
Spirostanol Saponins from Flowers of Allium Porrum and Related Compounds Indicating Cytotoxic Activity and Affecting Nitric Oxide Production Inhibitory Effect in Peritoneal Macrophages


Abstract
:1. Introduction
2. Results and Discussion
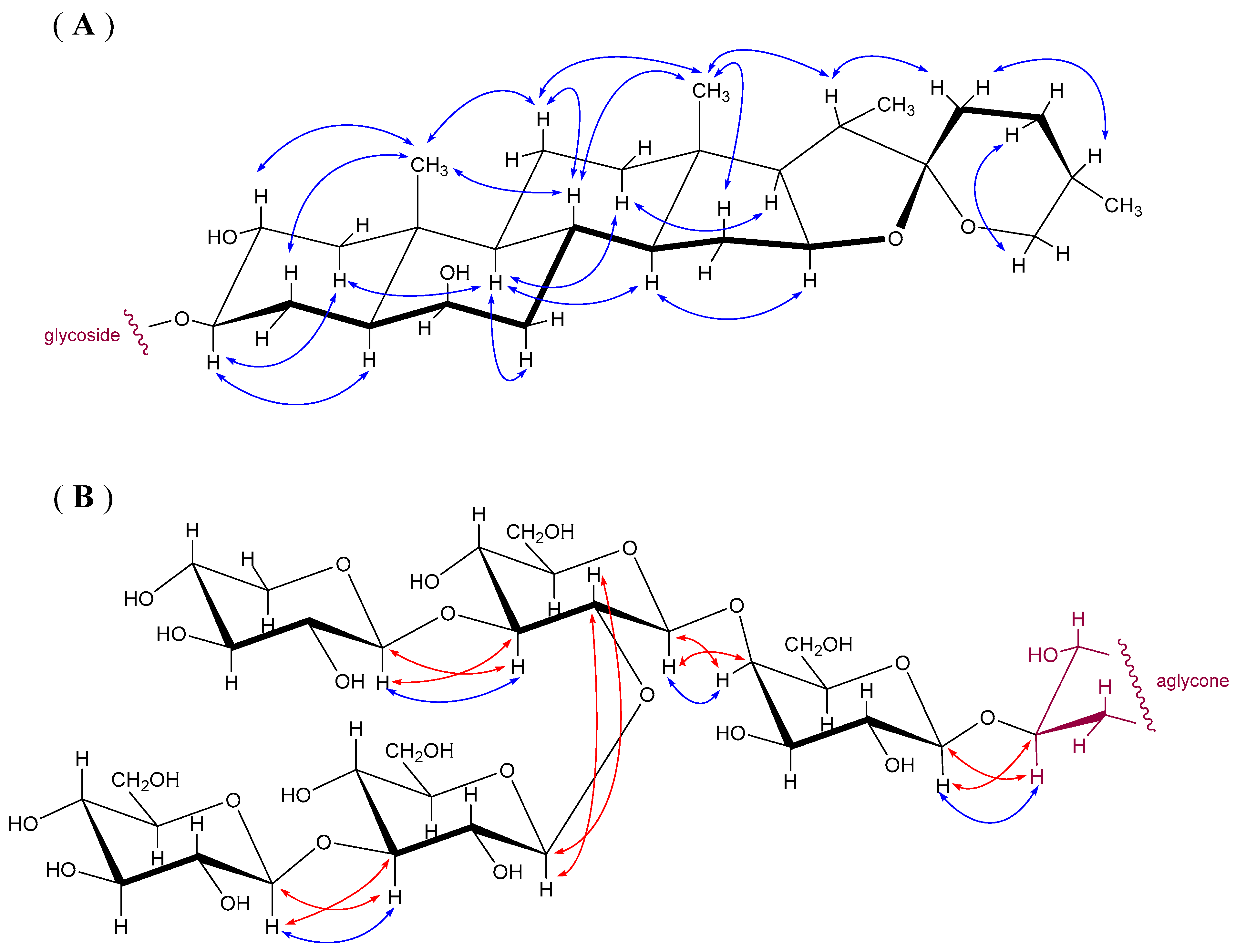
2.1. Chemical Identification and Structural Relations
2.2. Biological Activities of Selected Saponins
2.3. In Vitro Biological Effects
3. Materials and Methods
3.1. General Methods
3.2. Chemicals
3.3. Plant Material
3.4. Separation and Purification of Compounds
3.5. Identification of Compounds
3.5.1. Aginoside (1)
3.5.2. 6-Deoxy-Aginoside (2)
3.5.3. Yayoisaponin A (3), Previously Known also as Alliporin
3.5.4. Agigenin (4)
3.6. Biological Assays
3.6.1. Nitric Oxide (NO) Production
3.6.2. Cell Viability
3.6.3. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Hostettmann, K.; Marston, A. Book, Saponins. Chemistry and Pharmacology of Natural Products; Cambridge University Press: Cambridge, UK, 1995; pp. 287–306. [Google Scholar]
- Waller, G.R.; Yamasaki, K. (Eds.) Saponins Used in Traditional and Modern Medicine. Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 1996; Volume 404. [Google Scholar]
- Harmatha, J. Chemo-ecological role of spirostanol saponins in the interaction between plants and insects. In Book Saponins in Food, Feedstuffs and Medicinal Plants; Oleszek, W., Marston, A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; Volume 45, pp. 129–141. [Google Scholar]
- Lanzotti, V. Bioactive saponins from Allium and Aster plants. Phytochem. Rev. 2005, 4, 95–110. [Google Scholar] [CrossRef]
- Lanzotti, V. Bioactive polar natural compounds from garlic and onions. Phytochem. Rev. 2012, 11, 179–196. [Google Scholar] [CrossRef]
- Sobolewska, D.; Michalska, K.; Podolak, I.; Grabowska, K. Steroidal saponins from the genus Allium. Phytochem. Rev. 2016, 15, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Harmatha, J.; Vokáč, K.; Kmoníčková, E.; Zídek, Z. Lack of interference of common phytoecdysteroids with production of nitric oxide by immune-activated mammalian macrophages. Steroids 2008, 73, 466–471. [Google Scholar] [CrossRef]
- Harmatha, J.; Buděšínský, M.; Vokáč, K.; Kostecká, P.; Kmoníčková, E.; Zídek, Z. Trilobolide and related sesquiterpene lactones from Laser trilobum possessing immunobiological properties. Fitoterapia 2013, 89, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Harmatha, J.; Vokáč, K.; Buděšínský, M.; Zídek, Z.; Kmoníčková, E. Immunobiological properties of sesquiterpene lactones obtained by chemically transformed structural modifications of trilobolide. Fitoterapia 2015, 107, 90–99. [Google Scholar] [CrossRef]
- Harmatha, J.; Buděšínský, M.; Jurášek, M.; Zimmermann, T.; Drašar, P.; Zídek, Z.; Kmoníčková, E.; Vejvodová, L. Structural modification of trilobolide for upgrading its immunobiological properties and reducing its cytotoxic action. Fitoterapia 2019, 134, 88–95. [Google Scholar] [CrossRef]
- Arnault, C.; Harmatha, J.; Mauchamp, B.; Sláma, K. Influence of allelochemical substances of the host plant (Allium porrum) on development and moulting of Acrolepiopsis assectella (Lepidoptera). In Their Role as Selective Factor; Labeyrie, V., Fabres, G., Lachaise, D., Eds.; Insects-Plants, Junk Publishers: Dordrecht, The Netherlands, 1987; pp. 249–255. [Google Scholar]
- Harmatha, J.; Mauchamp, B.; Arnault, C.; Sláma, K. Identification of a spirostane-type saponin in the flowers of leek with inhibitory effects on growth of leek-moth larvae. Biochem. Syst. Ecol. 1987, 15, 113–116. [Google Scholar] [CrossRef]
- Arnault, C.; Mauchamp, B. Ecdysis inhibition in Acrolepiopsis assectella larvae by digitoxin: Antagonistic effects of cholesterol. Experientia 1985, 41, 1074–1077. [Google Scholar] [CrossRef]
- Harmatha, J.; Dinan, L. Interaction of dimeric ecdysteroids, glycosidic ecdysteroid conjugates and ecdysis-disturbing saponins with the ecdysteroid receptor assessed by means of the Drosophila melanogaster B-II bioassay. In Book Arthropods: Chemical, Physiological and Environmental Aspects; Konopinska, D., Ed.; Wroclaw Technology University Press: Wroclaw, Poland, 2002; pp. 79–84. [Google Scholar]
- Sata, N.; Matsunaga, S.; Fusetani, N.; Nushikawa, H.; Takamura, S.; Saito, T. New antifungal and cytotoxic steroidal saponins from the bulbs of elephant garlic mutant. Biosci. Biotechnol. Biochem. 1998, 62, 1904–1911. [Google Scholar] [CrossRef] [PubMed]
- Kelginbayev, A.N.; Gorovits, M.B.; Gorovits, T.T.; Abubakirov, N.K. Allium steroid saponins and sapogenins IX—Structure of aginosid. Khim. Prir. Soedin. 1976, 4, 480–486. [Google Scholar]
- Dinan, L.; Bourne, P.C.; Meng, Y.; Sarker, S.D.; Tolentino, R.B. Whiting, Assessment of natural products in the Drosophila melanogaster BII cell bioassay for ecdysteroid agonist and antagonist activities. CMLS Cell. Mol. Life Sci. 2001, 58, 321–342. [Google Scholar] [CrossRef]
- Carotenuto, A.; Fattorusso, E.; Lanzotti, V.; Magno, S. Spirostanol saponins of Allium porrum L. Phytochemistry 1999, 51, 1077–1082. [Google Scholar] [CrossRef]
- Fattorusso, E.; Lanzotti, V.; Taglialatela-Scafati, O.; Di Rosa, M.; Ianaro, A. Cytotoxic saponins from bulbs of Allium porrum L. J. Agric. Food Chem. 2000, 48, 3455–3462. [Google Scholar] [CrossRef] [PubMed]
- Fattorusso, E.; Lanzotti, V.; Magno, S.; Taglialatela-Scafati, O. Sapogenins of Allium porrum L. J. Agric. Food Chem. 1998, 46, 4904–4908. [Google Scholar] [CrossRef]
- Gvazava, L.N.; Skhirtladze, A.V. Steroidal saponin from Allium porrum. Chem. Nat. Comp. 2017, 53, 1093–1095. [Google Scholar] [CrossRef]
- Gvazava, L.N.; Skhirtladze, A.V. Steroidal glycoside from Allium porrum. Chem. Nat. Comp. 2018, 54, 487–489. [Google Scholar] [CrossRef]
- Maisashvili, M.R.; Kuchukhidze, D.K.; Kikoladze, V.S.; Gvazava, L.N. Steroidal glycosides of gitogenin from Allium rotundum. Chem. Nat. Comp. 2012, 48, 86–90. [Google Scholar] [CrossRef]
- Tolkacheva, N.V.; Shashkov, A.S.; Chirva, V.Y. Steroidal glycosides from Allium cyrillii bulbs. Chem. Nat. Comp. 2012, 48, 272–275. [Google Scholar] [CrossRef]
- Mimaki, Y.; Kuroda, M.; Fukasawa, T.; Sashida, Y. Steroidal glycosides from bulbs of Allium jesdianum. J. Nat. Prod. 1999, 62, 194–197. [Google Scholar] [CrossRef]
- Kawashima, K.; Minaki, Y.; Sashida, Y. Steroidal saponins from the bulbs of Allium schubertii. Phytochemistry 1993, 32, 1267–1272. [Google Scholar] [CrossRef]
- Kravets, S.D.; Vollerner, Y.S.; Gorovits, M.B.; Abubakirov, N.K. Steroids of the spirostan and furostan series from plants of the genus Allium. Khim. Prir. Soed. 1990, 26, 359–373. [Google Scholar]
- Selim, S.; Al Jaouni, S. Anticancer and apoptotic effects on cell proliferation of diosgenin isolated from Costus speciosus (Koen.) Sm. BMC Complemet. Altern. Med. 2015, 15, 301. [Google Scholar] [CrossRef] [Green Version]
- Carotenuto, A.; Fattorusso, E.; Lanzotti, V.; Magno, S.; De Feo, V.; Carnuccio, R.; D’Acquisto, F. Porrigenins A and B, Novel Cytotoxic and Antiproliferative Sapogenins Isolated from Allium porrum. J. Nat. Prod. 1997, 60, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Rezgui, A.; Mitaine-Offer, A.C.; Paululat, T.; Delemasure, S.; Dutartre, P.; Lacaille-Dubois, M.-A. Cytotoxic steroidal glycosides from Allium flavum. Fitoterapia 2014, 93, 121–125. [Google Scholar] [CrossRef]
- Lafont, R.; Harmatha, J.; Marion-Poll, F.; Dinan, L.; Wilson, I.D. The Ecdysone Handbook, 3rd ed.; 2002; Continuously Updated; Available online: http://ecdybase.org/ (accessed on 20 September 2021).
- Rodrigues Adao, C.; Pereira da Silva, B.; Wanderley Tinoco, L.; Paz Parente, J. Haemolytic Activity and Immunological Adjuvant Effect of a New Steroidal Saponin from Allium ampeloprasum var. porrum. Chem. Biodivers. 2012, 9, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Rafieian-Kopaei, M. New concepts in nutraceuticals as alternative for pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487–1499. [Google Scholar]
- De Geyter, E.; Lambert, E.; Geelen, D.; Smagghe, G. Novel advances with plant saponins as natural insecticides to control pest insects. Pest Technol. 2007, 1, 96–105. [Google Scholar]
- Chaieb, I. Saponins as Insecticides: A Review. Tunisian J. Plant Protect. 2010, 5, 39–50. [Google Scholar]
- Singh, B.; Kaur, A. Control of insect pests in crop plants and stored food grains using plant saponins: A review. Food Sci. Technol. 2018, 87, 93–101. [Google Scholar] [CrossRef]
- Barile, E.; Bonanomi, G.; Antignani, V.; Zolfaghari, B.; Sajjadi, S.E.; Scala, F.; Lanzotti, V. Saponins from Allium minutiflorum with antifungal activity. Phytochemistry 2007, 68, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Lanzotti, V.; Barile, E.; Antignani, V.; Bonanomi, G.; Scala, F. Antifungal saponins from bulbs of garlic, Allium sativum L. var. Voghiera. Phytochemistry 2012, 78, 126–134. [Google Scholar] [CrossRef]
- Mostafa, A.; Sudisha, J.; El-Sayed, M.; Ito, S.; Ikeda, T.; Yamauchi, N.; Shigyo, M. Aginoside saponin, a potent antifungal compound, and secondary metabolite analyses from Allium nigrum L. Phytochem. Lett. 2013, 6, 274–280. [Google Scholar] [CrossRef]
- Sparg, S.G.; Light, M.E.; van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharm. 2004, 94, 219–243. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Xiang, L.; Huang, W.; He, X. Spirostanol saponins from Chinese onion (Allium chinense) exert pronounced anti-inflammatory and anti-proliferative activities. J. Funcion. Foods 2016, 25, 208–219. [Google Scholar] [CrossRef]
- Lacaille-Dubois, M.-A. Bioactive saponins with cancer related and immunomodulatory activity: Recent developments. Stud. Nat. Prod. Chem. (Part L) 2005, 32, 209–246. [Google Scholar]
- Jabrane, A.; Ben Jannet, H.; Miyamoto, T.; Mirjolet, J.-F.; Duchamp, O.; Harzallah-Skhiri, F.; Lacaille-Dubois, M.A. Spirostane and cholestane glycosides from the bulbs of Allium nigrum L. Food Chem. 2011, 125, 447–455. [Google Scholar] [CrossRef]
- Upadhyay, R.K. Nutritional and therapeutic potential of Allium vegetables. J. Nutr. Therap. 2017, 6, 18–37. [Google Scholar] [CrossRef]
- Dawid, C.; Weber, D.; Musiol, E.; Janas, V.; Baur, S.; Lang, R.; Fromme, T. Comparative assessment of purified saponins as permeabilization agents during respiratory. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148251. [Google Scholar] [CrossRef] [PubMed]
- Karassina, N.; Hofsteen, P.; Cali, J.J.; Vidugiriene, J. Time- and dose-depentendt toxicity studies in 3D cultures using a luminiscent lactate dehydrogenase assay. Methods Mol. Biol. 2021, 2255, 77–86. [Google Scholar]
- Mskhiladze, L.; Legault, J.; Lavoie, S.; Mshvildadze, V.; Kuchukhidze, J.; Elias, R.; Pichette, A. Cytotoxic steroidal saponins from the flowers of Allium leucanthum. Molecules 2008, 13, 2925–2934. [Google Scholar] [CrossRef] [Green Version]
- Kintia, P.K. Chemistry and Biological Activity of Steroid Saponins from Moldavian Plants. In Saponins Used in Traditional and Modern Medicine. Advances in Experimental Medicine and Biology; Waller, G.R., Yamasaki, K., Eds.; Springer: Boston, MA, USA, 1996; Volume 404, pp. 309–334. [Google Scholar]
- Dinan, L.; Harmatha, J.; Lafont, R. Chromatographic procedures for the isolation of plant steroids. J. Chromatogr. A 2001, 935, 105–123. [Google Scholar] [CrossRef]
- Dinan, L.; Harmatha, J.; Lafont, R. HPLC of Steroids. In High Performance Liquid Chromatography in Phytochemical Analysis; Waksmundzka-Hajnos, M., Sherma, J., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2011; Volume 102, pp. 679–708. [Google Scholar]
- Vennemann, A.; Alessandrini, F.; Wiemann, M. Differential Effects of Surface-Functionalized Zirconium Oxide Nanoparticles on Alveolar Macrophages, Rat Lung, and a Mouse Allergy Model. Nanomaterials 2017, 7, 280. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.K.; Rather, M.A.; Jha, A.K.; Shashank, A.; Singhal, S.; Sharma, M.; Pathak, U.; Sharma, D.; Mastinu, A. Artocarpus lakoocha Roxb. and Artocarpus heterophyllus Lam. Flowers: New Sources of Bioactive Compounds. Plants 2020, 9, 1329. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Cytotoxicity CC50 (µM) (95% Limits of Confidence) | Nitric Oxide, IC50 (µM) (95% Limits of Confidence) |
|---|---|---|
| 1. aginoside | 11.13 (2.76–44.91) | 7.84 (2.16–28.51) |
| 2. 6-deoxy-aginoside | 5.70 (1.83–17.71) | 5.21 (1.17–23.14) |
| 3. yayoisaponin A (alliporin) | 7.21 (1.64–31.47) | 5.62 (1.59–19.24) |
| 4. agigenin | >100 | >100 |
| 5. digitonin | 5.83 (1.72–19.75) | 5.52 (1.57–19.46) |
| 6. tomatonin | 9.90 (2.80–35.03) | 7.19 (2.99–17.29) |
| 7. diosgenin | >100 | >100 |
| Position | Aginoside (1) | 6-Deoxyaginoside (2) | Alliporin (3) | Agigenin (4) | ||||
|---|---|---|---|---|---|---|---|---|
| Aglycone | 13C | 1H | 13C | 1H | 13C | 1H | 13C | 1H |
| 1 | 46.94 | 1.22; 2.18 | 45.38 | 1.12; 2.15 | 46.89 | 1.20; 2.17 | 48.10 | 1.37; 2.28 |
| 2 | 70.36 | 4.07 | 70.28 | 3.93 | 70.35 | 4.07 | 73.21 | 4.19 |
| 3 | 84.36 | 4.01 | 84.01 | 3.85 | 84.42 | 4.00 | 77.33 | 3.97 |
| 4 | 31.72 | 2.12; 2.36 | 33.83 | 1.43; 1.82 | 31.60 | 2.12; 2.36 | 35.10 | 2.10; 2.53 |
| 5 | 47.64 | 1.13 | 44.38 | 0.96 | 47.60 | 1.12 | 48.49 | 1.40 |
| 6 | 69.80 | 3.96 | 27.90 | 0.98; 1.11 | 69.79 | 3.95 | 70.31 | 4.09 |
| 7 | 40.67 | 1.13; 1.99 | 31.93 | b ; 1.45 | 40.65 | 1.12; 1.98 | 40.88 | 1.21; 2.04 |
| 8 | 29.83 | 2.14 | 34.37 | 1.33 | 29.80 | 2.13 | 30.08 | 2.23 |
| 9 | 54.32 | 0.71 | 54.15 | 0.53 | 54.30 | 0.69 | 54.75 | 0.83 |
| 10 | 36.84 | -- | 36.67 | -- | 36.81 | -- | 37.71 | -- |
| 11 | 21.16 | 1.35; 1.52 | 21.23 | b ; 1.40 | 21.16 | 1.35; 1.50 | 21.46 | 1.47; 1.64 |
| 12 | 39.90 | 1.04; 1.65 | 39.86 | 0.96; 1.59 | 39.90 | 1.05; 1.64 | 40.20 | 1.14; 1.71 |
| 13 | 40.50 | -- | 40.59 | -- | 40.47 | -- | 40.93 | -- |
| 14 | 56.03 | 1.10 | 56.13 | 0.98 | 56.02 | 1.08 | 56.29 | 1.16 |
| 15 | 32.04 | 1.40; 2.05 | 32.00 | 1.36; 1.98 | 32.03 | 1.40; 2.05 | 32.24 | 1.44; 2.10 |
| 16 | 80.95 | 4.55 | 80.98 | 4.53 | 80.94 | 4.53 | 81.11 | 4.56 |
| 17 | 62.82 | 1.81 | 62.79 | 1.76 | 62.79 | 1.80 | 63.04 | 1.85 |
| 18 | 16.40 | 0.82 | 16.42 | 0.77 | 16.38 | 0.81 | 16.62 | 0.87 |
| 19 | 17.01 | 1.25 | 13.22 | 0.66 | 16.98 | 1.24 | 17.55 | 1.44 |
| 20 | 41.82 | 1.91 | 41.80 | 1.91 | 41.79 | 1.90 | 41.99 | 1.95 |
| 21 | 14.83 | 1.12 | 14.83 | 1.10 | 14.82 | 1.10 | 15.03 | 1.14 |
| 22 | 109.09 | -- | 109.12 | -- | 109.08 | -- | 109.21 | -- |
| 23 | 31.58 | 1.56; 1.63 | 31.62 | 1.63 (2H) | 31.55 | 1.57; 1.63 | 31.74 | 1.60; 1.67 |
| 24 | 29.04 | 1.24; 1.54 | 29.08 | 1.23; 1.53 | 29.01 | 1.50 (2H) | 29.22 | 1.53 (2H) |
| 25 | 30.39 | 1.54 | 30.41 | 1.54 | 30.37 | 1.52 | 30.58 | 1.55 |
| 26 | 66.68 | 3.47; 3.57 | 66.70 | 3.48; 3.57 | 66.66 | 3.45; 3.55 | 66.83 | 3.49; 3.57 |
| 27 | 17.12 | 0.66 | 17.15 | 0.67 | 17.11 | 0.64 | 17.30 | 0.67 |
| Gal (I) | ||||||||
| 1 | 102.92 | 4.96 | 103.08 | 4.90 | 102.87 | 4.94 | ||
| 2 | 72.35 | 4.54 | 72.41 | 4.51 | 72.34 | 4.50 | ||
| 3 | 75.57 | 4.03 | 75.58 | 4.03 | 75.54 | 4.02 | ||
| 4 | 79.22 | 4.58 | 79.23 | 4.57 | 78.99 | 4.56 | ||
| 5 | 75.36 | 4.12 | 75.35 | 4.11 | 75.39 | 4.12 | ||
| 6 | 60.51 | 4.21; 4.59 | 60.50 | 4.20; 4.59 | 60.55 | 4.20; 4.57 | ||
| Glc (II) | ||||||||
| 1 | 104.45 | 5.19 | 104.52 | 5.18 | 104.66 | 5.13 | ||
| 2 | 81.05 | 4.33 | 81.06 | 4.33 | 80.94 | 4.53 | ||
| 3 | 86.83 | 4.11 | 86.81 | 4.11 | 86.89 | 4.06 | ||
| 4 | 70.19 | 3.77 | 70.22 | 3.78 | 70.11 | 3.76 | ||
| 5 | 77.40 | 3.83 | 77.42 | 3.82 | 77.33 | 3.80 | ||
| 6 | 62.72 | 4.04; 4.47 | 62.74 | 4.03; 4.47 | 62.65 | 4.04; 4.45 | ||
| Xyl (III) | ||||||||
| 1 | 104.76 | 5.23 | 104.78 | 5.23 | 104.11 | 5.16 | ||
| 2 | 74.94 | 3.94 | 74.96 | 3.94 | 74.99 | 4.05 | ||
| 3 | 78.27 | 3.90 | 78.30 | 3.90 | 78.23 | 3.84 | ||
| 4 | 70.60 | 4.10 | 70.62 | 4.09 | 70.47 | 4.07 | ||
| 5 | 67.12 | 3.65; 4.20 | 67.14 | 3.64; 4.20 | 67.01 | 3.62; 4.17 | ||
| Glc (IV) | ||||||||
| 1 | 104.59 | 5.57 | 104.62 | 5.57 | 103.68 | 5.57 | ||
| 2 | 75.87 | 4.04 | 75.92 | 4.03 | 75.28 | 3.92 | ||
| 3 | 77.98 | 4.14 | 77.98 | 4.13 | 87.70 | 4.10 | ||
| 4 | 71.20 | 4.04 | 71.20 | 4.08 | 69.54 | 3.86 | ||
| 5 | 78.50 | 4.07 | 78.52 | 4.07 | 77.83 | 4.12 | ||
| 6 | 62.54 | 4.40; 4.53 | 62.53 | 4.39; 4.54 | 62.26 | 4.20; 4.45 | ||
| Glc (V) | ||||||||
| 1 | 105.22 | 5.08 | ||||||
| 2 | 77.56 | 3.80 | ||||||
| 3 | 78.19 | 4.05 | ||||||
| 4 | 71.33 | 4.09 | ||||||
| 5 | 80.39 | 4.28 | ||||||
| 6 | 62.20 | 4.25; 4.40 | ||||||
| Aglycone | Glycosidic Part | ||||
|---|---|---|---|---|---|
| Position | 13C | 1H | 13C | 1H | |
| 1 | 47.14 | H-1α: 0.93; H-1β: 1.905 | Gal (I) | ||
| 2 | 71.34 | 3.67 | 1 | 102.84 | 4.385 d (J = 7.8) |
| 3 | 85.02 | 3.56 | 2 | 72.86 | 3.72 dd (J = 9.7; 7.8) |
| 4 | 31.47 | H-4α: 1.755; H-4β: 1.84 | 3 | 75.50 | 3.55 dd (J = 9.7; 3.3) |
| 5 | 48.46 | 1.205 | 4 | 79.94 | 4.05 dd (J = 3.3; 1.0) |
| 6 | 71.52 | 3.805 | 5 | 75.67 | 3.56 ddd (7.4; 6.3; 1.0) |
| 7 | 40.71 | H-7α: 1.185; H-4β: 1.83 | 6 | 61.40 | 3.86 dd (J = 11.3; 7.4); 3.675 (J = 11.3; 6.3) |
| 8 | 30.74 | 1.95 | Glc (II) | ||
| 9 | 55.52 | 0.79 | 1 | 104.41 | 4.63 d (J = 7.6) |
| 10 | 37.77 | -- | 2 | 80.80 | 3.76 dd (J = 9.8; 7.6) |
| 11 | 22.13 | H-11α: 1.57; H-11β: 1.44 | 3 | 87.59 | 3.74 dd (J = 9.8; 8.2) |
| 12 | 41.02 | H-12α: 1.185; H-12β: 1.76 | 4 | 70.42 | 3.30 dd (J = 8.2; 9.8) |
| 13 | 41.79 | -- | 5 | 70.25 | 3.46 ddd (9.8; 7.0; 2.3) |
| 14 | 57.13 | 1.18 | 6 | 63.04 | 3.90 dd (J = 11.6; 2.3); 3.595 dd (J = 11.6; 7.0) |
| 15 | 32.70 | H-15α: 2.00; H-15β: 1.30 | Xyl (III) | ||
| 16 | 82.18 | 4.39 | 1 | 104.88 | 4.64 d (J = 7.7) |
| 17 | 63.83 | 1.76 | 2 | 75.31 | 3.26 dd (J = 9.6; 7.7) |
| 18 | 16.98 | 0.829 | 3 | 75.47 | 3.29 dd (J = 9.6; 8.0) |
| 19 | 17.24 | 1.073 | 4 | 70.96 | 3.53 ddd (J = 10.3; 8.0; 5.9) |
| 20 | 42.94 | 1.91 | 5 | 67.19 | 3.92 dd (J = 11.6; 5.9); 3.265 dd (11.6; 10.3) |
| 21 | 14.88 | 0.963 | Glc (IV) | ||
| 22 | 110.56 | -- | 1 | 103.70 | 5.01 d (J = 8.0) |
| 23 | 32.41 | H-23α: 1.70; H-23β: 1.57 | 2 | 75.01 | 3.42 dd (J = 9.4; 8.0) |
| 24 | 29.88 | H-24α: 1.43; H-24β: 1.63 | 3 | 87.66 | 3.58 dd (J = 9.4; 8.3) |
| 25 | 31.44 | 1.60 | 4 | 71.57 | 3.29 dd (J = 8.3; 9.7) |
| 26 | 67.85 | H-26α: 3.32; H-26β:3.45 | 5 | 78.10 | 3.34 ddd (J = 9.7; 6.3; 2.0) |
| 27 | 17.49 | 0.794 | 6 | 62.87 | 3.92 (J = 12.2; 2.0); 3.83 dd (J = 12.2; 6.3) |
| Glc (V) | |||||
| 1 | 105.21 | 4.57 d (J = 7.8) | |||
| 2 | 77.51 | 3.34 dd (J = 9.0; 7.8) | |||
| 3 | 77.78 | 3.40 t (J ~ 9.0; 9.0) | |||
| 4 | 77.96 | 3.345 dd (~9.0; 9.0) | |||
| 5 | 78.22 | 3.315 ddd (J ~9.0; 6.3; 2.3) | |||
| 6 | 62.63 | 3.89 dd (J = 11.8; 2.3); 3.645 dd (J = 11.8; 6.3) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harmatha, J.; Buděšínský, M.; Zídek, Z.; Kmoníčková, E. Spirostanol Saponins from Flowers of Allium Porrum and Related Compounds Indicating Cytotoxic Activity and Affecting Nitric Oxide Production Inhibitory Effect in Peritoneal Macrophages. Molecules 2021, 26, 6533. https://doi.org/10.3390/molecules26216533
Harmatha J, Buděšínský M, Zídek Z, Kmoníčková E. Spirostanol Saponins from Flowers of Allium Porrum and Related Compounds Indicating Cytotoxic Activity and Affecting Nitric Oxide Production Inhibitory Effect in Peritoneal Macrophages. Molecules. 2021; 26(21):6533. https://doi.org/10.3390/molecules26216533
Chicago/Turabian StyleHarmatha, Juraj, Miloš Buděšínský, Zdeněk Zídek, and Eva Kmoníčková. 2021. "Spirostanol Saponins from Flowers of Allium Porrum and Related Compounds Indicating Cytotoxic Activity and Affecting Nitric Oxide Production Inhibitory Effect in Peritoneal Macrophages" Molecules 26, no. 21: 6533. https://doi.org/10.3390/molecules26216533