
Priming Maritime Pine Megagametophytes during Somatic Embryogenesis Improved Plant Adaptation to Heat Stress

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effect of Priming on Maritime Pine Somatic Embryogenesis
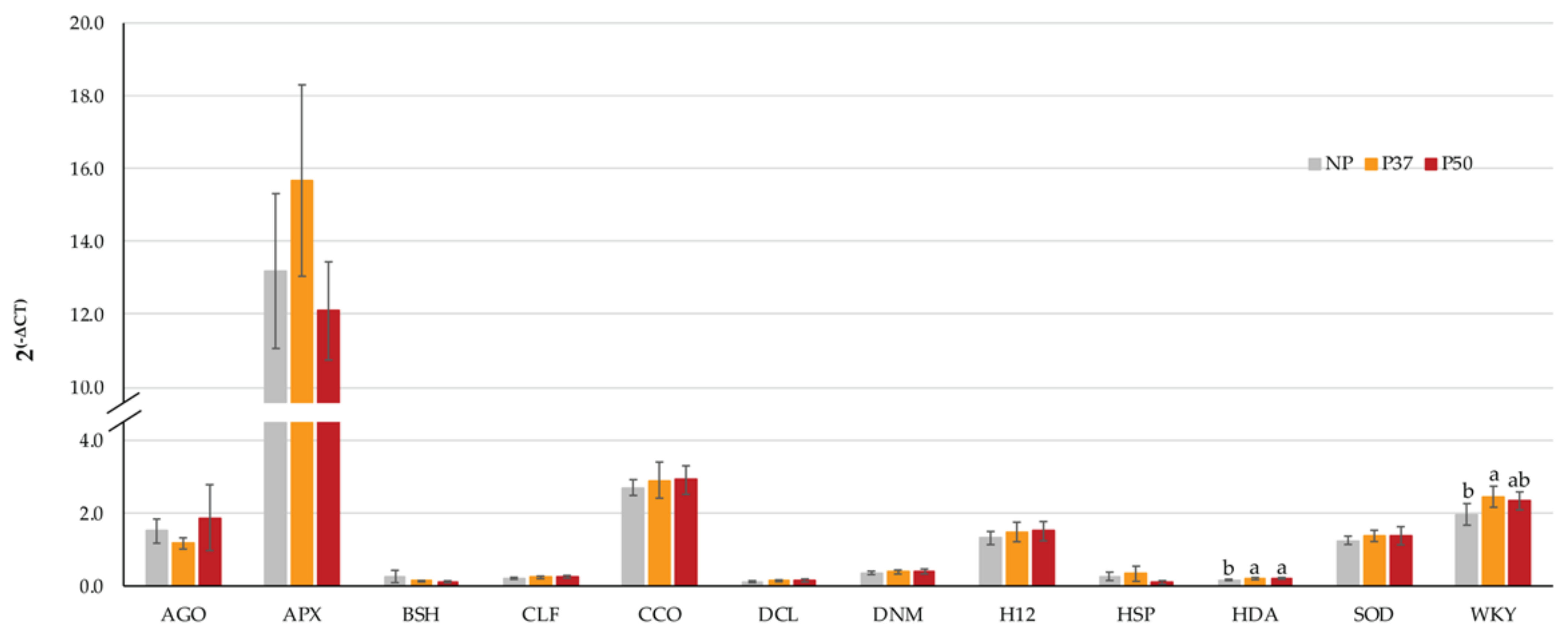
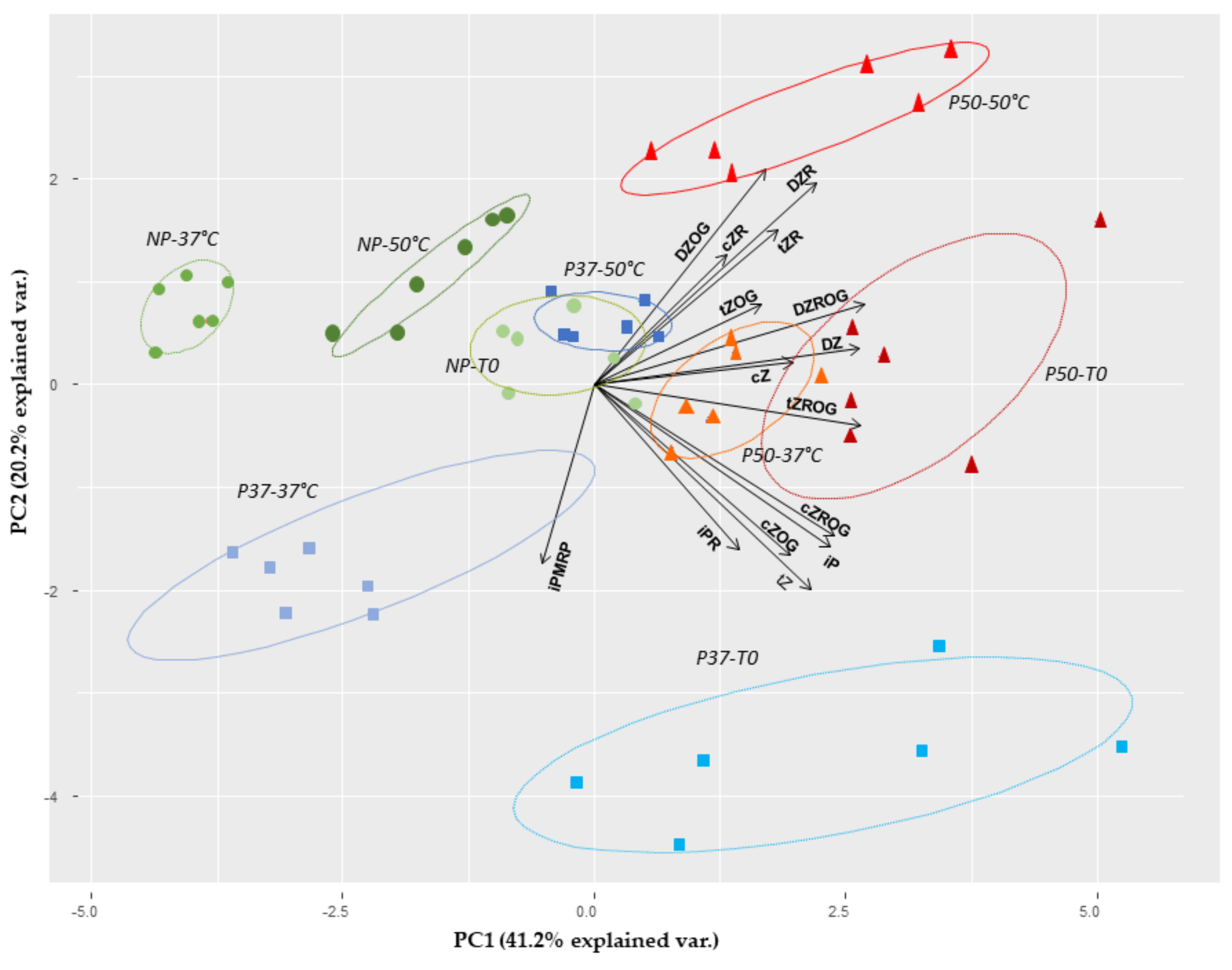
2.2. Effect of Priming on Gene Expression Profile of Embryogenic Lines
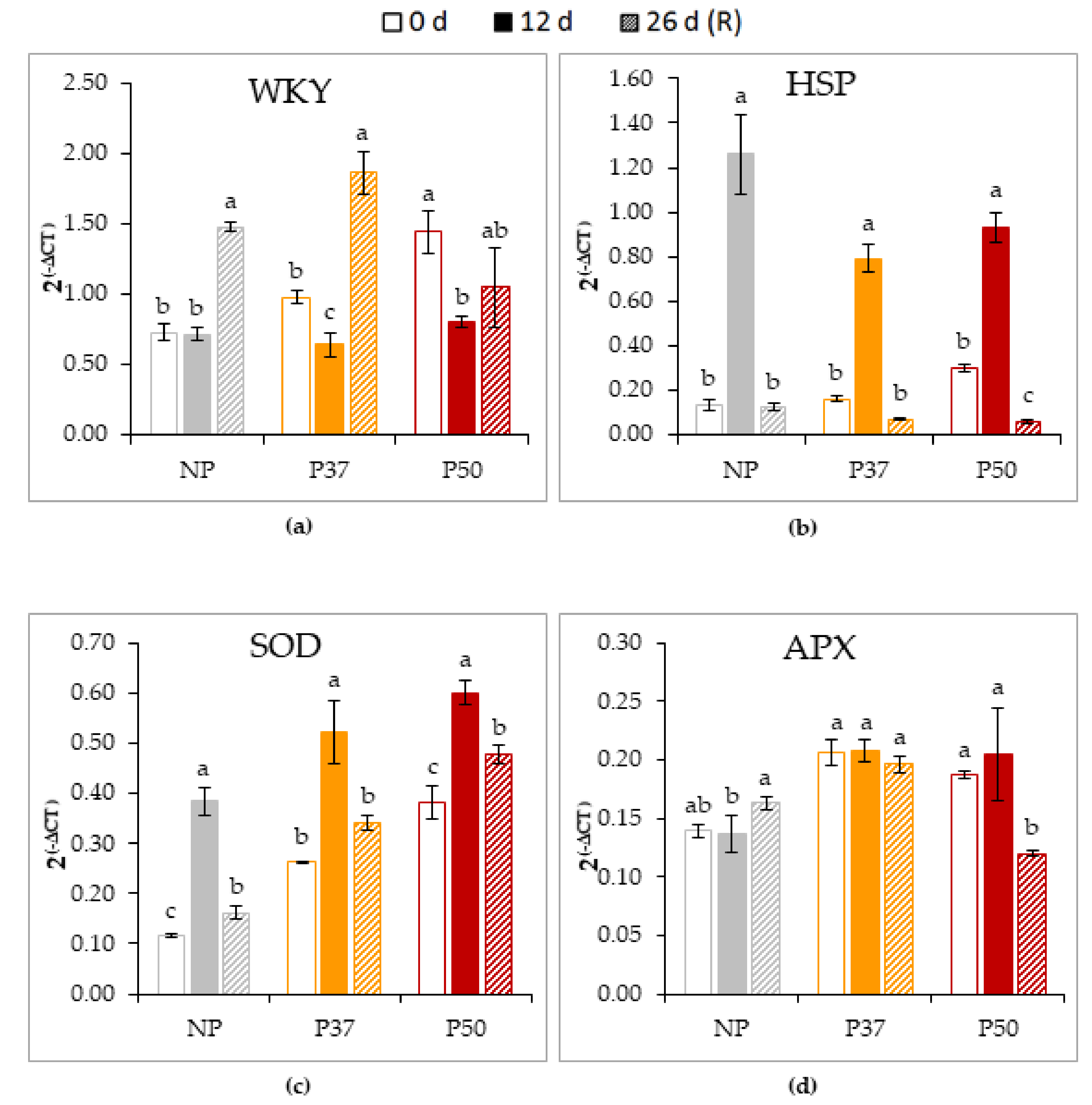
2.3. Response to Heat Stress of in Vitro Growing Maritime Pine Plants Derived from Primed Embryogenic Lines
2.4. Response of Primed Maritime Pine Plants to High Temperature Stress in the Greenhouse
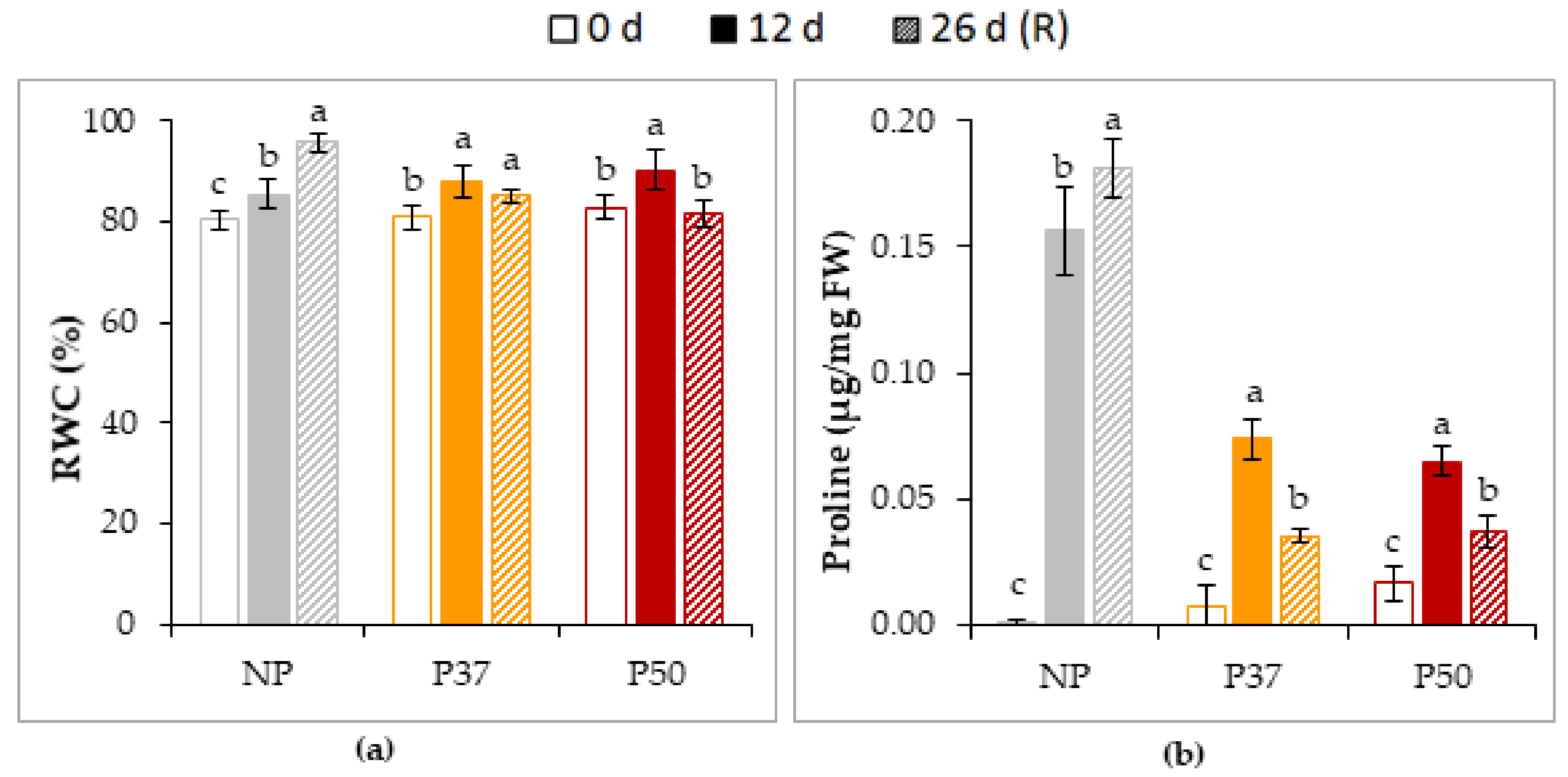
2.4.1. Osmotic Adjustment
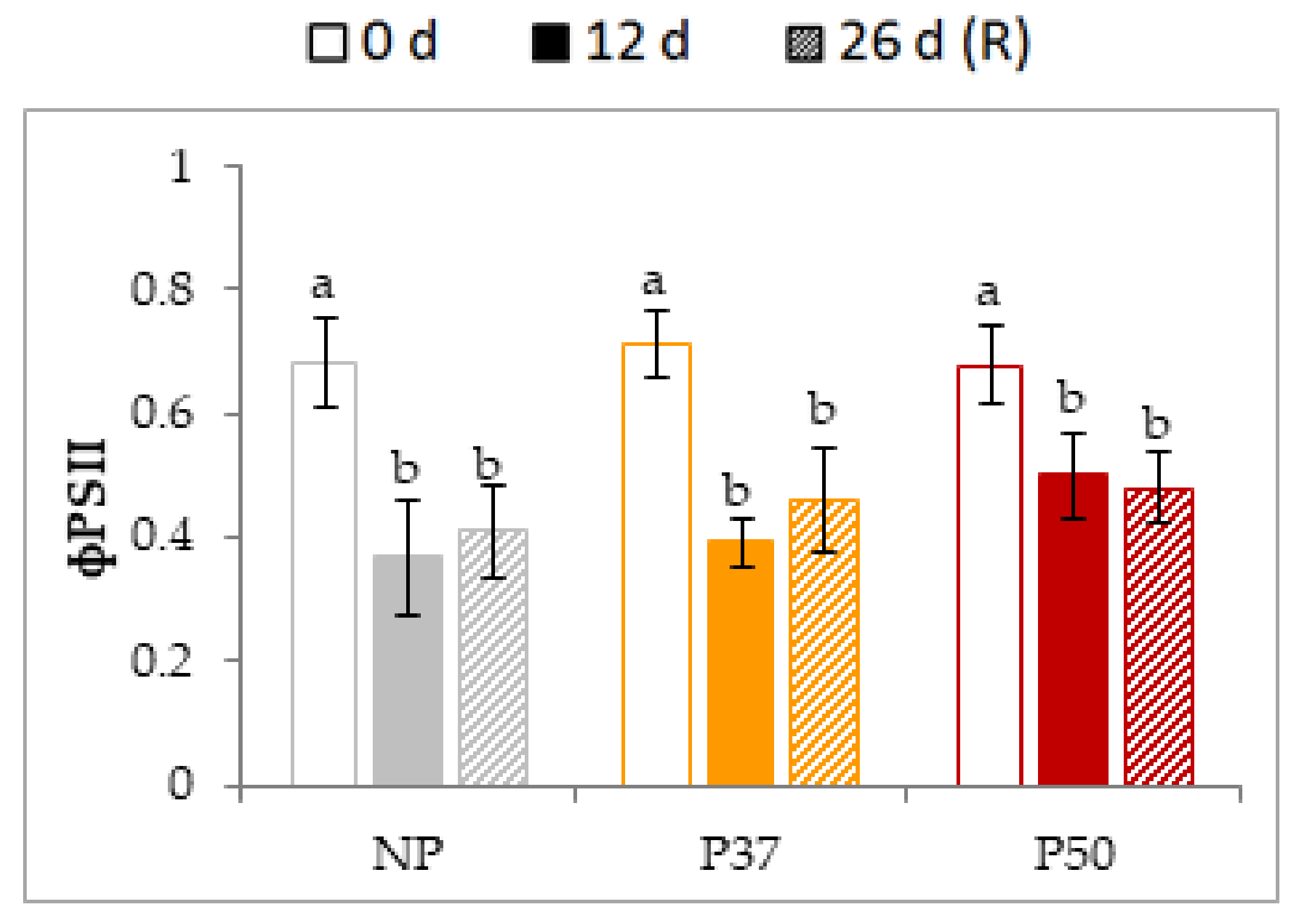
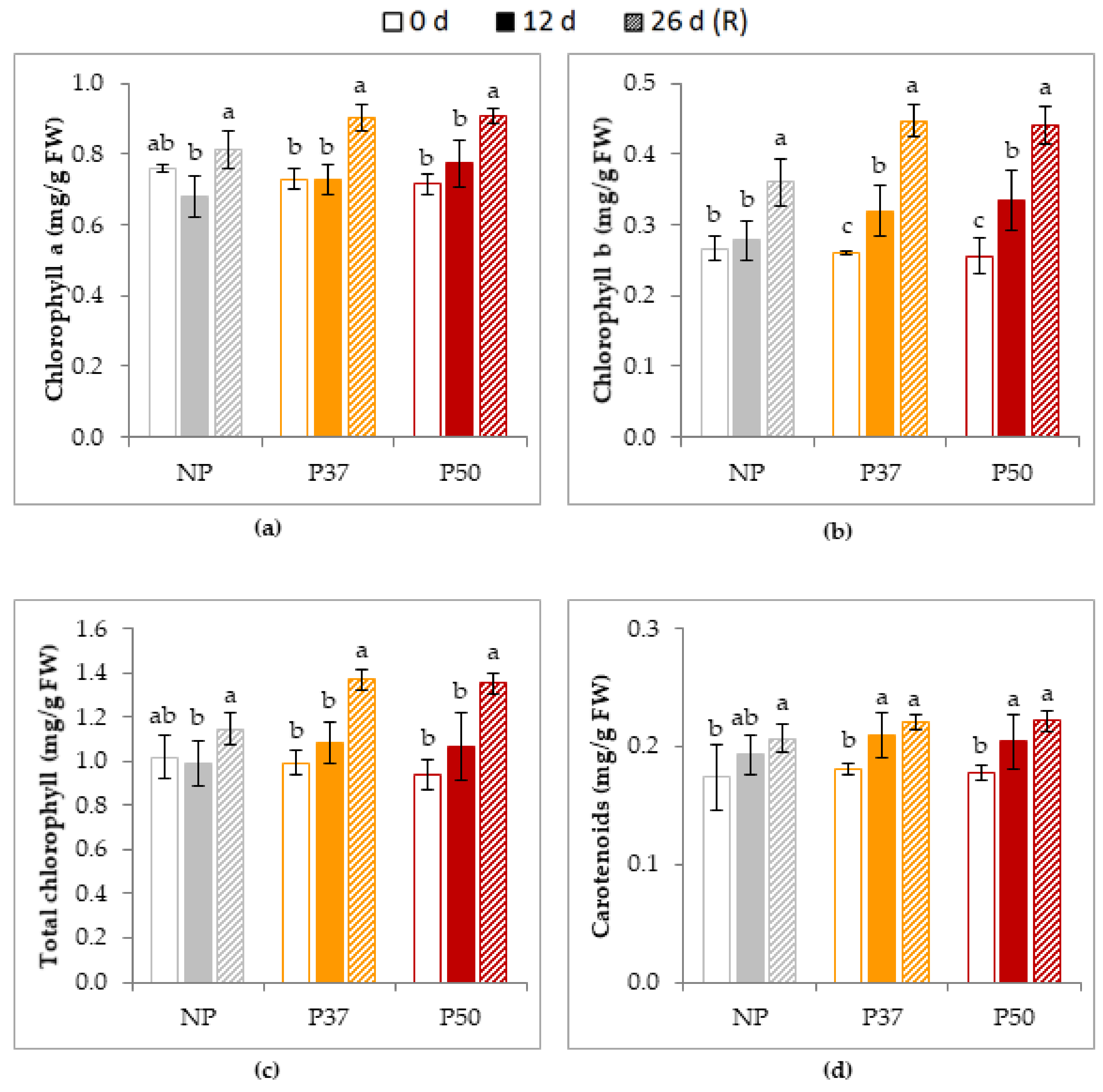
2.4.2. Photosynthetic Parameters
3. Discussion
4. Materials and Methods
4.1. Plant Material and Production of Primed Plants
4.2. Heat Stress Experiments
4.3. Gene Expression Analyses
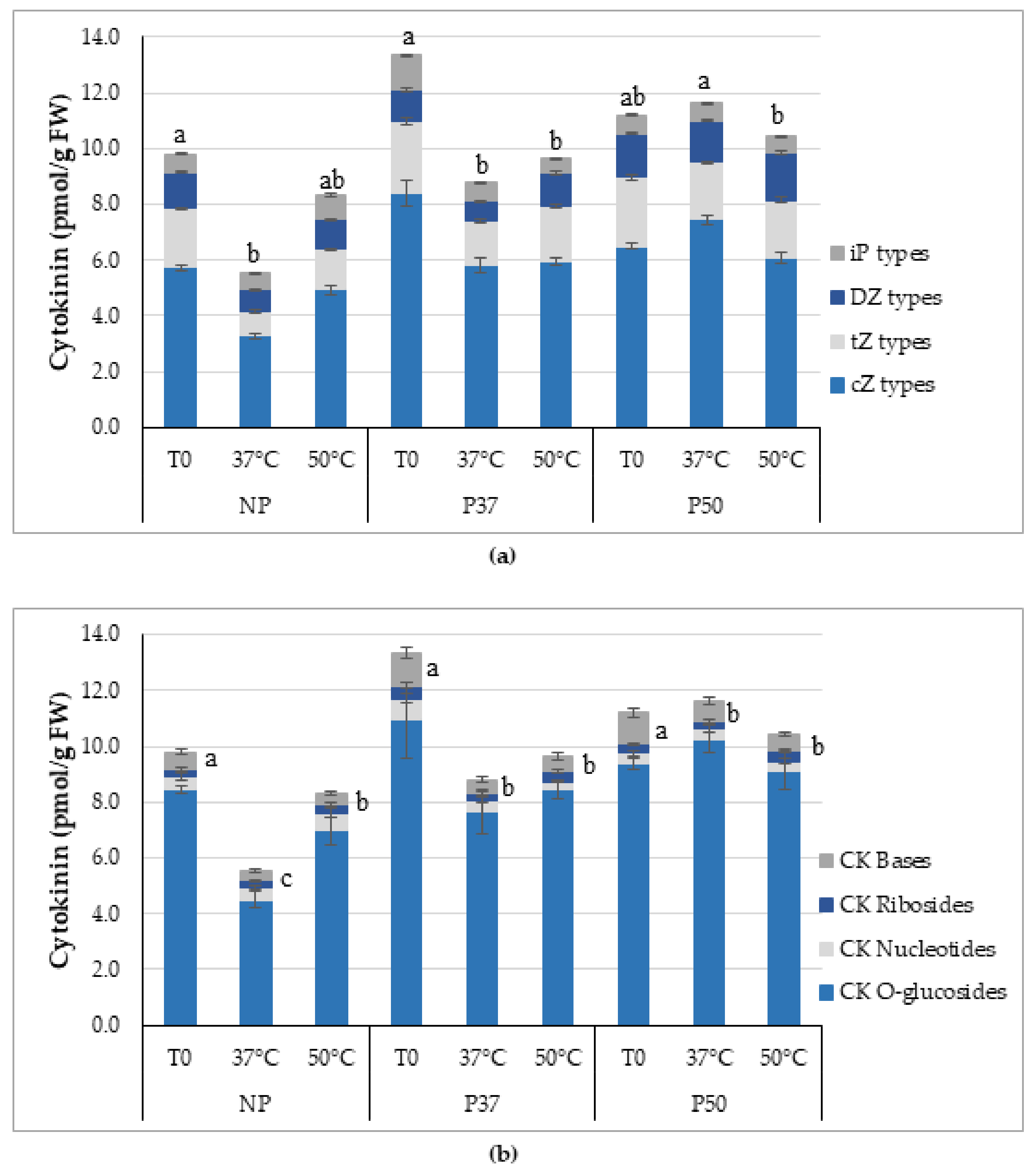
4.4. Plant Hormone Analyses
4.5. Characterization of 2-Year-Old Maritime Pine Plants after Heat Stress in Greenhouse Conditions
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Fitzgerald, J.; Jacobsen, J.B.; Blennow, K.; Thorsen, B.J.; Lindner, M. Climate change in European forests: How to adapt. In EFI Policy Brief; Fitzgerald, J., Jacobsen, J.B., Blennow, K., Thorsen, B.J., Lindner, M., Eds.; European Forest Institute: Joensuu, Finland, 2013; p. 16. [Google Scholar]
- Lebedev, V.G.; Lebedeva, T.N.; Chernodubov, A.I.; Shestibratov, K.A. Genomic selection for forest tree improvement: Methods, achievements and perspectives. Forests 2020, 11, 1190. [Google Scholar] [CrossRef]
- Bose, A.K.; Moser, B.; Rigling, A.; Lehmann, M.M.; Milcu, A.; Peter, M.; Rellstab, C.; Wohlgemuth, T.; Gessler, A. Memory of environmental conditions across generations affects the acclimation potential of scots pine. Plant Cell Environ. 2020, 43, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Turgut-Kara, N.; Arikan, B.; Celik, H. Epigenetic memory and priming in plants. Genetica 2020, 148, 47–54. [Google Scholar] [CrossRef]
- Galviz, Y.C.F.; Ribeiro, R.V.; Souza, G.M. Yes, plants do have memory. Theor. Exp. Plant Physiol. 2020, 32, 195–202. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, N.; Virlouvet, L.; Riethoven, J.J.; Fromm, M.; Avramova, Z. Four distinct types of dehydration stress memory genes in Arabidopsis thaliana. BMC Plant Biol. 2013, 13, 229. [Google Scholar] [CrossRef] [Green Version]
- Yakovlev, I.A.; Carneros, E.; Lee, Y.; Olsen, J.E.; Fossdal, C.G. Transcriptional profiling of epigenetic regulators in somatic embryos during temperature induced formation of an epigenetic memory in Norway spruce. Planta 2016, 243, 1237–1249. [Google Scholar] [CrossRef]
- Schmitz, R.J.; Schultz, M.D.; Lewsey, M.G.; O’Malley, R.C.; Urich, M.A.; Libiger, O.; Schork, N.J.; Ecker, J.R. Transgenerational epigenetic instability is a source of novel methylation variants. Science 2011, 334, 369–373. [Google Scholar] [CrossRef] [Green Version]
- Raj, S.; Bräutigam, K.; Hamanishi, E.; Wilkins, O.; Thomas, B.R.; Schroeder, W.R.; Mansfield, S.D.; Plant, A.L.; Campbell, M.M. Clone history shapes Populus drought responses. Proc. Natl. Acad. Sci. USA 2011, 108, 12521–12526. [Google Scholar] [CrossRef] [Green Version]
- Yakovlev, I.A.; Fossdal, C.G.; Johnsen, Ø. MicroRNAs, the epigenetic memory and climatic adaptation in Norway spruce. New Phytol. 2010, 187, 1154–1169. [Google Scholar] [CrossRef] [PubMed]
- Baum, S.; Reimer-Michalski, E.-M.; Bolger, A.; Andrea, J.; Mantai, A.J.; Benes, V.; Usadel, B.; Conratha, U. Isolation of Open Chromatin Identifies Regulators of Systemic Acquired Resistance. Plant Physiol. 2019, 181, 817–833. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Li, Z.; Hoque, T.S.; Burritt, D.J.; Fujita, M.; Munné-Bosch, S. Heat or cold priming-induced cross-tolerance to abiotic stresses in plants: Key regulators and posible mechanisms. Protoplasma 2018, 255, 399–412. [Google Scholar] [CrossRef]
- Wang, X.; Liu, F.; Jiang, D. Priming: A promising strategy for crop production in response to future climate. J. Integr. Agric. 2017, 16, 2709–2716. [Google Scholar] [CrossRef]
- Mishra, D.; Shekhar, S.; Singh, D.; Chakraborty, S.; Chakraborty, N. Heat Shock Proteins and Abiotic Stress Tolerance in Plants. In Regulation of Heat Shock Protein Responses; Asea, A., Kaur, P., Eds.; Springer: Cham, Switzerland, 2018; pp. 41–69. [Google Scholar]
- Kurjak, D.; Konôpková, A.; Kmeť, J.; Macková, M.; Frýdl, J.; Živčák, M.; Palmroth, S.; Ditmarová, L.; Gömör, D. Variation in the performance and thermostability of photosystem II in European beech (Fagus sylvatica L.) provenances is influenced more by acclimation than by adaptation. Eur. J. For. Res. 2019, 13892, 79–92. [Google Scholar] [CrossRef]
- Marias, D.E.; Meinzer, F.C.; Woodruff, D.R.; McCulloh, K.A. Thermotolerance and heat stress responses of Douglas-fir and ponderosa pine seedling populations from contrasting climates. Tree Physiol. 2016, 37, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Escandón, M.; Cañal, M.; Pascual, J.; Pinto, G.; Correia, B.; Amaral, J.; Meijón, M. Integrated physiological and hormonal profile of heat-induced thermotolerance in Pinus radiata. Tree Physiol. 2016, 36, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Escandón, M.; Valledor, L.; Pascual, J.; Pinto, G.; Cañal, M.J.; Meijón, M. System-wide analysis of short-term response to high temperature in Pinus radiata. J. Exp. Bot. 2017, 68, 3629–3641. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Kramna, B.; Gaudinova, A.; Knirsch, V.; Spichal, L.; Zatloukal, M.; Vankova, R. Heat Acclimation and Inhibition of Cytokinin Degradation Positively Affect Heat Stress Tolerance of Arabidopsis. Front. Plant Sci. 2020, 11, 87. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162. [Google Scholar] [CrossRef] [Green Version]
- Zwack, P.J.; Rashotte, A.M. Interactions between cytokinin signalling and abiotic stress responses. J. Exp. Bot. 2015, 66, 4863–4871. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Benková, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Černý, M.; Jedelský, P.L.; Novák, J.; Schlosser, A.; Brzobohatý, B. Cytokikin modulates proteomic, transcriptomic and growth responses to temperature shocks in Arabidopsis. Plant Cell Environ. 2014, 37, 1641–1655. [Google Scholar] [CrossRef]
- Lelu-Walter, M.A.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic embryogenesis in forestry with a focus on Europe: State-of-the-art, benefits, challenges and future direction. Tree Genet. Genomes 2013, 9, 883–899. [Google Scholar] [CrossRef]
- Bonga, J.M. Can explant choice help resolve recalcitrance problems in in vitro propagation, a problem still acute especially for adult conifers? Trees 2017, 31, 781–789. [Google Scholar] [CrossRef]
- Arrillaga, I.; Morcillo, M.; Zanón, I.; Lario, F.; Segura, J.; Sales, E. New approaches to optimize somatic embryogenesis in maritime pine. Front. Plant Sci. 2019, 10, 138. [Google Scholar] [CrossRef] [Green Version]
- Humánez, A.; Blasco, M.; Brisa, C.; Segura, J.; Arrillaga, I. Somatic embryogenesis from different tissues of Spanish populations of maritime pine. Plant Cell Tissue Organ Cult. 2012, 111, 373–383. [Google Scholar] [CrossRef]
- Alía, R.; Martín, S. EUFORGEN Technical guidelines for genetic conservation and use for maritime pine (Pinus pinaster). In International Plant Genetic Resources Institute; Bioversity International: Rome, Italy, 2003; p. 6. [Google Scholar]
- Zas, R.; Sampedro, L.; Solla, A.; Vivas, M.; Lombardero, M.J.; Alía, R.; Rozas, V. Dendroecology in common gardens: Population differentiation and plasticity in resistance, recovery and resilience to extreme drought events in Pinus pinaster. Agric. For. Meteorol. 2020, 291, 108060. [Google Scholar] [CrossRef]
- García-Mendiguren, O.; Montalbán, I.A.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. Environmental conditions at the initial stages of Pinus radiata somatic embryogenesis affect the production of somatic embryos. Trees 2016, 30, 949–958. [Google Scholar] [CrossRef]
- Moncaleán, P.; García-Mendiguren, O.; Novak, O.; Strnad, M.; Goicoa, T.; Ugarte, M.D.; Montalbán, I.A. Temperature and water availability during maturation affect the cytokinins and auxins profile of radiata pine somatic embryos. Front. Plant Sci. 2018, 9, 1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Mendiguren, O.; Montalbán, I.A.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. Are we able to modulate the response of somatic embryos of pines to drought stress? Acta Hortic. 2017, 1155, 77–84. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Montalbán, I.A.; De Medeiros Oliveira, E.; Dell’Aversana, E.; D’Amelia, L.; Carillo, P.; Steiner, N.; Fraga, H.P.F.; Guerra, M.P.; Goicoa, T.; et al. Effect of thermal stress on tissue ultrastructure and metabolite profiles during initiation of radiata pine somatic embryogenesis. Front. Plant Sci. 2019, 9, 2004. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Pereira, C.; Montalbán, I.A.; Pěnčík, A.; Petřík, I.; Pavlovic, I.; Novák, O.; Strnad, M.; Moncaleán, P. Quantification of endogenous aromatic cytokinins in Pinus radiata embryonal masses after application of heat stress during initiation of somatic embryogenesis. Trees 2020. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Moncaleán, P.; Pereira, C.; Pěnčík, A.; Petřík, I.; Pavlovic, I.; Novák, O.; Strnad, M.; Goicoa, T.; Ugarte, M.D.; et al. Cytokinins are involved in drought tolerance of Pinus radiata plants originating from embryonal masses induced at high temperatures. Tree Physiol. 2020, tpaa055. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Pereira, C.; Sales, E.; Meijón, M.; Arrillaga, I.; Cañal, M.J.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P.; Montalbán, I.A. Induction of radiata pine somatic embryogenesis at high temperatures provokes a long-term decrease in DNA methylation/hydroxymethylation and differential expression of stress-related genes. Plants 2020, 9, 1762. [Google Scholar] [CrossRef]
- Do Nascimento, A.M.M.; Barroso, P.A.; Do Nascimento, N.F.F.; Goicoa, T.; Ugarte, M.D.; Montalbán, I.A.; Moncaleán, P. Pinus spp. Somatic Embryo Conversion under High Temperature: Effect on the Morphological and Physiological Characteristics of Plantlets. Forests 2020, 11, 1181. [Google Scholar] [CrossRef]
- Pereira, C.; Castander-Olarieta, A.; Montalbán, I.A.; Pěnčík, A.; Petřík, I.; Pavlovic, I.; De Medeiros Oliveira, E.; Fraga, H.; Guerra, M.P.; Novak, O.; et al. Embryonal masses induced at high temperatures in Aleppo pine: Cytokinin profile and cytological characterization. Forests 2020, 11, 807. [Google Scholar] [CrossRef]
- Vega-Bartol, J.J.; Simões, M.; Lorenz, W.W.; Rodrigues, A.S.; Alba, R.; Dean, J.F.; Miguel, C.M. Transcriptomic analysis highlights epigenetic and transcriptional regulation during zygotic embryo development of Pinus pinaster. BMC Plant Biol. 2013, 13, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, A.S.; De Vega, J.J.; Miguel, C.M. Comprehensive assembly and analysis of the transcriptome of maritime pine developing embryos. BMC Plant Biol. 2018, 18, 379. [Google Scholar] [CrossRef] [PubMed]
- Perdiguero, P.; Barbero, M.; Cervera, M.; Collada, C.; Soto, Á. MoLecular response to water stress in two contrasting Mediterranean pines (Pinus pinaster and Pinus pinea). Plant Physiol. Biochem. 2013, 67, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Cano, M.; Morcillo, A.; Humánez, A.; Mendoza-Poudereux, I.; Alborch, A.; Segura, J.; Arrillaga, I. Maritime pine (Pinus pinaster Aiton). In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants, 2nd ed.; Jain, S.M., Gupta, P., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1, pp. 167–179. [Google Scholar]
- Luo, M.; Cheng, K.; Xu, Y.; Yang, S.; Wu, K. Plant Responses to Abiotic Stress Regulated by Histone Deacetylases. Front. Plant Sci. 2017, 8, 2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.U.; Ali, A.; Khan, H.A.; Baek, D.; Park, J.; Lim, C.J.; Zareen, S.; Jan, M.; Lee, S.Y.; Pardo, J.M.; et al. PWR/HDA9/ABI4 Complex Epigenetically Regulates ABA Dependent Drought Stress Tolerance in Arabidopsis. Front. Plant Sci. 2020, 11, 623. [Google Scholar] [CrossRef] [PubMed]
- de Rooij, P.G.H.; Perrella, G.; Kaiserli, E.; Van Zanten, M. The diverse and unanticipated roles of histone deacetylase 9 in coordinating plant development and environmental acclimation. J. Exp. Bot. 2020, 71, 6211–6225. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Sinha, K.; Bhunia, R. Can wheat survive in heat? Assembling tools towards successful development of heat stress tolerance in Triticum aestivum L. Mol. Biol. Rep. 2019, 46, 2577–2593. [Google Scholar] [CrossRef]
- Wu, X.; Shiroto, Y.; Kishitani, S.; Ito, Y.; Toriyama, K. Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 2009, 28, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Azeem, F.; Nawaz, M.A.; Acet, T.; Abbas, A.; Imran, Q.M.; Shah, K.H.; Rehman, H.M.; Chhung, G.; Yang, S.H.; et al. Transcription factors WRKY11 and WRKY17 are involved in abiotic stress responses in Arabidopsis. J. Plant Physiol. 2018, 226, 12–21. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Ismail, M.R.; Malek, M.A.; Latif, M.A.; Oladosu, Y. Heat Shock Proteins: Functions And Response Against Heat Stress In Plants. Int. J. Sci. Technol. Res. 2014, 3, 204–218. [Google Scholar]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants feel the heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef]
- Lohmann, C.; Eggers-Schumacher, G.; Wunderlich, M.; Schöffl, F. Two different heat shock transcription factors regulate immediate early expression of stress genes in Arabidopsis. MoL. Genet. Genom. 2004, 271, 11–21. [Google Scholar] [CrossRef]
- Byung-Hoon, K.; Fritz, S. Interaction between Arabidopsis heat shock transcription factor 1 and 70 kDa heat shock proteins. J. Exp. Bot. 2002, 53, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.J.; Vierling, E. A small heat shock protein cooperates with heat shock protein 70 systems to reactivate a heat-denatured protein. Plant Physiol. 2000, 122, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, N.K.; Kim, Y.K.; Grover, A. Rice sHsp genes: Genomic organization and expression profiling under stress and development. BMC Genom. 2009, 10, 393. [Google Scholar] [CrossRef] [Green Version]
- Dobrá, J.; Černý, M.; Štorchová, H.; Dobrev, P.; Skalák, J.; Jedelský, P.L.; Lukšanová, H.; Gaudinová, A.; Pešek, B.; Malbeck, J.; et al. The impact of heat stress targeting on the hormonal and transcriptomic response in Arabidopsis. Plant Sci. 2015, 231, 52–61. [Google Scholar] [CrossRef]
- Schäfer, M.; Brütting, C.; Meza-Canales, I.D.; Großkinsky, D.K.; Vankova, R.; Baldwin, I.T.; Meldau, S. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J. Exp. Bot. 2015, 66, 4873–4884. [Google Scholar] [CrossRef] [Green Version]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Asensi-Fabado, M.A.; Olivan, A.; Munne-Bosch, S. A comparative study of the hormonal response to high temperatures and stress reiteration in three Labiatae species. Environ. Exp. Bot. 2013, 94, 57–65. [Google Scholar] [CrossRef]
- Gosavi, G.U.; Jadhav, A.; Kale, A.A.; Gadakh, S.R.; Pawar, B.D.; Chimote, V.P. Effect of heat stress on proline, chlorophyll content, heat shock proteins and antioxidant enzyme activity in sorghum (Sorghum bicolor) at seedlings stage. Indian J. Biotechnol. 2014, 13, 356–363. [Google Scholar]
- Raja, V.; Qadir, S.U.; Alyemeni, M.N.; Ahmad, P. Impact of drought and heat stress individually and in combination on physio-biochemical parameters, antioxidant responses, and gene expression in Solanum lycopersicum. 3 Biotech 2020, 10, 208. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef] [PubMed]
- Arbona, V.; Hossain, Z.; López-Climent, M.F.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Antioxidant enzymatic activity is linked to waterlogging stress tolerance in citrus. Physiol. Plant. 2008, 132, 452–466. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Env. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Wang, B.; Chen, J.; Chen, L.; Wang, X.; Wang, R.; Ma, L.; Peng, S.; Luo, J.; Chen, Y. Combined drought and heat stress in Camellia oleifera cultivars: Leaf characteristics, soluble sugar and protein contents, and Rubisco gene expression. Trees 2015, 29, 1483–1492. [Google Scholar] [CrossRef]
- Ahmed, C.; Rouina, B.; Boukhris, M. Changes in water relations, photosynthetic activity and proline accumulation in one-year-old olive trees (Olea europaea L. cv. ChemLali) in response to NaCl salinity. Acta Physiol. Plant. 2008, 30, 553–560. [Google Scholar] [CrossRef]
- MoLinari, H.; Marur, C.; Daros, E.; de Campos, M.; de Carvalho, J.; Filho, J.; Pereira, L.; Vieira, L. Evaluation of the stress-inducible production of proline in transgenic sugarcane (Saccharum spp.): Osmotic adjustment, chlorophyll fluorescence and oxidative stress. Physiol. Plant. 2007, 130, 218–229. [Google Scholar] [CrossRef]
- Borgo, L.; Marur, C.J.; Vieira, L.G. Effects of high proline accumulation on chloroplast and mitochondrial ultrastructure and on osmotic adjustment in tobacco plants. Acta Sci. Agron. 2015, 37, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Kong, L.; Wu, Z.; Rosenqvist, E.; Wanga, Y.; Zhaoa, L.; Zhaoa, T.; Ottosen, C. Physiological response of tomatoes at drought, heat and their combination followed by recovery. Physiol. Plant. 2019, 165, 144–154. [Google Scholar] [CrossRef]
- Adams III, W.; Demmig-Adams, B. Carotenoid composition and down regulation of photosystem II in three conifer species during the winter. Physiol. Plant. 1994, 92, 451–458. [Google Scholar] [CrossRef]
- Grover, A.; Mittal, D.; Negi, M.; Lavania, D. Generating high temperature tolerant transgenic plants: Achievements and challenges. Plant Sci. 2013, 205–206, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Irigoyen, J.; Einerich, D.; Sanchez-Diaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Wollenweber, B.; Jiang, D. Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities in wheat seedlings. Plant Physiol. Bioch. 2014, 74, 185–192. [Google Scholar] [CrossRef]
- Harsh, A.; Sharma, Y.; Joshi, U.; Rampuria, S.; Singh, G.; Kumar, S.; Sharma, R. Effect of short-term heat stress on total sugars, proline and some antioxidant enzymes in moth bean (Vigna aconitifolia). Ann. Agric. Sci. 2016, 61, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Vinocur, B.; Altman, A. Recent Advances in Engineering Plant Tolerance to Abiotic Stress: Achievements and Limitations. Curr. Opin. Biotechnol. 2005, 16, 123–132. [Google Scholar] [CrossRef]
- Correia, B.; Hancock, R.D.; Amaral, J.; Gomez-Cadenas, A.; Valledor, L.; Pinto, G. Combined drought and heat activates protective responses in Eucalyptus globulus that are not activated when subjected to drought or heat stress alone. Front. Plant Sci. 2018, 9, 819. [Google Scholar] [CrossRef] [Green Version]
- Canales, J.; Bautista, R.; Label, P.; Gómez-Maldonado, J.; Lesur, I.; Fernández-Pozo, N.; Rueda-López, M.; Guerrero-Fernández, D.; Castro-Rodríguez, V.; Hicham, B.; et al. Gene expression profiling in the stem of young maritime pine trees: Detection of ammonium stress-responsive genes in the apex. Trees 2012, 26, 609–619. [Google Scholar] [CrossRef]
- Vega-Bartol, J.J.; Santos, R.R.; Simoes, M.; Miguel, C.M. Normalizing gene expression by quantitative PCR during somatic embryogenesis in two representative conifer species: Pinus pinaster and Picea abies. Plant Cell Rep. 2013, 32, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Svačinová, J.; Novák, O.; Plačková, L.; Lenobel, R.; Holík, J.; Strnad, M.; Doležal, K. A new approach for cytokinin isolation from Arabidopsis tissues using miniaturized purification: Pipette tip solid-phase extraction. Plant Methods 2012, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Šimura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant hormonomics: Multiple phytohormone profiling by targeted metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–208. [Google Scholar] [CrossRef]
- Nebauer, S.; Renau-Morata, B.; Guardiola, J.; MoLina, R. Photosynthesis down-regulation precedes carbohydrate accumulation under sink limitation in Citrus. Tree Physiol. 2011, 31, 169–177. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1997, 148, 350–382. [Google Scholar] [CrossRef]
- Rodríguez, O.; Vilasó, J.E.; Aguilera, I.; Pérez, R.M.; Ábalos, A. Validación por verificación del método colorimétrico de la antrona para la cuantificación de ramnolípidos. Revista Cubana de Química 2013, 25, 287–294. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Provenance | Family | % of Explants with Embryogenic Masses (EMs) | ||||
|---|---|---|---|---|---|---|
| Megagametophyte Priming (°C) | ||||||
| 23 °C (NP) | 30 °C | 37 °C | 50 °C | Mean z | ||
| Galicia Mean y | 1007 | 2.5 ± 2.5 | 3.0 ± 1.5 | NT | 2.5 ± 1.5 | 2.0 ± 0.8 b |
| 1046 | 9.2 ± 3.2 | 8.2 ± 3.0 | NT | 7.3 ± 2.4 | 6.2 ± 1.3 a | |
| 1058 | 21.2 ± 3.6 | 10.2 ± 3.3 | NT | 12.0 ± 4.7 | 10.9 ± 2.1 a | |
| 10.9 ± 2.3 | 7.2 ± 1.6 | - | 7.1 ± 1.9 | |||
| Soria-Burgos Mean y | B5 | 48.4 ± 3.9 | NT | 10.1 ± 2.3 | 29.7 ± 6.4 | 22.1 ± 3.8 a |
| B14 | 23.9 ± 4.8 | NT | 0.9 ± 0.9 | 16.8 ± 4.7 | 10.4 ± 2.9 b | |
| B50 | 18.2 ± 6.2 | NT | 0.0 ± 0.0 | 2.4 ± 2.4 | 5.1 ± 3.0 c | |
| 30.9 ± 3.6 a | - | 4.0 ± 1.5 c | 18.7 ± 3.7 b | |||
| Provenance | Priming (°C) | Number of Lines at Maturation | Lines that Produced Mature SE (%) | Number of Recovered Plants |
|---|---|---|---|---|
| Galicia | 23 (NP) | 11 | 72.7 | 46 |
| 30 | 13 | 76.9 | 99 | |
| 50 | 12 | 91.7 | 66 | |
| Soria-Burgos | 23 (NP) | 15 | 86.7 | 64 |
| 37 | 7 | 100.0 | 95 | |
| 50 | 12 | 91.7 | 125 |
| Gene | Gene Annotation | Reference/NCBI Accession |
|---|---|---|
| APX | Ascorbate Peroxidase | AY485994 |
| CCO | Caffeoyl-CoA O-Methyltransferase (CCOMT) | AM502291.1, JN013969.1 |
| HSP | Heat Shock Protein 70 (HSP70) | CT577590 |
| SOD | Cu-Zn-superoxide dismutase precursor | AF434186 |
| WKY | Transcription factor WRKY11 | CT582155 |
| AGO | Argonaute 9 (AGO9) | de Vega-Bartol et al. [40] |
| BSH | Bushy Growth (BSH) | |
| CLF | Curly Leaf (CLF) | |
| DCL | Dicer-Like 1 (DCL1) | |
| HDA | Histone Deacetilase 9 (HDA9) | |
| DNM | DNA (Cytosine-5)-Methyltransferase 1 (DNMT1) | Rodrigues et al. [41] |
| H12 | Histone 1.2 (H1.2) | |
| TUB | α-Tubuline (α-TUB) | de Vega-Bartol et al. [78] |
| HIS | Histone 3 (HIS3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Oliver, M.A.; Haro, J.G.; Pavlović, I.; Novák, O.; Segura, J.; Sales, E.; Arrillaga, I. Priming Maritime Pine Megagametophytes during Somatic Embryogenesis Improved Plant Adaptation to Heat Stress. Plants 2021, 10, 446. https://doi.org/10.3390/plants10030446
Pérez-Oliver MA, Haro JG, Pavlović I, Novák O, Segura J, Sales E, Arrillaga I. Priming Maritime Pine Megagametophytes during Somatic Embryogenesis Improved Plant Adaptation to Heat Stress. Plants. 2021; 10(3):446. https://doi.org/10.3390/plants10030446
Chicago/Turabian StylePérez-Oliver, María Amparo, Juan Gregorio Haro, Iva Pavlović, Ondřej Novák, Juan Segura, Ester Sales, and Isabel Arrillaga. 2021. "Priming Maritime Pine Megagametophytes during Somatic Embryogenesis Improved Plant Adaptation to Heat Stress" Plants 10, no. 3: 446. https://doi.org/10.3390/plants10030446