
Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials



1
School of Biological Sciences, University of Canterbury, Christchurch 8140, New Zealand
2
School of Life Sciences, Yantai University, Yantai 264005, China
3
Laboratory of Growth Regulators, Institute of Experimental Botany of the Czech Academy of Sciences & Faculty of Science of Palacký University, CZ-783 71 Olomouc, Czech Republic
*
Author to whom correspondence should be addressed.
Plants 2021, 10(11), 2309; https://doi.org/10.3390/plants10112309
Submission received: 7 October 2021
/
Accepted: 22 October 2021
/
Published: 27 October 2021
(This article belongs to the Special Issue 10th Anniversary of Plants—Recent Advances and Perspectives)
Abstract
:Using plant growth regulators to alter cytokinin homeostasis with the aim of enhancing endogenous cytokinin levels has been proposed as a strategy to increase yields in wheat and barley. The plant growth regulators INCYDE and CPPU inhibit the cytokinin degrading enzyme cytokinin oxidase/dehydrogenase (CKX), while TD-K inhibits the process of senescence. We report that the application of these plant growth regulators in wheat and barley field trials failed to enhance yields, or change the components of yields. Analyses of the endogenous cytokinin content showed a high concentration of trans-zeatin (tZ) in both wheat and barley grains at four days after anthesis, and statistically significant, but probably biologically insignificant, increases in cisZ-O-glucoside, along with small decreases in cZ riboside (cZR), dihydro Z (DHZ), and DHZR and DHZOG cytokinins, following INCYDE application to barley at anthesis. We discuss possible reasons for the lack of efficacy of the three plant growth regulators under field conditions and comment on future approaches to manipulating yield in the light of the strong homeostatic mechanisms controlling endogenous cytokinin levels.
Keywords:
cytokinin; TD-K; thidiazuron; INCYDE; CPPU; isopentenyl transferase; IPT; cytokinin oxidase/dehydrogenase; CKX; wheat; barley; yield1. Introduction
Food producers face a range of challenges in addressing global food security in the 21st century. These include continuing growth in food consumption in developing nations [1] and the effects of climate change, which will likely have significant and adverse effects on the environment and agriculture [2,3,4]. Increasing the yield of cereal crops, including wheat and barley, is fundamental to ensuring food security. In the 2019/2020 season, global production of wheat was more than 770 million tonnes, while for barley it was more than 150 million tonnes [5]. Several traits in cereals have been identified as important components of, and contributors to, overall yields, including having more productive tillers [6,7,8], a greater proportion of fertile grain-containing florets, larger grains, and leaf senescence occurring at an optimal time [9]. Notably, there can also be trade-offs between different components of yield, where increasing grain number can result in a decrease in grain weight [10,11,12,13,14]. Likewise, the production of more tillers is not necessarily beneficial, as small, unproductive tillers could direct resources away from productive tillers and negatively impact yield [15,16].
The cytokinins are a plant hormone group involved in many aspects of growth and development, including root and shoot growth [17,18,19], flower development [20,21], nitrogen signaling [22,23,24], senescence [25,26], stress response [27], seed yield components [28], and seed development [29,30,31,32], making them an important contributor to cereal yield.
Cytokinins are often grouped into three biologically active forms: naturally occurring, substituted adenines with either an N6 isoprenoid side chain or an aromatic side chain; and the synthetic diphenyl ureas. Briefly, isopentenyl transferase (IPT) catalyses the first committed step towards the formation of the isoprenoid cytokinins. The first formed cytokinins are the nucleotides that are converted by LOG (LONELY GUY) to the active free base forms, trans-zeatin (tZ), N6-isopentenyladenine (iP), cis-zeatin (cZ), and dihydrozeatin (DHZ), which are detected by a two-component signaling system. Cytokinin levels are controlled through destruction by cytokinin oxidase/dehydrogenase (CKX) or inactivation by cytokinin glucosyl transferase to O- or N-glucosides [32].
Previous attempts at manipulating yield and endogenous cytokinin have included the direct application of cytokinin itself to both wheat [33,34] and barley [35,36]. These approaches have involved direct injection into plant organs [34,37,38] or, more practically, through irrigation and spraying ([39], and references therein). However, success in field trials has been mixed, with findings in controlled experiments often hard to replicate in the field given the range of environmental factors and the complexity of analyzing their effects [12,39,40].
An alternative to the application of cytokinin has been the targeting of the enzymes that either deactivate cytokinin through glucosylation [41], or irreversibly degrade cytokinin via CKX [42,43]. Targeting CKX expression and/or activity has been suggested as a potential strategy to enhance yield [28,31,44,45,46], and CKX gene family members (GFMs) have been identified as being important for determining yield in both wheat and barley ([9], and references therein).
Given the challenge of increasing yield in the field using cytokinin [31], there has been a search for alternative compounds that might impact components of yield, including compounds that target CKX and compounds that might affect yield through other processes, including senescence. Such compounds include CPPU, TDZ, and the novel plant growth regulators (PGRs) INCYDE and TD-K [46,47,48,49]. These compounds became the focus for this research.
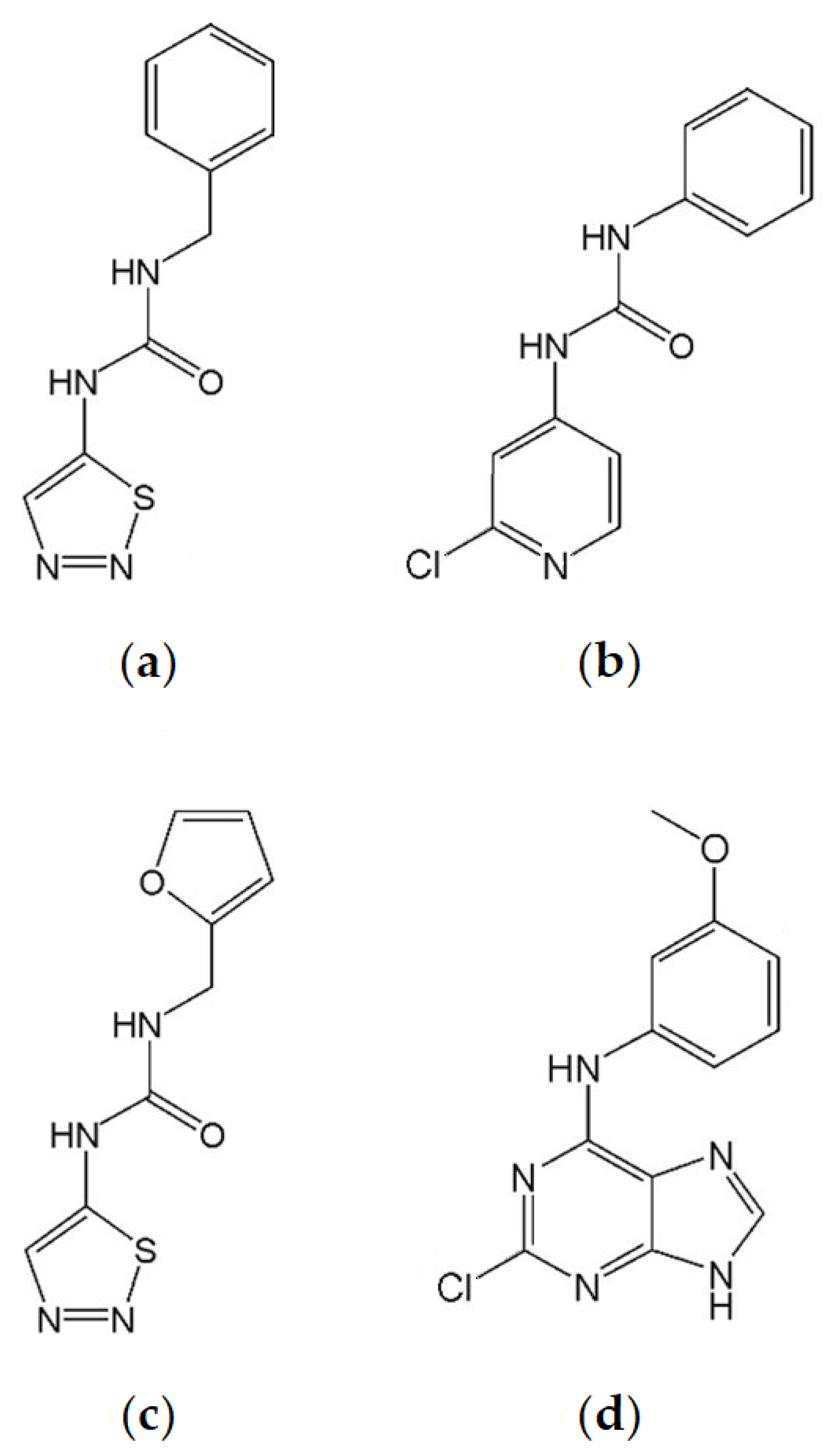
Thidiazuron is a substituted phenylurea (Figure 1a) that has been shown to inhibit CKX [50,51,52]. Thidiazuron has strong cytokinin activity [53,54,55]. It is able to activate cytokinin receptors [48,53,54,56] and has anti-senescence properties [46,57] that are stronger than trans-zeatin (tZ) and 6-benzylaminopurine (BAP) [48]. It is also able to promote shoot growth [58,59,60,61], increase fruit size [62], and produce ethylene when applied to leaves [48]. The latter property makes it desirable as a cotton defoliant [63].
CPPU (N-(2-chloro-4-pyridyl)-N’-phenylurea) is a diphenylurea derivative (Figure 1b) which is able to inhibit CKX [64,65] more strongly than TDZ [66]. Although it activates cytokinin receptors AHK3/AHK4, it does so more weakly than TDZ [52]. CPPU is also reported to be able to delay senescence [67], promote shoot formation [68], enhance fruit size [69,70,71,72], promote earlier flowering [73], and provide resistance to drought stress [74].
TD-K (N-furfuryl-N’-1,2,3-thiadiazol-5-yl-urea) is a diphenylurea thidiazuron derivative (Figure 1c) which has strong cytokinin activity comparable to BA in Amaranthus and tobacco callus bioassays [49]. TD-K has strong anti-senescence capacity, relative to TDZ and BA [48,49]. Compared to TDZ, it more weakly activates cytokinin receptors [49,53,54,56], is less able to promote ethylene production in mung bean hypocotyls [75], and, in contrast to TDZ, does not inhibit root growth [48].
INCYDE (2-chloro-6-(3-methoxyphenyl)aminopurine) is a substituted 6-anilinopurine derivative (Figure 1d). It is a stronger inhibitor of cytokinin oxidase/dehydrogenase than TDZ, while more weakly activating cytokinin receptors compared to TDZ and tZ [55]. It activated the cytokinin responsive reporter gene ARR5:GUS [76] in a dose-dependent manner. INCYDE was shown to enhance yield of Rapid Cycling Brassica rapa but only under specific, controlled conditions [49]. INCYDE increased shoot FW in CKX1-overexpressing Arabidopsis thaliana seedlings [76]. INCYDE application has been reported to increase flower production in tomatoes [77], shoot production when applied with BA [78], and has a dose-dependent inhibition of shoot and/or root growth in Bulbine natalensis and Rumex crispus [45] and micropropagated Eucomis autumnalis [78]. INCYDE is also reported to alleviate the effects of biotic [79] and abiotic stress [45,77]. Additionally, when applied in the field to barley, analogue INCYDE-F was responsible for altering the endogenous cytokinin content [80].
Three PGRs with different properties and modes of action were selected for this investigation: INCYDE, TD-K, and CPPU. These compounds were applied to wheat and barley in field trials and components of the yields were analyzed. The effects of these compounds on endogenous cytokinins was also examined.
2. Results
2.1. Field Trial Analyses
Analyses carried out on the harvested wheat and barley from the field trials did not reveal any statistically significant difference in the yield (T/ha), thousand grain weight (TGW) in grams (g), or protein composition between any of the treatments and the controls for either wheat or barley (Table 1). The Orator wheat (2013/14) field trial was broadly infected with Septoria during a critical time in development, which negatively impacted the yield. Given the lack of evidence for any change in yield, the field trials were discontinued. Additional trials were carried out using outdoor pot trials where the same treatments and growth stages described for the field trials were used, but no statistically significant differences in yield or yield components were found for these trials either [81].
2.2. LC–MS/MS Analyses in Grain
LC–MS/MS analyses of wheat and barley grains from control plants assessed four days after anthesis (4 DAA) show that the concentration of tZ was much greater than the concentration of the other free bases iP, cZ, or DHZ (Table 2 and Table 3). Inactivation by glucosylation is clearly evident, as shown by the elevated levels of cZ- and cZ riboside-O-glucosides (cZOG and cZROG) in barley, and in wheat by elevated levels of tZ 9-glucoside (tZ9G), cZOG, and cZROG.
In wheat grains, neither TD-K nor CPPU treatment resulted in a significant change in any of the cytokinin metabolites compared to the control (Table 2). At four days following INCYDE treatment/anthesis in barley grains, there was a significant increase in the content of cZ O-glucoside (cZOG), cZ-types overall and the total O-glucoside cytokinins (Table 3). Conversely, there were small but statistically significant decreases in the concentration of cZR, DHZ, DHZR, DHZOG, and the total base and ribosides of cZ and DHZ cytokinins following INCYDE application.
3. Discussion
The region where our field trials were conducted, Canterbury, New Zealand, is known for world record cereal production (17.398 tonnes per hectare of wheat crop (Guinness World Records, 2020)). Our trials were conducted under optimal field conditions of water and fertilizer, which we recognize as a potentially challenging environment to assess PGR efficacy, a comment also made by Nisler et al. [66] with respect to their PGR field trials in the Czech Republic.
The lack of yield enhancement following INCYDE application (Table 1) suggests that this compound had little effect in our field trials on either wheat or barley. Positive trends in yield in field trials of wheat and barley treated with cytokinin derivatives similar to INCYDE have been reported but these failed to reach statistical significance [80]. Consequently, our field data are not in conflict with this. While the Orator wheat (2013/14) field trial, where INCYDE was applied, was impacted by Septoria, there was no evidence of INCYDE ameliorating the effect of this disease, in contrast to the report by Reusche et al. [79]. This is not to imply that INCYDE is not efficacious under other conditions, as changes in gene expression occur following application [83], and responses are clearly evident under more controlled environments, including in bioassays [55], in in vitro culture settings [45,77,78,79], and in pot trials with Rapid Cycling Brassica rapa [49].
The statistically significant increase in cZOG following INCYDE application to barley may show a mechanism in common with previous in vitro experiments, where INCYDE (with BA) enhanced O-glucoside accumulation in banana plantlets [84]. It is possible that active cytokinin forms may have been channelled into inactivated O-glucosides as a consequence of reduced inactivation by CKX, due to inhibition of CKX by INCYDE. Because of the activation of homeostatic mechanisms, and also because of the very high endogenous levels of active tZ immediately after anthesis, any transitory increases in active cytokinins, if they had occurred, are likely to be biologically insignificant.
Neither of the two diphenylurea-derivatives, TD-K or CPPU, enhanced yield (Table 1). This is in contrast with an increase of 120.9% for oilseed rape yield (6.038 vs. 4.99 T/ha), and 106% (7.02 vs. 7.49 T/ha) for spring barley reported in the TD-K patent for PGR application at BBCH50 (extension growth) [48]. Details of statistical significance are not, however, provided for the different crops. More recently, a diphenylurea derivative was applied to barley and wheat under field conditions in the Czech Republic [66]. Although these studies targeted earlier growth stages, including at BBCH 20–25, as well as seed treatments, they also targeted the emergence of the inflorescence (BBCH 51), and at a concentration range between 5 and 50 µM, which is comparable to that used in our study. However, the field data for wheat and barley treated with diphenylurea-derivative Compound 19 are only presented as percent of control without statistical analyses available [66]. The variability apparent between years (particularly in tiller number and 1000 grain weight) makes it essential that statistical analysis of the yield data (0.7 to 6.6% yield increase compared to control) is presented.
Likewise, CPPU, despite having success with enhancing fruit size, has not had much success when used to target cereals in the field ([32], and references therein). The difficulties of replicating findings from controlled environments onto the field have been reported [12,40], with field trials introducing a multitude of uncontrolled or difficult to control factors, many of which could affect cytokinin homeostasis.
An increased tiller number is not necessarily seen as desirable in wheat [9], so we specifically targeted the PGRs at later stages of development: for TD-K this was from anthesis onwards, due to its strong anti-senescence properties [48,49]; and for INCYDE and CPPU from GS39, when florets are being established, and/or GS51, when ears are particularly susceptible to stress [85,86], and or across anthesis, the latter chosen due to the rapidly changing cytokinin content and elevated CKX expression associated with this stage in development ([9], and references therein). Indeed, a high level of tZ cytokinin was identified in wheat four days after anthesis (DAA) (Table 2). This aligns with previous reports of high levels of zeatin in wheat early in grain development [87,88,89,90,91], and, moreover, confirms that this cytokinin is tZ. The transient nature of this narrow developmental window that is associated with cell division is also a possible reason for the lack of yield enhancement by cytokinins in cereal field trials ([31], and references therein), since in the field environment, anthesis is spread across several days, although we attempted to cover this by applications at GS61 and 65.
The high concentration of tZ in barley at 4 DAA (Table 3) has also been reported [92]. In contrast, the low concentration of cZ contrasts with the high peak of cZ reported previously in developing barley kernels [93]. This suggests that 4 DAA is possibly after the cZ peak. The high concentration of cZOG suggests active deactivation of cZ within days post-anthesis.
Our research suggests that INCYDE, TD-K, and CPPU have little to no effect on components of harvestable yield in wheat and barley grown under optimal field conditions. Additionally, this research highlights some of the difficulties and issues of conducting field trials with PGRs, with any attempt to manipulate cytokinin made more difficult not only by strong homeostatic responses but also by the complex, pleiotropic nature of cytokinin [31,39]. Feedback responses following the disturbance of cytokinin homeostasis have been observed or suggested elsewhere in the form of an increase in CKX expression and/or activity [14,94,95,96,97,98,99]. An increase in cytokinin following CKX inhibition might also be responsible for an enhancement in the deactivation of cytokinins, which could explain the stronger production of cis-type O-glucosides seen in barley (Table 3). Feedback mechanisms might also involve IPT GFMs, with HvIPT1 and HvIPT2 both being downregulated in response to a local increase in cytokinin following the knockout of HvCKX1 [100].
Despite these difficulties, targeting CKX is still an important strategy for manipulating cytokinin and yield [9,31,45,46], and arguably more suitable than alternative strategies, including the direct application of cytokinin, or targeting IPT, given that CKX is considered a more moderate or ‘softer’ regulator of cytokinin compared to IPT [101]. Future research could focus on determining if the endogenous changes in barley (Table 3) and, indeed, the lack of change in wheat, were the result of changes in expression of genes associated with cytokinin homeostasis, including biosynthesis (IPTs), degradation (CKXs), and glucosylation (CGTs), and whether these results could help explain the lack of yield in the field trials. Additionally, with the identification of the key CKX gene family members that affect yield in wheat (reviewed in [9]), and with interesting results in wheat [13,14,102], barley [12,87,103,104], and rice [44,105] trials, transgenic approaches hold significant potential for enhancing yield in cereals.
However, whether the resulting cereal is a result of genetic modification or gene editing, in some jurisdictions such plants are subject to legal and social restrictions which make their cultivation, processing, and marketing difficult or impossible [106,107,108]. In this context, non-transgenic approaches, such as the Targeting Induced Local Lesions In Genome (TILLING) strategy, offer numerous advantages, including overcoming the limits imposed by the lack of genetic variability in traditional breeding, the acceleration of breeding programs, and, above all, the possibility of developing new varieties that do not have the limitations that characterize transgenic organisms [107]. Both the CRISPR/Cas9-mediated gene editing technology and the TILLING approach have their own merits and demerits relating to the initial investment by researchers, the access to the requisite technology, the range of mutations that are either targeted (in gene editing) or identified (multiple point mutations in TILLING) and their use in breeding [106].
More recently, two in silico TILLING resources have been generated and made publicly available. These include the whole exome sequencing of over 1200 TILLING mutant lines of a well-known European bread wheat variety Cadenza [109,110]. Similarly, an in silico TILLING resource is being generated for the most widely grown Chinese bread wheat variety, Jimai 22. Within this population, multiple point mutants for not only all CKX GFMs but also the zeatin O-glucosyl transferase (ZOGT) GFMs have been identified [9,41]. Importantly, while CKX GFMs have been the target of much research [9], the high levels of cytokinin glucosides in wheat and barley, and the negative relationship of ZOGT gene expression with yield in wheat [41,91], indicate that the ZOGT GFMs warrant further investigation, which is beyond the tools offered by the CKX inhibiting PGRs.
4. Materials and Methods
4.1. Field Trials
Wheat and barley field trials were carried out over two seasons, near Lincoln, New Zealand (43°36′15.7″ S 172°25′56.0″ E and 43°37′04.7″ S 172°27′09.4″ E). Autumn-sown wheat (cultivar Orator) was grown in the 2013/14 season, while barley (cultivar Quench) and wheat (cultivar Torch) were grown in 2014/15. Sowing spacing was kept constant, to prevent any confounding effect on tiller number. Field trials were carried out in a farmer’s paddock and subject to standard field management including regular irrigation, fertilizer application, and application of compounds, including herbicide, insecticide, and fungicides, where necessary. Field trials were planted in 10 m × 2.5 m plots, arranged in a randomized complete block design with four replicates for each treatment. Plant growth regulators INCYDE, TD-K, and CPPU were applied at concentrations between 10 and 100 µM at growth stages (GS), defined according to the Zadoks scale [82], including GS 39 (the appearance of flag leaf ligule), GS 51 (appearance of the spikelet), and at GS 61 to 69 (defined as anthesis). Plant growth regulators were applied at rates of 187 L/ha for the 2013/14 trial, and 170 L/ha for the 2014/15 trial.
INCYDE, TD-K and CPPU were prepared by dissolving compounds in dimethylsulfoxide (DMSO) (Scharlab), diluted with water and then, prior to application, mixed with surfactant (Yates Sprayfix, Yates) at 0.5% (v/v). Two controls were used in the field trials, ‘untreated controls’ where no application was made, and ‘DMSO controls’ where the amount of DMSO used was equivalent to the highest PGR concentration for each respective trial, unless stated otherwise in the results. Applications were made by New Zealand Arable using CO2 pressurized hand-hand plot booms for applications rates between 170–190 L/ha.
4.2. Plant Material
4.2.1. Yield and Protein Composition Analyses
Once wheat and barley plants had senesced completely, plants were harvested with a Sampo combine harvester (Sampo Rosenlew Ltd., Pori, Finland) and protein content was analyzed by New Zealand Grainlab. Onboard weighing provided the analysis of yield (tonnes per hectare) and, using 20 g screened samples of grain, the TGW was calculated with a Numigral I seed counter (Sinar). Protein composition was analyzed using an Instalab® 700 NIR Analyzer (DICKEY-john). The thousand grain weight was calculated for each plot, using 20 g of screened grain samples.
4.2.2. LC–MS/MS Analyses
Grain material for LC–MS/MS analyses was sampled from the field trials, following anthesis-targeted application of either INCYDE (50 µM), TD-K (50 µM), CPPU (100 µM), or water + DMSO. Following treatment, whole heads were sampled at day 4 after anthesis, which was 4 days after treatment. Wheat and barley heads were frozen by immediately submerging the samples in liquid nitrogen and storing at −80 °C. Wheat and barley grains were dissected from the middle third section of the spike, with basal florets within the spikelet targeted in wheat [9]. Grains were then organized, based on the developmental stages as described in [111]. INCYDE-treated wheat grains were not sampled for LC–MS/MS analyses, given that this trial (wheat cv. Orator, 2013/14) was infected with Septoria at a critical time during grain development.
Grains were ground under liquid nitrogen and freeze dried with a Savant™ SPD131DDA SpeedVac™ Concentrator (Thermo Fisher Scientific) to produce samples weighing between 8 to 22 mg. For each treatment, three replicates were prepared. Samples were then analyzed according to [112]. Sample extraction was carried out with a modified Bieleski solution (60% MeOH, 10% HCOOH, and 30% H2O), and [13C5]cZ, [13C5]tZ, [2H5]tZR, [2H5]tZ7G, [2H5]tZ9G, [2H5]tZOG, [2H5]tZROG, [2H5]tZMP, [2H3]DHZ, [2H3]DHZR, [2H3]DHZ9G, [2H7]DHZOG, [2H3]DHZMP, [2H6]iP, [2H6]iPR, [2H6]iP7G, [2H6]iP9G, [2H6]iPMP stable isotope-labelled standards (0.25 pmol of cytokinin bases, ribosides, N-glucosides, 0.5 pmol of cytokinin O-glucosides and nucleotides; Olchemim) were added to each sample to validate phytohormone determination. Sample purification was carried out with mixed-mode cation-exchange (MCX) cartridges (Oasis MCX, 30 mg/1 mL; Waters). Analytes were eluted by two-step elution using a 0.35 M NH4OH aqueous solution and 0.35 M NH4OH in 60% (v/v) methanol solution. The resulting eluate was subsequently evaporated to dryness and then dissolved in the mobile phase (15 mM ammonium formate pH 4.0 in 5% (v/v) methanol). LC–MS/MS analyses were carried out using a Acquity UPLC® System (Waters) and a triple quadrupole mass spectrometer XevoTM TQ MS (Waters). The mass spectrometry data was then processed utilizing MassLynx™ Mass Spectrometry Software with TargetLynx™ (Waters).
4.3. Statistical Analyses
For yield and protein composition from the field trials, the mean was generated using four replicates for each treatment and the data presented with standard errors. Statistically significant differences, where p ≤ 0.05, were determined between PGR treatments and the respective DMSO control using a two-way ANOVA. A logit transformation was made to protein composition data prior to ANOVA analysis. Similarly, statistically significant differences for LC–MS/MS data were determined between PGR treatments and the control using two-way ANOVA (significance level: 0.05), with a post hoc two-sided Dunnett test (Confidence Interval: 95%). To ensure the assumptions of the ANOVA were met, an examination of Q-Q plots of standardized residuals was made, and where necessary the equality of variances ensured through a Levene’s test and plot of standardized residuals and predicted values.
Author Contributions
Conceptualization, P.E.J. and J.S.; methodology, J.S., P.E.J. and O.N.; validation, J.S., M.J.v.V. and O.N.; formal analysis, M.J.v.V. and P.E.J.; investigation, M.J.v.V., J.S. and P.E.J.; resources, P.E.J., J.S. and O.N.; data curation, M.J.v.V. and O.N.; writing—original draft preparation, M.J.v.V.; writing—review and editing, P.E.J., M.J.v.V. and J.S.; visualization, M.J.v.V.; supervision, P.E.J. and J.S.; project administration, P.E.J.; funding acquisition, P.E.J., J.S. and O.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Foundation for Arable Research, New Zealand in the provision of a PhD scholarship (M.J.v.V.) and expendables (P.E.J., M.J.v.V. and J.S.); and by the Ministry of Education, Youth and Sports of the Czech Republic through the European Regional Development Fund-Project ‘Plants as a tool for sustainable global development’ (CZ.02.1.01/0.0/0.0/16_019/0000827, O.N.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are contained within the article.
Acknowledgments
The authors sincerely thank Hana Martínková and Petra Amakorová for their help with the analysis of phytohormones. We acknowledge Marek Zatloukal for INCYDE and Jaroslav Nisler for TD-K. We acknowledge New Zealand Arable for application of the PGRs and the New Zealand Grainlab for the yield and composition analyses.
Conflicts of Interest
The authors of this manuscript declare no conflict of interest.
References
- Baldos, U.L.C.; Hertel, T.W. Global food security in 2050: The role of agricultural productivity and climate change. Aust. J. Agric. Econ. 2014, 58, 554–570. [Google Scholar] [CrossRef] [Green Version]
- Schmidhuber, J.; Tubiello, F.N. Global food security under climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19703–19708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nellemann, C.; MacDevette, M.; Manders, T.; Eickhout, B.; Svihus, B.; Prins, A.G.; Kaltenborn, B.P. The Environmental Food Crisis: The Environment’s Role in Averting Future Food Crises: A UNEP Rapid Response Assessment; UNEP/Earthprint: Stevenage, UK, 2009. [Google Scholar]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- USDA Foreign Agricultural Service. Grain: World Markets and Trade; USDA Foreign Agricultural Service: Washington, DC, USA, 2021.
- Langer, R.H.M.; Prasad, P.C.; Laude, H.M. Effects of kinetin on tiller bud elongation in wheat (Triticum aestivum L.). Ann. Bot. 1973, 37, 565–571. [Google Scholar] [CrossRef]
- Darwinkel, A. Ear development and formation of grain yield in winter wheat. Neth. J. Agric. Sci. 1980, 28, 156–163. [Google Scholar] [CrossRef]
- Harrison, M.A.; Kaufman, P.B. Hormonal regulation of lateral bud (tiller) release in oats (Avena sativa L.). Plant Physiol. 1980, 66, 1123–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhao, J.; Song, J.; Jameson, P.E. Cytokinin dehydrogenase: A genetic target for yield improvement in wheat. Plant Biotechnol. J. 2020, 18, 614–630. [Google Scholar] [CrossRef] [PubMed]
- Van Daele, I.; Gonzalez, N.; Vercauteren, I.; de Smet, L.; Inzé, D.; Roldán-Ruiz, I.; Vuylsteke, M. A comparative study of seed yield parameters in Arabidopsis thaliana mutants and transgenics. Plant Biotechnol. J. 2012, 10, 488–500. [Google Scholar] [CrossRef]
- Guo, T.; Chen, K.; Dong, N.-Q.; Shi, C.-L.; Ye, W.-W.; Gao, J.-P.; Shan, J.-X.; Lin, H.-X. GRAIN SIZE AND NUMBER1 negatively regulates the OsMKKK10-OsMKK4-OsMPK6 cascade to coordinate the trade-off between grain number per panicle and grain size in rice. Plant Cell 2018, 30, 871–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holubová, K.; Hensel, G.; Vojta, P.; Tarkowski, P.; Bergougnoux, V.; Galuszka, P. Modification of barley plant productivity through regulation of cytokinin content by reverse-genetics approaches. Front. Plant Sci. 2018, 9, 1676. [Google Scholar] [CrossRef] [Green Version]
- Jabłoński, B.; Ogonowska, H.; Szala, K.; Bajguz, A.; Orczyk, W.; Nadolska-Orczyk, A. Silencing of TaCKX1 mediates expression of other TaCKX genes to increase yield parameters in wheat. Int. J. Mol. Sci. 2020, 21, 4809. [Google Scholar] [CrossRef]
- Jabłoński, B.; Szala, K.; Przyborowski, M.; Bajguz, A.; Chmur, M.; Gasparis, S.; Orczyk, W.; Nadolska-Orczyk, A. TaCKX2.2 genes coordinate expression of other TaCKX family members, regulate phytohormone content and yield-related traits of wheat. Int. J. Mol. Sci. 2021, 22, 4142. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, P.W.; Kirkegaard, J.A.; Lilley, J.M.; Gregory, P.J.; Rebetzke, G.J. A tillering inhibition gene influences root–shoot carbon partitioning and pattern of water use to improve wheat productivity in rainfed environments. J. Exp. Bot. 2016, 67, 327–340. [Google Scholar] [CrossRef]
- Kebrom, T.H.; Chandler, P.M.; Swain, S.M.; King, R.W.; Richards, R.A.; Spielmeyer, W. Inhibition of tiller bud outgrowth in the tin mutant of wheat is associated with precocious internode development. Plant Physiol. 2012, 160, 308–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef] [Green Version]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H.; Schmülling, T. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2003, 15, 2532–2550. [Google Scholar] [CrossRef] [Green Version]
- Brenner, W.G.; Schmülling, T. Transcript profiling of cytokinin action in Arabidopsis roots and shoots discovers largely similar but also organ-specific responses. BMC Plant Biol. 2012, 12, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef] [Green Version]
- D’Aloia, M.; Bonhomme, D.; Bouché, F.; Tamseddak, K.; Ormenese, S.; Torti, S.; Coupland, G.; Périlleux, C. Cytokinin promotes flowering of Arabidopsis via transcriptional activation of the FT paralogue TSF. Plant J. 2011, 65, 972–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takei, K.; Ueda, N.; Aoki, K.; Kuromori, T.; Hirayama, T.; Shinozaki, K.; Yamaya, T.; Sakakibara, H. AtIPT3 is a key determinant of nitrate-dependent cytokinin biosynthesis in Arabidopsis. Plant Cell Physiol. 2004, 45, 1053–1062. [Google Scholar] [CrossRef] [Green Version]
- Shtratnikova, V.Y.; Kudryakova, N.V.; Kudoyarova, G.R.; Korobova, A.V.; Akhiyarova, G.R.; Danilova, M.N.; Kusnetsov, V.V.; Kulaeva, O.N. Effects of nitrate and ammonium on growth of Arabidopsis thaliana plants transformed with the ARR5::GUS construct and a role for cytokinins in suppression of disturbances induced by the presence of ammonium. Russ. J. Plant Physiol. 2015, 62, 741–752. [Google Scholar] [CrossRef]
- Gu, J.; Li, Z.; Mao, Y.; Struik, P.C.; Zhang, H.; Liu, L.; Wang, Z.; Yang, J. Roles of nitrogen and cytokinin signals in root and shoot communications in maximizing of plant productivity and their agronomic applications. Plant Sci. 2018, 274, 320–331. [Google Scholar] [CrossRef]
- Gan, S.; Amasino, R.M. Cytokinins in plant senescence: From spray and pray to clone and play. Bioessays 1996, 18, 557–565. [Google Scholar] [CrossRef]
- Zwack, P.J.; Rashotte, A.M. Cytokinin inhibition of leaf senescence. Plant Signal. Behav. 2013, 8, e24737. [Google Scholar] [CrossRef]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Schwarz, I.; Scheirlinck, M.T.; Otto, E.; Bartrina, I.; Schmidt, R.C.; Schmülling, T. Cytokinin regulates the activity of the inflorescence meristem and components of seed yield in oilseed rape. J. Exp. Bot. 2020, 71, 7146–7159. [Google Scholar] [CrossRef]
- Riefler, M.; Novak, O.; Strnad, M.; Schmülling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 2006, 18, 40–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Jameson, P.E.; Song, J. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2016, 67, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jameson, P.E.; Song, J. Will cytokinins underpin the second ‘Green Revolution’? J. Exp. Bot. 2020, 71, 6872–6875. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, W.; Dai, T.; Zhou, Q. Effects of exogenous hormones on floret development and grain setting in wheat. Plant Growth Regul. 2001, 35, 225–231. [Google Scholar] [CrossRef]
- Gupta, N.K.; Gupta, S.; Shukla, D.S.; Deshmukh, P.S. Differential responses of BA injection on yield and specific grain growth in contrasting genotypes of wheat (Triticum aestivum L.). Plant Growth Regul. 2003, 40, 201–205. [Google Scholar] [CrossRef]
- Williams, R.H.; Cartwright, P.M. The effect of applications of a synthetic cytokinin on shoot dominance and grain yield in spring barley. Ann. Bot. 1980, 46, 445–452. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Poustini, K.; Ahmadi, A. Effects of foliar application of BAP on source and sink strength in four six-rowed barley (Hordeum vulgare L.) cultivars. Plant Growth Regul. 2008, 54, 231–239. [Google Scholar] [CrossRef]
- Warrier, A.; Bhardwaj, S.N.; Pande, P.C. Effect of benzyladenine on grain growth in Aestivum wheat. Plant Cell Physiol. 1987, 28, 735–739. [Google Scholar] [CrossRef]
- Sivakumar, T.; Srivastava, G.C. Effects of benzyl adenine and abscisic acid on grain yield and yield components in Triticale and wheat. J. Agron. Crop. Sci. 2001, 186, 43–46. [Google Scholar] [CrossRef]
- Koprna, R.; De Diego, N.; Dundálková, L.; Spíchal, L. Use of cytokinins as agrochemicals. Bioorg. Med. Chem. 2016, 24, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Nagel, L.; Brewster, R.; Riedell, W.E.; Reese, R.N. Cytokinin regulation of flower and pod set in soybeans (Glycine max (L.) Merr.). Ann. Bot. 2001, 88, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhao, J.; Song, J.; Jameson, P.E. Cytokinin glucosyl transferases, key regulators of cytokinin homeostasis, have potential value for wheat improvement. Plant Biotechnol. J. 2021, 19, 878–896. [Google Scholar] [CrossRef] [PubMed]
- McGaw, B.A.; Horgan, R. Cytokinin oxidase from Zea mays kernels and Vinca rosea crown-gall tissue. Planta 1983, 159, 30–37. [Google Scholar] [CrossRef]
- Galuszka, P.; Frébort, I.; Šebela, M.; Peč, P. Degradation of cytokinins by cytokinin oxidases in plants. Plant Growth Regul. 2000, 32, 315–327. [Google Scholar] [CrossRef]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Plant science: Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Gemrotová, M.; Kulkarni, M.G.; Stirk, W.A.; Strnad, M.; Van Staden, J.; Spíchal, L. Seedlings of medicinal plants treated with either a cytokinin antagonist (PI-55) or an inhibitor of cytokinin degradation (INCYDE) are protected against the negative effects of cadmium. Plant Growth Regul. 2013, 71, 137–145. [Google Scholar] [CrossRef]
- Nisler, J.; Kopečný, D.; Končitíková, R.; Zatloukal, M.; Bazgier, V.; Berka, K.; Zalabák, D.; Briozzo, P.; Strnad, M.; Spíchal, L. Novel thidiazuron-derived inhibitors of cytokinin oxidase/dehydrogenase. Plant Mol. Biol. 2016, 92, 235–248. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Wardle, D.A.; Zurowski, C.; Looney, N.E. Phenylureas CPPU and thidiazuron affect yield components, fruit composition, and storage potential of four seedless grape selections. J. Am. Soc. Hortic. Sci. 1992, 117, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Nisler, J.; Zatloukal, M.; Spíchal, L.; Koprna, R.; Doležal, K.; Strnad, M. 1,2,3-thiadiazol-5yl-urea Derivatives, Use Thereof for Regulating Plant Senescence and Preparations Containing These Derivatives. U.S. Patent 9,993,002, 12 June 2018. [Google Scholar]
- Van Voorthuizen, M.J.; Nisler, J.; Song, J.; Spíchal, L.; Jameson, P.E. Targeting cytokinin homeostasis in rapid cycling Brassica rapa with plant growth regulators INCYDE and TD-K. Plants 2021, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, J.M.; Armstrong, D.J. Regulation of cytokinin oxidase activity in callus tissues of Phaseolus vulgaris L. cv Great Northern. Plant Physiol. 1986, 80, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Hare, P.D.; Van Staden, J. Inhibitory effect of thidiazuron on the activity of cytokinin oxidase isolated from soybean callus. Plant Cell Physiol. 1994, 35, 1121–1125. [Google Scholar] [CrossRef]
- Kopečný, D.; Briozzo, P.; Popelková, H.; Šebela, M.; Končitíková, R.; Spíchal, L.; Nisler, J.; Madzak, C.; Frébort, I.; Laloue, M.; et al. Phenyl-and benzylurea cytokinins as competitive inhibitors of cytokinin oxidase/dehydrogenase: A structural study. Biochimie 2010, 92, 1052–1062. [Google Scholar] [CrossRef]
- Yamada, H.; Suzuki, T.; Terada, K.; Takei, K.; Ishikawa, K.; Miwa, K.; Yamashino, T.; Mizuno, T. The Arabidopsis AHK4 histidine kinase is a cytokinin-binding receptor that transduces cytokinin signals across the membrane. Plant Cell Physiol. 2001, 42, 1017–1023. [Google Scholar] [CrossRef]
- Spíchal, L.; Rakova, N.Y.; Riefler, M.; Mizuno, T.; Romanov, G.A.; Strnad, M.; Schmülling, T. Two cytokinin receptors of Arabidopsis thaliana, CRE1/AHK4 and AHK3, differ in their ligand specificity in a bacterial assay. Plant Cell Physiol. 2004, 45, 1299–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatloukal, M.; Gemrotová, M.; Doležal, K.; Havlíček, L.; Spíchal, L.; Strnad, M. Novel potent inhibitors of A. thaliana cytokinin oxidase/dehydrogenase. Bioorg. Med. Chem. 2008, 16, 9268–9275. [Google Scholar] [CrossRef]
- Mok, M.C.; Mok, D.W.S.; Armstrong, D.J.; Shudo, K.; Isogai, Y.; Okamoto, T. Cytokinin activity of N-phenyl N’-1,2,3-thiadiazol-5-ylurea (Thidiazuron). Phytochemistry 1982, 21, 1509–1511. [Google Scholar] [CrossRef]
- Ferrante, A.; Hunter, D.A.; Hackett, W.P.; Reid, M.S. Thidiazuron—A potent inhibitor of leaf senescence in Alstroemeria. Postharvest Biol. Technol. 2002, 25, 333–338. [Google Scholar] [CrossRef]
- Malik, K.A.; Saxena, P.K. Regeneration in Phaseolus vulgaris L.: High-frequency induction of direct shoot formation in intact seedlings by N6-benzylaminopurine and thidiazuron. Planta 1992, 186, 384–389. [Google Scholar] [CrossRef]
- Singh, N.D.; Sahoo, L.; Sarin, N.B.; Jaiwal, P.K. The effect of TDZ on organogenesis and somatic embryogenesis in pigeonpea (Cajanus cajan L. Millsp). Plant Sci. 2003, 164, 341–347. [Google Scholar] [CrossRef]
- Zhihui, S.; Tzitzikas, M.; Raemakers, K.; Zhengqiang, M.; Visser, R. Effect of TDZ on plant regeneration from mature seeds in pea (Pisum sativum). Vitr. Cell. Dev. Biol. Plant 2009, 45, 776. [Google Scholar] [CrossRef]
- Guo, B.; Abbasi, B.H.; Zeb, A.; Xu, L.L.; Wei, Y.H. Thidiazuron: A multi-dimensional plant growth regulator. Afr. J. Biotechnol. 2011, 10, 8984–9000. [Google Scholar]
- Piao, Y.L.; Wang, H.T.; Zhang, H.M. Effects of TDZ on grapes quality in cold area. J. Agric. Sci. Yanbian Univ. 2006, 4, 012. [Google Scholar]
- Suttle, J.C. Involvement of ethylene in the action of the cotton defoliant thidiazuron. Plant Physiol. 1985, 78, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Hayata, Y.; Niimi, Y.; Iwasaki, N. Synthetic cytokinin-1-(2chloro4pyridyl)-3-phenylurea (CPPU)-promotes fruit set and induces parthenocarpy in watermelon. J. Am. Soc. Hort. Sci. 1995, 120, 997–1000. [Google Scholar] [CrossRef] [Green Version]
- Bilyeu, K.D.; Cole, J.L.; Laskey, J.G.; Riekhof, W.R.; Esparza, T.J.; Kramer, M.D.; Morris, R.O. Molecular and biochemical characterization of a cytokinin oxidase from maize. Plant Physiol. 2001, 125, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Nisler, J.; Kopečný, D.; Pěkná, Z.; Končitíková, R.; Koprna, R.; Murvanidze, N.; Werbrouck, S.P.O.; Havlíček, L.; De Diego, N.; Kopečná, M.; et al. Diphenylurea-derived cytokinin oxidase/dehydrogenase inhibitors for biotechnology and agriculture. J. Exp. Bot. 2021, 72, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Piao, Y.L.; Lv, H.; Bai, H.J.; Li, M.D. Effect of TDZ on fruiting and fruit growth of Xiaoxiangshui pear of Yanbian [J]. J. Agric. Sci. Yanbian Univ. 2009, 3, 007. [Google Scholar]
- Guo, D.P.; Zhu, Z.J.; Hu, X.X.; Zheng, S.J. Effect of cytokinins on shoot regeneration from cotyledon and leaf segment of stem mustard (Brassica juncea var. tsatsai). Plant Cell Tissue Organ. Cult. 2005, 83, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Biasi, R.; Costa, G.; Giuliani, R.; Succi, F.; Sansavini, S. Effects of CPPU on kiwifruit performance. In II International Symposium on Kiwifruit; International Society for Horticultural Science: Korbeek-Lo, Belgium, 1991; Volume 297, pp. 367–374. [Google Scholar]
- Flaishman, M.A.; Shargal, A.; Shlizerman, L.; Stern, R.A. The synthetic cytokinins CPPU and TDZ prolong the phase of cell division in developing pear (Pyrus communis L.) fruit. Acta Hortic. 2006, 671, 151–157. [Google Scholar] [CrossRef]
- Stern, R.A.; Ben-Arie, R.; Neria, O.; Flaishman, M. CPPU and BA increase fruit size of ‘Royal Gala’ (Malus domestica) apple in a warm climate. J. Hort. Sci. Biotechnol. 2003, 78, 297–302. [Google Scholar] [CrossRef]
- Ferrara, G.; Mazzeo, A.; Netti, G.; Pacucci, C.; Matarrese, A.M.S.; Cafagna, I.; Mastrorilli, P.; Vezzoso, M.; Gallo, V. Girdling, gibberellic acid, and forchlorfenuron: Effects on yield, quality, and metabolic profile of table grape cv Italia. Am. J. Enol. Vitic. 2014, 65, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, N.; Oba, H.; Mizuta, D.; Yoshida, H.; Olsen, J.E. Timing of blue and red light exposure and CPPU application during the raising of seedlings can control flowering timing of petunia. Acta. Hortic. 2016, 1134, 171–178. [Google Scholar] [CrossRef]
- Jianchang, H.; Yan, X.; Chunxiang, Z.; Hongbin, L. Protective effect of CPPU in papaya plant under drought stress. J. Fruit Sci. 2003, 3, 013. [Google Scholar]
- Yip, W.K.; Yang, S.F. Effect of thidiazuron, a cytokinin-active urea derivative, in cytokinin-dependent ethylene production systems. Plant Physiol. 1986, 80, 515–519. [Google Scholar] [CrossRef] [Green Version]
- Zatloukal, M.; Plihalova, L.; Klaskova, J.; Spíchal, L.; Koprna, R.; Dolezal, K.; Strnad, M. Substituted 6-Anilino-9-heterocyclylpurine Derivatives for Inhibition of Plant Stress. U.S. Patent 10,662,194, 26 May 2020. [Google Scholar]
- Aremu, A.O.; Masondo, N.A.; Sunmonu, T.O.; Kulkarni, M.G.; Zatloukal, M.; Spichal, L.; Doležal, K.; Van Staden, J. A novel inhibitor of cytokinin degradation (INCYDE) influences the biochemical parameters and photosynthetic apparatus in NaCl-stressed tomato plants. Planta 2014, 240, 877–889. [Google Scholar] [CrossRef]
- Aremu, A.O.; Stirk, W.A.; Masondo, N.A.; Plačková, L.; Novák, O.; Pěnčík, A.; Zatloukal, M.; Nisler, J.; Spíchal, L.; Doležal, K.; et al. Dissecting the role of two cytokinin analogues (INCYDE and PI-55) on in vitro organogenesis, phytohormone accumulation, phytochemical content and antioxidant activity. Plant Sci. 2015, 238, 81–94. [Google Scholar] [CrossRef]
- Reusche, M.; Klásková, J.; Thole, K.; Truskina, J.; Novák, O.; Janz, D.; Strnad, M.; Spíchal, L.; Lipka, V.; Teichmann, T. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium longisporum. Am. Phytopath. Soc. 2013, 26, 850–860. [Google Scholar] [CrossRef] [Green Version]
- Koprna, R.; Humplík, J.F.; Špíšek, Z.; Bryksová, M.; Zatloukal, M.; Mik, V.; Novák, O.; Nisler, J.; Doležal, K. Improvement of tillering and grain yield by application of cytokinin derivatives in wheat and barley. Agronomy 2021, 11, 67. [Google Scholar] [CrossRef]
- Van Voorthuizen, M.J. Novel plant growth regulator effects on yield, senescence and cytokinin homeostasis in wheat, barley and rapid cycling Brassica rapa. Ph.D. Thesis, University of Canterbury, Christchurch, New Zealand, 2018. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. Konzak: “A decimal code for the growth stages of cereals”. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Berková, V.; Kameniarová, M.; Ondrisková, V.; Berka, M.; Menšíková, S.; Kopecká, R.; Luklová, M.; Novák, J.; Spíchal, L.; Rashotte, A.M.; et al. Arabidopsis response to inhibitor of cytokinin degradation INCYDE: Modulations of cytokinin signaling and plant proteome. Plants 2020, 9, 1563. [Google Scholar] [CrossRef] [PubMed]
- Aremu, A.O.; Bairu, M.W.; Novák, O.; Plačková, L.; Zatloukal, M.; Doležal, K.; Finnie, J.F.; Strnad, M.; Van Staden, J. Physiological responses and endogenous cytokinin profiles of tissue-cultured “Williams” bananas in relation to roscovitine and an inhibitor of cytokinin oxidase/dehydrogenase (INCYDE) treatments. Planta 2012, 236, 1775–1790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Wang, J.Q.; Huang, Z.L.; Mi, L.; Xu, K.F.; Wu, J.J.; Fan, Y.H.; Ma, S.Y.; Jiang, D.G. Effects of low temperature at booting stage on sucrose metabolism and endogenous hormone contents in winter wheat spikelet. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Erena, M.F.; Lohraseb, I.; Munoz-Santa, I.; Taylor, J.D.; Emebiri, L.C.; Collins, N.C. The WtmsDW locus on wheat chromosome 2B controls major natural variation for floret sterility responses to heat stress at booting stage. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Jameson, P.E.; McWha, J.A.; Wright, G.J. Cytokinins and changes in their activity during the development of grains of wheat (Triticum aestivum L.). Zeit. Pflanzenphysiol. 1982, 106, 27–36. [Google Scholar] [CrossRef]
- Lenton, J.R.; Appleford, N.E.J. Cytokinins and Early Growth in Wheat; Monograph, British Plant Growth Regulator Group: Wantage, UK, 1986; Volume 14, pp. 99–113. [Google Scholar]
- Banowetz, G.M.; Ammar, K.; Chen, D.D. Postanthesis temperatures influence cytokinin accumulation and wheat kernel weight. Plant Cell Environ. 1999, 22, 309–316. [Google Scholar] [CrossRef]
- Hess, J.R.; Carman, J.G.; Banowetz, G.M. Hormones in wheat kernels during embryony. J. Plant Physiol. 2002, 159, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.N.; Perry, L.; Kisiala, A.; Olechowski, H.; Emery, R.N. Cytokinin activity during early kernel development corresponds positively with yield potential and later stage ABA accumulation in field-grown wheat (Triticum aestivum L.). Planta 2020, 252, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Faix, B.; Radchuk, V.; Nerlich, A.; Hümmer, C.; Radchuk, R.; Emery, R.J.N.; Keller, H.; Götz, K.-P.; Weschke, W.; Geigenberger, P.; et al. Barley grains, deficient in cytosolic small subunit of ADP-glucose pyrophosphorylase, reveal coordinate adjustment of C: N metabolism mediated by an overlapping metabolic-hormonal control. Plant J. 2012, 69, 1077–1093. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.F.; Paleczny, A.R.; Olechowski, H.; Emery, R.N. Changes in cytokinin form and concentration in developing kernels correspond with variation in yield among field-grown barley cultivars. Plant Physiol. Biochem. 2013, 64, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Motyka, V.; Faiss, M.; Strnad, M.; Kamínek, M.; Schmulling, T. Changes in cytokinin content and cytokinin oxidase activity in response to derepression of ipt gene transcription in transgenic tobacco calli and plants. Plant Physiol. 1996, 112, 1035–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motyka, V.; Vaňková, R.; Čapková, V.; Petrášek, J.; Kamínek, M.; Schmülling, T. Cytokinin-induced upregulation of cytokinin oxidase activity in tobacco includes changes in enzyme glycosylation and secretion. Physiol. Plant 2003, 117, 11–21. [Google Scholar] [CrossRef]
- Brugière, N.; Jiao, S.; Hantke, S.; Zinselmeier, C.; Roessler, J.A.; Niu, X.; Jones, R.J.; Habben, J.E. Cytokinin oxidase gene expression in maize is localized to the vasculature, and is induced by cytokinins, abscisic acid, and abiotic stress. Plant Physiol. 2003, 132, 1228–1240. [Google Scholar] [CrossRef] [Green Version]
- Blagoeva, E.; Dobrev, P.I.; Malbeck, J.; Motyka, V.; Gaudinová, A.; Vaňková, R. Effect of exogenous cytokinins, auxins and adenine on cytokinin N-glucosylation and cytokinin oxidase/dehydrogenase activity in de-rooted radish seedlings. Plant Growth Regul. 2004, 44, 15–23. [Google Scholar] [CrossRef]
- Hirose, N.; Takei, K.; Kuroha, T.; Kamada-Nobusada, T.; Hayashi, H.; Sakakibara, H. Regulation of cytokinin biosynthesis, compartmentalization and translocation. J. Exp. Bot. 2008, 59, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Vyroubalová, Š.; Václavíková, K.; Turečková, V.; Novák, O.; Šmehilová, M.; Hluska, T.; Ohnoutková, L.; Frébort, I.; Galuszka, P. Characterization of new maize genes putatively involved in cytokinin metabolism and their expression during osmotic stress in relation to cytokinin levels. Plant Physiol. 2009, 151, 433–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparis, S.; Przyborowski, M.; Kała, M.; Nadolska-Orczyk, A. Knockout of the HvCKX1 or HvCKX3 gene in barley (Hordeum vulgare L.) by RNA-Guided Cas9 nuclease affects the regulation of cytokinin metabolism and root morphology. Cells 2019, 8, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Bai, W.; Zeng, Q.; Song, S.; Zhang, M.; Li, X.; Hou, L.; Xiao, Y.; Luo, M.; Li, D.; et al. Moderately enhancing cytokinin level by down-regulation of GhCKX expression in cotton concurrently increases fiber and seed yield. Mol. Breed. 2015, 35, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Song, G.; Gao, J.; Zhang, S.; Zhang, R.; Li, W.; Chen, M.; Liu, M.; Xia, X.; Risacher, T.; et al. Enhancement of grain number per spike by RNA interference of cytokinin oxidase 2 gene in bread wheat. Hereditas 2018, 155, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalewski, W.; Galuszka, P.; Gasparis, S.; Orczyk, W.; Nadolska-Orczyk, A. Silencing of the HvCKX1 gene decreases the cytokinin oxidase/dehydrogenase level in barley and leads to higher plant productivity. J. Exp. Bot. 2010, 61, 1839–1851. [Google Scholar] [CrossRef] [Green Version]
- Zalewski, W.; Gasparis, S.; Boczkowska, M.; Rajchel, I.K.; Kała, M.; Orczyk, W.; Nadolska-Orczyk, A. Expression patterns of HvCKX genes indicate their role in growth and reproductive development of barley. PLoS ONE 2014, 9, e115729. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.Y.; Chen, H.W.; Ng, C.Y.; Lin, C.Y.; Tseng, T.H.; Li, W.H.; Ku, M.S. Down-regulation of cytokinin oxidase 2 expression increases tiller number and improves rice yield. Rice 2015, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Uauy, C.; Wulff, B.B.; Dubcovsky, J. Combining traditional mutagenesis with new high-throughput sequencing and genome editing to reveal hidden variation in polyploid wheat. Annu. Rev. Genet. 2017, 51, 435–454. [Google Scholar] [CrossRef] [Green Version]
- Gil, J.; Andrade-Martínez, J.S.; Duitama, J. Accurate, efficient and user-friendly mutation calling and sample identification for TILLING experiments. Front. Genet. 2021, 12, 624513. [Google Scholar] [CrossRef]
- Lassoued, R.; Phillips, P.W.; Macall, D.M.; Hesseln, H.; Smyth, S.J. Expert opinions on the regulation of plant genome editing. Plant Biotechnol. J. 2021, 19, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Wheat Tilling. Available online: https://wheat-tilling.com/ (accessed on 25 October 2021).
- Krasileva, K.V.; Vasquez-Gross, H.A.; Howell, T.; Bailey, P.; Paraiso, F.; Clissold, L.; Dubcovsky, J. Uncovering hidden variation in polyploid wheat. Proc. Natl. Acad. Sci. USA 2017, 114, E913–E921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Jiang, L.; Jameson, P.E. Co-ordinate regulation of cytokinin gene family members during flag leaf and reproductive development in wheat. BMC Plant Biol. 2012, 12, 78. [Google Scholar] [CrossRef] [Green Version]
- Svačinová, J.; Novák, O.; Plačková, L.; Lenobel, R.; Holík, J.; Strnad, M.; Doležal, K. A new approach for cytokinin isolation from Arabidopsis tissues using miniaturized purification: Pipette tip solid-phase extraction. Plant Methods 2012, 8, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Structures of plant growth regulators. (a) Thidiazuron. (b) CPPU. (c) TD-K. (d) INCYDE.


{kind=link}
Table 1.
Yield and protein composition in wheat (cv. Orator and cv. Torch) and barley (cv. Quench).
| Wheat Cultivar Orator (2013/14) | |||
| Treatment | Yield (T/ha) | TGW (g) | Protein (%) |
| Untreated Control | 10.8 ± 0.2 | 46.0 ± 1.1 | 11.2 ± 0.2 |
| DMSO 50 µM Control (GS 39, 51, 61, 65) | 11.1 ± 0.1 | 46.6 ± 0.2 | 11.2 ± 0.1 |
| DMSO 25 µM Control (GS 61, 65, 65 + 13 d) | 11.2 ± 0.1 | 47.2 ± 0.7 | 11.0 ± 0.1 |
| INCYDE 10 µM (GS 65) | 11.4 ± 0.2 | 45.9 ± 1.1 | 11.4 ± 0 |
| INCYDE 25 µM (GS 39, 51, 61, 65) | 11.3 ± 0.2 | 46.2 ± 0.7 | 11.1 ± 0.1 |
| INCYDE 25 µM (GS 39) | 11.2 ± 0.2 | 45.0 ± 1.1 | 11.2 ± 0.1 |
| INCYDE 25 µM (GS 51) | 11.0 ± 0.1 | 45.4 ± 0.8 | 11.1 ± 0.2 |
| INCYDE 25 µM (GS 61) | 11.1 ± 0.2 | 45.6 ± 0.4 | 11.1 ± 0.1 |
| INCYDE 25 µM (GS 65) | 11.1 ± 0.2 | 45.1 ± 0.5 | 11.2 ± 0.1 |
| INCYDE 50 µM (GS 61) | 11.1 ± 0.1 | 45.5 ± 1.2 | 11.4 ± 0.1 |
| INCYDE 50 µM (GS 65) | 11.1 ± 0.2 | 46.9 ± 0.3 | 11.4 ± 0.1 |
| TD-K 10 µM (GS 61, 65, 65 + 13 d) | 11.1 ± 0.1 | 46.3 ± 1.1 | 11.1 ± 0.1 |
| TD-K 25 µM (GS 61, 65, 65 + 13 d) | 11.4 ± 0.1 | 46.0 ± 0.5 | 11.1 ± 0.1 |
| Wheat Cultivar Torch (2014/15) | |||
| Treatment | Yield (T/ha) | TGW (g) | Protein (%) |
| Untreated Control | 14.5 ± 0.1 | 46.8 ± 1.1 | 9.8 ± 0.2 |
| DMSO Control (GS 51, 61, 65, 65 + 15 d) | 14.5 ± 0.3 | 48.1 ± 0.2 | 9.9 ± 0.2 |
| TD-K 10 µM (GS 61, 65, 65 + 15 d) | 14.5 ± 0.2 | 47.4 ± 0.5 | 10.0 ± 0.2 |
| TD-K 50 µM (GS 61, 65, 65 + 15 d) | 14.7 ± 0.3 | 49.1 ± 0.5 | 9.8 ± 0.05 |
| CPPU 10 µM (GS 61, 65) | 14.4 ± 0.3 | 47.1 ± 0.4 | 10.0 ± 0.2 |
| CPPU 30 µM (GS 61, 65) | 14.5 ± 0.3 | 48.8 ± 0.2 | 9.7 ± 0.1 |
| CPPU 100 µM (GS 61, 65) | 14.6 ± 0.2 | 48.4 ± 0.3 | 9.7 ± 0.3 |
| CPPU 10 µM (GS 51, 65) | 14.5 ± 0.2 | 47.8 ± 0.4 | 9.6 ± 0.1 |
| CPPU 30 µM (GS 51, 65) | 14.7 ± 0.1 | 47.2 ± 0.9 | 9.9 ± 0.1 |
| CPPU 100 µM (GS 51, 65) | 14.4 ± 0.2 | 48.1 ± 0.8 | 9.9 ± 0.2 |
| Barley Cultivar Quench (2014/15) | |||
| Treatment | Yield (T/ha) | TGW (g) | Protein (%) |
| Untreated Control | 10.8 ± 0.2 | 52.3 ± 0.5 | 14.3 ± 0.1 |
| DMSO Control (GS 51, 61, 65, 65 + 15 d) | 11.0 ± 0.1 | 52.0 ± 0.4 | 13.9 ± 0.4 |
| INCYDE 10 µM (GS 65) | 11.0 ± 0.2 | 53.0 ± 0.2 | 13.8 ± 0.3 |
| INCYDE 25 µM (GS 39, 51, 61, 65) | 11.2 ± 0.2 | 51.8 ± 0.5 | 14.0 ± 0.2 |
| INCYDE 25 µM (GS 39) | 11.3 ± 0.2 | 52.5 ± 0.5 | 13.8 ± 0.2 |
| INCYDE 25 µM (GS 51) | 11.3 ± 0.1 | 52.4 ± 0.9 | 14.0 ± 0.4 |
| INCYDE 25 µM (GS 61) | 11.1 ± 0.1 | 52.2 ± 0.9 | 13.9 ± 0.5 |
| INCYDE 25 µM (GS 65) | 11.3 ± 0.1 | 52.5 ± 0.7 | 14.2 ± 0.4 |
| INCYDE 50 µM (GS 61) | 11.2 ± 0.1 | 52.5 ± 0.8 | 14.1 ± 0.4 |
| INCYDE 50 µM (GS 65) | 11.2 ± 0.3 | 52.3 ± 0.4 | 13.9 ± 0.5 |
Data were analyzed using an ANOVA, with protein percentage data logit-transformed prior to ANOVA. Data are presented as the means ± standard error (n = 4). Yield is provided in tonnes per hectare (T/ha), thousand grain weight (TGW) in grams (g) and protein as a percentage (%). Concentration of each treatment is given in µM, with growth stage (GS) indicating the growth stage (Zadoks scale [82]) targeted for treatment, and ‘d’ indicating the number of days after the respective growth stage. The dimethylsulfoxide (DMSO) controls list the GS targeted, with volumes equivalent to the DMSO used in the highest concentration within each field trial, with the exception of Orator (2013/14), where DMSO Control (GS 61, 65, 65 + 13 d) was provided at a volume equivalent to 25 µM applications.
Table 2.
LC–MS/MS analyses of the quantity of cytokinins in wheat (cultivar Torch, 2014/15) grains treated at anthesis with TD-K or CPPU. Measurements were made at four days after anthesis.
Table 2.
LC–MS/MS analyses of the quantity of cytokinins in wheat (cultivar Torch, 2014/15) grains treated at anthesis with TD-K or CPPU. Measurements were made at four days after anthesis.
| Wheat Cytokinin Concentrations (pmol/g DW) | |||
|---|---|---|---|
| Type | Control | TD-K 50 µM | CPPU 100 µM |
| tZ | 794.5 ± 71.1 | 705.3 ± 75.7 | 889.2 ± 73.7 |
| tZR | 60.0 ± 6.7 | 60.3 ± 3.2 | 63.5 ± 2.3 |
| tZOG | 20.4 ± 2.8 | 24.7 ± 0.5 | 22.8 ± 1.3 |
| tZROG | 5.8 ± 0.7 | 6.8 ± 0.2 | 6.5 ± 0.5 |
| tZRMP | 115.9 ± 3.6 | 106.5 ± 2.8 | 103.1 ± 10.5 |
| tZ7G | <LOD | <LOD | <LOD |
| tZ9G | 247.3 ± 21.3 | 286.1 ± 3.5 | 268.7 ± 11.5 |
| Total tZ types | 1244.0 ± 104.8 | 1189.7 ± 85.2 | 1353.8 ± 57.0 |
| iP | 1.6 ± 0.2 | 1.3 ± 0.1 | 1.3 ± 0.1 |
| iPR | 2.2 ± 0.2 | 1.8 ± 0.1 | 2.5 ± 0.4 |
| iPRMP | 22.9 ± 1.5 | 22.1 ± 3.8 | 25.9 ± 4.0 |
| iP7G | <LOD | <LOD | <LOD |
| iP9G | <LOD | <LOD | <LOD |
| Total iP types | 26.8 ± 1.7 | 25.1 ± 3.9 | 29.7 ± 4.1 |
| cZ | 9.6 ± 0.5 | 7.8 ± 1.1 | 8.6 ± 1.2 |
| cZR | 33.9 ± 3.9 | 24.7 ± 3.5 | 30.5 ± 4.0 |
| cZOG | 114.3 ± 12.4 | 130.8 ± 11.7 | 116.1 ± 12.9 |
| cZROG | 139.1 ± 13.2 | 154.5 ± 6.6 | 150.6 ± 8.9 |
| cZRMP | 10.0 ± 0.8 | 7.7 ± 1.3 | 10.8 ± 1.2 |
| cZ9G | <LOD | <LOD | <LOD |
| Total cZ types | 306.5 ± 26.5 | 325.5 ± 12.2 | 316.5 ± 17.0 |
| DHZ | 0.23 ± 0.01 | 0.20 ± 0.03 | 0.20 ± 0.03 |
| DHZR | 2.9 ± 0.1 | 2.4 ± 0.3 | 2.6 ± 0.3 |
| DHZOG | 1.4 ± 0.2 | 1.7 ± 0.1 | 1.5 ± 0.1 |
| DHZROG | 9.5 ± 1.1 | 10.7 ± 0.6 | 10.0 ± 0.8 |
| DHZRMP | <LOD | <LOD | <LOD |
| DHZ7G | 15.0 ± 0.3 | 13.3 ± 0.9 | 14.6 ± 2.2 |
| DHZ9G | 0.07 ± 0.003 | 0.06 ± 0.01 | 0.07 ± 0.01 |
| Total DHZ types | 29.1 ± 1.5 | 28.2 ± 0.7 | 28.9 ± 2.6 |
| Total CK bases | 806.0 ± 71.4 | 714.6 ± 76.9 | 899.3 ± 74.9 |
| Total CK ribosides | 98.7 ± 6.8 | 89.1 ± 7.0 | 99.0 ± 7.0 |
| Total CK nucleotides | 148.9 ± 3.7 | 136.3 ± 2.0 | 139.8 ± 12.8 |
| Total CK O-glucosides | 290.5 ± 30.2 | 329.2 ± 18.8 | 307.5 ± 23.7 |
| Total CK N-glucosides | 262.3 ± 21.6 | 299.4 ± 3.9 | 283.4 ± 13.6 |
| Total cytokinins | 1606.4 ± 117.3 | 1568.5 ± 69.5 | 1728.9 ± 43.4 |
Treatments were compared to the control using a two-sided ANOVA. Data are presented as the means ± standard error (n = 3). LOD indicates below limit of detection. Treatments were made at anthesis (GS 60). Cytokinin abbreviations: CK (cytokinins), tZ (trans-zeatin), iP (N6-isopentenyladenine), cZ (cis-zeatin), DHZ (dihydrozeatin), R (riboside), OG (O-glucoside), RMP (riboside-5′-monophosphate), 7G (7-N-glucoside), 9G (9-N-glucoside).
Table 3.
LC–MS/MS analyses of the quantity of cytokinins in barley (cultivar Quench, 2014/15) grains treated at anthesis with INCYDE. Measurements were made at four days after anthesis.
Table 3.
LC–MS/MS analyses of the quantity of cytokinins in barley (cultivar Quench, 2014/15) grains treated at anthesis with INCYDE. Measurements were made at four days after anthesis.
| Barley Cytokinin Concentrations (pmol/g DW) | ||
|---|---|---|
| Type | Control | INCYDE 50 µM |
| tZ | 759.5 ± 66.8 | 642.8 ± 30.0 |
| tZR | 432.0 ± 44.3 | 458.9 ± 36.6 |
| tZOG | 89.5 ± 2.7 | 97.6 ± 7.9 |
| tZROG | 28.1 ± 1.8 | 30.2 ± 0.6 |
| tZRMP | 455.3 ± 27.0 | 454.6 ± 13.0 |
| tZ7G | <LOD | <LOD |
| tZ9G | 46.2 ± 2.6 | 65.0 ± 9.1 |
| Total tZ types | 1810.6 ± 142.1 | 1749.1 ± 61.6 |
| iP | 2.0 ± 0.2 | 2.3 ± 0.3 |
| iPR | 4.5 ± 0.6 | 4.7 ± 0.3 |
| iPRMP | 86.8 ± 12.8 | 89.0 ± 11.9 |
| iP7G | <LOD | <LOD |
| iP9G | <LOD | <LOD |
| Total iP types | 93.3 ± 13.6 | 96.0 ± 12.2 |
| cZ | 3.6 ± 0.1 | 3.2 ± 0.1 |
| cZR | 23.4 ± 0.5 | 20.1 ± 0.5 * |
| cZOG | 328.9 ± 11.8 | 417.1 ± 9.7 * |
| cZROG | 242.5 ± 8.4 | 256.8 ± 8.6 |
| cZRMP | 20.0 ± 1.8 | 20.6 ± 1.9 |
| cZ9G | <LOD | <LOD |
| Total cZ types | 618.4 ± 18.8 | 717.9 ± 12.8 * |
| DHZ | 0.9 ± 0.02 | 0.7 ± 0.04 * |
| DHZR | 6.5 ± 0.1 | 5.1 ± 0.3 * |
| DHZOG | 12.6 ± 0.6 | 10.6 ± 0.4 * |
| DHZROG | 47.3 ± 3.1 | 43.0 ± 2.8 |
| DHZRMP | <LOD | <LOD |
| DHZ7G | 6.0 ± 0.3 | 5.0 ± 0.7 |
| DHZ9G | 0.06 ± 0.01 | 0.06 ± 0.01 |
| Total DHZ types | 73.0 ± 3.7 | 64.5 ± 3.8 |
| Total CK bases | 766.0 ± 67.0 | 649.0 ± 30.4 |
| Total CK ribosides | 466.3 ± 44.6 | 488.8 ± 37.1 |
| Total CK nucleotides | 562.2 ± 38.6 | 564.2 ± 11.1 |
| Total CK O-glucosides | 748.9 ± 27.8 | 855.4 ± 25.2 * |
| Total CK N-glucosides | 51.9 ± 2.6 | 70.1 ± 8.7 |
| Total cytokinins | 2595.2 ± 177.1 | 2627.5 ± 68.9 |
* Indicates a statistically significant (p ≤ 0.05) difference for the treatment compared to the control using a two-sided ANOVA and post hoc two-sided Dunnett test (CI: 95%). Significant differences are provided in bold. Data are presented as the means ± standard error (n = 3). LOD indicates below limit of detection. Treatments were made at anthesis (GS 60). Cytokinin abbreviations: CK (cytokinins), tZ (trans-zeatin), iP (N6-isopentenyladenine), cZ (cis-zeatin), DHZ (dihydrozeatin), R (riboside), OG (O-glucoside), RMP (riboside-5′-monophosphate), 7G (7-N-glucoside), 9G (9-N-glucoside).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
van Voorthuizen, M.J.; Song, J.; Novák, O.; Jameson, P.E. Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials. Plants 2021, 10, 2309. https://doi.org/10.3390/plants10112309
AMA Style
van Voorthuizen MJ, Song J, Novák O, Jameson PE. Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials. Plants. 2021; 10(11):2309. https://doi.org/10.3390/plants10112309
Chicago/Turabian Stylevan Voorthuizen, Matthew J., Jiancheng Song, Ondřej Novák, and Paula E. Jameson. 2021. "Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials" Plants 10, no. 11: 2309. https://doi.org/10.3390/plants10112309
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.