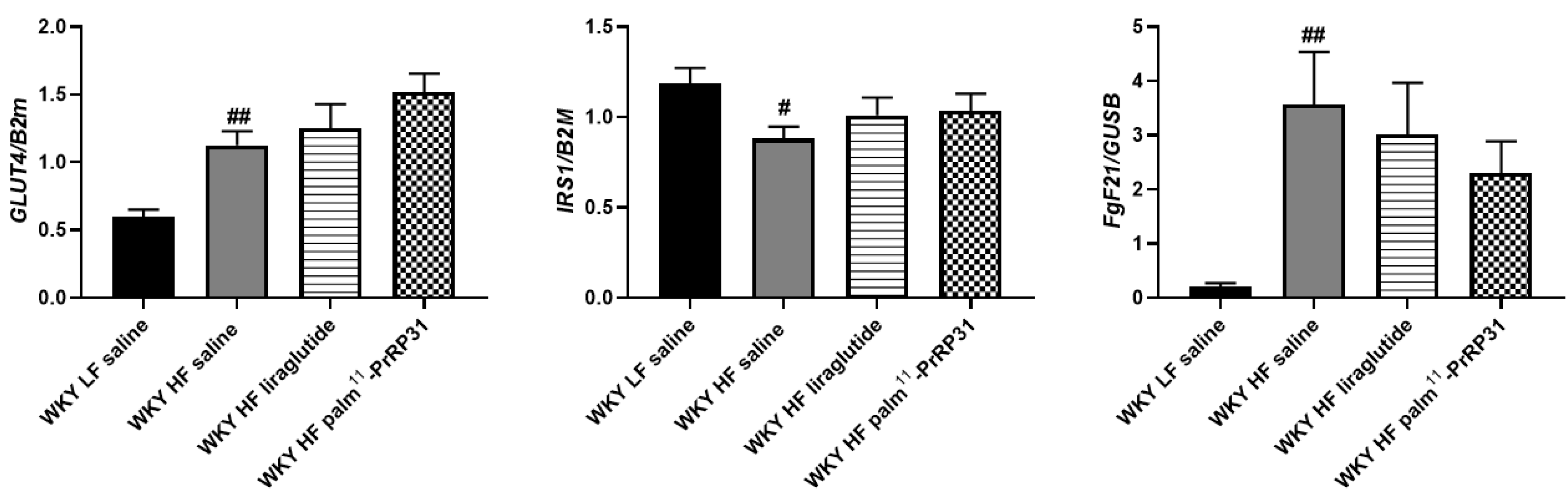
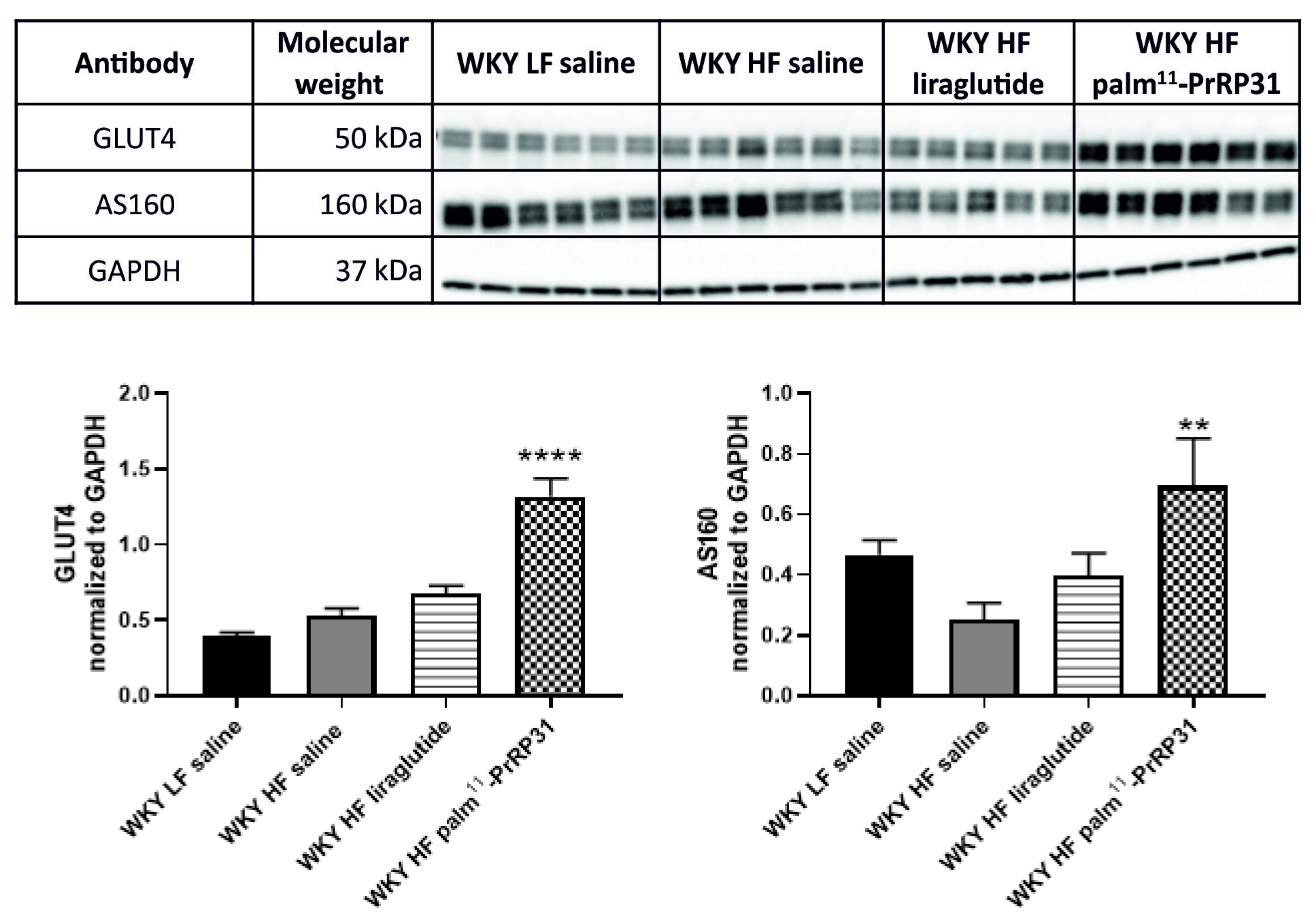
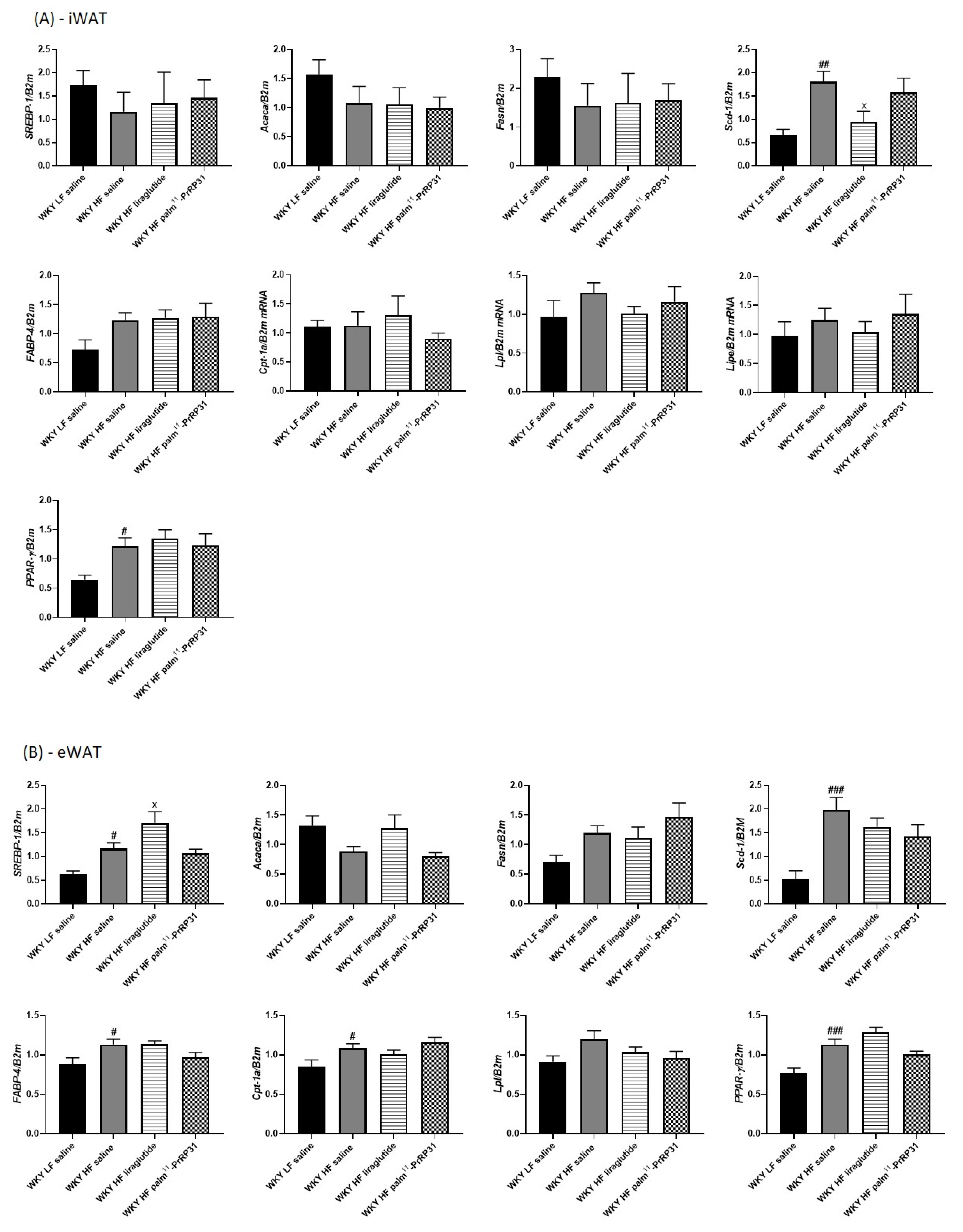
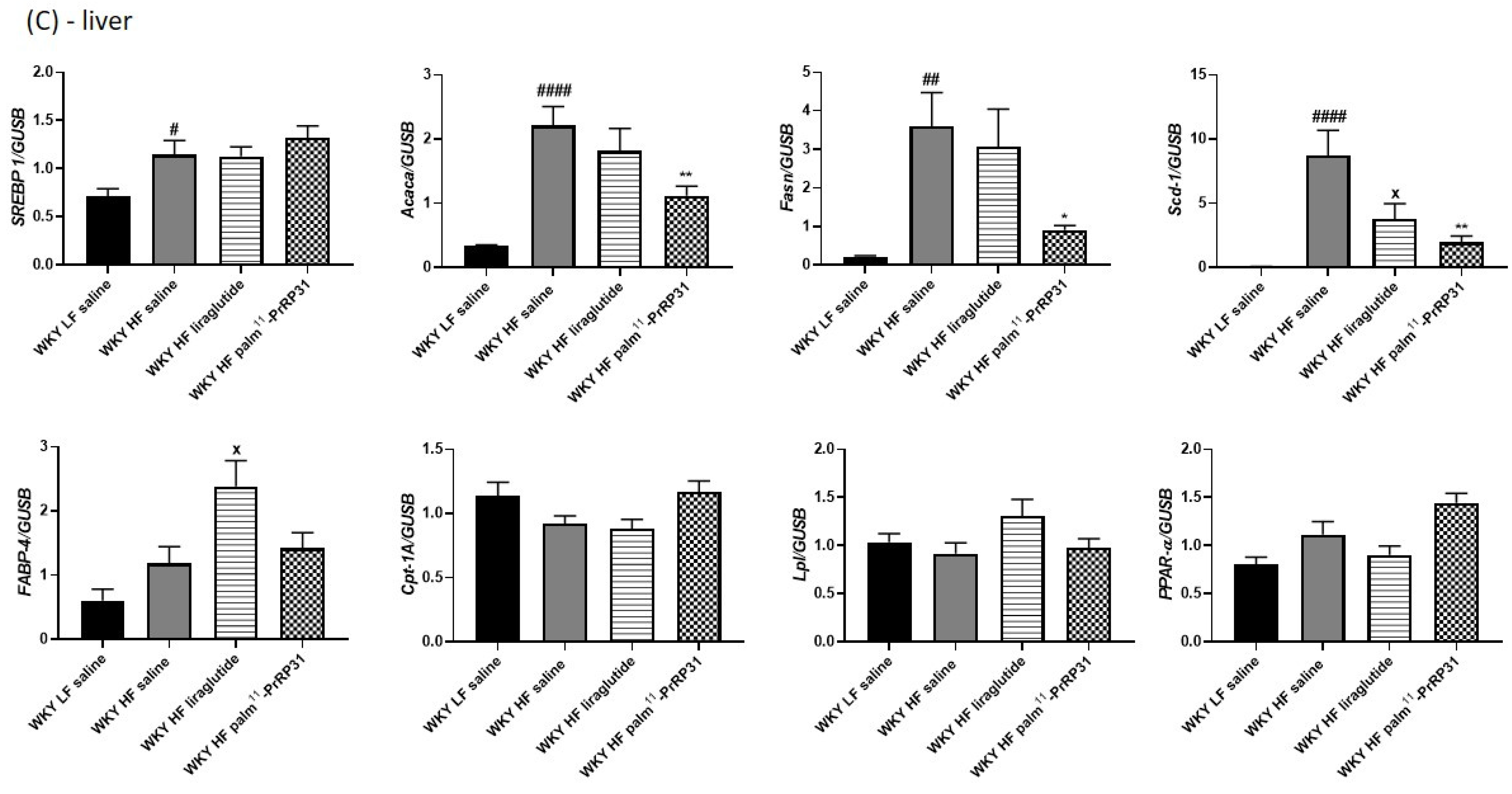
3.6. Liver Lipogenesis Was Stimulated in 66-Week-Old WKY Rats Fed a HF Diet, and Palm11-PrRP31 Treatment Attenuated the mRNA Expression of Genes Regulating Liver Lipogenesis
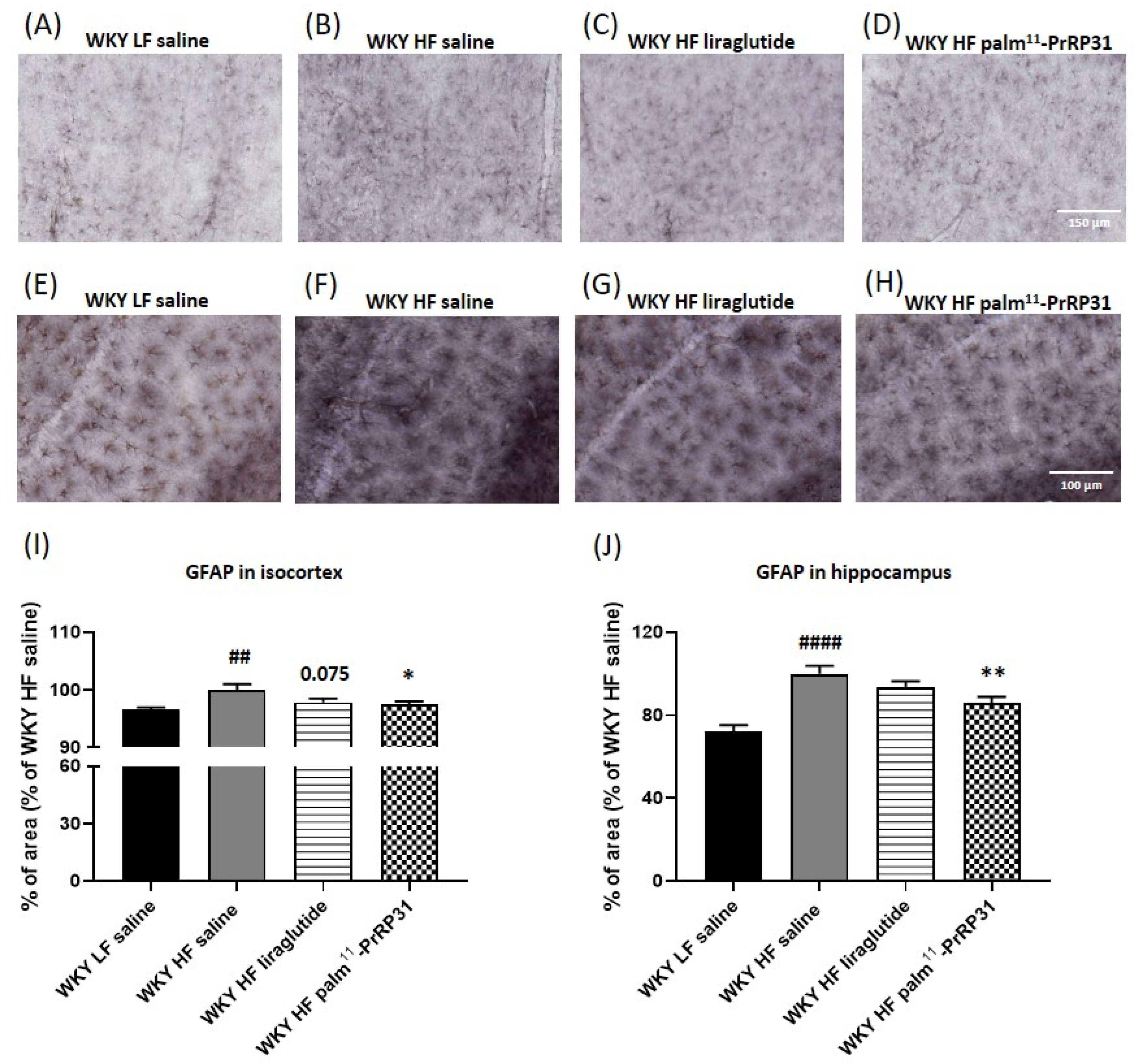
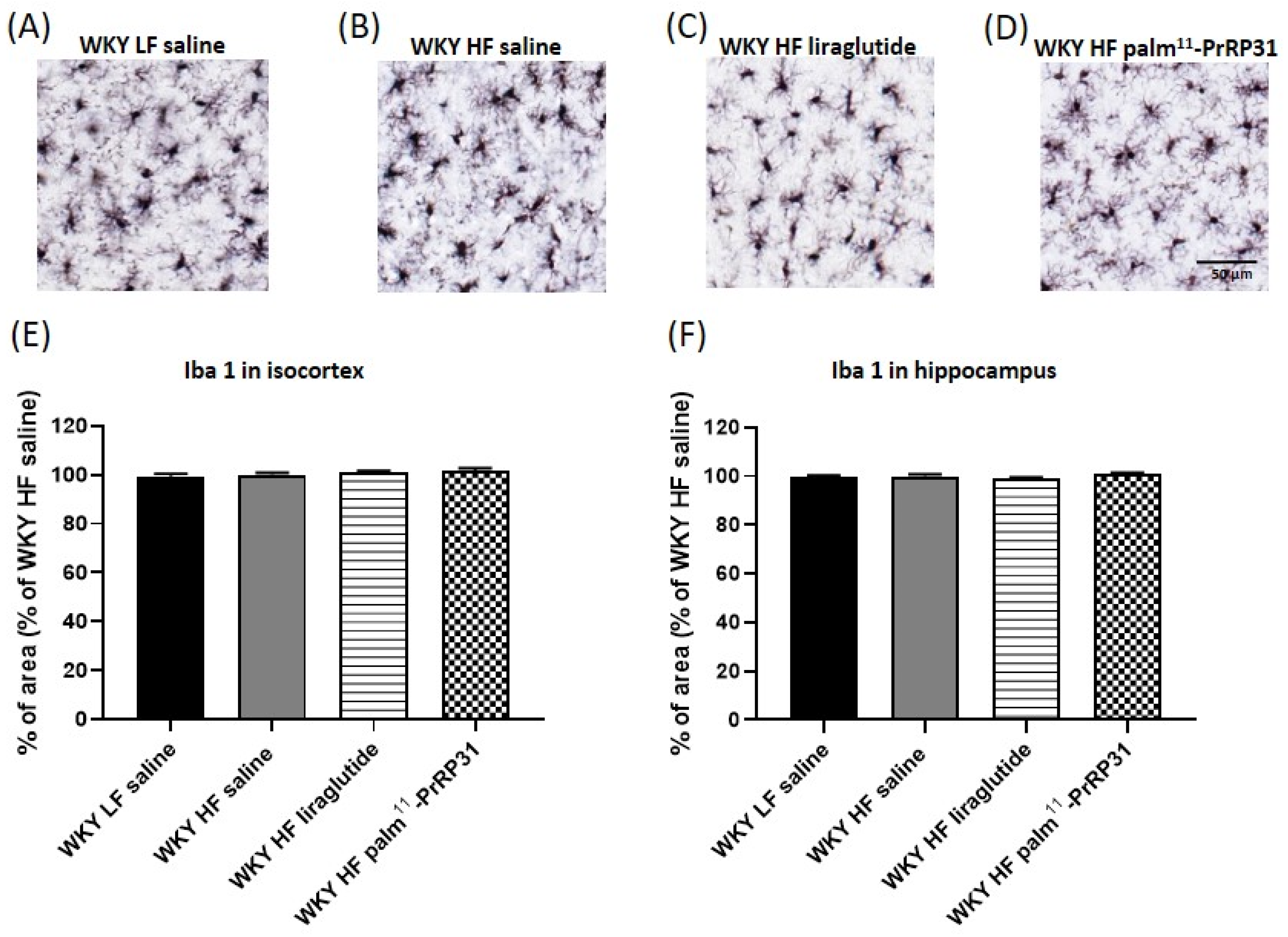
3.7. Palm11-PrRP31 Attenuated Astrocytosis in the Isocortex and Hippocampus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Git, K.C.G.; Peterse, C.; Beerens, S.; Luijendijk, M.C.M.; van der Plasse, G.; la Fleur, S.E.; Adan, R.A.H. Is leptin resistance the cause or the consequence of diet-induced obesity? Int. J. Obes. 2018, 42, 1445–1457. [Google Scholar] [CrossRef] [PubMed]
- Engin, A. The Definition and Prevalence of Obesity and Metabolic Syndrome. Adv. Exp. Med. Biol. 2017, 960, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Satoh, H.; Watanabe, T. Liraglutide enhances insulin sensitivity by activating AMP-activated protein kinase in male Wistar rats. Endocrinology 2014, 155, 3288–3301. [Google Scholar] [CrossRef] [PubMed]
- Quinn, L. Mechanisms in the Development of Type 2 Diabetes Mellitus. J. Cardiovasc. Nurs. 2002, 16. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Dey, D.; Roy, S.S. Molecular mechanism of insulin resistance. J. Biosci. 2007, 32, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Chawla, S.; Guchhait, P. Type-2 diabetes: Current understanding and future perspectives. IUBMB Life 2015, 67, 506–513. [Google Scholar] [CrossRef]
- Rojano, A.; Sena, E.; Manzano-Nuñez, R.; Pericàs, J.M.; Ciudin, A. NAFLD as the metabolic hallmark of obesity. Intern. Emerg. Med. 2022. [Google Scholar] [CrossRef]
- Arch, J.R. Horizons in the Pharmacotherapy of Obesity. Curr. Obes. Rep. 2015, 4, 451–459. [Google Scholar] [CrossRef]
- Patel, D. Pharmacotherapy for the management of obesity. Metabolism 2015, 64, 1376–1385. [Google Scholar] [CrossRef]
- Bray, G.A.; Frühbeck, G.; Ryan, D.H.; Wilding, J.P.H. Management of obesity. Lancet 2016, 387, 1947–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dailey, M.J.; Moran, T.H. Glucagon-like peptide 1 and appetite. Trends Endocrinol. Metab. 2013, 24, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladenheim, E.E. Liraglutide and obesity: A review of the data so far. Drug Des. Devel. Ther. 2015, 9, 1867–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, L.B.; Lau, J. The Discovery and Development of Liraglutide and Semaglutide. Front. Endocrinol. 2019, 10, 155. [Google Scholar] [CrossRef] [Green Version]
- Krieger, J.P. Intestinal glucagon-like peptide-1 effects on food intake: Physiological relevance and emerging mechanisms. Peptides 2020, 131, 170342. [Google Scholar] [CrossRef]
- Horowitz, M.; Flint, A.; Jones, K.L.; Hindsberger, C.; Rasmussen, M.F.; Kapitza, C.; Doran, S.; Jax, T.; Zdravkovic, M.; Chapman, I.M. Effect of the once-daily human GLP-1 analogue liraglutide on appetite, energy intake, energy expenditure and gastric emptying in type 2 diabetes. Diabetes Res. Clin. Pract. 2012, 97, 258–266. [Google Scholar] [CrossRef]
- Decara, J.; Arrabal, S.; Beiroa, D.; Rivera, P.; Vargas, A.; Serrano, A.; Pavón, F.J.; Ballesteros, J.; Dieguez, C.; Nogueiras, R.; et al. Antiobesity efficacy of GLP-1 receptor agonist liraglutide is associated with peripheral tissue-specific modulation of lipid metabolic regulators. Biofactors 2016, 42, 600–611. [Google Scholar] [CrossRef]
- Mráziková, L.; Neprašová, B.; Mengr, A.; Popelová, A.; Strnadová, V.; Holá, L.; Železná, B.; Kuneš, J.; Maletínská, L. Lipidized Prolactin-Releasing Peptide as a New Potential Tool to Treat Obesity and Type 2 Diabetes Mellitus: Preclinical Studies in Rodent Models. Front. Pharmacol. 2021, 12, 779962. [Google Scholar] [CrossRef]
- Maletínská, L.; Nagelová, V.; Tichá, A.; Zemenová, J.; Pirník, Z.; Holubová, M.; Špolcová, A.; Mikulášková, B.; Blechová, M.; Sýkora, D.; et al. Novel lipidized analogs of prolactin-releasing peptide have prolonged half-lives and exert anti-obesity effects after peripheral administration. Int. J. Obes. 2015, 39, 986–993. [Google Scholar] [CrossRef]
- Pražienková, V.; Holubová, M.; Pelantová, H.; Bugáňová, M.; Pirník, Z.; Mikulášková, B.; Popelová, A.; Blechová, M.; Haluzík, M.; Železná, B.; et al. Impact of novel palmitoylated prolactin-releasing peptide analogs on metabolic changes in mice with diet-induced obesity. PLoS ONE 2017, 12, e0183449. [Google Scholar] [CrossRef]
- Čermáková, M.; Pelantová, H.; Neprašová, B.; Šedivá, B.; Maletínská, L.; Kuneš, J.; Tomášová, P.; Železná, B.; Kuzma, M. Metabolomic Study of Obesity and Its Treatment with Palmitoylated Prolactin-Releasing Peptide Analog in Spontaneously Hypertensive and Normotensive Rats. J. Proteome Res. 2019, 18, 1735–1750. [Google Scholar] [CrossRef] [PubMed]
- Holubová, M.; Zemenová, J.; Mikulášková, B.; Panajotova, V.; Stöhr, J.; Haluzík, M.; Kuneš, J.; Železná, B.; Maletínská, L. Palmitoylated PrRP analog decreases body weight in DIO rats but not in ZDF rats. J. Endocrinol. 2016, 229, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Holubová, M.; Hrubá, L.; Neprašová, B.; Majerčíková, Z.; Lacinová, Z.; Kuneš, J.; Maletínská, L.; Železná, B. Prolactin-releasing peptide improved leptin hypothalamic signaling in obese mice. J. Mol. Endocrinol. 2018, 60, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikulaskova, B.; Holubova, M.; Prazienkova, V.; Zemenova, J.; Hruba, L.; Haluzik, M.; Zelezna, B.; Kunes, J.; Maletinska, L. Lipidized prolactin-releasing peptide improved glucose tolerance in metabolic syndrome: Koletsky and spontaneously hypertensive rat study. Nutr. Diabetes 2018, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Spolcova, A.; Mikulaskova, B.; Holubova, M.; Nagelova, V.; Pirnik, Z.; Zemenova, J.; Haluzik, M.; Zelezna, B.; Galas, M.C.; Maletinska, L. Anorexigenic lipopeptides ameliorate central insulin signaling and attenuate tau phosphorylation in hippocampi of mice with monosodium glutamate-induced obesity. J. Alzheimers Dis. 2015, 45, 823–835. [Google Scholar] [CrossRef]
- Kořínková, L.; Holubová, M.; Neprašová, B.; Hrubá, L.; Pražienková, V.; Bencze, M.; Haluzík, M.; Kuneš, J.; Maletínská, L.; Železná, B. Synergistic effect of leptin and lipidized PrRP on metabolic pathways in ob/ob mice. J. Mol. Endocrinol. 2020, 64, 77–90. [Google Scholar] [CrossRef]
- Pražienková, V.; Funda, J.; Pirník, Z.; Karnošová, A.; Hrubá, L.; Kořínková, L.; Neprašová, B.; Janovská, P.; Benzce, M.; Kadlecová, M.; et al. GPR10 gene deletion in mice increases basal neuronal activity, disturbs insulin sensitivity and alters lipid homeostasis. Gene 2021, 774, 145427. [Google Scholar] [CrossRef]
- Prochazkova, M.; Budinska, E.; Kuzma, M.; Pelantova, H.; Hradecky, J.; Heczkova, M.; Daskova, N.; Bratova, M.; Modos, I.; Videnska, P.; et al. Vegan Diet Is Associated With Favorable Effects on the Metabolic Performance of Intestinal Microbiota: A Cross-Sectional Multi-Omics Study. Front. Nutr. 2021, 8, 783302. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Strnad, Š.; Vrkoslav, V.; Klimšová, Z.; Zemenová, J.; Cvačka, J.; Maletínská, L.; Sýkora, D. Application of matrix-assisted laser desorption/ionization mass spectrometry imaging in combination with LC-MS in pharmacokinetic study of metformin. Bioanalysis 2018, 10, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Stoeckli, M.; Staab, D.; Wetzel, M.; Brechbuehl, M. iMatrixSpray: A free and open source sample preparation device for mass spectrometric imaging. Chimia 2014, 68, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Strnad, Š.; PraŽienková, V.; Holubová, M.; Sýkora, D.; Cvačka, J.; Maletínská, L.; Železná, B.; Kuneš, J.; Vrkoslav, V. Mass spectrometry imaging of free-floating brain sections detects pathological lipid distribution in a mouse model of Alzheimer’s-like pathology. Analyst 2020, 145, 4595–4605. [Google Scholar] [CrossRef] [PubMed]
- Löfgren, L.; Forsberg, G.-B.; Ståhlman, M. The BUME method: A new rapid and simple chloroform-free method for total lipid extraction of animal tissue. Sci. Rep. 2016, 6, 27688. [Google Scholar] [CrossRef] [Green Version]
- Lansang, M.C.; Williams, G.H.; Carroll, J.S. Correlation between the glucose clamp technique and the homeostasis model assessment in hypertension. Am. J. Hypertens. 2001, 14, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Zemenova, J.; Sykora, D.; Freislebenova, A.; Maletinska, L. LC-MS/MS analysis of lipidized analogs of prolactin-releasing peptide utilizing a monolithic column and simple sample preparation. Bioanalysis 2017, 9, 1319–1328. [Google Scholar] [CrossRef]
- Hayes, M.R.; Kanoski, S.E.; Alhadeff, A.L.; Grill, H.J. Comparative effects of the long-acting GLP-1 receptor ligands, liraglutide and exendin-4, on food intake and body weight suppression in rats. Obesity 2011, 19, 1342–1349. [Google Scholar] [CrossRef]
- Hansen, G.; Jelsing, J.; Vrang, N. Effects of liraglutide and sibutramine on food intake, palatability, body weight and glucose tolerance in the gubra DIO-rats. Acta Pharmacol. Sin. 2012, 33, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Szczepańska, E.; Gietka-Czernel, M. FGF21: A Novel Regulator of Glucose and Lipid Metabolism and Whole-Body Energy Balance. Horm. Metab. Res. 2022, 54, 203–211. [Google Scholar] [CrossRef]
- Cartland, S.P.; Tamer, N.; Patil, M.S.; Di Bartolo, B.A.; Kavurma, M.M. A “Western Diet” promotes symptoms of hepatic steatosis in spontaneously hypertensive rats. Int. J. Exp. Pathol. 2020, 101, 152–161. [Google Scholar] [CrossRef]
- Jani, S.; Da Eira, D.; Stefanovic, M.; Ceddia, R.B. The ketogenic diet prevents steatosis and insulin resistance by reducing lipogenesis, diacylglycerol accumulation and protein kinase C activity in male rat liver. J. Physiol. 2022, 600, 4137–4151. [Google Scholar] [CrossRef] [PubMed]
- Pelantova, H.; Bartova, S.; Anyz, J.; Holubova, M.; Zelezna, B.; Maletinska, L.; Novak, D.; Lacinova, Z.; Sulc, M.; Haluzik, M.; et al. Metabolomic profiling of urinary changes in mice with monosodium glutamate-induced obesity. Anal. Bioanal. Chem. 2016, 408, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Kim, I.Y.; Kim, Y.N.; Kim, J.S.; Shin, J.H.; Jang, Z.H.; Lee, H.S.; Hwang, G.S.; Seong, J.K. 1H NMR-based metabolite profiling of diet-induced obesity in a mouse mode. BMB Rep. 2012, 45, 419–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Zhang, L.; Dong, F.; Liu, Y.; Li, N.; Li, H.; Lei, H.; Hao, F.; Wang, Y.; Zhu, Y.; et al. Metabonomic Changes Associated with Atherosclerosis Progression for LDLR(-/-) Mice. J. Proteome Res. 2015, 14, 2237–2254. [Google Scholar] [CrossRef] [PubMed]
- Vairetti, M.; Carini, R.; De Cesaris, M.G.; Splendore, R.; Richelmi, P.; Bertè, F.; Albano, E. Beta-alanine protection against hypoxic liver injury in the rat. Biochim. Biophys. Acta 2002, 1587, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterfield, C.J.; Turton, J.A.; Scales, M.D.; Timbrell, J.A. Taurine, a possible urinary marker of liver damage: A study of taurine excretion in carbon tetrachloride-treated rats. Arch. Toxicol. 1991, 65, 548–555. [Google Scholar] [CrossRef]
- Sun, Y.J.; Wang, H.P.; Liang, Y.J.; Yang, L.; Li, W.; Wu, Y.J. An NMR-based metabonomic investigation of the subacute effects of melamine in rats. J. Proteome Res. 2012, 11, 2544–2550. [Google Scholar] [CrossRef]
- Gaster, M.; Rustan, A.C.; Beck-Nielsen, H. Differential utilization of saturated palmitate and unsaturated oleate: Evidence from cultured myotubes. Diabetes 2005, 54, 648–656. [Google Scholar] [CrossRef] [Green Version]
- Alkhouri, N.; Dixon, L.J.; Feldstein, A.E. Lipotoxicity in nonalcoholic fatty liver disease: Not all lipids are created equal. Expert Rev. Gastroenterol. Hepatol. 2009, 3, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Solon-Biet, S.M.; Cogger, V.C.; Pulpitel, T.; Heblinski, M.; Wahl, D.; McMahon, A.C.; Warren, A.; Durrant-Whyte, J.; Walters, K.A.; Krycer, J.R.; et al. Defining the Nutritional and Metabolic Context of FGF21 Using the Geometric Framework. Cell Metab. 2016, 24, 555–565. [Google Scholar] [CrossRef]
- Flippo, K.H.; Potthoff, M.J. Metabolic Messengers: FGF21. Nat. Metab. 2021, 3, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Ooi, G.J.; Meikle, P.J.; Huynh, K.; Earnest, A.; Roberts, S.K.; Kemp, W.; Parker, B.L.; Brown, W.; Burton, P.; Watt, M.J. Hepatic lipidomic remodeling in severe obesity manifests with steatosis and does not evolve with non-alcoholic steatohepatitis. J. Hepatol. 2021, 75, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N.; Nagana Gowda, G.A.; Djukovic, D.; Raftery, D. Distinguishing NASH Histological Severity Using a Multiplatform Metabolomics Approach. Metabolites 2020, 10, 168. [Google Scholar] [CrossRef] [PubMed]
- Holubova, M.; Hruba, L.; Popelova, A.; Bencze, M.; Prazienkova, V.; Gengler, S.; Kratochvilova, H.; Haluzik, M.; Zelezna, B.; Kunes, J.; et al. Liraglutide and a lipidized analog of prolactin-releasing peptide show neuroprotective effects in a mouse model of beta-amyloid pathology. Neuropharmacology 2019, 144, 377–387. [Google Scholar] [CrossRef]
- Prazienkova, V.; Ticha, A.; Blechova, M.; Spolcova, A.; Zelezna, B.; Maletinska, L. Pharmacological characterization of lipidized analogs of prolactin-releasing peptide with a modified C-terminal aromatic ring. J. Physiol. Pharmacol. 2016, 67, 121–128. [Google Scholar]
- Mikulaskova, B.; Zemenova, J.; Pirnik, Z.; Prazienkova, V.; Bednarova, L.; Zelezna, B.; Maletinska, L.; Kunes, J. Effect of palmitoylated prolactin-relea sing peptide on food intake and neural activation after different routes of peripheral administration in rats. Peptides 2016, 75, 109–117. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ADIPOSE TISSUE | Acaca | LIVER | Acaca |
| Cpt-1a | Cpt-1 | ||
| FABP-4 | FABP-4 | ||
| Fasn | Fasn | ||
| GLUT-4 | Lpl | ||
| Irs-1 | Ppar-α | ||
| Lipe | Scd-1 | ||
| Lpl | SREBP-1 | ||
| Ppar-γ | |||
| Scd-1 | |||
| SREBP-1 |
| Age | 60 Weeks | |
|---|---|---|
| Group | WKY LF | WKY HF |
| BW (g) | 439.25 ± 6.85 | 589.42 ± 6.81 **** |
| Glucose (mmol/L) | 5.00 ± 0.25 | 6.11 ± 0.09 **** |
| Insulin (ng/mL) | 0.53 ± 0.12 | 2.95 ± 0.19 *** |
| HOMA index | 20.85 ± 5.07 | 133.46 ± 11.60 **** |
| Leptin (ng/mL) | 2.29 ± 0.28 | 33.02 ± 1.75 **** |
| TG (mmol/L) | 0.46 ± 0.06 | 0.47 ± 0.02 |
| CHOL (mmol/L) | 3.51 ± 0.25 | 3.20 ± 0.09 |
| Age | 66 Weeks | |||
|---|---|---|---|---|
| Group | WKY LF Saline | WKY HF Saline | WKY HF Liraglutide | WKY HF Palm11-PrRP31 |
| BW (g) | 444.00 ± 8.55 | 591.00 ± 12.37 #### | 456.83 ± 11.77 xxxx | 519.14 ± 9.05 *** |
| Liver weight (g) | 10.81 ± 0.55 | 11.20 ± 0.435 | 9.76 ± 0.40 | 10.42 ± 0.34 |
| Leptin (ng/mL) | 3.13 ± 0.60 | 24.52 ± 2.22 #### | 12.60 ± 2.13 xxx | 15.14 ± 2.07 ** |
| TG (mmol/L) | 0.54 ± 0.06 | 0.56 ± 0.03 | 0.38 ± 0.03 | 0.59 ± 0.05 |
| CHOL (mmol/L) | 2.79 ± 0.12 | 2.64 ± 0.15 | 2.47 ± 0.13 | 2.50 ± 0.15 |
| Glucose (mmol/L) | 5.49 ± 0.17 | 5.98 ± 0.17 | 6.62 ± 0.65 | 6.56 ± 0.28 |
| HbA1c (mmol/mol) | 2.00 ± 0.09 | 2.62 ± 0.13 ## | 1.86 ± 0.13 xxx | 2.27 ± 0.08 |
| Insulin (ng/mL) | 0.48 ± 0.16 | 2.77 ± 0.27 #### | 2.07 ± 0.25 | 2.09 ± 0.35 |
| HOMA index | 20.65 ± 6.69 | 127.25 ± 13.29 #### | 104.73 ± 15.00 | 116.58 ± 19.38 |
| Glucagon (pg/mL) | 4.33 ± 0.05 | 4.58 ± 0.04 ## | 4.38 ± 0.04 x | 4.41 ± 0.07 |
| FGF21 (pg/mL) | 262.96 ± 36.90 | 607.63 ± 147.23 # | 173.85 ± 31.75 x | 512.64 ± 149.38 |
| Palm11-PrRP31 vs. HF Saline | Liraglutide vs. HF Saline | LF Saline vs. HF Saline | ||||
|---|---|---|---|---|---|---|
| Metabolite | ∆ [%] | ∆ [%] | ∆ [%] | |||
| URINE | ||||||
| 1-Methylnicotinamide | −15.18 | 43.10 | * | −28.33 | ||
| Sucrose | 78.01 | * | 10.19 | −60.34 | ||
| Creatinine | −18.73 | * | −19.69 | * | −27.38 | ** |
| Taurine | −30.07 | −53.85 | ** | −78.03 | * | |
| β-alanine | −40.72 | *** | −44.33 | *** | −58.79 | *** |
| 2-Hydroxyisobutyrate | −12.87 | 75.28 | * | −2.03 | ||
| PLASMA | ||||||
| Glucose | −30.89 | ** | −28.50 | * | −32.94 | ** |
| Arabinose | −32.88 | ** | −31.52 | * | −37.79 | ** |
| Sucrose | −22.62 | × | −23.25 | × | −33.34 | ** |
| Unidentified saccharide | −26.47 | ** | −27.30 | ** | −29.65 | ** |
| 3-Hydroxybutyrate | −15.88 | −49.10 | * | −31.68 | ||
| Creatine | −31.10 | ** | −29.09 | * | −33.61 | ** |
| Fumarate | −38.62 | * | −29.62 | −24.14 | ||
| Lactate | −45.52 | × | −11.94 | −40.01 | ||
| Glycine | 41.66 | ** | 7.44 | 23.63 | ||
| Serine | 37.94 | ** | 14.39 | 3.06 | ||
| Tryptophan | −21.25 | * | −0.17 | −5.27 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mráziková, L.; Hojná, S.; Vaculová, P.; Strnad, Š.; Vrkoslav, V.; Pelantová, H.; Kuzma, M.; Železná, B.; Kuneš, J.; Maletínská, L. Lipidized PrRP Analog Exhibits Strong Anti-Obesity and Antidiabetic Properties in Old WKY Rats with Obesity and Glucose Intolerance. Nutrients 2023, 15, 280. https://doi.org/10.3390/nu15020280
Mráziková L, Hojná S, Vaculová P, Strnad Š, Vrkoslav V, Pelantová H, Kuzma M, Železná B, Kuneš J, Maletínská L. Lipidized PrRP Analog Exhibits Strong Anti-Obesity and Antidiabetic Properties in Old WKY Rats with Obesity and Glucose Intolerance. Nutrients. 2023; 15(2):280. https://doi.org/10.3390/nu15020280
Chicago/Turabian StyleMráziková, Lucia, Silvie Hojná, Petra Vaculová, Štěpán Strnad, Vladimír Vrkoslav, Helena Pelantová, Marek Kuzma, Blanka Železná, Jaroslav Kuneš, and Lenka Maletínská. 2023. "Lipidized PrRP Analog Exhibits Strong Anti-Obesity and Antidiabetic Properties in Old WKY Rats with Obesity and Glucose Intolerance" Nutrients 15, no. 2: 280. https://doi.org/10.3390/nu15020280