In Vivo Reporters for Visualizing Alternative Splicing of Hormonal Genes


1
Laboratory of Hormonal Regulations in Plants, Institute of Experimental Botany, Czech Academy of Sciences, 165 02 Prague, Czech Republic
2
Functional Genomics and Proteomics of Plants, Central European Institute of Technology and National Centre for Biomolecular Research, Masaryk University, 625 00 Brno, Czech Republic
*
Author to whom correspondence should be addressed.
Plants 2020, 9(7), 868; https://doi.org/10.3390/plants9070868
Submission received: 10 June 2020
/
Revised: 2 July 2020
/
Accepted: 7 July 2020
/
Published: 8 July 2020
(This article belongs to the Special Issue Advances in Auxin Research)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Rapid progress in plant molecular biology in recent years has uncovered the main players in hormonal pathways and characterized transcriptomic networks associated with hormonal response. However, the role of RNA processing, in particular alternative splicing (AS), remains largely unexplored. Here, using example genes involved in cytokinin signaling, brassinosteroid synthesis and auxin transport, we present a set of reporters devised to visualize their AS events in vivo. These reporters show a differential tissue-specific expression of certain transcripts and reveal that expression of some of the them can be changed by the application of the exogenous hormone. Finally, based on the characterized AS event of the PIN7 auxin efflux carrier, we designed a system that allows a rapid genetic screening for the factors upstream of this AS event. Our innovative toolset can be therefore highly useful for exploring novel regulatory nodes of hormonal pathways and potentially helpful for plant researchers focusing on developmental aspects of AS.
Keywords:
alternative splicing; auxin; cytokinins; brassinosteroids; AHP6; DWF4; PIN7; D-amino acid oxidase; genetic screen; Arabidopsis1. Introduction
Advanced sequencing has revealed a stunning complexity of eukaryotic (and plant) transcriptomes, shaped by alternative splicing (AS). In contrast to the increasing depth and refinement of the transcriptomic approaches, very few individual AS events have been experimentally characterized in detail. In multicellular organisms, a crucial feature of the AS events is the temporal and tissue-specific extent of their expression. Fluorescent AS sensors represent a powerful tool that allows for AS detection in vivo and in situ. These reporters are widely used in animal systems [1,2,3,4] and are also helpful in deciphering the upstream stimuli modifying the transcript levels inside cells [2,3,5,6,7]. Nonetheless, their usage in plant systems appears to be rare [8,9,10].
Phytohormones integrate developmental programs and environmental cues during the whole plant lifecycle. Although the key factors governing hormonal signaling, transport, and their direct internal levels have been deeply characterized, only a little work has been dedicated to the role of AS in these pathways. Though a few examples underlining the functional role of AS have been described for abscisic acid [11,12,13,14,15], knowledge about the involvement of AS in hormonal regulations of cytokinin, brassinosteroid, and auxin pathways remains sporadic [13,16,17,18,19,20].
Here, we reveal the advantage of fluorescent sensors directed for the visualization of various types of AS and illustrate their usage on several genes involved in phytohormonal processes. To this end, we examined the splice variants of the genes involved in cytokinin signaling (AHP6 [21]), brassinosteroid synthesis (DWF4 [22,23]), and auxin transport (PIN4 and PIN7 [24,25,26]), and investigated their expression in the Arabidopsis seedling root tip. We reveal that some of the splice isoforms are differentially expressed and their expression levels can be altered following hormonal treatment. Finally, based on the PIN7 AS reporter, we also present a system for an efficient forward genetic screening aimed for the identification of the regulators upstream of the PIN7 AS event.
2. Results
2.1. Reporters for Visualizing Common Types of AS Events in Plant Hormonal Genes
Previously, Mähönen and collaborators [21] found that AHP6, a negative regulator of cytokinin signaling, is processed into two splice variants. The amino acid sequence encoded by the canonical transcript (AHP6b) corresponds to that of a common AHP transducer of its class [21,27,28], while the minor AHP6a isoform lacks a large part of the first exon (Figure 1a). The weak ahp6-2 allele contains a mutation leading to a cryptic AS site inside the first intron, which leads to the exclusive expression of the alternative AHP6a transcript [21]. To visualize the expression of the AHP6a isoform, we fused the ahp6-2 sequence with the mVenus tag under the control of the native AHP6 promoter (Figure 1b). Wild type AHP6-mCherry, attributable to the more abundant canonical AHP6b transcript [21] (Figure 1c), is synthesized in the protoxylem founder cells and the adjacent pericycle and is actively distributed within the primary root stele [21,29]. In contrast, ahp6-2-mVenus appears to be retained at subcellular compartments within the AHP6 promoter [21] expression domain, and it fails to migrate from the protoxylem poles of the meristem stele (Figure 1d). Cytokinin treatment leads, similar to that of the AHP6:GFP promoter fusion [21], to the decrease in expression of the wild type DNA-based AHP6:AHP6-mCherry reporter (Figure 1c). An even stronger signal drop was seen for AHP6:ahp6-2-mVenus (Figure 1d). This collectively indicates that such AS reporters can also be used in plants. Moreover, AS could change the properties of the AHP6 protein, conceivably including its ability to act non-cell autonomously in the root meristem and one can also speculate whether AHP6 splice isoforms can be differentially regulated at the post-transcriptional level.
Detecting the expression of the minor AHP6a isoform in vivo, we turned to another type of hormonal AS event. We selected DWF4, an essential component of the brassinosteroid synthesis pathway [22]. Here, the alternative DWF4.2 variant contains a premature stop codon in the last intron retained, compared to the canonical DWF4.1 transcript (Figure 1e) [23]. To visualize a simultaneous expression of both splice variants, we inserted the GFP and RFP tags into coding regions prior to the termination codons of DWF4.1 and DWF4.2, respectively (Figure 1f). Adapting the usage from the animal field [1], we named the reporter D4.1G.2R (for DWF4.1-GFP + DWF4.2-RFP). We observed that the D4.1G expression is identifiable starting from the root tip transition zone and continuing to the proximal regions of the root, while the D4.2R signal prevails in the root meristem (Figure 1g). Although we were unable to distinguish, due to the low signal intensity, whether AS also changes the subcellular localization of the resulting protein, the patchy character of the D4.2R signal in the meristem suggests that it might be a short-living protein possibly linked to specific phases of the cell cycle [30]. The low stability of DWF4 was corroborated by the treatment with brassinolide (Figure 1g). This leads to a rapid decrease in the protein expression [31] and the decline in the fluorescence signal for both variants already after 4 h of treatment (Figure 1g). It suggests that AS can change the overall expression pattern of the DWF4 protein and that the in vivo reporters can be used in Arabidopsis for monitoring various types of AS, such as alternative donor site selection or intron retention.
2.2. Reporters Monitoring AS of Auxin Transporters PIN4 and PIN7
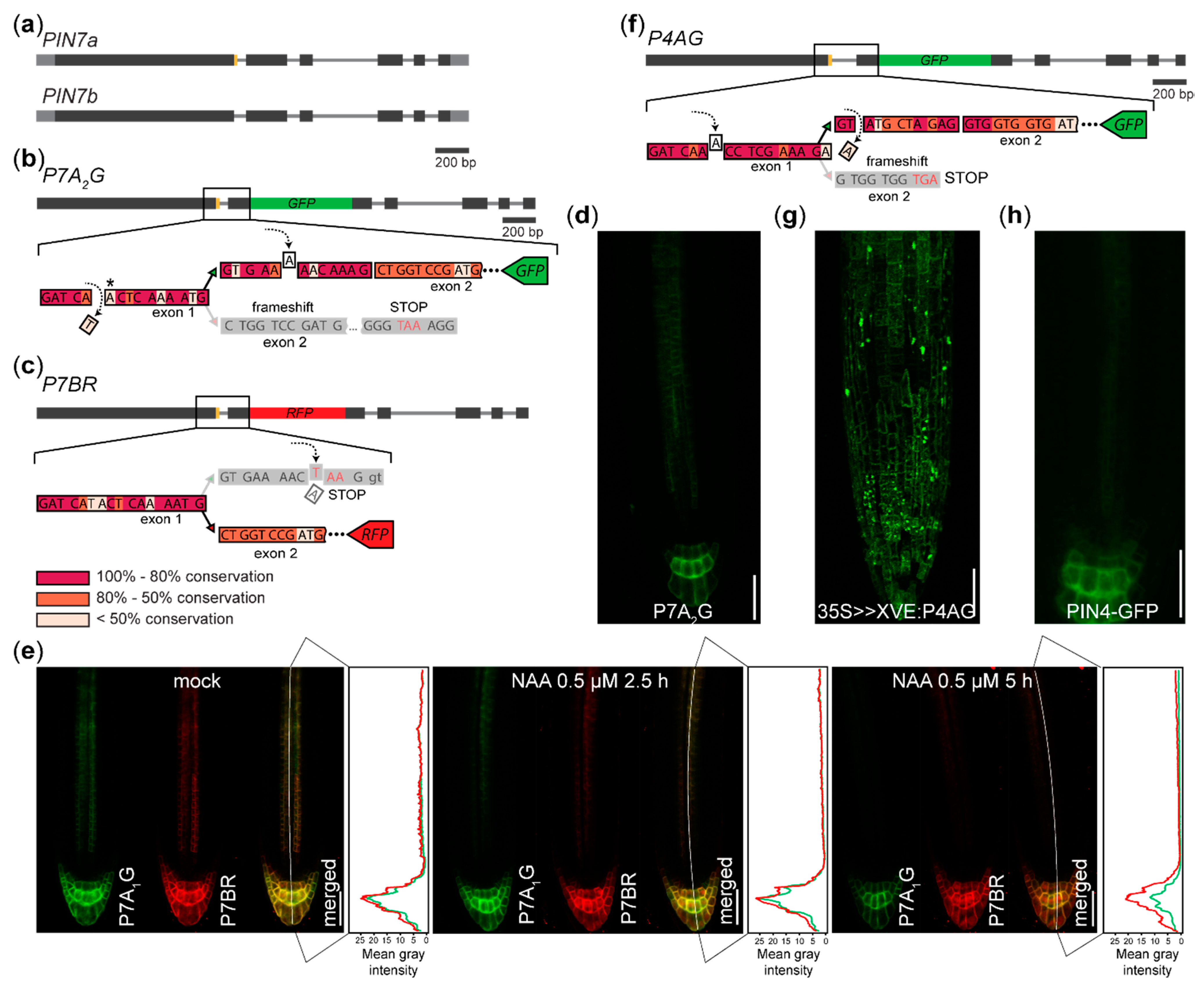
We also generated the P7A2G (for PIN7a-GFP, version 2) reporter for AS of the gene encoding an auxin efflux carrier PIN7 [16,24], designed as an alternative to the P7A1G sensor visualizing the AS of the PIN7a/b event reported earlier [10] (Figure 2a–c). We observed the expression pattern and the subcellular protein localization similar to that previously reported [10] (Figure 2d), but with a lower signal intensity (data not shown). It reveals that one can design several versions of the reporters, obtaining similar results.
We treated the described P7A1G x P7BR (for PIN7b-RFP) AS sensor [10] with the synthetic auxin 1-naphthylacetic acid (NAA). We observed that the signal in the stele decreased after 2.5 h [32] for both PIN7 variants, while the signal in the columella persisted at nearly similar levels for P7BR, in contrast to a noticeable drop of P7A1G intensity after 5 h (Figure 2e). This suggests that hormonal treatment can differentially change the expression of the splice isoforms and that auxin can directly or indirectly affect gene expression also at the post-transcriptional level.
The fluorescence was also seen in the case of the P4AG (PIN4a-GFP) reporter (Figure 2f), whose expression was controlled by the β-estradiol inducible 35S promoter [33]. Here, the P4AG signal was also present in the cells outside the native PIN4 expression domain (Figure 2g,h). These reporters can therefore also be used to test whether the expression of the splice variant is tissue-specifically predetermined.
2.3. A System for Genetic Screening for the Factors Upstream of the PIN7 AS Event
Although a few upstream regulators of some AS events have been identified in plants [12,13,15], no systematic effort has been made in order to explore such regulators using a classical forward genetic methodology. To design a genetic screening system for the regulators of the PIN7a/b AS event, we considered some of the previously proposed positive or negative selection markers, e.g., genes coding for barnase, diphtheria toxin, RNAse T1 [34,35], phosphonate monoesterase (pehA) [34,36] or D-amino acid oxidase (DAAO) [34,37,38]. DAAO, in particular, turned out to be suitable for designing such system, as the external application of the substrate can conditionally control the expected lethality of the selection marker [36,37]. We modified the P7A1G sensor [10] by replacing the GFP tag with DAAO [37] and generated stable lines carrying this transgene (Figure 3a).
The lines containing the DAAO construct termed P7A1D (for PIN7a-DAAO), which were indistinguishable from the wild type on the control media (Figure 3b). However, the application of D-isoleucine [37] had a dramatic impact on the seedling viability in these genotypes (Figure 3c). These experiments thus suggest that the DAAO gene can be used, along with fluorescent reporters, as a basis for a rapid genetic screen aimed for the identification of positive regulators of the PIN7 AS event.
3. Discussion
Here we reveal that various types of AS can be visualized in vivo and in situ in plants. As the AS reporters provide fine spatiotemporal information about the splice isoform expression, they can be highly useful in situations when quantitative (e.g., qRT-PCR) or qualitative (e.g., RNA hybridization in situ) approaches could not be reliably used. In extraordinary cases, such reporters can also extend the information delivered by the standard gene expression reporters, such as genomic DNA-based translational fusions with the reporter tag expressed only inside a subset of splice variants.
This system has certain limits, however, and a careful interpretation of isoform expression is desirable. For example, from the public transcriptomic databases, the native DWF4.2 transcript levels in the root appear to be significantly lower than that of DWF4.1 [23,39,40]. It can be, on the one hand, explained by the high ratio of mature versus meristematic cells present in the original RNA-sequencing experiments. On the other hand, inserting the region coding for the fluorescent tag can interfere with putative exonic elements required for the regulation of the AS event and lead to isoform expression artifact. Where possible, considering evolutional conservation is therefore highly useful, as demonstrated by both versions of the PIN7a reporter (P7A1G and P7A2G, respectively, Figure 3b), which show similar expression patterns in the root tip. This technique is, of course, limited in use only for monitoring protein-coding regions and practically not suitable, e.g., for assessing alternative polyadenylation or AS of non-coding RNAs.
In vivo sensors are commonly used in animal systems to demonstrate the developmental impact of AS. They have been used for the detection of tissue-specific expression of AS events e.g., in C. elegans [1,2,41], D. melanogaster [7], mouse [4,42] and human [3,5,6]. Moreover, these systems have also been instrumental in identifying the factors upstream of some of these AS events [2,3,5,6,7]. The usage of AS reporters in plant model organisms is scarce and has practically never been purposed for assessing cell-specific expression of splice isoforms [9,43,44]. To our knowledge, genetic screening with the marker, such as DAAO, has never been used in plants for such purpose at all. The fluorescent reporters can be used in the forward genetic screens as well. Exemplifying P7A1D, we even provide a more effective tool for the rapid and effective isolation of mutants with defects in this AS event.
An increasing number of pieces of evidence show that AS might be an important (and largely neglected) regulator of hormonal pathways, specifically in auxin-dependent processes [10,16,18,19,20]. The differential PIN7 AS reporter expression in columella following auxin treatment brings another interesting hint into these schemes. Altogether, our technological improvements can be therefore highly useful for plant biologists studying the post-transcriptional molecular mechanisms in hormonal and developmental pathways.
4. Materials and Methods
4.1. Plant Growth Conditions and Microscopy
All plant material was in Arabidopsis thaliana (L.) Heynh., Col-0 ecotype. Usually, the seeds were surface sterilized and, after 2 days of stratification in 4°C, cultivated under a 16 h:8 h photoperiod, 22:18°C, light:dark, on the 0.5 × MS medium [45] with 1% sucrose. For the reporter gene analyses, primary roots of 4–6 day-old vertically grown seedlings were used. Inducible transgene expression was controlled by the cultivation of the seedlings for six days (along with appropriate controls) on sterile media containing 5 µM 17-β-estradiol (est). The following chemicals were used for treatments: N6-benzyladenine (BA, diluted as 0.2 µM in DMSO), brassinolide (BL, 0.1 µM in DMSO), 1-naphthylacetic acid (NAA, 0.5 mM in DMSO), 17-β-estradiol (5 mM in DMSO), D-isoleucine (1 M in water), all from Sigma (Sigma-Aldrich, St. Louis, MO, USA). An equivalent amount of solvent was added to the control media.
The hormonal treatments were done under indicated conditions in the liquid 0.5 × MS medium supplemented with the appropriate compound and followed by the imaging. For examining the lines containing the DAAO transgenes, the seedlings were grown on the solid 0.5 × MS media supplemented with 30 mM D-isoleucine for 14–18 days and documented.
Confocal microscopy was conducted on inverted Zeiss Axio Observer.Z1 equipped with the standard confocal LSM880 and Airyscan modules with the 20×/0.8 DIC M27 air and 40×/1.2 W Kor FCS M27 air. For each experiment, 8 to 20 roots were analyzed in two independent biological replicates.
4.2. DNA Manipulations and Transgenic Work
The AHP6:ahp6-2-mVenus and AHP6:AHP6-mCherry constructs were made using the Multisite Gateway procedure (Invitrogen, Life Technologies, Carlsbad, CA, USA), inserted into the pHm43GW,0 vector as described [46,47], respecting the design of the original translational fusion [21]. The D4.1G.2R reporter was made using the Gibson Assembly method [48] (New England Biolabs, Ipswich, MA, USA) by recombination of the 2032-nucleotide sequence upstream of the start codon to the last coding codon of the isoform DWF4.2 (1), RFP with a stop codon (2) and the last intron plus last exon of the DWF4.1 isoform (3), all inserted into the pMDC83 [33] backbone containing GFP. PIN7A2G was made by the analogous procedure, as described previously [10]. Cloning of the P4AG reporter was done as for PIN7 [10], the entry pDONR202 vector containing the genomic DNA-based PIN4-GFP [49] construct was modified by inverse PCR and recombined in the LR Gateway reaction with the pMDC7 destination vector [33]. The P7A1D coding polynucleotides were custom synthesized (Gen9, Ginkgo Bioworks, Boston, MA, USA), cloned into the pDONR221 P5-P2 vector and together with the PIN7 promoter [10] in the pDONR221 P1-P5r vector recombined with the pH7WG destination vector [47] in the Multisite Gateway LR reaction. The validated binary constructs were transformed into Arabidopsis by floral dipping and T2 or T3 generation of transformants showing the respective signal examined.
Author Contributions
Conceptualization, K.R., I.K. and K.T.; methodology, K.R., I.K., K.T. and E.K.; validation, I.K., K.T. and E.K.; investigation, I.K., K.T. and E.K.; writing—original draft preparation, K.R.; writing—review and editing, K.R., I.K., and K.T.; visualization, I.K. and K.T.; supervision, K.R.; project administration, K.R.; funding acquisition, K.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Czech Science Foundation (16-26428S) to I. K., K. T. and K. R, and the Ministry of Education, Youth and Sports of the Czech Republic (MEYS, CZ.02.1.01/0.0/0.0/16_019/0000738 and LTC18073) to K. R. Imaging Facility of the Institute of Experimental Botany is supported by MEYS (projects LM2018129—Czech BioImaging, ERDF CZ.02.1.01/0.0/0.0/16_013/0001775 and OPPK CZ.2.16/3.1.00/21519).
Acknowledgments
We thank Jozef Mravec for suggesting initial concepts, the members of Jan Hejátko lab for support during the early phases of the project and Elena Zemlyanskaya for help with editing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Kuroyanagi, H.; Kobayashi, T.; Mitani, S.; Hagiwara, M. Transgenic alternative-splicing reporters reveal tissue-specific expression profiles and regulation mechanisms in vivo. Nat. Methods 2006, 3, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Norris, A.D.; Gao, S.; Norris, M.L.; Ray, D.; Ramani, A.K.; Fraser, A.G.; Morris, Q.; Hughes, T.R.; Zhen, M.; Calarco, J.A. A pair of RNA-binding proteins controls networks of splicing events contributing to specialization of neural cell types. Mol. Cell 2014, 54, 946–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Damoiseaux, R.; Chen, L.; Black, D.L. A broadly applicable high-throughput screening strategy identifies new regulators of Dlg4 (Psd-95) alternative splicing. Genome Res. 2013, 23, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Bonano, V.I.; Oltean, S.; Garcia-Blanco, M.A. A protocol for imaging alternative splicing regulation in vivo using fluorescence reporters in transgenic mice. Nat. Protoc. 2007, 2, 2166–2181. [Google Scholar] [CrossRef] [PubMed]
- Stoilov, P.; Lin, C.-H.; Damoiseaux, R.; Nikolic, J.; Black, D.L. A high-throughput screening strategy identifies cardiotonic steroids as alternative splicing modulators. PNAS 2008, 105, 11218–11223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, M.J.; Wang, Q.; Kennedy, C.J.; Silver, P.A. An Alternative Splicing Network Links Cell-Cycle Control to Apoptosis. Cell 2010, 142, 625–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.S.S.; Millard, S.S. Deterministic splicing of Dscam2 is regulated by Muscleblind. Sci. Adv. 2019, 5, eaav1678. [Google Scholar] [CrossRef] [Green Version]
- Göhring, J.; Jacak, J.; Barta, A. Imaging of Endogenous Messenger RNA Splice Variants in Living Cells Reveals Nuclear Retention of Transcripts Inaccessible to Nonsense-Mediated Decay in Arabidopsis. Plant Cell 2014, 26, 754–764. [Google Scholar] [CrossRef] [Green Version]
- Ushijima, T.; Hanada, K.; Gotoh, E.; Yamori, W.; Kodama, Y.; Tanaka, H.; Kusano, M.; Fukushima, A.; Tokizawa, M.; Yamamoto, Y.Y.; et al. Light Controls Protein Localization through Phytochrome-Mediated Alternative Promoter Selection. Cell 2017, 171, 1316–1325.e12. [Google Scholar] [CrossRef]
- Kashkan, I.; Hrtyan, M.; Filepova, R.; Vondrakova, Z.; Hejatko, J.; Simon, S.; Rombaut, D.; Jacobs, T.B.; Frilander, M.J.; Friml, J.; et al. Mutually opposing activity of PIN7 splicing isoforms is required for auxin-mediated tropic responses in Arabidopsis thaliana. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wang, Z.; Ji, H.; Yuan, B.; Wang, S.; Su, C.; Yao, B.; Zhao, H.; Li, X. ABA signalling is fine-tuned by antagonistic HAB1 variants. Nat. Commun. 2015, 6, 8138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, X.; Qian, B.; Cao, F.; Wu, W.; Yang, L.; Guan, Q.; Gu, X.; Wang, P.; Okusolubo, T.A.; Dunn, S.L.; et al. An Arabidopsis PWI and RRM motif-containing protein is critical for pre-mRNA splicing and ABA responses. Nat. Commun. 2015, 6, 8139. [Google Scholar] [CrossRef] [Green Version]
- Szakonyi, D.; Duque, P. Alternative Splicing as a Regulator of Early Plant Development. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, R.F.; Carvalho, S.D.; Duque, P. The plant-specific SR45 protein negatively regulates glucose and ABA signaling during early seedling development in Arabidopsis. Plant Physiol. 2010, 154, 772–783. [Google Scholar] [CrossRef] [Green Version]
- Sugliani, M.; Brambilla, V.; Clerkx, E.J.M.; Koornneef, M.; Soppe, W.J.J. The conserved splicing factor SUA controls alternative splicing of the developmental regulator ABI3 in Arabidopsis. Plant Cell 2010, 22, 1936–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrtyan, M.; Šliková, E.; Hejátko, J.; Růžička, K. RNA processing in auxin and cytokinin pathways. J. Exp. Bot. 2015, 66, 4897–4912. [Google Scholar] [CrossRef] [Green Version]
- Fujikura, U.; Jing, R.; Hanada, A.; Takebayashi, Y.; Sakakibara, H.; Yamaguchi, S.; Kappel, C.; Lenhard, M. Variation in Splicing Efficiency Underlies Morphological Evolution in Capsella. Dev. Cell 2018, 44, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Kriechbaumer, V.; Wang, P.; Hawes, C.; Abell, B.M. Alternative splicing of the auxin biosynthesis gene YUCCA4 determines its subcellular compartmentation. Plant J. 2012, 70, 292–302. [Google Scholar] [CrossRef]
- Ghelli, R.; Brunetti, P.; Napoli, N.; Paolis, A.D.; Cecchetti, V.; Tsuge, T.; Serino, G.; Matsui, M.; Mele, G.; Rinaldi, G.; et al. A Newly Identified Flower-Specific Splice Variant of AUXIN RESPONSE FACTOR8 Regulates Stamen Elongation and Endothecium Lignification in Arabidopsis. Plant Cell 2018, 30, 620–637. [Google Scholar] [CrossRef] [Green Version]
- Remy, E.; Cabrito, T.R.; Baster, P.; Batista, R.A.; Teixeira, M.C.; Friml, J.; Sá-Correia, I.; Duque, P. A major facilitator superfamily transporter plays a dual role in polar auxin transport and drought stress tolerance in Arabidopsis. Plant Cell 2013, 25, 901–926. [Google Scholar] [CrossRef] [Green Version]
- Mähönen, A.P.; Bishopp, A.; Higuchi, M.; Nieminen, K.M.; Kinoshita, K.; Törmäkangas, K.; Ikeda, Y.; Oka, A.; Kakimoto, T.; Helariutta, Y. Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 2006, 311, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.; Dilkes, B.P.; Fujioka, S.; Takatsuto, S.; Sakurai, A.; Feldmann, K.A. The DWF4 Gene of Arabidopsis Encodes a Cytochrome P450 That Mediates Multiple 22α-Hydroxylation Steps in Brassinosteroid Biosynthesis. Plant Cell 1998, 10, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jürgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Benková, E.; Blilou, I.; Wisniewska, J.; Hamann, T.; Ljung, K.; Woody, S.; Sandberg, G.; Scheres, B.; Jürgens, G.; et al. AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 2002, 108, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Benková, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertová, D.; Jürgens, G.; Friml, J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Skerker, J.M.; Perchuk, B.S.; Siryaporn, A.; Lubin, E.A.; Ashenberg, O.; Goulian, M.; Laub, M.T. Rewiring the specificity of two-component signal transduction systems. Cell 2008, 133, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Mähönen, A.P.; Higuchi, M.; Törmäkangas, K.; Miyawaki, K.; Pischke, M.S.; Sussman, M.R.; Helariutta, Y.; Kakimoto, T. Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis. Curr. Biol. 2006, 16, 1116–1122. [Google Scholar] [CrossRef] [Green Version]
- Besnard, F.; Refahi, Y.; Morin, V.; Marteaux, B.; Brunoud, G.; Chambrier, P.; Rozier, F.; Mirabet, V.; Legrand, J.; Lainé, S.; et al. Cytokinin signalling inhibitory fields provide robustness to phyllotaxis. Nature 2014, 505, 417–421. [Google Scholar] [CrossRef]
- Doerner, P.; Jørgensen, J.-E.; You, R.; Steppuhn, J.; Lamb, C. Control of root growth and development by cyclin expression. Nature 1996, 380, 520–523. [Google Scholar] [CrossRef]
- Bancoş, S.; Nomura, T.; Sato, T.; Molnár, G.; Bishop, G.J.; Koncz, C.; Yokota, T.; Nagy, F.; Szekeres, M. Regulation of Transcript Levels of the Arabidopsis Cytochrome P450 Genes Involved in Brassinosteroid Biosynthesis. Plant Physiol. 2002, 130, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieten, A.; Vanneste, S.; Wisniewska, J.; Benková, E.; Benjamins, R.; Beeckman, T.; Luschnig, C.; Friml, J. Functional redundancy of PIN proteins is accompanied by auxin-dependent cross-regulation of PIN expression. Development 2005, 132, 4521–4531. [Google Scholar] [CrossRef] [Green Version]
- Curtis, M.D.; Grossniklaus, U. A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol. 2003, 133, 462–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempe, K.; Gils, M. Pollination control technologies for hybrid breeding. Mol. Breed. 2011, 27, 417–437. [Google Scholar] [CrossRef]
- Debeaujon, I.; Nesi, N.; Perez, P.; Devic, M.; Grandjean, O.; Caboche, M.; Lepiniec, L. Proanthocyanidin-accumulating cells in Arabidopsis testa: Regulation of differentiation and role in seed development. Plant Cell 2003, 15, 2514–2531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dotson, S.B.; Lanahan, M.B.; Smith, A.G.; Kishore, G.M. A phosphonate monoester hydrolase from Burkholderia caryophilli PG2982 is useful as a conditional lethal gene in plants. Plant J. 1996, 10, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Erikson, O.; Hertzberg, M.; Näsholm, T. A conditional marker gene allowing both positive and negative selection in plants. Nat. Biotechnol. 2004, 22, 455–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollegioni, L.; Molla, G. New biotech applications from evolved D-amino acid oxidases. Trends Biotechnol. 2011, 29, 276–283. [Google Scholar] [CrossRef]
- Li, S.; Yamada, M.; Han, X.; Ohler, U.; Benfey, P.N. High-Resolution Expression Map of the Arabidopsis Root Reveals Alternative Splicing and lincRNA Regulation. Dev. Cell 2016, 39, 508–522. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Calixto, C.P.G.; Marquez, Y.; Venhuizen, P.; Tzioutziou, N.A.; Guo, W.; Spensley, M.; Entizne, J.C.; Lewandowska, D.; ten Have, S.; et al. A high quality Arabidopsis transcriptome for accurate transcript-level analysis of alternative splicing. Nucleic Acids Res. 2017. [Google Scholar] [CrossRef]
- Kuroyanagi, H.; Ohno, G.; Mitani, S.; Hagiwara, M. The Fox-1 family and SUP-12 coordinately regulate tissue-specific alternative splicing in vivo. Mol. Cell. Biol. 2007, 27, 8612–8621. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, A.; Hosokawa, M.; Nojima, T.; Hagiwara, M. Splicing Reporter Mice Revealed the Evolutionally Conserved Switching Mechanism of Tissue-Specific Alternative Exon Selection. PLoS ONE 2010, 5, e10946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanno, T.; Lin, W.-D.; Fu, J.L.; Chang, C.-L.; Matzke, A.J.M.; Matzke, M. A Genetic Screen for Pre-mRNA Splicing Mutants of Arabidopsis thaliana Identifies Putative U1 snRNP Components RBM25 and PRP39a. Genetics 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanno, T.; Venhuizen, P.; Wen, T.-N.; Lin, W.-D.; Chiou, P.; Kalyna, M.; Matzke, A.J.M.; Matzke, M. PRP4KA, a Putative Spliceosomal Protein Kinase, Is Important for Alternative Splicing and Development in Arabidopsis thaliana. Genetics 2018, 210, 1267–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Siligato, R.; Wang, X.; Yadav, S.R.; Lehesranta, S.; Ma, G.; Ursache, R.; Sevilem, I.; Zhang, J.; Gorte, M.; Prasad, K.; et al. MultiSite Gateway-Compatible Cell Type-Specific Gene-Inducible System for Plants. Plant Physiol. 2016, 170, 627–641. [Google Scholar] [CrossRef]
- Karimi, M.; Bleys, A.; Vanderhaeghen, R.; Hilson, P. Building Blocks for Plant Gene Assembly. Plant Physiol. 2007, 145, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
Figure 1.
Reporters visualizing AS of AHP6 and DWF4 in the root tip: (a) Diagram of the coding regions of the AHP6 gene. (b) Scheme of the ahp6-2-mVenus reporter for visualizing the AHP6a-mVenus isoform. (c) AHP6:AHP6-mCherry line, containing the genomic AHP6 sequence, shows that the AHP6 protein is distributed throughout the root tip stele [21] (left); the treatment with the synthetic cytokinin benzyladenine (BA) decreases the expression of the wild type AHP6:AHP6-mCherry. Note the predominantly nuclear presence of the AHP6-mCherry foci (inset). (d) AHP6:ahp6-2-mVenus is present only in the protoxylem founder cells and neighboring pericycle (left); following the treatment with BA, the reporter expression decreases even more notably. Inset: a perinuclear cytoplasmic or endocompartment localization of ahp6-2-mVenus. (e) Diagram of the coding regions of the DWF4 gene. (f) Scheme of the D4.1G.2R reporter visualizing the DWF4.1 and DWF4.2 expression. (g) Following the treatment with brassinolide (BL), the fluorescence signal drops below the detection limit of the microscope. Bars, 100 µm, on (c), (d) and (g). Black—coding regions, grey—UTRs, thin grey line—introns, yellow—regions changed by AS, red—RFP, green—GFP, on (a), (b), (e) and (f).
Figure 1.
Reporters visualizing AS of AHP6 and DWF4 in the root tip: (a) Diagram of the coding regions of the AHP6 gene. (b) Scheme of the ahp6-2-mVenus reporter for visualizing the AHP6a-mVenus isoform. (c) AHP6:AHP6-mCherry line, containing the genomic AHP6 sequence, shows that the AHP6 protein is distributed throughout the root tip stele [21] (left); the treatment with the synthetic cytokinin benzyladenine (BA) decreases the expression of the wild type AHP6:AHP6-mCherry. Note the predominantly nuclear presence of the AHP6-mCherry foci (inset). (d) AHP6:ahp6-2-mVenus is present only in the protoxylem founder cells and neighboring pericycle (left); following the treatment with BA, the reporter expression decreases even more notably. Inset: a perinuclear cytoplasmic or endocompartment localization of ahp6-2-mVenus. (e) Diagram of the coding regions of the DWF4 gene. (f) Scheme of the D4.1G.2R reporter visualizing the DWF4.1 and DWF4.2 expression. (g) Following the treatment with brassinolide (BL), the fluorescence signal drops below the detection limit of the microscope. Bars, 100 µm, on (c), (d) and (g). Black—coding regions, grey—UTRs, thin grey line—introns, yellow—regions changed by AS, red—RFP, green—GFP, on (a), (b), (e) and (f).

Figure 2.
Reporters visualizing AS of PIN7 and PIN4 in the root tip: (a) Diagram of the coding regions of the PIN7 gene. The canonical PIN7b isoform differs from PIN7a in the presence of the 12 nucleotide region (orange). (b) Scheme of the P7A2G reporter for visualizing the PIN7a isoform. Removing the nucleotide prior to the alternative splice site leads to a frameshift in the PIN7b transcript. The frame is restored by insertion of the nucleotide into the region exclusively encoded in the P7A2G sequence following the alternative splice site. Asterisk: a neighboring A nucleotide is removed in the P7A1G sensor [10] (c) The P7BR reporter, as outlined in [10]. The visualization of P7BR is accomplished by introducing the termination codon into the protruding region encoded entirely by PIN7a. P7AG and P7BR are encoded by two constructs that are separately transformed and then crossed. (d) Expression pattern of the P7A2G reporter. (e) Following the treatment with the synthetic auxin NAA, a differential decrease of expression is observed in the root columella cells, while nearly no difference in their expression change is observed in the vascular cylinder. (f) Scheme of the P4AG reporter for visualizing the PIN4a isoform (PIN4 shows an identical exon/intron structure with PIN7 [16]). Following the induction of the 35S>>XVE:P4AG transgene, the GFP fluorescence is also seen outside (g) the PIN4:PIN4-GFP expression domain (h). Bars, 100 µm on (e), (d), (g) and (h). Black—coding regions, grey—UTRs, thin grey line—introns, yellow—regions modified by AS, red—RFP, green—GFP on (a), (b), (c) and (f). The color shading of the evolutional conservation code outlined (c) was assessed as a percentage of the preserved nucleotides on the alignment of the PIN7-like sequences within the Brassicaceae family.
Figure 2.
Reporters visualizing AS of PIN7 and PIN4 in the root tip: (a) Diagram of the coding regions of the PIN7 gene. The canonical PIN7b isoform differs from PIN7a in the presence of the 12 nucleotide region (orange). (b) Scheme of the P7A2G reporter for visualizing the PIN7a isoform. Removing the nucleotide prior to the alternative splice site leads to a frameshift in the PIN7b transcript. The frame is restored by insertion of the nucleotide into the region exclusively encoded in the P7A2G sequence following the alternative splice site. Asterisk: a neighboring A nucleotide is removed in the P7A1G sensor [10] (c) The P7BR reporter, as outlined in [10]. The visualization of P7BR is accomplished by introducing the termination codon into the protruding region encoded entirely by PIN7a. P7AG and P7BR are encoded by two constructs that are separately transformed and then crossed. (d) Expression pattern of the P7A2G reporter. (e) Following the treatment with the synthetic auxin NAA, a differential decrease of expression is observed in the root columella cells, while nearly no difference in their expression change is observed in the vascular cylinder. (f) Scheme of the P4AG reporter for visualizing the PIN4a isoform (PIN4 shows an identical exon/intron structure with PIN7 [16]). Following the induction of the 35S>>XVE:P4AG transgene, the GFP fluorescence is also seen outside (g) the PIN4:PIN4-GFP expression domain (h). Bars, 100 µm on (e), (d), (g) and (h). Black—coding regions, grey—UTRs, thin grey line—introns, yellow—regions modified by AS, red—RFP, green—GFP on (a), (b), (c) and (f). The color shading of the evolutional conservation code outlined (c) was assessed as a percentage of the preserved nucleotides on the alignment of the PIN7-like sequences within the Brassicaceae family.

Figure 3.
A screening system for the factors genetically upstream of the PIN7a/b splicing event: (a) A scheme of the P7A1D reporter. Physiologically inactive D-isoleucine is converted by DAAO to the toxic 3-methyl-2-oxopentanoate when P7A1D is expressed [37]. (b) The 6-days-old transgenic seedlings harboring the P7A1D reporter on control media. (c) The 12-days-old seedlings carrying the P7A1D reporter germinated on the media supplemented with 30 mM D-isoleucine, which leads to severe tissue lesions and loss of viability.
Figure 3.
A screening system for the factors genetically upstream of the PIN7a/b splicing event: (a) A scheme of the P7A1D reporter. Physiologically inactive D-isoleucine is converted by DAAO to the toxic 3-methyl-2-oxopentanoate when P7A1D is expressed [37]. (b) The 6-days-old transgenic seedlings harboring the P7A1D reporter on control media. (c) The 12-days-old seedlings carrying the P7A1D reporter germinated on the media supplemented with 30 mM D-isoleucine, which leads to severe tissue lesions and loss of viability.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kashkan, I.; Timofeyenko, K.; Kollárová, E.; Růžička, K. In Vivo Reporters for Visualizing Alternative Splicing of Hormonal Genes. Plants 2020, 9, 868. https://doi.org/10.3390/plants9070868
AMA Style
Kashkan I, Timofeyenko K, Kollárová E, Růžička K. In Vivo Reporters for Visualizing Alternative Splicing of Hormonal Genes. Plants. 2020; 9(7):868. https://doi.org/10.3390/plants9070868
Chicago/Turabian StyleKashkan, Ivan, Ksenia Timofeyenko, Eva Kollárová, and Kamil Růžička. 2020. "In Vivo Reporters for Visualizing Alternative Splicing of Hormonal Genes" Plants 9, no. 7: 868. https://doi.org/10.3390/plants9070868
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.