
Characterization of the Molecular Mechanisms of Resistance against DMI Fungicides in Cercospora beticola Populations from the Czech Republic

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Fungicide Sensitivity Assay
2.3. Preparation of Liquid Cultures
2.4. C. beticola Cyp51 Gene Expression Analysis
2.5. Cloning of the Whole Cyp51 Gene from C. beticola
2.6. Three-Dimensional Structural Validation and Visualization
2.7. Molecular Docking and Simulation Studies
2.8. MD Trajectories Analysis
3. Results
3.1. Fungicide Sensitivity Assays
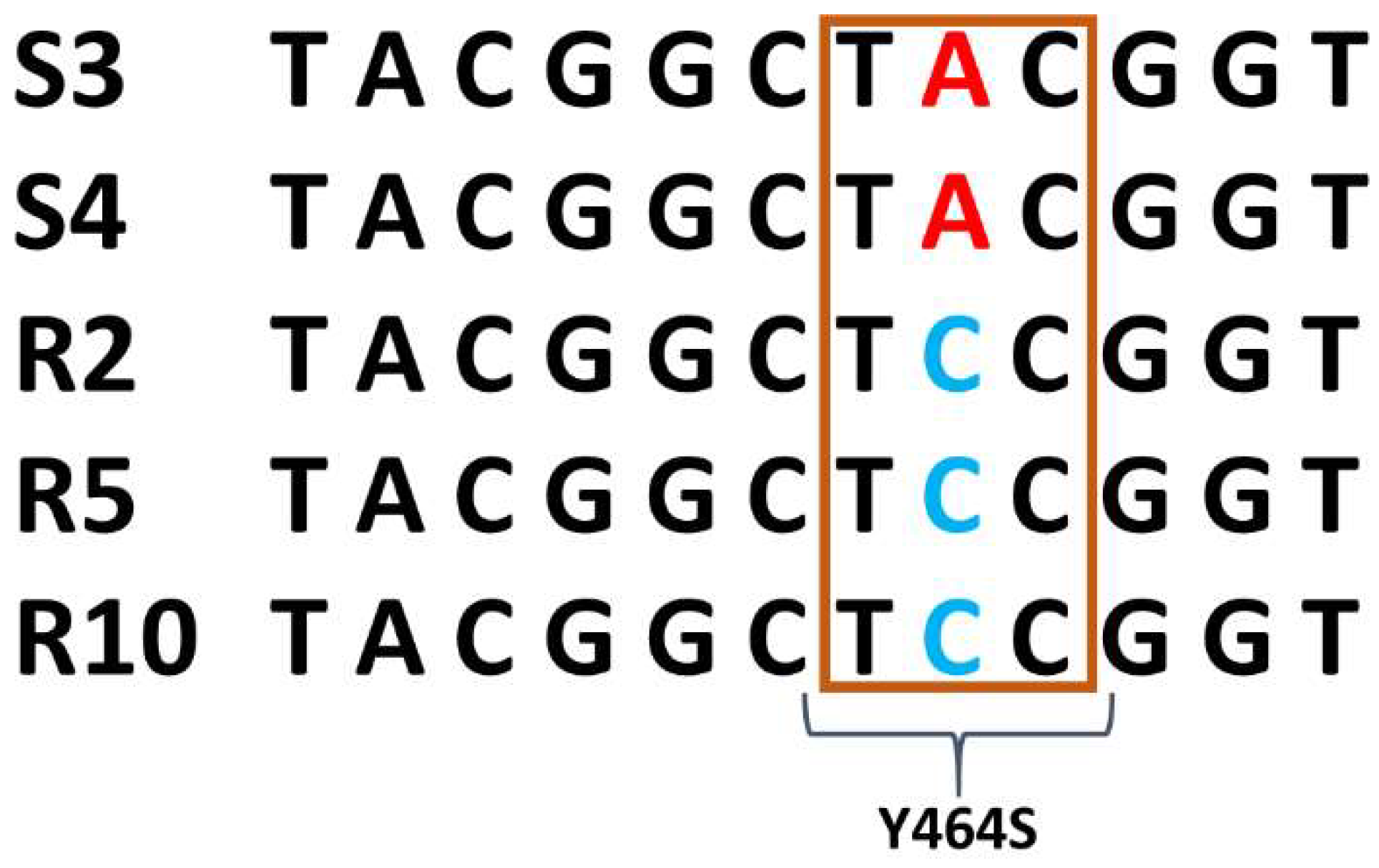
3.2. CbCyp51 Gene Mutations
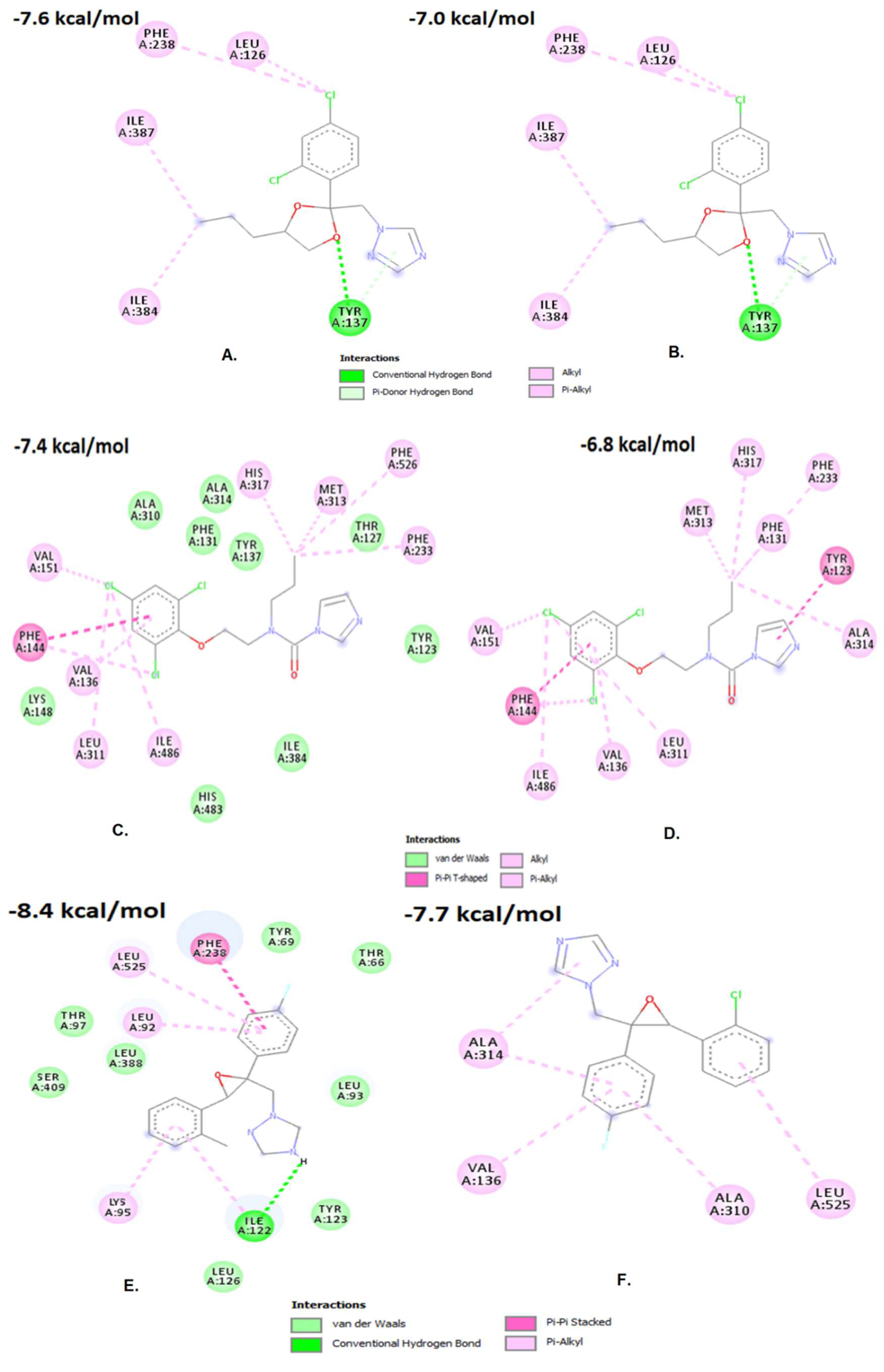
3.3. Impact of the Y464S Mutation on Fungicide Binding
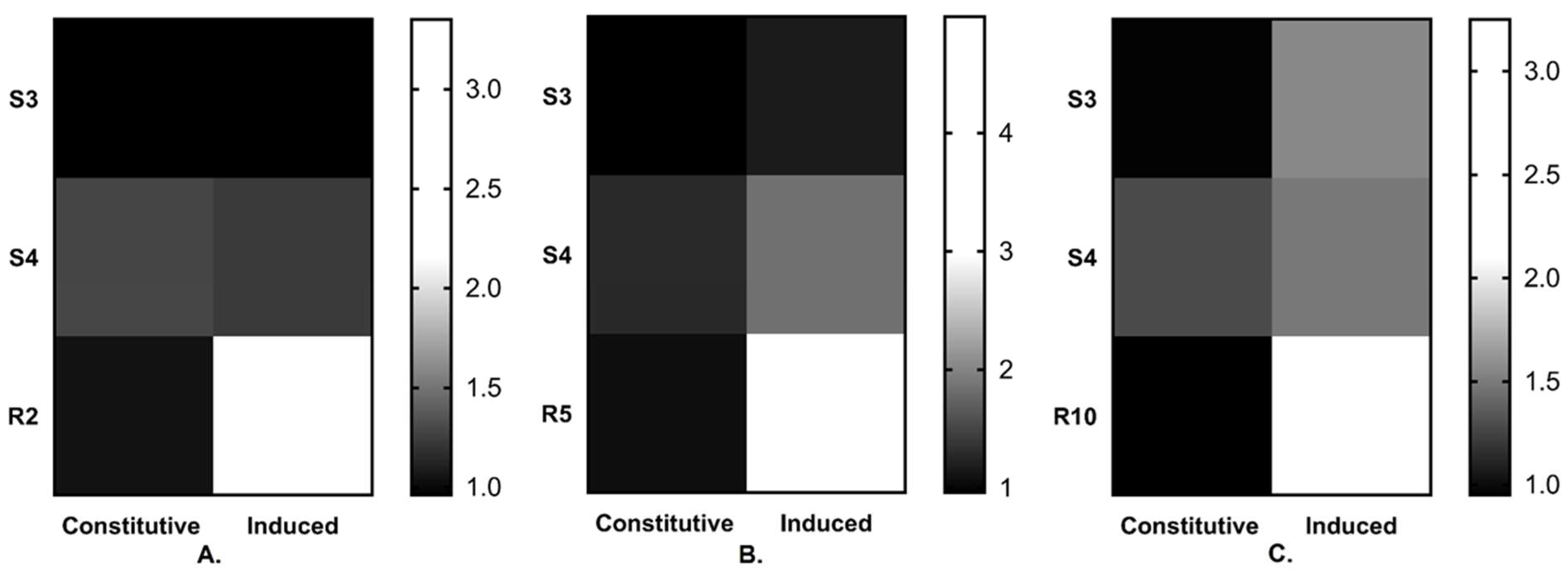
3.4. Cyp51 Gene Expression Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Compliance with Ethics Requirements
References
- Weiland, J.; Koch, G. Sugar beet leaf spot disease (Cercospora beticola Sacc.). Mol. Plant Pathol. 2004, 5, 157–166. [Google Scholar] [CrossRef]
- Rangel, L.I.; Spanner, R.E.; Ebert, M.K.; Pethybridge, S.J.; Stukenbrock, E.H.; de Jonge, R.; Secor, G.A.; Bolton, M.D. Cercospora beticola: The intoxicating lifestyle of the leaf spot pathogen of sugar beet. Mol. Plant Pathol. 2020, 21, 1020–1041. [Google Scholar] [CrossRef] [PubMed]
- Skaracis, G.N.; Biancardi, E. Breeding for Cercospora resistance in sugar beet. Cercospora beticola Sacc Biol. Agron. Influ. Control Meas. Sugar Beet 2000, 2, 177–195. [Google Scholar]
- Lollis, M.; King, S.; Jacobsen, B.; Lollis, M. Influence of glyphosate on Fusarium wilt, Cercospora leaf spot, and Rhizoctonia root and Crown rot diseases of sugar beets. Phytopathology 2009, 99, S76. [Google Scholar]
- Skaracis, G.N.; Pavli, O.I.; Biancardi, E. Cercospora leaf spot disease of sugar beet. Sugar Tech. 2010, 12, 220–228. [Google Scholar] [CrossRef]
- Khan, J.; Del Río, L.E.; Nelson, R.; Khan, M.F.R. Improving the Cercospora leaf spot management model for sugar beet in Minnesota and North Dakota. Plant Dis. 2007, 91, 1105–1108. [Google Scholar] [CrossRef] [PubMed]
- Wolf, P.F.J.; Verreet, J.A. Quaternary concept of Integrated Pest Management (IPM) developed for the control of Cercospora leaf spot in sugar beet. Cercospora Leaf Spot Sugar Beet Relat. Species Am. Phytopathol. Soc. St Paul Minn. USA 2010, 223–233. [Google Scholar]
- Muellender, M.M.; Mahlein, A.-K.; Stammler, G.; Varrelmann, M. Evidence for the association of target-site resistance in Cyp51 with reduced DMI sensitivity in European Cercospora beticola field isolates. Pest Manag. Sci. 2021, 77, 1765–1774. [Google Scholar] [CrossRef]
- Bolton, M.D.; Rivera, V.; Secor, G. Identification of the G143A mutation associated with QoI resistance in Cercospora beticola field isolates from Michigan, United States. Pest Manag. Sci. 2013, 69, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Secor, G.A.; Rivera, V.V.; Khan, M.F.R.; Gudmestad, N.C. Monitoring fungicide sensitivity of Cercospora beticola of sugar beet for disease management decisions. Plant Dis. 2010, 94, 1272–1282. [Google Scholar] [CrossRef] [Green Version]
- Kayamori, M.; Zakharycheva, A.; Saito, H.; Komatsu, K. Resistance to demethylation Inhibitors in Cercospora beticola, a pathogen of sugar beet in Japan, and development of unique Cross-resistance patterns. Eur. J. Plant Pathol. 2021, 160, 39–52. [Google Scholar] [CrossRef]
- Georgopoulos, S.G.; Dovas, C. A Serious outbreak of strains of Cercospora beticola resistant to benzimidazole fungicides in Northern Greece. Plant Dis. Report. 1973, 57, 321–324. [Google Scholar]
- Karaoglanidis, G.S.; Ioannidis, P.M.; Thanassoulopoulos, C.C. Reduced sensitivity of Cercospora beticola isolates to sterol-demethylation-inhibiting fungicides. Plant Pathol. 2000, 49, 567–572. [Google Scholar] [CrossRef]
- Trkulja, N.R.; Milosavljević, A.G.; Mitrović, M.S.; Jović, J.B.; Toševski, I.T.; Khan, M.F.; Secor, G.A. Molecular and experimental evidence of multi-resistance of Cercospora beticola field populations to MBC, DMI and QoI fungicides. Eur. J. Plant Pathol. 2017, 149, 895–910. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Lv, Q.; Yan, L.; Wang, Y.; Jiang, Y. The fungal CYP51s: Their functions, structures, related drug resistance, and inhibitors. Front. Microbiol. 2019, 10, 691. [Google Scholar] [CrossRef] [PubMed]
- Ziogas, B.N.; Malandrakis, A.A. Sterol biosynthesis inhibitors: C-14 demethylation (DMIs). In Fungicide Resistance in Plant Pathogens: Principles and a Guide to Practical Management; Springer: Tokyo, Japan, 2015; pp. 199–216. ISBN 978-4-431-55642-8. [Google Scholar]
- Spanner, R.; Taliadoros, D.; Richards, J.; Rivera-Varas, V.; Neubauer, J.; Natwick, M.; Hamilton, O.; Vaghefi, N.; Pethybridge, S.; Secor, G.A. Genome-wide association and selective sweep studies reveal the complex genetic architecture of DMI fungicide resistance in Cercospora beticola. Genome Biol. Evol. 2021, 13, evab209. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. Chapter two—The evolution of fungicide resistance. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2015; Volume 90, pp. 29–92. [Google Scholar]
- Ma, Z.; Proffer, T.J.; Jacobs, J.L.; Sundin, G.W. Overexpression of the 14α-demethylase target gene (CYP51) mediates fungicide resistance in Blumeriella jaapii. Appl. Environ. Microbiol. 2006, 72, 2581–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, N.J.; Bass, C.; Dixon, A.; Neve, P. The evolutionary origins of pesticide resistance. Biol. Rev. 2019, 94, 135–155. [Google Scholar] [CrossRef]
- Marichal, P.; Bossche, H.V.; Odds, F.C.; Nobels, G.; Warnock, D.W.; Timmerman, V.; Broeckhoven, C.V.; Fay, S.; Mose-Larsen, P. Molecular biological characterization of an azole-resistant Candida glabrata Isolate. Antimicrob. Agents Chemother. 1997, 41, 2229–2237. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Chen, S. Non-target site mechanisms of fungicide resistance in crop pathogens: A Review. Microorganisms 2021, 9, 502. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.D.; Ebert, M.K.; Faino, L.; Rivera-Varas, V.; de Jonge, R.; Van de Peer, Y.; Thomma, B.P.; Secor, G.A. RNA-sequencing of Cercospora beticola DMI-sensitive and resistant isolates after treatment with tetraconazole identifies common and contrasting pathway induction. Fungal Genet. Biol. 2016, 92, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakaune, R.; Hamamoto, H.; Imada, J.; Akutsu, K.; Hibi, T. A novel ABC transporter gene, PMR5, is involved in multidrug resistance in the phytopathogenic fungus Penicillium digitatum. Mol. Genet. Genomics 2002, 267, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Schoonbeek, H.; Del Sorbo, G.; De Waard, M.A. The ABC transporter BcatrB affects the sensitivity of Botrytis cinerea to the phytoalexin resveratrol and the fungicide fenpiclonil. Mol. Plant. Microbe Interact. 2001, 14, 562–571. [Google Scholar] [CrossRef] [Green Version]
- Deising, H.B.; Reimann, S.; Pascholati, S.F. Mechanisms and significance of fungicide resistance. Braz. J. Microbiol. 2008, 39, 286–295. [Google Scholar] [CrossRef] [Green Version]
- Kitchen, J.L.; van den Bosch, F.; Paveley, N.D.; Helps, J.; van den Berg, F. The evolution of fungicide resistance resulting from combinations of foliar-acting systemic seed treatments and foliar-applied fungicides: A modeling analysis. PLoS ONE 2016, 11, e0161887. [Google Scholar] [CrossRef]
- Krug, J.C. Moist chambers for the development of fungi. In Biodiversity of fungi; Academic Press: Burlington, ON, USA, 2004; pp. 589–593. [Google Scholar]
- Bolton, M.D.; Birla, K.; Rivera-Varas, V.; Rudolph, K.D.; Secor, G.A. Characterization of CbCyp51 from field isolates of Cercospora beticola. Phytopathology 2012, 102, 298–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, F.P.; Wilcox, W.F. Sensitivity to azoxystrobin among isolates of Uncinula necator: Baseline distribution and relationship to myclobutanil sensitivity. Plant Dis. 2002, 86, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Palli, S.R. Epigenetic modifications acetylation and deacetylation play important roles in juvenile hormone Action. BMC Genomics 2018, 19, 934. [Google Scholar] [CrossRef]
- Sen, M.K.; Hamouzová, K.; Mikulka, J.; Bharati, R.; Košnarová, P.; Hamouz, P.; Roy, A.; Soukup, J. Enhanced metabolism and target gene overexpression confer resistance against acetolactate synthase-inhibiting herbicides in Bromus sterilis. Pest Manag. Sci. 2021, 77, 2122–2128. [Google Scholar] [CrossRef]
- Amir, M.; Mohammad, T.; Kumar, V.; Alajmi, M.F.; Rehman, M.; Hussain, A.; Alam, P.; Dohare, R.; Islam, A.; Ahmad, F. Structural analysis and conformational dynamics of STN1 gene mutations involved in coat plus syndrome. Front. Mol. Biosci. 2019, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Mazumdar, A.; Haddad, Y.; Milosavljevic, V.; Michalkova, H.; Guran, R.; Bhowmick, S.; Moulick, A. Peptide-carbon quantum dots conjugate, derived from human retinoic acid receptor responder protein 2, against antibiotic-resistant gram positive and gram negative pathogenic bacteria. Nanomaterials 2020, 10, 325. [Google Scholar] [CrossRef] [Green Version]
- Holtschulte, B. Cercospora beticola–Worldwide distribution and incidence. Cercospora beticola 2000, 2, 5–16. [Google Scholar]
- Rossi, V. Effect of host resistance in decreasing infection rate of Cercospora leaf spot epidemics on sugar beet. Phytopathol. Mediterr. 1995, 34, 149–156. [Google Scholar]
- Jacobsen, B.J.; Franc, G.D. Cercospora leaf spot. Compend. Beet Dis. Pests 2009, 2, 7–10. [Google Scholar]
- Rosenzweig, N.; Hanson, L.E.; Mambetova, S.; Jiang, Q.W.; Guza, C.; Stewart, J.; Somohano, P. Fungicide sensitivity monitoring of Alternaria spp. causing leaf spot of sugar beet (Beta vulgaris) in the upper Great Lakes. Plant Dis. 2019, 103, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Nikou, D.; Malandrakis, A.; Konstantakaki, M.; Vontas, J.; Markoglou, A.; Ziogas, B. Molecular characterization and detection of overexpressed C-14 alpha-demethylase-based DMI resistance in Cercospora beticola field isolates. Pestic. Biochem. Physiol. 2009, 95, 18–27. [Google Scholar] [CrossRef]
- Shrestha, S.; Neubauer, J.; Spanner, R.; Natwick, M.; Rios, J.; Metz, N.; Secor, G.A.; Bolton, M.D. Rapid detection of Cercospora beticola in sugar beet and mutations associated with fungicide resistance using LAMP or probe-sased qPCR. Plant Dis. 2020, 104, 1654–1661. [Google Scholar] [CrossRef]
- Robbertse, B.; Van Der Rijst, M.; Van Aarde, I.M.R.; Lennox, C.; Crous, P.W. DMI sensitivity and cross-resistance patterns of Rhynchosporium secalis isolates from South Africa. Crop Prot. 2001, 20, 97–102. [Google Scholar] [CrossRef]
- Huf, A.; Rehfus, A.; Lorenz, K.H.; Bryson, R.; Voegele, R.T.; Stammler, G. Proposal for a new nomenclature for CYP51 haplotypes in Zymoseptoria tritici and analysis of their distribution in Europe. Plant Pathol. 2018, 67, 1706–1712. [Google Scholar] [CrossRef]
- Heick, T.M.; Justesen, A.F.; Jørgensen, L.N. Resistance of wheat pathogen Zymoseptoria tritici to DMI and QoI fungicides in the Nordic-Baltic Region-a status. Eur. J. Plant Pathol. 2017, 149, 669–682. [Google Scholar] [CrossRef]
- Cools, H.J.; Mullins, J.G.; Fraaije, B.A.; Parker, J.E.; Kelly, D.E.; Lucas, J.A.; Kelly, S.L. Impact of recently emerged sterol 14α-demethylase (CYP51) variants of Mycosphaerella graminicola on azole fungicide sensitivity. Appl. Environ. Microbiol. 2011, 77, 3830–3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamamoto, H.; Hasegawa, K.; Nakaune, R.; Lee, Y.J.; Makizumi, Y.; Akutsu, K.; Hibi, T. Tandem repeat of a transcriptional enhancer upstream of the sterol 14α-demethylase Gene (CYP51) in Penicillium digitatum. Appl. Environ. Microbiol. 2000, 66, 3421–3426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnabel, G.; Jones, A.L. The 14α-demethylase (CYP51A1) gene is overexpressed in Venturia inaequalis strains resistant to myclobutanil. Phytopathology 2001, 91, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prelich, G. Gene overexpression: Uses, mechanisms, and interpretation. Genetics 2012, 190, 841–854. [Google Scholar] [CrossRef] [Green Version]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein–ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, L.; Hu, J.; Duan, H.; Lin, D.; Liu, P.; Meng, Q.; Li, B.; Si, N.; Liu, C.; et al. Resistance mechanisms and molecular docking studies of four novel QoI fungicides in Peronophythora litchii. Sci. Rep. 2015, 5, 17466. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Diao, Y.; Wang, W.; Hao, J.; Imran, M.; Duan, H.; Liu, X. Assessing the risk for resistance and elucidating the genetics of Colletotrichum truncatum that is only sensitive to some DMI fungicides. Front. Microbiol. 2017, 8, 1779. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Ishida, K.; Konagai, A.; Maebashi, K.; Asaoka, T. Strong antifungal activity of SS750, a new triazole derivative, is based on its selective binding affinity to cytochrome P450 of fungi. Antimicrob. Agents Chemother. 2002, 46, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Chen, W.; Zhao, W.; Wang, J.; Wang, B.; Li, F.; Wei, M.; Guo, J.; Chen, C.; Zheng, J.; et al. Mutations and overexpression of CYP51 associated with DMI-resistance in Colletotrichum gloeosporioides from chili. Plant Dis. 2019, 104, 668–676. [Google Scholar] [CrossRef]
- Hubbard, R.E.; Haider, M.K. Hydrogen bonds in proteins: Role and Strength. eLS 2010. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9780470015902.a0003011.pub2 (accessed on 22 October 2021).
- Mullins, J.G.L.; Parker, J.E.; Cools, H.J.; Togawa, R.C.; Lucas, J.A.; Fraaije, B.A.; Kelly, D.E.; Kelly, S.L. Molecular modelling of the emergence of azole resistance in Mycosphaerella graminicola. PLoS ONE 2011, 6, e20973. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, Y.; Schnabel, G.; Peng, C.A.; Lagishetty, S.; Smith, K.; Luo, C.; Yuan, H. Inherent resistance to 14α-demethylation inhibitor fungicides in Colletotrichum truncatum is likely linked to CYP51A and/or CYP51B gene variants. Phytopathology 2018, 108, 1263–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5’ to 3’) | Amplicon Length (bp) | Annealing Temperature (°C) |
|---|---|---|---|
| F1 | TCGTCTTCCACTTCGTACCC | 172 | 58 |
| R1 | CCGTTCAGGATGAAGTCGTT | ||
| F2 | ACGGAGTTACCCACGTTGTC | 174 | 58 |
| R2 | TCTCCTTGATGTCACGAACG | ||
| F3 | TCGTCTTCCACTTCGTACCC | 1453 | 60 |
| R3 | CTCTCCCACTTCACAACAGC | ||
| F4 | GTGTTTGGCAAGGACGTCG | 1229 | 61 |
| R4 | CTCTCCCACTTCACAACAGC |
| Strain | Propiconazole | Prochloraz | Epoxiconazole |
|---|---|---|---|
| S3 | 0.17 (±0.02) | 0.19 (±0.02) | 0.05 (±0.01) |
| S4 | 0.05 (±0.01) | 0.32 (±0.22) | 0.45 (±0.32) |
| R2 | 14.27 (±2.94) | 2.29 (±0.23) | 2.19 (±0.17) |
| R5 | 5.43 (±0.54) | 44.01 (±5.85) | 2.78 (±0.13) |
| R10 | 3.11 (±0.14) | 4.36 (±0.42) | 39.53 (±6.23) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, R.; Mazakova, J.; Ali, A.; Sur, V.P.; Sen, M.K.; Bolton, M.D.; Manasova, M.; Rysanek, P.; Zouhar, M. Characterization of the Molecular Mechanisms of Resistance against DMI Fungicides in Cercospora beticola Populations from the Czech Republic. J. Fungi 2021, 7, 1062. https://doi.org/10.3390/jof7121062
Kumar R, Mazakova J, Ali A, Sur VP, Sen MK, Bolton MD, Manasova M, Rysanek P, Zouhar M. Characterization of the Molecular Mechanisms of Resistance against DMI Fungicides in Cercospora beticola Populations from the Czech Republic. Journal of Fungi. 2021; 7(12):1062. https://doi.org/10.3390/jof7121062
Chicago/Turabian StyleKumar, Ram, Jana Mazakova, Asad Ali, Vishma Pratap Sur, Madhab Kumar Sen, Melvin D. Bolton, Marie Manasova, Pavel Rysanek, and Miloslav Zouhar. 2021. "Characterization of the Molecular Mechanisms of Resistance against DMI Fungicides in Cercospora beticola Populations from the Czech Republic" Journal of Fungi 7, no. 12: 1062. https://doi.org/10.3390/jof7121062