Could Global Intensification of Nitrogen Fertilisation Increase Immunogenic Proteins and Favour the Spread of Coeliac Pathology?

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods

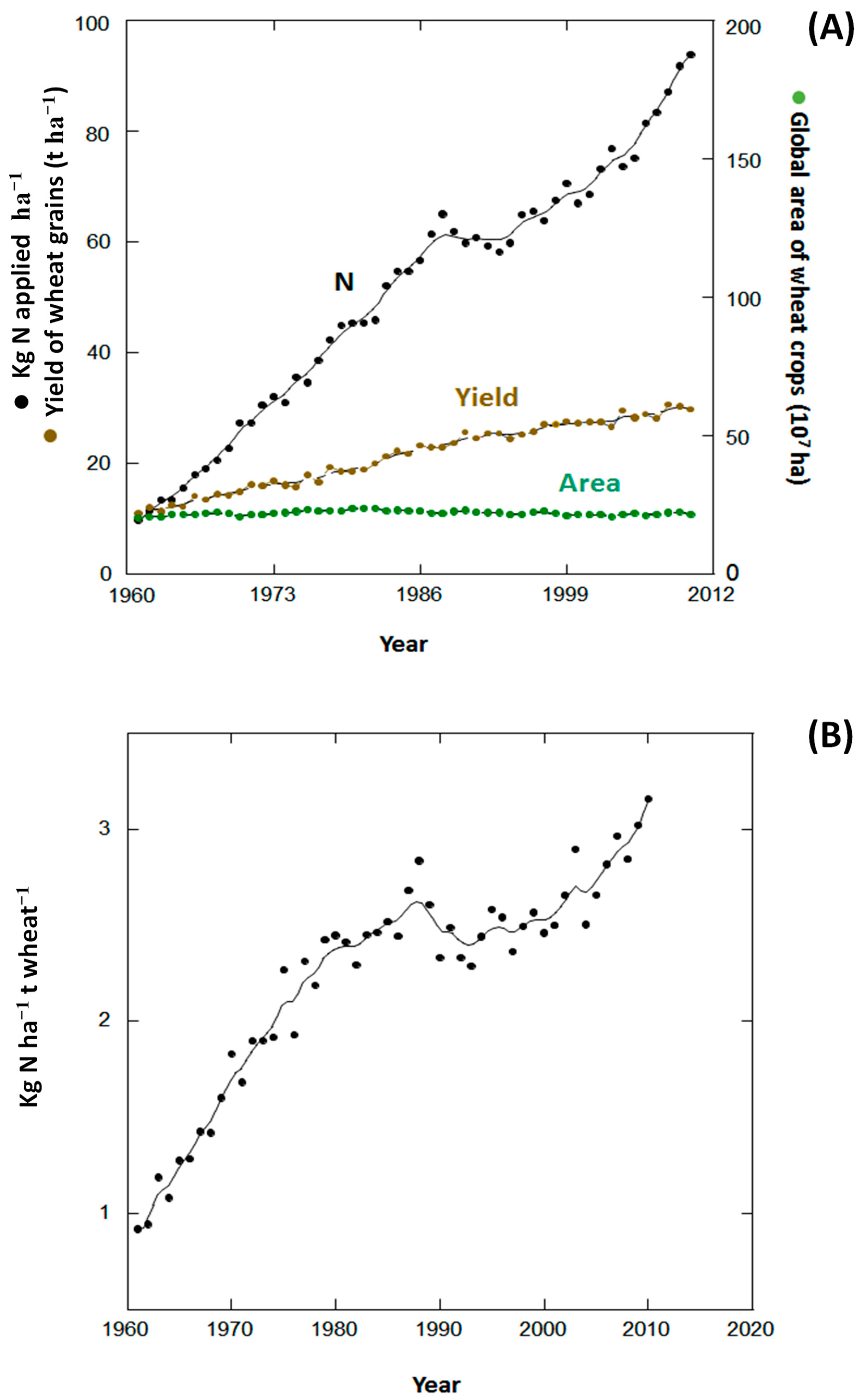{kind=link}
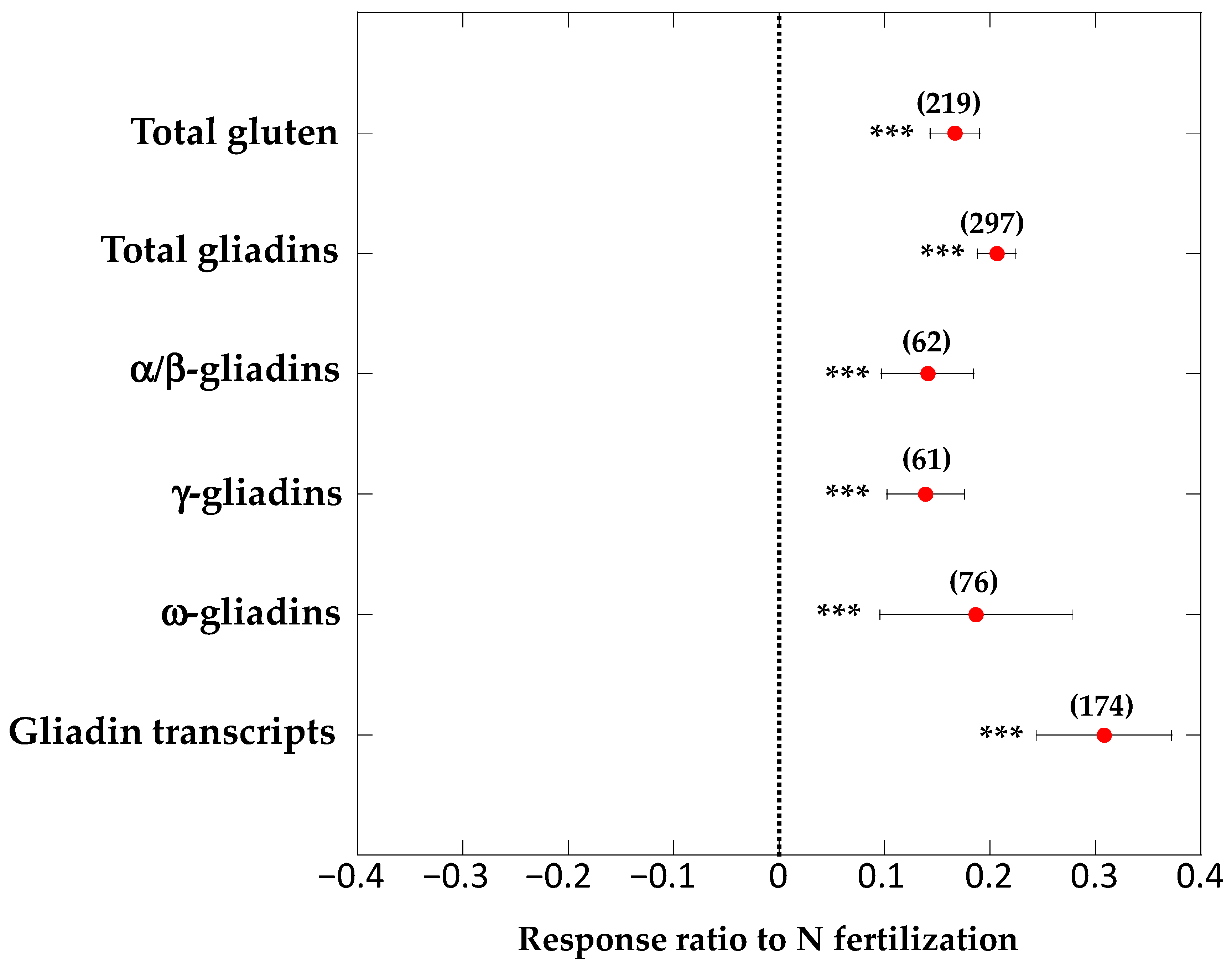{kind=link}
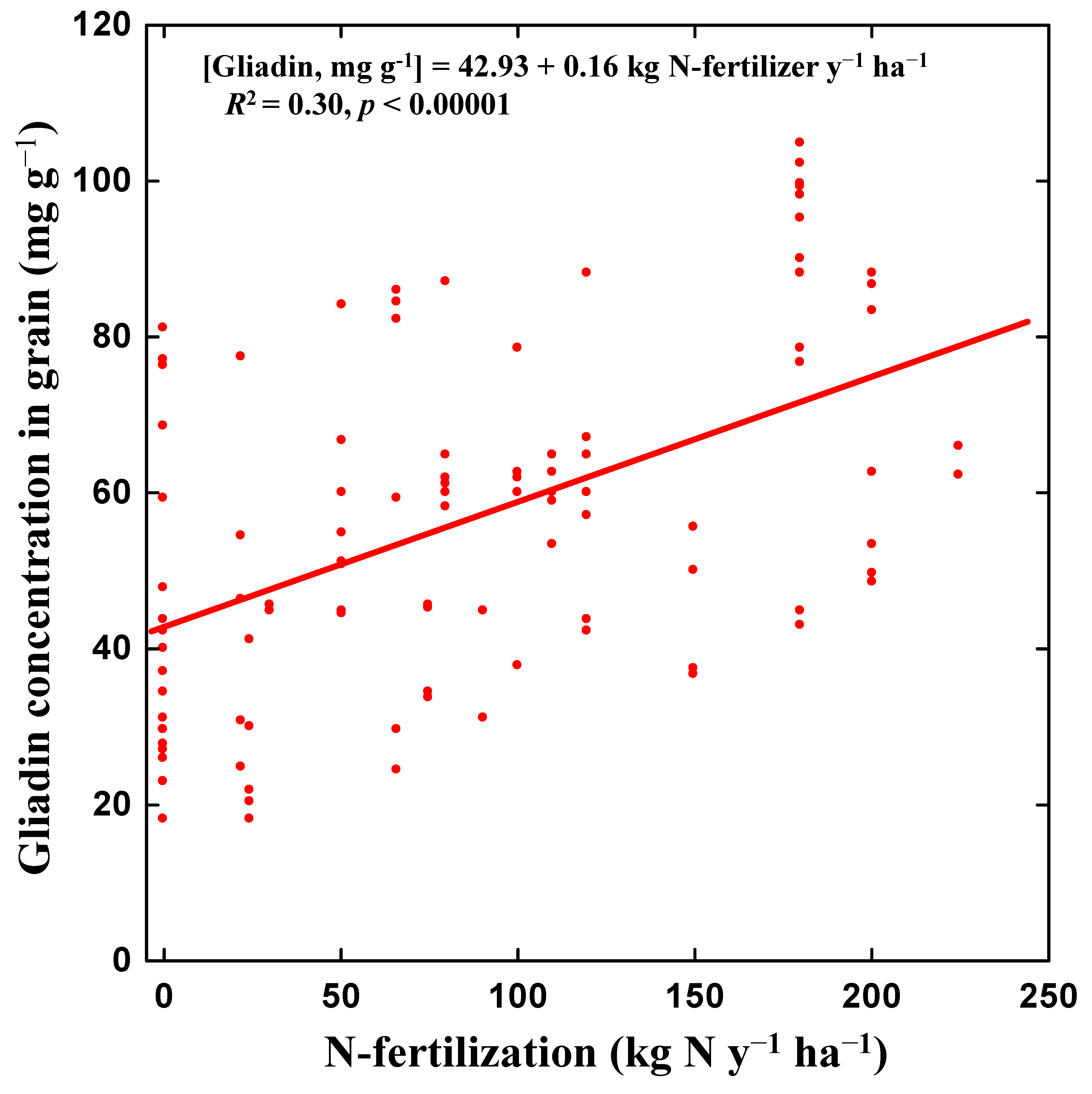{kind=link}
{kind=link}
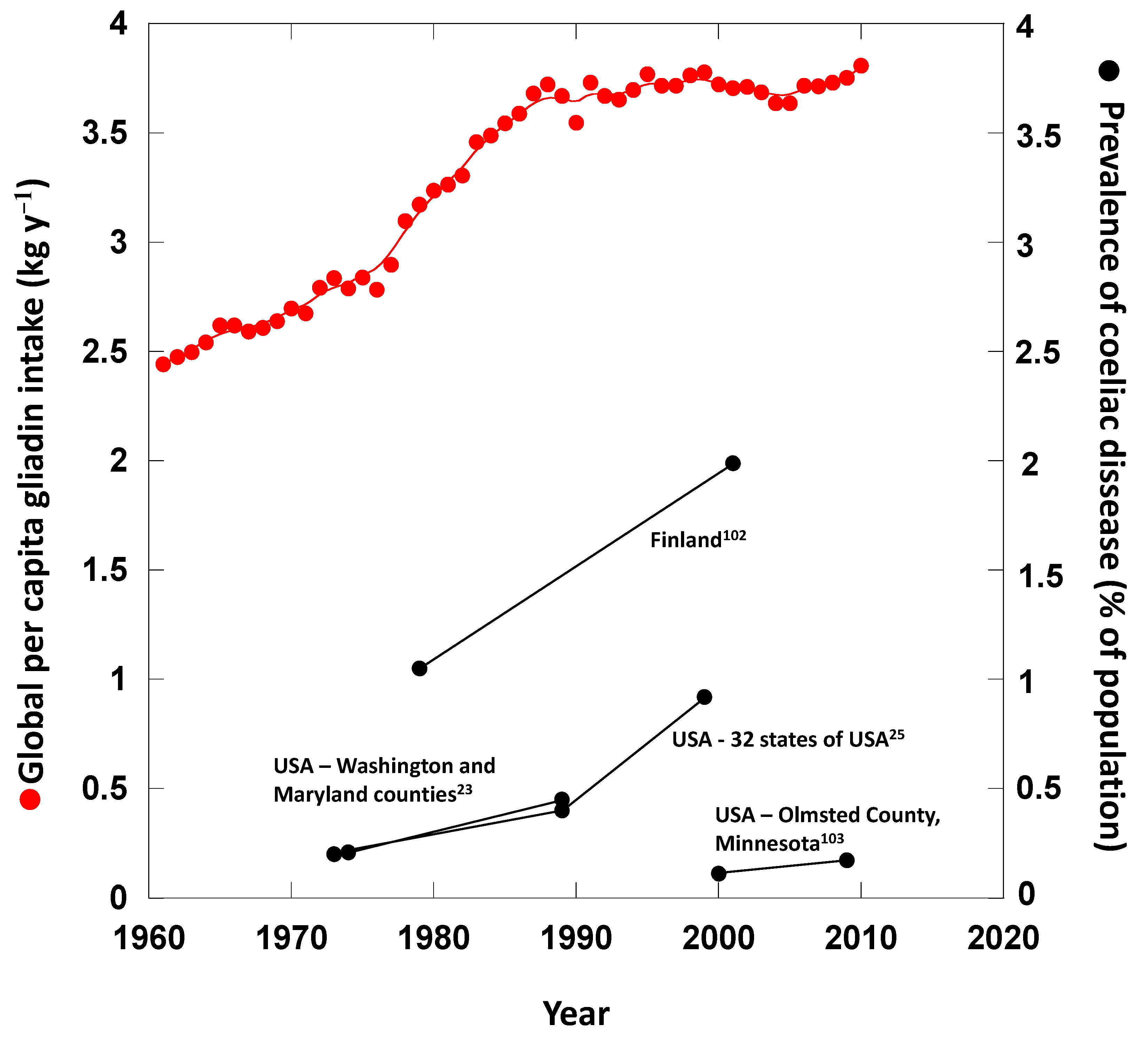{kind=link}
{kind=link}
| Bibliographic Source | Experimental Traits | Result | |||
|---|---|---|---|---|---|
| Reference | Site | Type of Experiment | Tested N-Fertilisation Rates (Kg N ha−1 y−1 if not Specified) | Genotypes | Changes in Concentrations in Grain and/or Flour in Response to Increasing N-Fertilisation |
| [45] | Experimental Farm Shiraz University (Iran) | Field | 0, 120, 240, 360 | Shiraz variety of winter wheat | Increases in gluten concentrations in grain |
| [46] | Bezek experimental station (Poland) | Field | 0, 50, 80 | Triticum aesticum ssp. spelta | Increases in total protein and gluten concentrations in grain |
| [47] | Two field areas of Tunis | Field | 0, 67 | Chili, Biskri, Mahmoudi, INRAT69, Karim, Razzak, Omrabiaa and Khiar varieties | Increases in gluten concentrations in grain |
| [48] | Experimental Station J. Hirschhorn (Argentina) | Field | 0, 70, 140 | Increases in gluten concentrations in grain | |
| [49] | Upland crop experimental farm of National Institute of crop Science (Korea) | Field | 25, 50, 75 | Five Korean wheat varieties | Increases in gluten concentrations in flour. Increases in α + β-gliadin and decreases in ω and γ-gliadin concentrations |
| [50] | Replicates in five field sites in U.K. | Field | 100, 200, 350 | Five breadmaking wheat varieties (Cordial, Hereward, Malacca, Marksman and Xi19) | Increases in total gliadin concentrations in grain |
| [50] | Canada | Field | 0, 100 | Neepawa variety | Increases in total gliadin concentrations in grain |
| [51] | Canada | Field | 0, 50, 100, 150, 200, 250, 300, 350, 400 | Neepawa variety | Increases in total protein and total gliadin concentrations in grain |
| [30] | Eight different site sources | Field | 0, 105, 165, 225 | Triticum aesticum ssp. spelta | Increases of total epipodes expression of a-gliadin in grain |
| [52] | Plant Breeding Station of Sladkovicovo-Novy (Slovakia) | Field | 120, 140 | Winter wheat | Increases in total protein and gluten concentrations in grain |
| [53] | Experimental Station J. Hirschhorn (Argentina) | Field | 0, 70, 140 | Increases in total gluten concentrations in flour | |
| [54] | Alava (Spain) | Field | 0, 100, 140, 180 | Soissons variety | Increases in total gliadin concentrations in grain |
| [55] | Spain | Pot experiment | 37, 48 mg ammonium or nitrate per pot | Cezanne variety | Increases in total protein and gliadins concentrations in flour |
| [56] | Experimental field station Teramo University (Italy) | Field | 50, 100, 150, 250 | Triticum turgidum L. subsp. durum | Increases in total protein, gluten and gliadins concentrations in flour |
| [57] | Spain | Greenhouse | 0, 22.2, 66.7, 200 | Bobwhite variety | Increases in total, α, ω and γ-gliadin and total protein concentrations in flour |
| [58] | National Center of Irrigation Technology station (Spain) | Field | 0, 120 | Winter wheat | Increases in total gliadins concentrations in flour |
| [59] | Field (Sweden) | Field | 0, 70, 140 | Sport, Dacke, Dragon and Thasos varieties | Increases in total proteins and gliadins concentrations in flour |
| [60] | UK | Field | 0, 40, 80, 120, 160, 200, 240 | Option and Riband varieties | Increases in total proteins and gliadins concentrations in grain |
| [61] | Malice (Poland) | Field | 0, 40, 80, 120 | Tybalt variety | Increases in gluten concentrations in grain |
| [62] | Agricultural experimental Staion of University of Technology and Life Sciences of Minikowo (Poland) | Field | 80, 120 | Spring wheat | Increases in gluten concentrations in grain |
| [6] | Peterlauki research and Study Farm (Latvia) | Field | 0, 60, 90, 120, 150, 180, 210, 240 | Skagen variety | Increases in gluten concentrations in grain |
| [63] | Henan Agricultural University Experimental Satation (China) | Field | 0, 90, 180, 270, 360, 450 | Yumai and Lanko Aizao varieties | Increases in total gliadins concentrations in flour |
| [64] | Swadzim Experimental Station (Poland) | Field | 0, 50, 100, 150 | Durabon, Durabonus, Duraprimus and Rusticano varieties | Increases in gluten concentrations in flour |
| [65] | Lincoln Research Farm (New Zealand) | Field | 0, 50, 100 | Batten, Kotare, Oroua, Rongotea, Ruapuna and Tui varieties | Increases in total gliadins concentrations in flour |
| [66] | Mira (Italy) | Field | 70, 120, 130, 160, 180, 200, 240 | Biensur variety | Increases in gluten concentrations in flour |
| [67] | Two different sites (Austria) | Field | 0, 180 | Three varieties: Capo, Renan and Lindos | Increases in total, α, ω and γ gliadin concentrations in flour |
| [68] | Experimental Farm of Helsinki University (Finland) | Field | 0, 110 | Scandinavian, Kadett, Ruso and Reno wheat varieties | Increases in total proteins concentrations but not changes in gliadin concentrations in flour |
| [69] | Hungary | Field | 30–300 | Winter wheat | Increases in gluten concentrations in flour |
| [70] | Chile | Field | 0, 220, 250 | Increases in gluten concentrations in flour | |
| [71] | France | Field | 40, 60 | Seedling from INRA | Increases in total gliadins concentrations in flour |
| [72] | Minokowo (Poland) | Field | 0, 60, 90, 120 | Zebra variety | Increases in gluten concentrations in flour |
| [73] | Field experimental Station of Mediterranean Agronomic Institute of Bari (Italy) | Field | 30, 40, 50, 70 | Triticum turgidum subsp. durum | Increases in total gluten concentrations in grain and flour |
| [74] | Brazil | Field | 0, 50, 100, 150 | Quartzo variety | Increases in gluten concentrations in grain |
| [75] | Experimental farm of INRA, Grignon, France | Field | 40, 60, 120 | Soissons variety | Increases in total gliadin concentrations in grain |
| [76] | Research field sation of Faculty of Agriculture (Croatia) | Field | 0–194 | Marija and Soissons varieties | Increases in gluten concentrations in grain |
| [77] | Rothamsted Research station (UK) | Field | 100, 200, 350 | Cordiale, Hereward, Istabraq, Malacca, Marksman and Xi 19 varieties | Increases in γ-gliadin gene expression |
| [78] | Rothamsted Research station (UK) | Field | 100, 200, 350 | Cordiale, Hereward, Istabraq, Malacca, Marksman and Xi 19 varieties | Increases in ω-gliadin gene expression |
| [17] | Germany | Field | 0, 40, 120, 180, 200 | Dozent, Monopol, Rektor, Apollo, Ares, Astron, Basalt, Bussard, Herzog, Ignaz, Kanzler, Monopol, Obelisk, Sperber varieties | Increases of α/β -gliadin, ω- and γ-gliadins, total gliadins and gluten concentrations in flour |
| [79] | Johann Heinrich von Thunen-Institute, Federal Research Institute for Rural Areas, Forestry and Fisheries, in Braunschweig, Germany | Field | 84, 168 | Batis variety | Increases of α/β -gliadin, ω- and γ-gliadins, total gliadins and gluten concentrations in flour |
| [80] | Research Station of Warmia and Mazury University (Poland) | Field | 80, 120 | Spring triticale cv. Andrus | Increases of α/β -gliadin, no clear effects on ω- and γ-gliadins in grain |
| [81] | Research Station of Warmia and Mazury University (Poland) | Field | 80, 120 | Spring triticale cv. Andrus | Increases of total gliadins concentrations in in grain |
| [82] | Uhrusk Experimental Station belonging to the University of Life Sciences in Lublin (Poland) | Field | 90, 150 | Opatka variety | Increases in gluten concentrations in grain |
| [83] | Fields research stations of Idaho and Monatana state Universities (USA) | Field | 168, 224, 280 | Spring wheat | Increases in gluten concentrations in flour |
| [84] | Futterkamp and Sonke-Nissen-Koog Northern Germany | Field | 220, 260 | Tobak and Asano varieties | Increases in total gliadin and gluten concentrations in flour |
| [85] | Grains Research Centre Kragujevac (Serbia) | Field | 60, 90, 120 | Increases in gluten concentrations in grain | |
| [86] | China Agricultural University Research Center field station, Hebei province, China | Field | 180, 240 | Zhongmai variety | Increases in gluten concentrations in grain |
| [87] | Chongzhou and Renshou experimental stations of Sichuan Agricultural University, China | Field | 0, 75, 150, 225 | Shumai 969, Shumai 482, Chuannong 16 and Mianmai 51 varieties | Increases of total, α/β-gliadin and ω-gliadins and gluten concentrations, no clear effects on ω-gliadins in flour |
| [88] | Germany | Pot experiment | 0.25, 1.0 and 2.5 g N/pot | Privileg variety | Increases of total gliadin concentrations |
3. Increasing Gluten and Gliadin Contents with N-Fertilisation
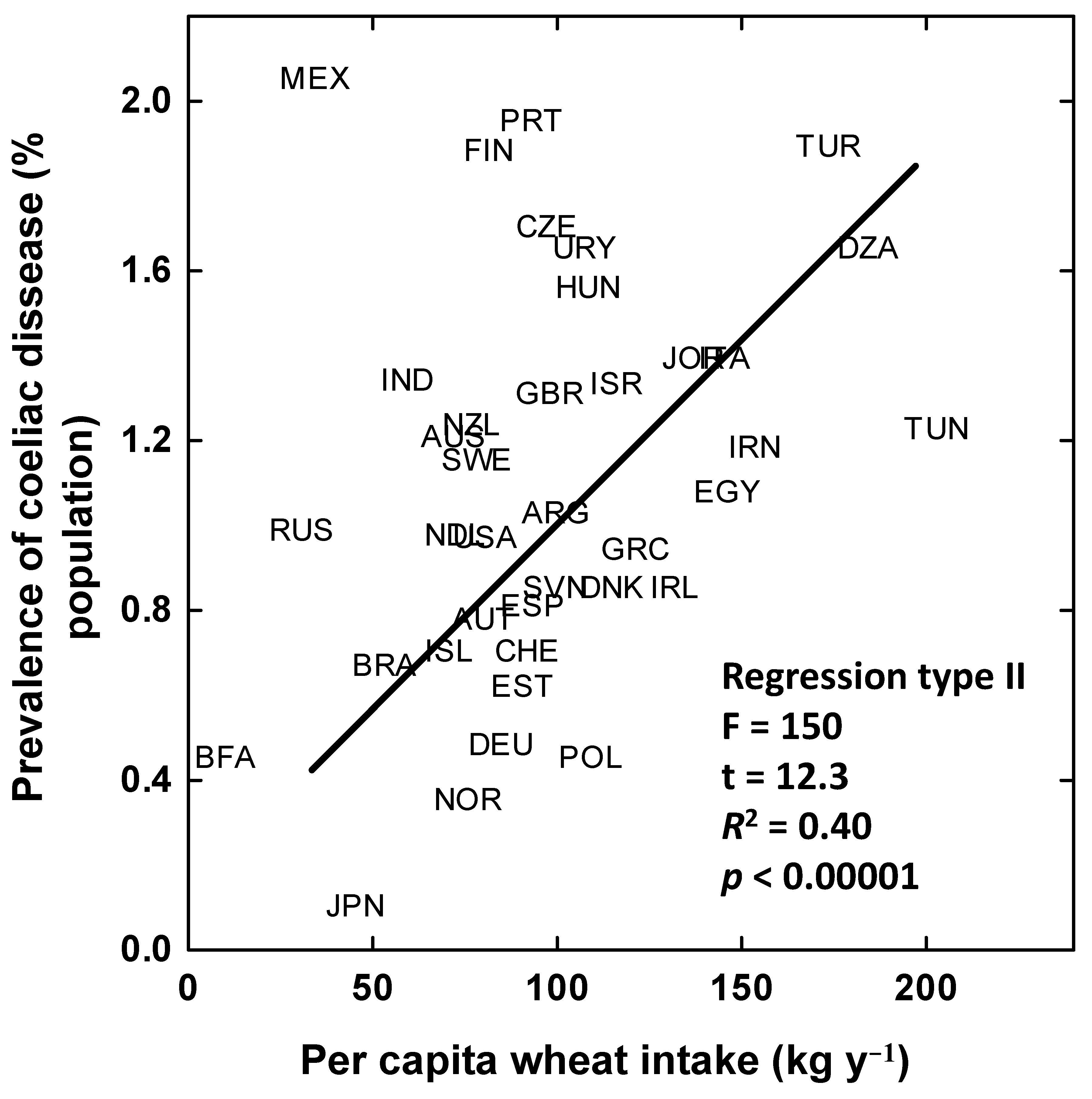
4. Increased Prevalence of Coeliac Disease
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lu, C.; Tian, H. Global nitrogen and phosphorus fertilizer use for agriculture production in the past half century: Shifted hot spots and nutrient imbalance. Earth Syst. Sci. Data 2017, 9, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Heffer, P.; Prud’homme, M. Global Nitrogen Fertilizer Demand and Supply: Trend, Current Level and Outlook. In Proceedings of the International Fertilizer Association (IFA), 7th International Nitrogen Initiative Conference, Melbourne, Australia, 4–8 December 2016; pp. 1–11. [Google Scholar]
- FAOSTAT. 2019. Available online: http://www.fao.org/faostat/en/#data (accessed on 11 July 2019).
- Food and Agriculture Organization of the United Nations (FAO). World Fertilizer Trends and Outlook to 2022; FAO: Rome, Italy, 2019. [Google Scholar]
- Xu, A.; Li, L.; Xie, J.; Wang, X.; Coulter, J.A.; Liu, C.; Wang, L. Effect of long-term nitrogen addition on wheat yield, nitrogen use efficiency, and esidual soil nitrate in a semiarid area of the loess Plateau of China. Sustainability 2019, 12, 1735. [Google Scholar] [CrossRef] [Green Version]
- Litke, L.; Gaile, Z.; Ruza, A. Effect of nitrogen fertilisation on winter wheat yield and yield quality. Agron. Res. 2018, 16, 500–509. [Google Scholar]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on Wheat Yield and Quality with Reduced Nitrogen Supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ierna, A.; Lombardo, G.M.; Mauromicales, G. Yield, nitrogen use efficiency and grain Quality in durum wheat as affected by nitrogen fertilisation under a Mediterranean Environment. Exp. Agric. 2015, 1, 314. [Google Scholar]
- Curtis, B.C. Wheat in the World. 2019. Available online: http://www.fao.org/3/y4011e/y4011e04.htm#TopOfPage (accessed on 13 September 2019).
- Kasarda, D.D. Can an Increase in Celiac Disease Be Attributed to an Increase in the Gluten Content of Wheat as a Consequence of Wheat Breeding? J. Agric. Food Chem. 2013, 61, 1155–1159. [Google Scholar] [CrossRef] [PubMed]
- USDA, United States Department of Agriculture. 2019. Available online: https://www.nass.usda.gov/Publications/Ag_Statistics/2019/index.php (accessed on 13 September 2019).
- Food and Agriculture Organization of the United Nations (FAO). Fertilizer Requirements in 2015 and 2030; FAO: Rome, Italy, 2004. [Google Scholar]
- Ladha, J.K.; Tirol-Padre, A.; Reddy, C.K.; Cassman, K.G.; Verma, S.; Powlson, D.S.; Van Kessel, C.; Richter, D.D.B.; Chakraborty, D.; Pathak, H. Global nitrogen budgets in cereals: A 50-year assessment for maize, rice and wheat production systems. Sci. Rep. 2016, 6, 19355. [Google Scholar] [CrossRef] [Green Version]
- DESA, United Nations, Department of Economic and Social Affairs. Population Dinamics. World Population Prospects. 2019. Available online: https://www.un.org/development/desa/dpad/tag/2019/ (accessed on 13 September 2019).
- FAOSTAT. Food and Agriculture Data. 2019. Available online: www.fao.org/faostat/en/#home (accessed on 13 July 2019).
- Garcia Del Moral, L.F.; Rharrabti, Y.; Martos, V.; Royo, C. Environmentally Induced Changes in Amino Acid Composition in the Grain of Durum Wheat Grown under Different Water and Temperature Regimes in a Mediterranean Environment. J. Agric. Food Chem. 2007, 55, 8144–8151. [Google Scholar] [CrossRef] [PubMed]
- Wieser, H.; Seilmeier, W. The influence of nitrogen fertilisation on quantities and proportions of different protein types in wheat flour. J. Sci. Food Agric. 1998, 76, 49–55. [Google Scholar] [CrossRef]
- Chope, G.A.; Wan, Y.; Penson, S.P.; Bhandari, D.G.; Powers, S.J.; Shewry, P.R.; Hawkesford, M.J. Effects of Genotype, Season, and Nitrogen Nutrition on Gene Expression and Protein Accumulation in Wheat Grain. J. Agric. Food Chem. 2014, 62, 4399–4407. [Google Scholar] [CrossRef]
- Zhen, S.; Deng, X.; Xu, X.; Liu, N.; Zhu, D.; Wang, Z.; Yan, Y. Effect of high-nitrogen fertilizer on gliadin and glutenin subproteomes during kernel development in wheat (Triticum aestivum L.). Crop. J. 2020, 8, 38–52. [Google Scholar] [CrossRef]
- Triboi, A.-M.; Martre, P.; Triboï-Blondel, A. Environmentally-induced changes in protein composition in developing grains of wheat are related to changes in total protein content. J. Exp. Bot. 2003, 54, 1731–1742. [Google Scholar] [CrossRef]
- Martre, P.; Jamieson, P.D.; Semenov, M.A.; Zyskowski, R.F.; Porter, J.R.; Triboi, E. Modelling protein content and composition in relation to crop nitrogen dynamics for wheat. Eur. J. Agron. 2006, 25, 138–154. [Google Scholar] [CrossRef]
- Piston, F.; Gil-Humanes, J.; Barro, F. Integration of promoters, inverted repeat sequences and proteomic data into a model for high silencing efficiency of coeliac disease related gliadins in bread wheat. BMC Plant Biol. 2013, 13, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasano, A.; Berti, I.; Gerarduzzi, T.; Not, T.; Colletti, R.B.; Drago, S.; Elitsur, Y.; Green, P.H.R.; Guandalini, S.; Hill, I.D.; et al. Prevalence of Celiac Disease in At-Risk and Not-At-Risk Groups in the United States. Arch. Intern. Med. 2003, 163, 286–292. [Google Scholar] [CrossRef]
- Mustalahti, K.; Catassi, C.; Reunanen, A.; Fabiani, E.; Heier, M.; McMillan, S.; Murray, L.; Metzger, M.-H.; Gasparin, M.; Bravi, E.; et al. The prevalence of celiac disease in Europe: Results of a centralized, international mass screening project. Ann. Med. 2010, 42, 587–595. [Google Scholar] [CrossRef]
- Catassi, C.; Kryszak, D.; Bhatti, B.; Sturgeon, C.; Helzlsouer, K.; Clipp, S.L.; Gelfond, D.; Puppa, E.; Sferruzza, A.; Fasano, A. Natural history of celiac disease autoimmunity in a USA cohort followed since 1974. Ann. Med. 2010, 42, 530–538. [Google Scholar] [CrossRef]
- Howdle, P. Gliadin, glutenin or both? The search for the Holy Grail in coeliac disease. Eur. J. Gastroenterol. Hepatol. 2006, 18, 703–706. [Google Scholar] [CrossRef]
- Beaudoin, K.; Willourhby, D.S. The role of the gluten-derived peptide gliadina in celíac disease. J. Nutr. Health Food Eng. 2014, 1, 229–232. [Google Scholar]
- Liester, M.G.; Liester, M. Drought’s potential influence on the increasing prevalence of celiac disease. Cogent Med. 2018, 5, 1529848. [Google Scholar] [CrossRef]
- Denery-Papini, S.; Nicolas, Y.; Popineau, Y. Efficiency and Limitations of Immunochemical Assays for the Testing of Gluten-free Foods. J. Cereal Sci. 1999, 30, 121–131. [Google Scholar] [CrossRef]
- Dubois, B.; Bertin, P.; Hautier, L.; Muhovski, Y.; Escarnot, E.; Mingeot, D. Genetic and environmental factors affecting the expression of α-gliadin canonical epitopes involved in celiac disease in a wide collection of spelt (Triticum aestivum ssp. spelta) cultivars and landraces. BMC Plant Biol. 2018, 18, 262. [Google Scholar] [CrossRef] [Green Version]
- Dupont, F.; Vensel, W.H.; Encarnacao, T.; Chan, R.; Kasarda, D.D. Similarities of omega gliadins from Triticum urartu to those encoded on chromosome 1A of hexaploid wheat and evidence for their post-translational processing. Theor. Appl. Genet. 2004, 108, 1299–1308. [Google Scholar] [CrossRef]
- Ensari, A.; Marsh, M.N.; Moriarty, K.J.; Moore, C.M.; Fido, R.J.; Tatham, A.S. Studies in vivo of ω-gliadins in gluten sensitivity (coeliac sprue disease). Clin. Sci. 1998, 95, 419. [Google Scholar] [CrossRef] [Green Version]
- Morita, E.; Matsuo, H.; Mihara, S.; Morimoto, K.; Savage, A.; Tatham, A. Fast ω-gliadin is a major allergen in wheat-dependent exercise-induced anaphylaxis. J. Dermatol. Sci. 2003, 33, 99–104. [Google Scholar] [CrossRef]
- Morrell, K.; Melby, M.K. Celiac Disease: The Evolutionary Paradox. Int. J. Celiac Dis. 2017, 5, 86–94. [Google Scholar]
- Palosuo, K.; Varjonen, E.; Kekki, O.-M.; Klemola, T.; Kalkkinen, N.; Alenius, H.; Reunala, T. Wheat ω-5 gliadin is a major allergen in children with immediate allergy to ingested wheat. J. Allergy Clin. Immunol. 2001, 108, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.; Van Bergen, J.; Loh, K.L.; Kooy-Winkelaar, Y.; Beringer, D.X.; Thompson, A.; Bakker, S.F.; Mulder, C.J.J.; Ladell, K.; McLaren, J.E.; et al. Determinants of Gliadin-Specific T Cell Selection in Celiac Disease. J. Immunol. 2015, 194, 6112–6122. [Google Scholar] [CrossRef] [Green Version]
- Salentijn, E.M.; Mitea, D.C.; Goryunova, S.V.; Van Der Meer, I.M.; Padioleau, I.; Gilissen, L.J.W.J.; Koning, F.; Smulders, M.J.M. Celiac disease T-cell epitopes from gamma-gliadins: Immunoreactivity depends on the genome of origin, transcript frequency, and flanking protein variation. BMC Genom. 2012, 13, 277. [Google Scholar] [CrossRef] [Green Version]
- Green, P.H.; Cellier, C. Celiac Disease. N. Engl. J. Med. 2007, 357, 1731–1743. [Google Scholar] [CrossRef]
- Samsel, A.; Seneff, S. Glyphosate, pathways to modern diseases II: Celiac sprue and gluten intolerance. Interdiscip. Toxicol. 2013, 6, 159–184. [Google Scholar] [CrossRef] [Green Version]
- Vader, W.; Kooy, Y.; Van Veelen, P.A.; De Ru, A.; Harris, D.; Benckhuijsen, W.; Peña, S.; Mearin, L.; Drijfhout, J.W.; Koning, F. The gluten response in children with celiac disease is directed toward multiple gliadin and glutenin peptides. Gastroenterology 2002, 122, 1729–1737. [Google Scholar] [CrossRef] [Green Version]
- Van De Wal, Y.; Kooy, Y.M.C.; Van Veelen, P.; Vader, W.; August, S.A.; Drijfhout, J.W.; Peña, S.A.; Koning, F. Glutenin is involved in the gluten-driven mucosal T cell response. Eur. J. Immunol. 1999, 29, 3133–3139. [Google Scholar] [CrossRef]
- Hedges, L.V.; Gurevitch, J.; Curtis, P. The Meta-Analysis of Response Ratios in Experimental Ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Viechtbauer, W.R. The Metafor Package: A Meta-Analysis Package for R. 2012. Available online: www.metafor-project.org/doku.php.MetaforPackage_2012 (accessed on 15 September 2019).
- Legendre, P. 2018 Packege “Lmodel2”. Available online: https://cran.r-project.org/web/packages/lmodel2/lmodel2.pdf (accessed on 16 September 2019).
- Abedi, T.; Alemzadeh, A.; Kazemeini, S.A. Wheat yield and grain protein response to nitrogen amount and timing. Aust. J. Crop Sci. 2011, 5, 330–336. [Google Scholar]
- Andruszczak, S. Spelt wheat grain yield and nutritional value response to sowing rate and nitrogen fertilisation. J. Anim. Plant Sci. 2018, 28, 1476–1484. [Google Scholar]
- Bouacha, O.D.; Nouaigui, S.; Rezgui, S. Effects of N and K fertilizers on durum wheat quality in different environments. J. Cereal Sci. 2014, 59, 9–14. [Google Scholar] [CrossRef]
- Castro, A.; Constanza, M.; Schierenbeck, M.; Sebastián, G.; Rosa, M. Evaluation of di ff erent fungicides and nitrogen rates on grain yield and bread-making quality in wheat a ff ected by Septoria tritici blotch and yellow spot. J. Cereal Sci. 2018, 83, 49–57. [Google Scholar] [CrossRef]
- Cho, S.-W.; Kang, C.-S.; Kang, T.-G.; Cho, K.-S.; Park, C.S. Influence of different nitrogen application on flour properties, gluten properties by HPLC and end-use quality of Korean wheat. J. Integr. Agric. 2018, 17, 982–993. [Google Scholar] [CrossRef]
- Daniel, C.; Triboi, E. Effects of Temperature and Nitrogen Nutrition on the Grain Composition of Winter Wheat: Effects on Gliadin Content and Composition. J. Cereal Sci. 2000, 32, 45–56. [Google Scholar] [CrossRef]
- Dubetz, S.; Gardiner, E.E.; Flynn, D.; De La Roche, A.I. Effect of nitrogen fertilizer on nitrogen fractions and amino acid composition of spring wheat. Can. J. Plant Sci. 1979, 59, 299–305. [Google Scholar] [CrossRef]
- Ducsay, L.; Ložek, O. Effect of topdressing with nitrogen on the yield and quality of winter wheat grain. Plant Soil Environ. 2011, 50, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Fleitas, M.C.; Schierenbeck, M.; Gerard, G.S.; Dietz, J.I.; Golik, S.I.; Campos, P.E.; Simón, M.R. How leaf rust disease and its control with fungicides affect dough properties, gluten quality and loaf volume under different N rates in wheat. J. Cereal Sci. 2018, 80, 119–127. [Google Scholar] [CrossRef]
- Fuertes-Mendizábal, T.; Aizpurua, A.; González-Moro, M.; Estavillo, J. Improving wheat breadmaking quality by splitting the N fertilizer rate. Eur. J. Agron. 2010, 33, 52–61. [Google Scholar] [CrossRef]
- Fuertes-Mendizábal, T.; González-Torralba, J.; Arregui, L.M.; González-Murua, C.; González-Moro, M.B.; Estavillo, J.M. Ammonium as sole N source improves grain quality in wheat. J. Sci. Food Agric. 2013, 93, 2162–2171. [Google Scholar] [CrossRef]
- Galieni, A.; Stagnari, F.; Visioli, G.; Marmiroli, N.; Speca, S.; Angelozzi, G.; D’Egidio, S.; Pisante, M. Nitrogen fertilisation of durum wheat: A case of study in Mediterranean area during transition to conservation agriculture. Ital. J. Agron. 2016, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- García-Molina, M.D.; Barro, F. Characterization of Changes in Gluten Proteins in Low-Gliadin Transgenic Wheat Lines in Response to Application of Different Nitrogen Regimes. Front. Plant Sci. 2017, 8, 257. [Google Scholar] [CrossRef] [Green Version]
- Guardia, G.; Sanz-cobena, A.; Sanchez-martín, L.; Fuertes-mendizábal, T.; González-murua, C.; Manuel, J.; Chadwick, D.; Vallejo, A. Agriculture, Ecosystems and Environment Urea-based fertilisation strategies to reduce yield-scaled N oxides and enhance bread-making quality in a rainfed Mediterranean wheat crop. Agric. Ecosyst. Environ. 2018, 265, 421–431. [Google Scholar] [CrossRef]
- Johansson, E.; Prieto-Linde, M.L.; Svensson, G. Influence of nitrogen application rate and timing on grain protein composition and gluten strength in Swedish wheat cultivars. J. Plant Nutr. Soil Sci. 2004, 167, 345–350. [Google Scholar] [CrossRef]
- Kindred, D.R.; Verhoeven, T.M.; Weightman, R.M.; Swanston, J.S.; Agu, R.C.; Brosnan, J.M.; Sylvester-Bradley, R. Effects of variety and fertiliser nitrogen on alcohol yield, grain yield, starch and protein content, and protein composition of winter wheat. J. Cereal Sci. 2008, 48, 46–57. [Google Scholar] [CrossRef]
- Klikocka, H.; Cybulska, M.; Barczak, B.; Narolski, B.; Szostak, B.; Kobiałka, A.; Nowak, A.; Wójcik, E. The effect of sulphur and nitrogen fertilization on grain yield and technological quality of spring wheat. Plant Soil Environ. 2016, 62, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Knapowski, T.; Ralcewicz, M.; Barczak, B.; Kozera, W. Effect of Nitrogen and Zinc Fertilizing on Bread-Making Quality of Spring Triticale Cultivated in Noteć Valley. Pol. J. Environ. Sustain. 2009, 18, 227–233. [Google Scholar]
- Ma, D.; Guo, T.; Wang, Z.; Wang, C.; Zhu, Y.; Wang, Y. Influence of nitrogen fertilizer application rate on winter wheat (Triticum aestivum L.) flour quality and Chinese noodle quality. J. Sci. Food Agric. 2009, 89, 1213–1220. [Google Scholar] [CrossRef]
- Makowska, A.; Obuchowski, W.; Sulewska, H.; Koziara, W.; Paschke, H. Agricultural University of Poznań. Acta Sci. Pol. 2008, 7, 29–39. [Google Scholar]
- Martin, R.; Sutton, K.; Moyle, T.; Hay, R.; Gillespie, R. Effect of nitrogen fertiliser on the yield and quality of six cultivars of autumn-sown wheat. N. Z. J. Crop. Hortic. Sci. 1992, 20, 273–282. [Google Scholar] [CrossRef]
- Morari, F.; Zanella, V.; Sartori, L.; Visioli, G.; Berzaghi, P.; Mosca, G. Optimising durum wheat cultivation in North Italy: Understanding the effects of site-specific fertilization on yield and protein content. Precis. Agric. 2018, 19, 257–277. [Google Scholar] [CrossRef]
- Pechanek, U.; Karger, A.; Gröger, S.; Charvat, B.; Schöggl, G.; Lelley, T. Effect of Nitrogen Fertilisation on Quantity of Flour Protein Components, Dough Properties, and Breadmaking Quality of Wheat. Cereal Chem. 1997, 74, 800–805. [Google Scholar] [CrossRef]
- Peltonen, J.; Virtanen, A. Effect of Nitrogen fertilizers differing in release characteristics on the quantity of storage proteins in wheat. Cereal Chem. 1994, 71, 1–5. [Google Scholar]
- Pepo, P.; Sipos, P.; Gyori, Z. Effects of fertilizer application on the baking quality of winter wheat varieties in a long term experiment under continental climatic conditions in Hungary. Cereal Res. Commun. 2005, 33, 825–832. [Google Scholar] [CrossRef]
- Pinilla-Quezada, H.; Herrera-Floody, L.E. Efecto de la fertilización nitrogenada tardía en aspectos de calidad panadera en trigo (Triticum aestivum L.). Idesia (Arica) 2008, 26, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Plessis, A.; Ravel, C.; Bordes, J.; Balfourier, F.; Martre, P. Association study of wheat grain protein composition reveals that gliadin and glutenin composition are trans-regulated by different chromosome regions. J. Exp. Bot. 2013, 64, 3627–3644. [Google Scholar] [CrossRef] [Green Version]
- Ralcewicz, M.; Knapowski, T.; Kozera, W.; Barczak, B. Technological value of spring wheat of zebra cultivar as related to the way of nitrogen and magnesium application. J. Cent. Eur. Agric. 2009, 10, 223–232. [Google Scholar]
- Rizzello, C.G.; Cavoski, I.; Turk, J.; Ercolini, D.; Nionelli, L.; Pontonio, E.; De Angelis, M.; De Filippis, F.; Gobbetti, M.; Di Cagno, R. Organic Cultivation of Triticum turgidum subsp. durum Is Reflected in the Flour-Sourdough Fermentation-Bread Axis. Appl. Environ. Microbiol. 2015, 81, 3192–3204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrighero, M.B.; Caires, E.F.; Lopes, R.B.; Zielinski, A.A.; Granato, D.; Demiate, I.M. Wheat technological quality as affected by nitrogen fertilization under a no-till system. Acta Sci. Technol. 2015, 37, 175. [Google Scholar] [CrossRef]
- Tea, I.; Genter, T.; Naulet, N.; Boyer, V.; Lummerzheim, M.; Kleiber, D. Effect of Foliar Sulfur and Nitrogen Fertilisation on Wheat Storage Protein Composition and Dough Mixing Properties. Cereal Chem. 2004, 81, 759–766. [Google Scholar] [CrossRef]
- Varga, B.; Svečnjak, Z.; Jurković, Z.; Pospíšil, M. Quality responses of winter wheat cultivars to nitrogen and fungicide applications in Croatia. Acta Agron. Hung. 2007, 55, 37–48. [Google Scholar] [CrossRef]
- Wan, Y.; Shewry, P.R.; Bitonti, M.B. A novel family of γ -gliadin genes induces are highly regulated by In Posidonia oceanica cadmium changes in DNA nitrogen supply developing wheat grain methylation and in chromatin patterning. J. Exp. Bot. 2013, 64, 161–168. [Google Scholar] [CrossRef]
- Wan, Y.; Gritsch, C.S.; Hawkesford, M.J.; Shewry, P. Effects of nitrogen nutrition on the synthesis and deposition of the ω-gliadins of wheat. Ann. Bot. 2014, 113, 607–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieser, H.; Manderscheid, R.; Erbs, M.; Weigel, H.-J. Effects of Elevated Atmospheric CO2 Concentrations on the Quantitative Protein Composition of Wheat Grain. J. Agric. Food Chem. 2008, 56, 6531–6535. [Google Scholar] [CrossRef]
- Wojtkowiak, K.; Stępień, A.; Warechowska, M.; Konopka, I.; Klasa, A. Effect of fertilisation technique on some indices of nutritional value of spring triticale grain. J. Elem. 2014, 229–242. [Google Scholar] [CrossRef]
- Wojtkowiak, K.; Stępień, A.; Tańska, M.; Konopka, I. Impact of nitrogen fertilization on the yield and content of protein fractions in spring triticale grain. Afr. J. Agric. Res. 2013, 8, 3778–3783. [Google Scholar] [CrossRef]
- Woźniak, A.; Makarski, B. Content of minerals, total protein and wet gluten in grain of spring wheat depending on cropping systems. J. Elem. 2012, 297–306. [Google Scholar] [CrossRef]
- Yang, R.; Liang, X.; Torrion, J.A.; Christiaens, R.J.; O’Brien, K.; Liu, Q. The Influence of Water and Nitrogen Availability on the Expression of End-Use Quality Parameters of Spring Wheat. Agronomy 2018, 8, 257. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Erley, G.S.A.; Rücker, S.; Koehler, P.; Obenauf, U.; Mühling, K.H. Late nitrogen application increased protein concentration but not baking quality of wheat. J. Plant Nutr. Soil Sci. 2016, 179, 591–601. [Google Scholar] [CrossRef]
- Zecevic, V.; Knežević, D.S.; Boskovic, J.; Milenkovic, S. Effect of nitrogen and ecological factors on quality of winter triticale cultivars. Genetika 2010, 42, 465–474. [Google Scholar] [CrossRef]
- Zhen, S.; Zhou, J.; Deng, X.; Zhu, G.; Cao, H.; Wang, Z.; Yan, Y. Metabolite pro fi ling of the response to high-nitrogen fertilizer during grain development of bread wheat (Triticum aestivum L.). J. Cereal Sci. 2016, 69, 85–94. [Google Scholar] [CrossRef]
- Zheng, T.; Qi, P.-F.; Cao, Y.-L.; Han, Y.-N.; Ma, H.-L.; Guo, Z.-R.; Wang, Y.; Qiao, Y.-Y.; Hua, S.-Y.; Yu, H.-Y.; et al. Mechanisms of wheat (Triticum aestivum) grain storage proteins in response to nitrogen application and its impacts on processing quality. Sci. Rep. 2018, 8, 11928. [Google Scholar] [CrossRef]
- Zörb, C.; Grover, C.; Steinfurth, D.; Mühling, K.H. Quantitative proteome analysis of wheat gluten as influenced by N and S nutrition. Plant Soil 2009, 327, 225–234. [Google Scholar] [CrossRef]
- Shewry, P.; Halford, N.G.; Tatham, A.S.; Popineau, Y.; Lafiandra, D.; Belton, P.S. The high molecular weight subunits of wheat glutenin and their role in determining wheat processing properties. Adv. Food Nutr. Res. 2003, 45, 219–302. [Google Scholar] [CrossRef]
- Altenbach, S.B.; Allen, P.V. Transformation of the US bread wheat ‘Butte 86’ and silencing of omega-5 gliadin genes. GM Crop. 2011, 2, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Gao, H.; Du, C.; Li, L.; Sun, W.; Liu, S.; Wang, C.; Xie, Y.; Kang, G. Transcriptomic and Metabolomics Analysis of Different Endosperm Region under Nitrogen Treatments. Int. J. Mol. Sci. 2019, 20, 4212. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Li, F.; Cao, S.; Zhang, K. Genomic and functional genomics analyses of gluten proteins and prospect for simultaneous improvement of end-use and health-related traits in wheat. Theor. Appl. Genet. 2020, 133, 1521–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewry, P.R.; Pellny, T.K.; Lovegrove, A. Is modern wheat bad for health? Nat. Plants 2016, 2, 16097. [Google Scholar] [CrossRef] [PubMed]
- Cichewicz, A.B.; Mearns, E.S.; Taylor, A.; Boulanger, T.; Gerber, M.; Leffler, D.A.; Drahos, J.; Sanders, D.S.; Craig, K.J.T.; Lebwohl, B. Diagnosis and Treatment Patterns in Celiac Disease. Dig. Dis. Sci. 2019, 64, 2095–2106. [Google Scholar] [CrossRef]
- Grode, L.; Bech, B.H.; Jensen, T.M.; Humaidan, P.; Agerholm, I.E.; Plana-Ripoll, O.; Ramlau-Hansen, C.H. Prevalence, incidence, and autoimmune comorbidities of coeliac disease: A nation-wide, population-based study in Denmark from 1977 to 2016. Eur. J. Gastroenterol. Hepatol. 2018, 30, 83–91. [Google Scholar] [CrossRef]
- Levinson-Castiel, R.; Eliakim, R.; Shinar, E.; Perets, T.-T.; Layfer, O.; Levhar, N.; Schvimer, M.; Marderfeld, L.; Ben-Horin, S.; Shamir, R. Rising prevalence of celiac disease is not universal and repeated testing is needed for population screening. United Eur. Gastroenterol. J. 2019, 7, 412–418. [Google Scholar] [CrossRef]
- Maiuri, L.; Troncone, R.; Mayer, M.; Coletta, S.; Picarelli, A.; De Vincenzi, M.; Pavone, V.; Auricchio, S. In vitro Activities of A-Gliadin-Related Synthetic Peptides Damaging Effect on the Atrophic Coeliac Mucosa and Activation of Mucosal Immune Response in the Treated Coeliac Mucosa. Scand. J. Gastroenterol. 1996, 31, 247–253. [Google Scholar] [CrossRef]
- Monguzzi, E.; Marabini, L.; Elli, L.; Vaira, V.; Ferrero, S.; Ferretti, F.; Branchi, F.; Gaudioso, G.; Scricciolo, A.; Lombardo, V.; et al. Gliadin effect on the oxidative balance and DNA damage: An in-vitro, ex-vivo study. Dig. Liver Dis. 2019, 51, 47–54. [Google Scholar] [CrossRef]
- Gil-Humanes, J.; Pistón, F.; Altamirano-Fortoul, R.; Real, A.; Comino, I.; Sousa, C.; Rosell, C.M.; Barro, F. Reduced-Gliadin Wheat Bread: An Alternative to the Gluten-Free Diet for Consumers Suffering Gluten-Related Pathologies. PLoS ONE 2014, 9, e90898. [Google Scholar] [CrossRef] [Green Version]
- FAO; FAOSTAT. DATA. Food Balance. 2020. Available online: http://doi.org/www.fao.org/faostat/en/#data/ (accessed on 6 June 2019).
- Makharia, G.K.; Verma, A.K.; Amarchand, R.; Bhatnagar, S.; Das, P.; Goswami, A.; Bhatia, V.; Ahuja, V.; Gupta, S.D.; Anand, K. Prevalence of celiac disease in the northern part of India: A community based study. J. Gastroenterol. Hepatol. 2011, 26, 894–900. [Google Scholar] [CrossRef]
- Lohi, S.; Mustalahti, K.; Kaukinen, K.; Laurila, K.; Collin, P.; Rissanen, H.; Lohi, O.; Bravi, E.; Gasparin, M.; Reunanen, A.; et al. Increasing prevalence of coeliac disease over time. Aliment. Pharmacol. Ther. 2007, 26, 1217–1225. [Google Scholar] [CrossRef]
- Ludvigsson, J.F.; Rubio-Tapia, A.; Van Dyke, C.T.; Melton, L.J.; Zinsmeister, A.R.; Lahr, B.D.; Murray, J.A. Increasing Incidence of Celiac Disease in a North American Population. Am. J. Gastroenterol. 2013, 108, 818–824. [Google Scholar] [CrossRef] [Green Version]
- Lindfors, K.; Cavallo, P.; Kurppa, K.; Lundin, K.E.A.; Makharia, G.; Mearin, M.L.; Murray, J.A.; Verdu, E.F.; Kaukinen, K. Coeliac disease. Nat. Rev. Dis. Prim. 2019, 5, 3. [Google Scholar] [CrossRef]
- Riskó, T.C.; Péntek, Á.; Wiwczaroski, T. Bread consumption habits in the gluten free diet. Appl. Stud. Agribus. Commer. 2017, 11, 113–119. [Google Scholar] [CrossRef]
- Abadie, V.; Sollid, L.M.; Barreiro, L.B.; Jabri, B. Integration of Genetic and Immunological Insights into a Model of Celiac Disease Pathogenesis. Annu. Rev. Immunol. 2011, 29, 493–525. [Google Scholar] [CrossRef] [Green Version]
- Cummins, A.G.; Roberts-Thomson, I.C. Prevalence of celiac disease in the Asia-Pacific region. J. Gastroenterol. Hepatol. 2009, 24, 1347–1351. [Google Scholar] [CrossRef]
- Kang, J.Y.; Kang, A.H.Y.; Green, A.; Gwee, K.A.; Ho, K.Y. Systematic review: Worldwide variation in the frequency of coeliac disease and changes over time. Aliment. Pharmacol. Ther. 2013, 38, 226–245. [Google Scholar] [CrossRef]
- Kucek, L.K.; Veenstra, L.D.; Amnuaycheewa, P.; Sorrells, M.E. A Grounded Guide to Gluten: How Modern Genotypes and Processing Impact Wheat Sensitivity. Compr. Rev. Food Sci. Food Saf. 2015, 14, 285–302. [Google Scholar] [CrossRef]
- Del Pozo, A.; Matus, I.; Serret, M.D.; Araus, J.L. Agronomic and physiological traits associated with breeding advances of wheat under high-productive Mediterranean conditions. The case of Chile. Environ. Exp. Bot. 2014, 103, 180–189. [Google Scholar] [CrossRef]
- Xynias, I.N.; Mylonas, I.; Korpetis, E.G.; Ninou, E.; Tsaballa, A.; Avdikos, I.D.; Mavromatis, A.G. Durum Wheat Breeding in the Mediterranean Region: Current Status and Future Prospects. Agronomy 2020, 10, 432. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, M.; Rodríguez-Quijano, M.; Giraldo, P.; Pinto, L.; Vázquez, J.F.; Carrillo, J.M.; Igrejas, G. Effect of allelic variation at glutenin and puroindoline loci on bread-making quality: Favorable combinations occur in less toxic varieties of wheat for celiac patients. Eur. Food Res. Technol. 2016, 243, 743–752. [Google Scholar] [CrossRef]
- Van den Broeck, H.C.; De Jong, H.C.; Salentijn, E.M.J.; Dekking, L.; Bosch, D.; Hamer, R.J.; Gilissen, L.J.W.J.; Van Der Meer, I.M.; Smulders, M.J.M. Presence of celiac disease epitopes in modern and old hexaploid wheat varieties: Wheat breeding may have contributed to increased prevalence of celiac disease. Theor. Appl. Genet. 2010, 121, 1527–1539. [Google Scholar] [CrossRef] [Green Version]
- Spisni, E.; Imbesi, V.; Giovanardi, E.; Petrocelli, G.; Alvisi, P.; Valerii, M.C. Differential Physiological Responses Elicited by Ancient and Heritage Wheat Cultivars Compared to Modern Ones. Nutrients 2019, 11, 2879. [Google Scholar] [CrossRef] [Green Version]
- Di Francesco, A.; Saletti, R.; Cunsolo, V.; Svensson, B.; Muccilli, V.; De Vita, P.; Foti, S. Qualitative proteomic comparison of metabolic and CM-like protein fractions in old and modern wheat Italian genotypes by a shotgun approach. J. Proteom. 2019, 211, 103530. [Google Scholar] [CrossRef] [PubMed]
- Pilolli, R.; Gadaleta, A.; Mamone, G.; Nigro, D.; De Angelis, E.; Montemurro, N.; Monaci, L. Scouting for Naturally Low-Toxicity Wheat Genotypes by a Multidisciplinary Approach. Sci. Rep. 2019, 9, 1646. [Google Scholar] [CrossRef] [PubMed]
- Prandi, B.; Bencivenni, M.; Faccini, A.; Tedeschi, T.; Dossena, A.; Marchelli, R.; Galaverna, G.; Sforza, S. Composition of peptide mixtures derived from simulated gastrointestinal digestion of prolamins from different wheat varieties. J. Cereal Sci. 2012, 56, 223–231. [Google Scholar] [CrossRef]
- Prandi, B.; Mantovani, P.; Galaverna, G.; Sforza, S. Genetic and environmental factors affecting pathogenicity of wheat as related to celiac disease. J. Cereal Sci. 2014, 59, 62–69. [Google Scholar] [CrossRef]
- Ribeiro, M.; Nunes, F.M. We might have got it wrong: Modern wheat is not more toxic for celiac patients. Food Chem. 2019, 278, 820–822. [Google Scholar] [CrossRef]
- Shewry, P.R. Do ancient types of wheat have health benefits compared with modern bread wheat? J. Cereal Sci. 2018, 79, 469–476. [Google Scholar] [CrossRef]
- Sievers, S.; Rohrbach, A.; Beyer, K. Wheat-induced food allergy in childhood: Ancient grains seem no way out. Eur. J. Nutr. 2019, 59, 2693–2707. [Google Scholar] [CrossRef]
- Simsek, S.; Budak, B.; Schwebach, C.S.; Ovando-Martínez, M. Historical vs. modern hard red spring wheat: Analysis of the chemical composition. Cereal Chem. J. 2019, 96, 937–949. [Google Scholar] [CrossRef]
- Pronin, D.; Börner, A.; Scherf, K.A. Old and modern wheat (Triticum aestivum L.) cultivars and their potential to elicit celiac disease. Food Chem. 2021, 339, 127952. [Google Scholar] [CrossRef]
- Colomba, M.S.; Gregorini, A. Are Ancient Durum Wheats Less Toxic to Celiac Patients? A Study ofα-Gliadin from Graziella Ra and Kamut. Sci. World J. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Grover, J.; Chhuneja, P.; Midha, V.; Ghia, J.E.; Deka, D.; Mukhopadhyay, C.S.; Sood, N.; Mahajan, R.; Singh, A.; Verma, R.; et al. Variable Immunogenic Potential of Wheat: Prospective for Selection of Innocuous Varieties for Celiac Disease Patients via in vitro Approach. Front. Immunol. 2019, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Malalgoda, M.; Meinhardt, S.W.; Simsek, S. Detection and quantitation of immunogenic epitopes related to celiac disease in historical and modern hard red spring wheat cultivars. Food Chem. 2018, 264, 101–107. [Google Scholar] [CrossRef]
- Šuligoj, T.; Gregorini, A.; Colomba, M.; Ellis, H.J.; Ciclitira, P.J. Evaluation of the safety of ancient strains of wheat in coeliac disease reveals heterogeneous small intestinal T cell responses suggestive of coeliac toxicity. Clin. Nutr. 2013, 32, 1043–1049. [Google Scholar] [CrossRef]
- Boukid, F.; Prandi, B.; Sforza, S.; Sayar, R.; Seo, Y.W.; Mejri, M.; Yacoubi, I. Understanding the Effects of Genotype, Growing Year, and Breeding on Tunisian Durum Wheat Allergenicity. 2. The Celiac Disease Case. J. Agric. Food Chem. 2017, 65, 5837–5846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prandi, B.; Tedeschi, T.; Folloni, S.; Galaverna, G.; Sforza, S. Peptides from gluten digestion: A comparison between old and modern wheat varieties. Food Res. Int. 2017, 91, 92–102. [Google Scholar] [CrossRef]
- Keirns, B.H.; Anderson, K.L.; Ojo, B.A.; Washburn, K.F.; El Rassi, G.D.; Lightfoot, S.A.; Carver, B.F.; Lucas, E.A.; Smith, B.J. A Comparative Study of Modern and Heirloom Wheat on Indicators of Gastrointestinal Health. J. Agric. Food Chem. 2019, 67, 14027–14037. [Google Scholar] [CrossRef]
- Swanson, N.L.; Leu, A.; Abrahamson, J.; Wallet, B. Genetically engineered crops, glyphosate and the deterioration of health in the United States of America. J. Org. Syst. 2014, 9, 6–37. [Google Scholar]
- Castellanos-Rubio, A.; Fernandez-Jimenez, N.; Kratchmarov, R.; Luo, X.; Bhagat, G.; Green, P.H.R.; Schneider, R.; Kiledjian, M.; Bilbao, J.R.; Ghosh, S. A long noncoding RNA associated with susceptibility to celiac disease. Science 2016, 352, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Bouziat, R.; Hinterleitner, R.; Brown, J.J.; Stencel-Baerenwald, J.E.; Ikizler, M.; Mayassi, T.; Meisel, M.; Kim, S.M.; Discepolo, V.; Pruijssers, A.J.; et al. Reovirus infection triggers inflammatory responses to dietary antigens and development of celiac disease. Science 2017, 356, 44–50. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penuelas, J.; Gargallo-Garriga, A.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Klem, K.; Urban, O.; Zhu, Y.-G.; Sardans, J. Could Global Intensification of Nitrogen Fertilisation Increase Immunogenic Proteins and Favour the Spread of Coeliac Pathology? Foods 2020, 9, 1602. https://doi.org/10.3390/foods9111602
Penuelas J, Gargallo-Garriga A, Janssens IA, Ciais P, Obersteiner M, Klem K, Urban O, Zhu Y-G, Sardans J. Could Global Intensification of Nitrogen Fertilisation Increase Immunogenic Proteins and Favour the Spread of Coeliac Pathology? Foods. 2020; 9(11):1602. https://doi.org/10.3390/foods9111602
Chicago/Turabian StylePenuelas, Josep, Albert Gargallo-Garriga, Ivan A. Janssens, Philippe Ciais, Michael Obersteiner, Karel Klem, Otmar Urban, Yong-Guan Zhu, and Jordi Sardans. 2020. "Could Global Intensification of Nitrogen Fertilisation Increase Immunogenic Proteins and Favour the Spread of Coeliac Pathology?" Foods 9, no. 11: 1602. https://doi.org/10.3390/foods9111602