
Cardiac Cx43 Signaling Is Enhanced and TGF-β1/SMAD2/3 Suppressed in Response to Cold Acclimation and Modulated by Thyroid Status in Hairless SHRM

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Biometric, Blood Samples and Cardiac Left Ventricular Tissue Parameters Monitoring
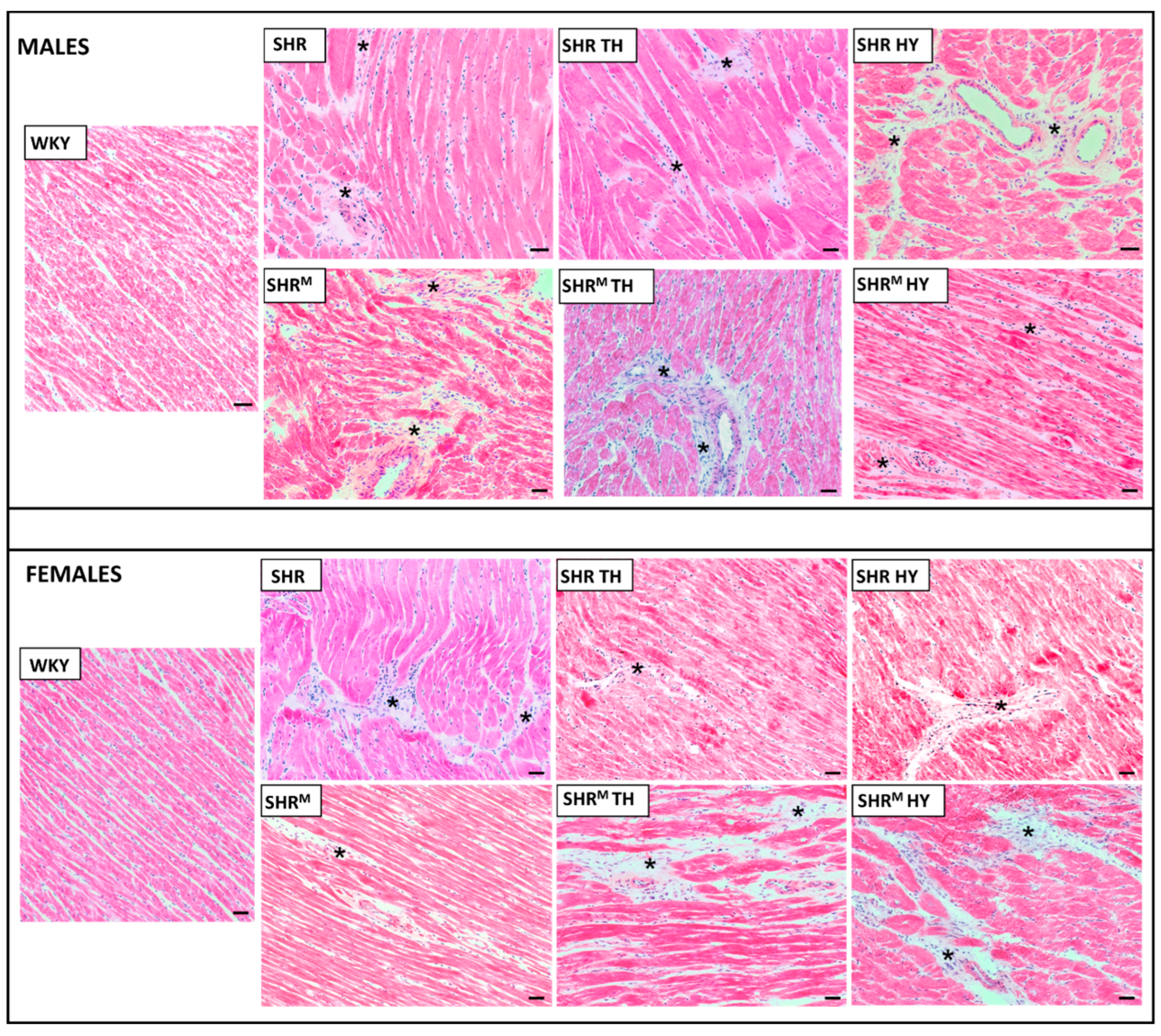
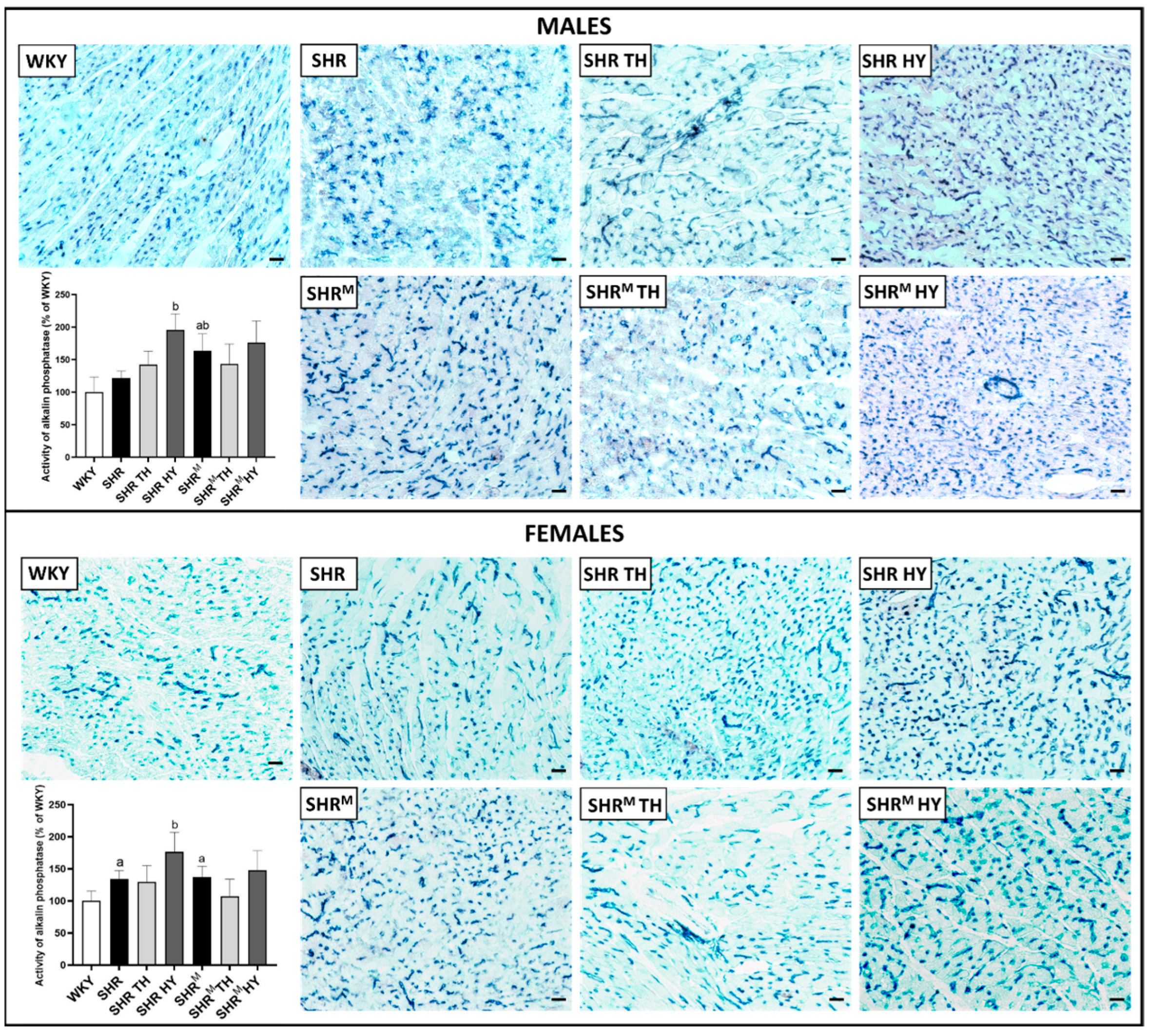
2.3. Myocardial Histology and Enzyme Histochemistry
2.4. Immunofluorescence Labelling of Myocardial Cx43, Cadherin, β-Catenin and Quantitative Image Analysis
2.5. Determination of Collagen Content by Hydroxyproline Measurement
2.6. Determination of Myocardial Protein Levels by Western Blotting
2.7. Statistical Evaluation
3. Results
3.1. Biometric, Blood Samples and Cardiac Left Ventricular Tissue Parameters of Experimental Rats
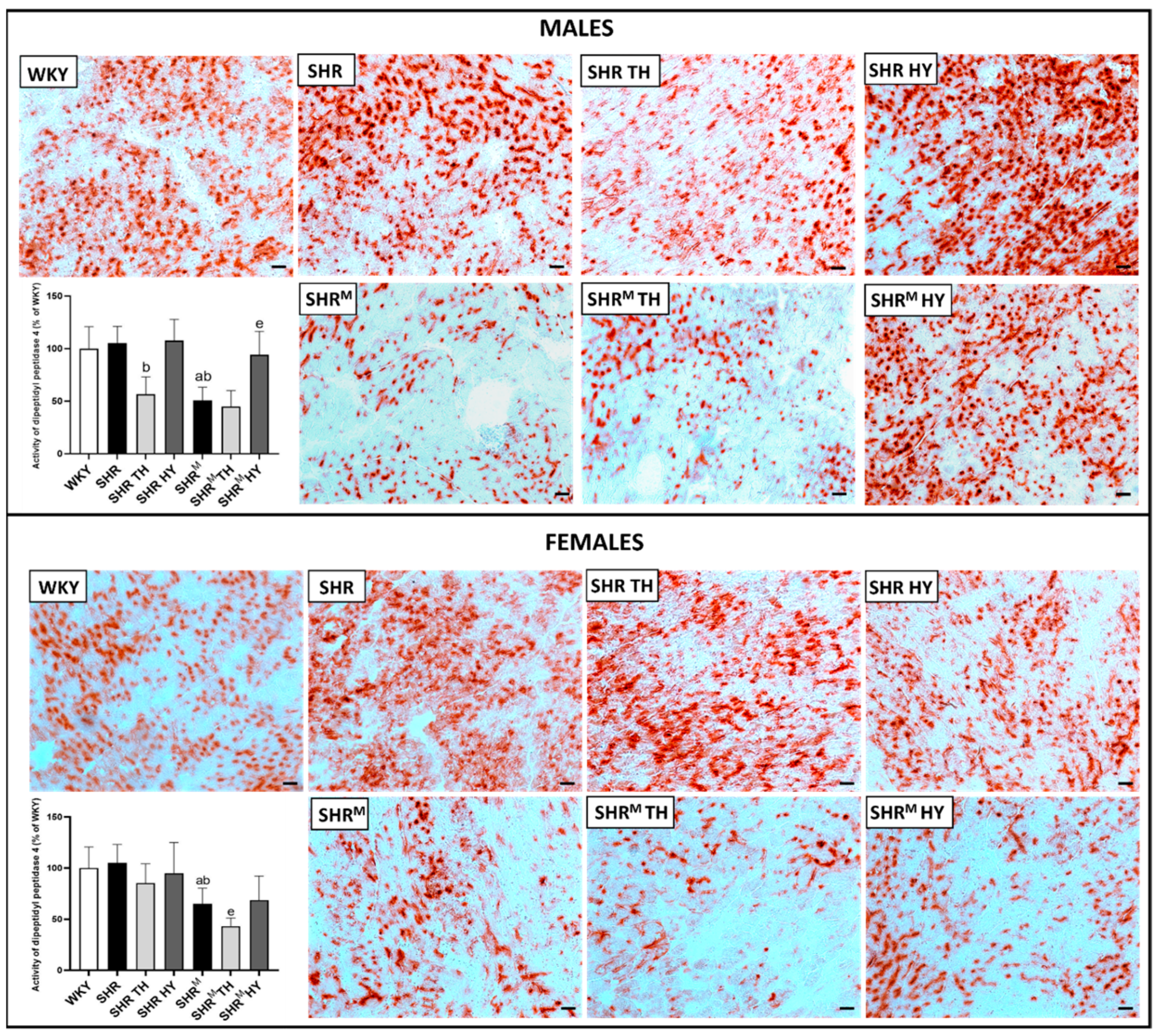
3.2. Myocardial Histology and Capillary Enzyme Histochemistry
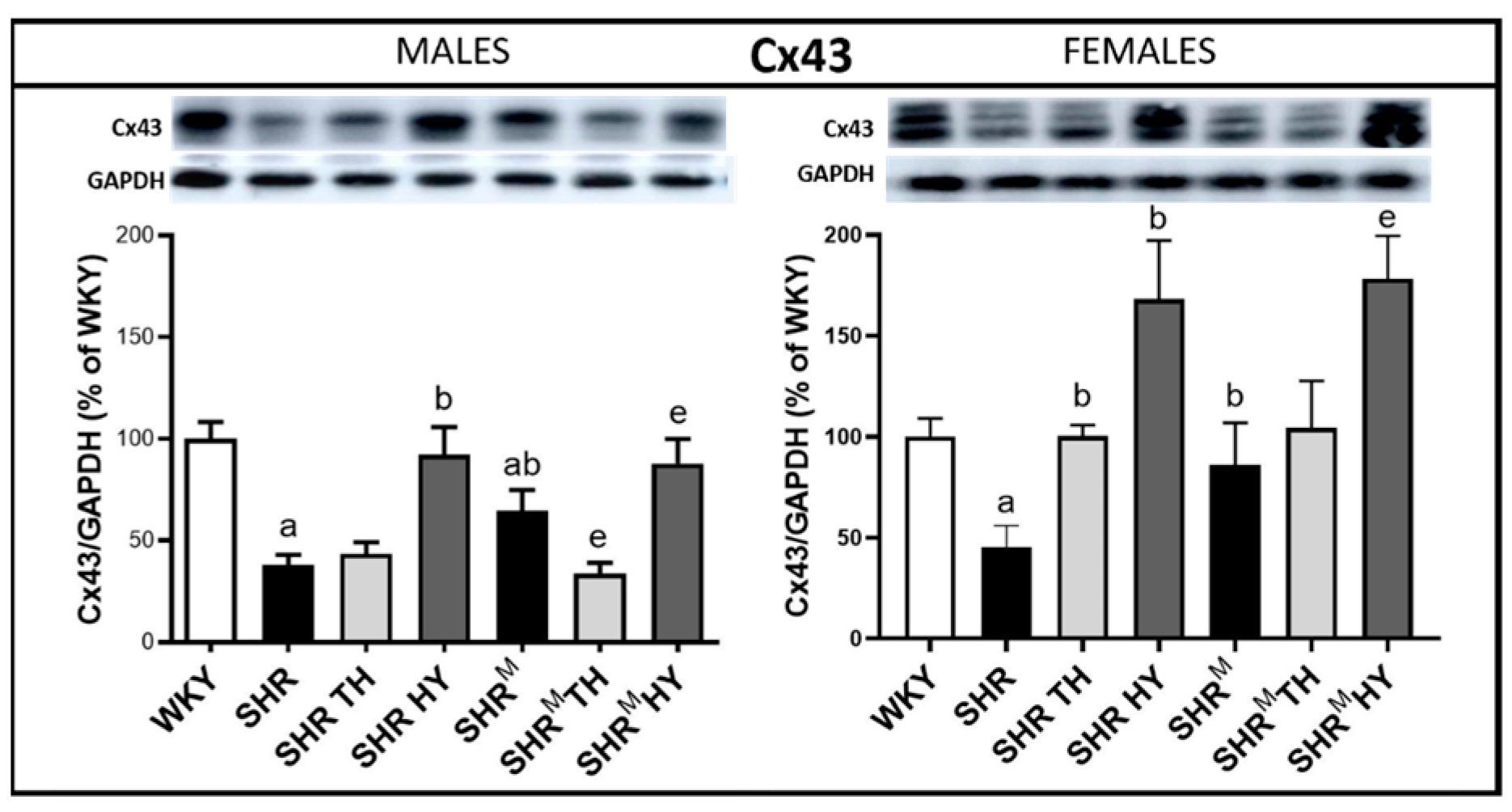
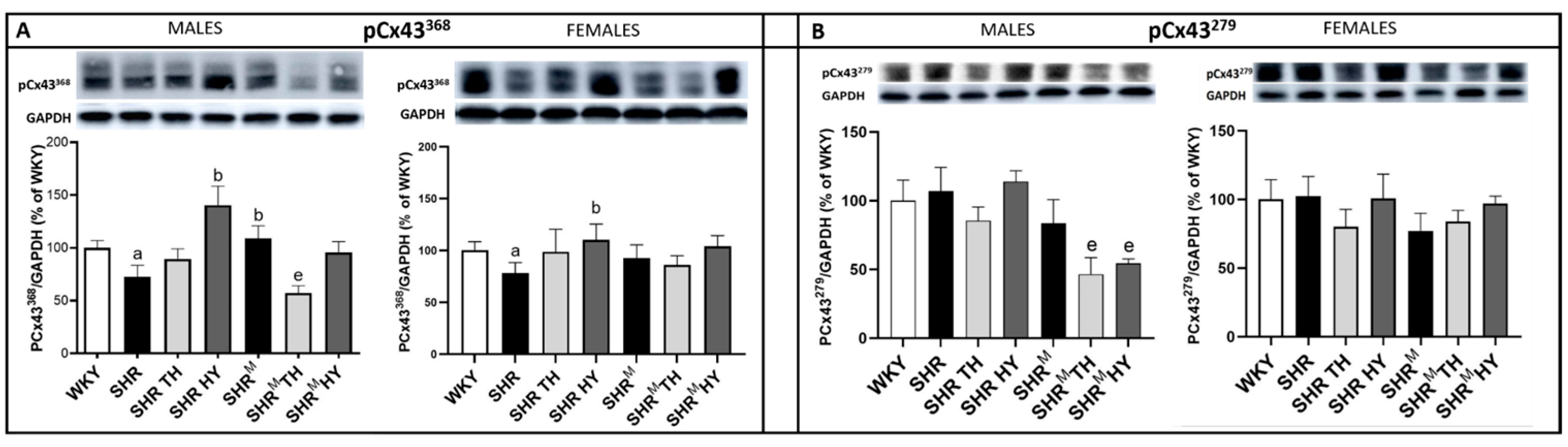
3.3. Myocardial Protein Levels of Cx43 and Its Variants
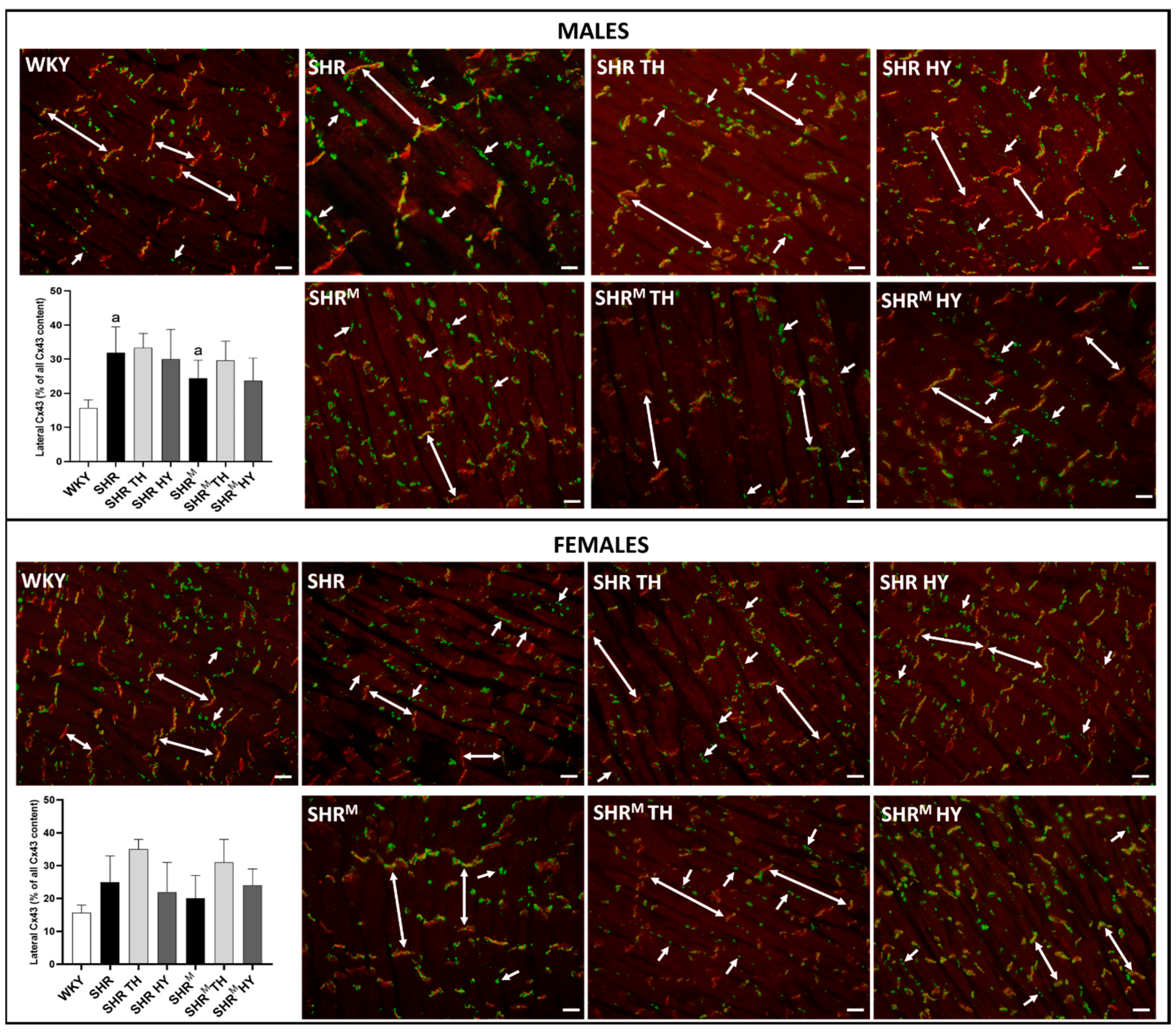
3.4. Myocardial Topology of Cx43 and Quantification of Its Abnormal Distribution
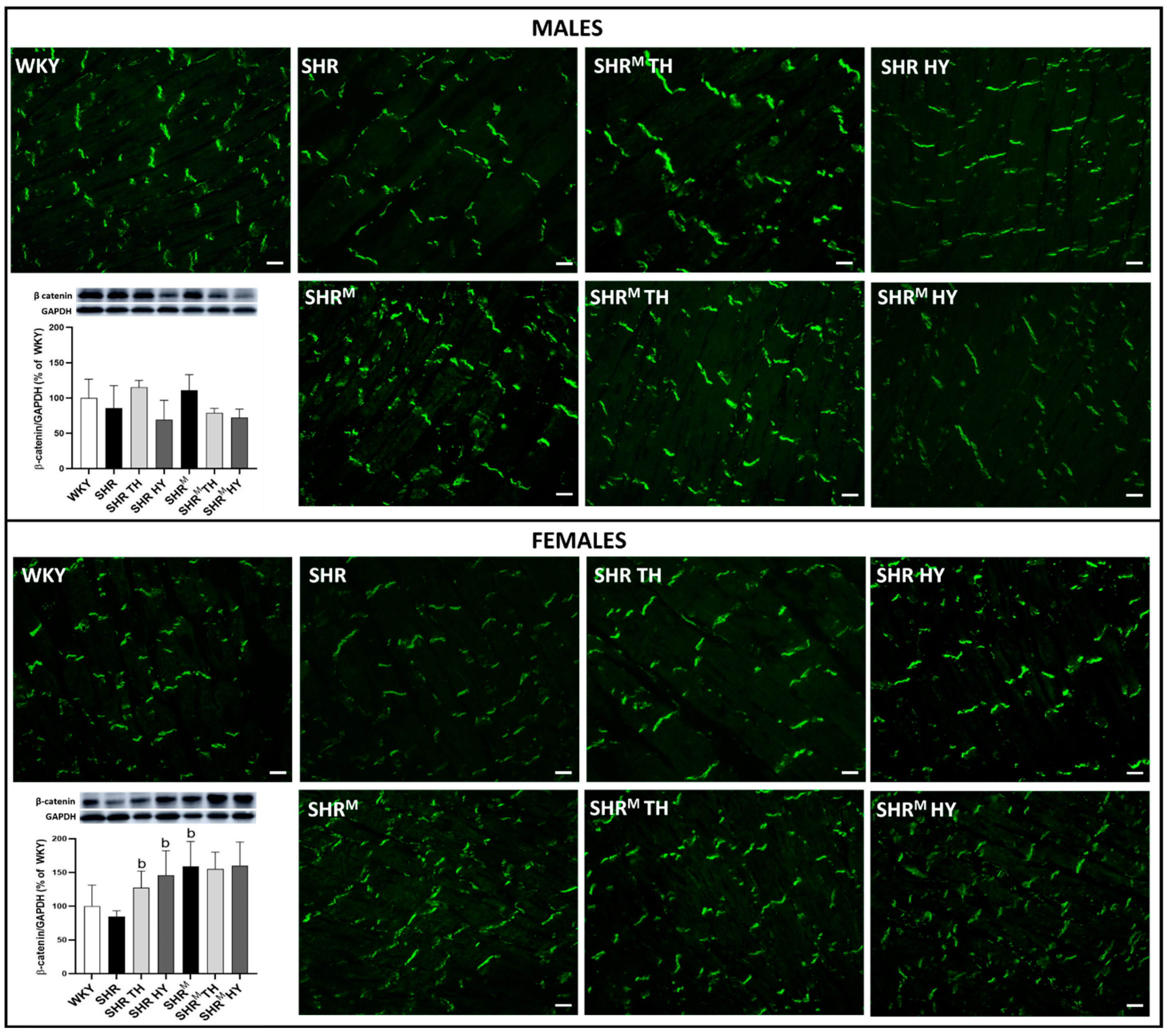
3.5. Myocardial Protein Levels and Topology of Cx43 Interacting Protein, β-Catenin
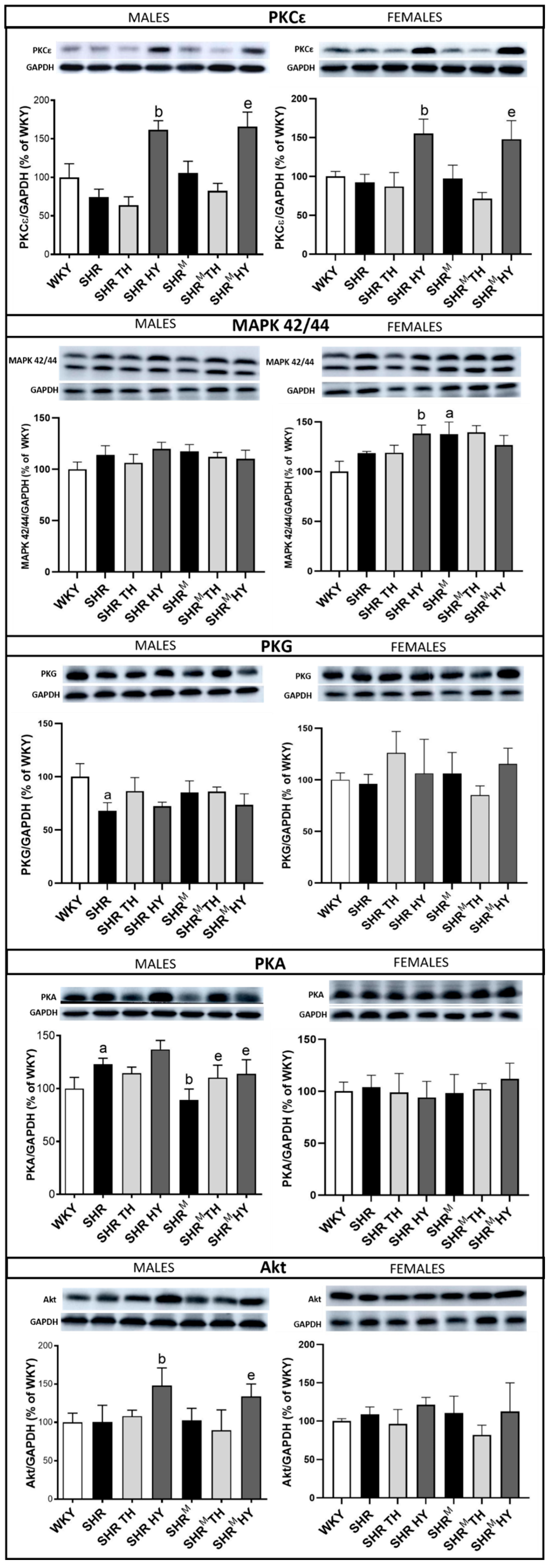
3.6. Myocardial Expression of Cx43 Interacting Protein Kinases
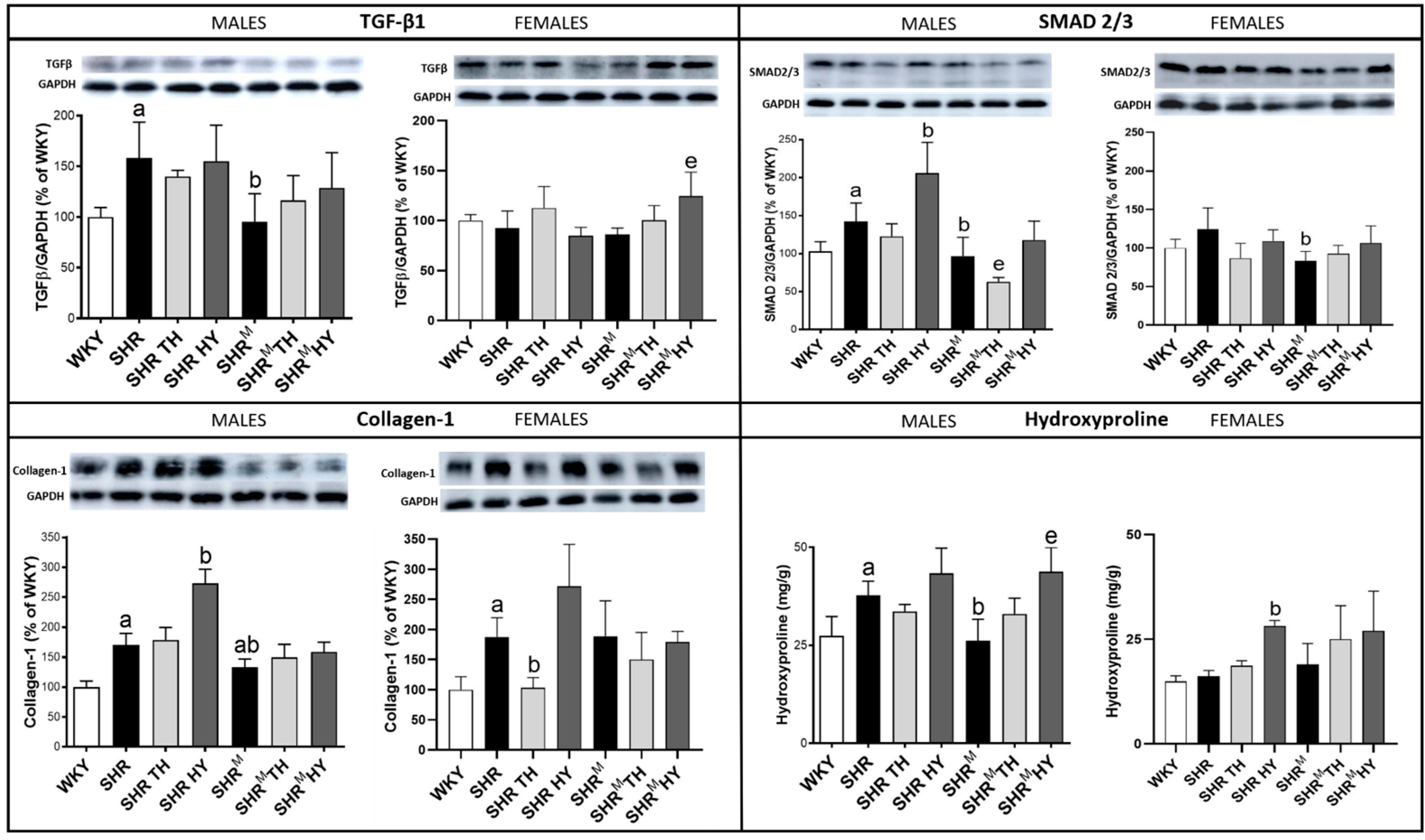
3.7. Myocardial Level of Profibrotic Markers TGF-β1, SMAD2/3, Collagen-1 and Hydroxyproline
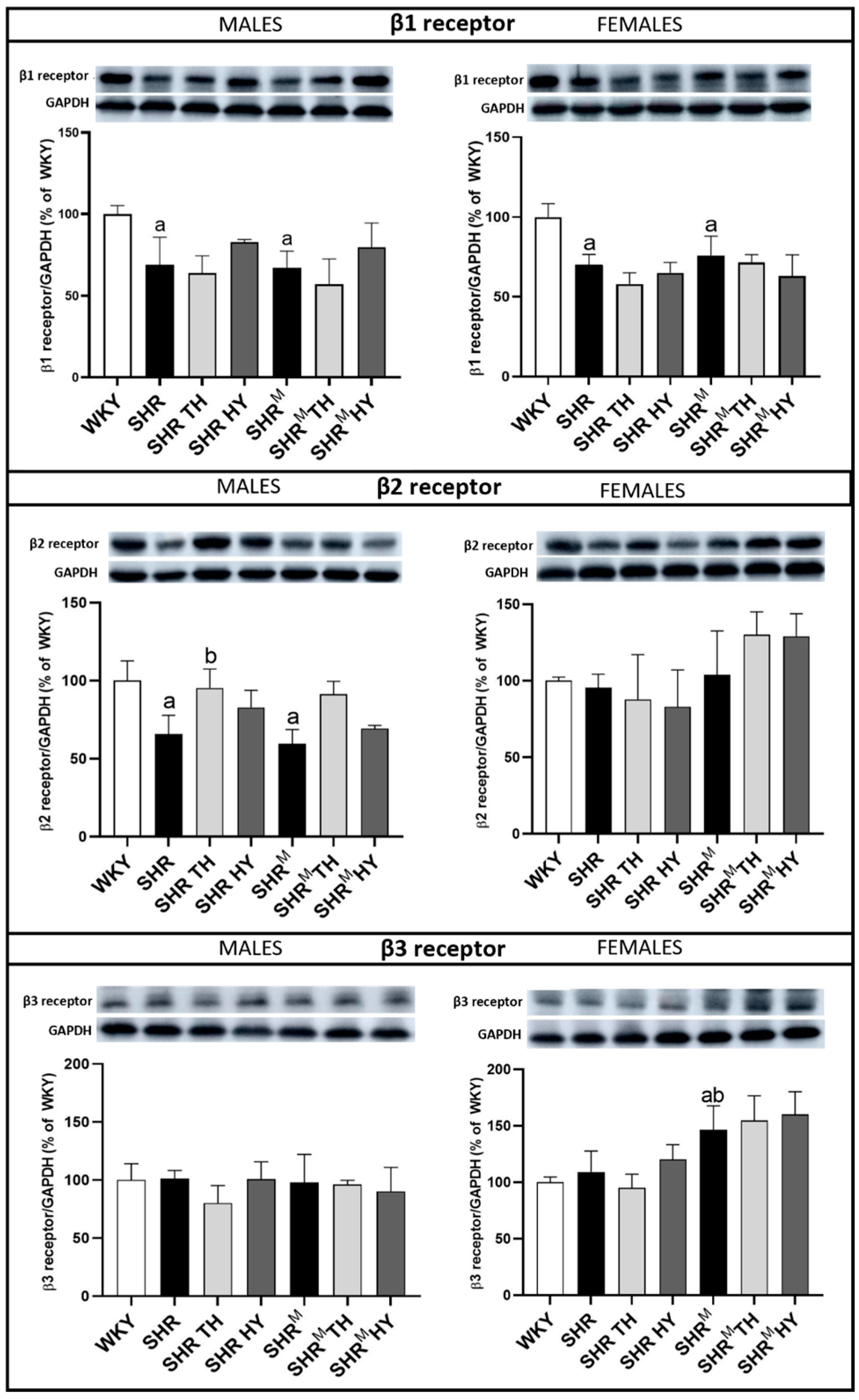
3.8. Myocardial Protein Levels of β1, β2 and β3-Adrenergic Receptors (AR)
4. Discussion
5. Conclusions
Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morrison, S.F. Central neural pathways for thermoregulation. Front. Biosci. 2011, 16, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, T.; Nakao, T.; Miyasaka, Y.; Nishino, T.; Matsumura, S.; Nakazeki, F.; Ide, Y.; Kimura, M.; Tsuji, S.; Rodriguez, R.R.; et al. microRNA-33 maintains adaptive thermogenesis via enhanced sympathetic nerve activity. Nat. Commun. 2021, 12, 843. [Google Scholar] [CrossRef] [PubMed]
- Yau, W.W.; Yen, P.M. Thermogenesis in adipose tissue activated by thyroid hormone. Int. J. Mol. Sci. 2020, 21, 3020. [Google Scholar] [CrossRef]
- Tsibulnikov, S.; Maslov, L.; Voronkov, N.; Oeltgen, P. Thyroid hormones and the mechanisms of adaptation to cold. Hormones 2020, 19, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Sentis, S.C.; Oelkrug, R.; Mittag, J. Thyroid hormones in the regulation of brown adipose tissue thermogenesis. Endocr. Connect. 2021, 10, R106–R115. [Google Scholar] [CrossRef]
- Kralova Lesna, I.; Rychlikova, J.; Vavrova, L.; Vybiral, S. Could human cold adaptation decrease the risk of cardiovascular disease? J. Therm. Biol. 2015, 52, 192–198. [Google Scholar] [CrossRef]
- Tibenska, V.; Benesova, A.; Vebr, P.; Liptakova, A.; Hejnová, L.; Elsnicová, B.; Drahota, Z.; Hornikova, D.; Galatík, F.; Kolar, D.; et al. Gradual cold acclimation induces cardioprotection without affecting β-adrenergic receptor-mediated adenylyl cyclase signaling. J. Appl. Physiol. 2020, 128, 1023–1032. [Google Scholar] [CrossRef]
- Rodríguez-Sinovas, A.; Sánchez, J.A.; Valls-Lacalle, L.; Consegal, M.; Ferreira-González, I. Connexins in the heart: Regulation, function and involvement in cardiac disease. Int. J. Mol. Sci. 2021, 22, 4413. [Google Scholar] [CrossRef]
- Andelova, K.; Benova, T.E.; Bacova, B.S.; Sykora, M.; Prado, N.J.; Diez, E.R.; Hlivak, P.; Tribulova, N. Cardiac connexin-43 hemichannels and pannexin1 channels: Provocative antiarrhythmic targets. Int. J. Mol. Sci. 2021, 22, 260. [Google Scholar] [CrossRef]
- Tribulova, N.; Szeiffova Bacova, B.; Benova, T.; Viczenczova, C. Can we protect from malignant arrhythmias by modulation of cardiac cell-to-cell coupling? J. Electrocardiol. 2015, 48, 434–440. [Google Scholar] [CrossRef]
- Dhein, S.; Salameh, A. Remodeling of cardiac gap junctional cell–cell coupling. Cells 2021, 10, 2422. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.A.; Van Veen, T.A.B.; De Jong, S.; Van Der Nagel, R.; Van Stuijvenberg, L.; Driessen, H.; Labzowski, R.; Oefner, C.M.; Bosch, A.A.; Nguyen, T.Q.; et al. Reduced Cx43 expression triggers increased fibrosis due to enhanced fibroblast activity. Circ. Arrhythmia Electrophysiol. 2012, 5, 380–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan Benova, T.; Szeiffova Bacova, B.; Viczenczova, C.; Diez, E.; Barancik, M.; Tribulova, N. Protection of cardiac cell-to-cell coupling attenuate myocardial remodeling and proarrhythmia induced by hypertension. Physiol. Res. 2016, 65, S29–S42. [Google Scholar] [CrossRef] [PubMed]
- Saitongdee, P.; Milner, P.; Becker, D.L.; Knight, G.E.; Burnstock, G. Increased connexin43 gap junction protein in hamster cardiomyocytes during cold acclimatization and hibernation. Cardiovasc. Res. 2000, 47, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Fedorov, V.V.; Li, L.; Glukhov, A.; Shishkina, I.; Aliev, R.R.; Mikheeva, T.; Nikolski, V.P.; Rosenshtraukh, L.V.; Efimov, I.R. Hibernator Citellus undulatus maintains safe cardiac conduction and is protected against tachyarrhythmias during extreme hypothermia: Possible role of Cx43 and Cx45 up-regulation. Heart Rhythm 2005, 2, 966–975. [Google Scholar] [CrossRef]
- Trnovská, J.; Šilhavỳ, J.; Zídek, V.; Šimáková, M.; Mlejnek, P.; Landa, V.; Eigner, S.; Eigner Henke, K.; Škop, V.; Oliyarnyk, O.; et al. Gender-related effects on substrate utilization and metabolic adaptation in hairless spontaneously hypertensive rat. Physiol. Res. 2015, 64, 51–60. [Google Scholar] [CrossRef]
- Sykora, M.; Bacova, B.S.; Benova, T.E.; Barancik, M.; Zurmanova, J.; Rauchova, H.; Weismann, P.; Pavelka, S.; Kurahara, L.H.; Slezak, J.; et al. Cardiac cx43 and ECM responses to altered thyroid status are blunted in spontaneously hypertensive versus normotensive rats. Int. J. Mol. Sci. 2019, 20, 3758. [Google Scholar] [CrossRef] [Green Version]
- Behuliak, M.; Vavrəínová, A.; Bencze, M.; Polgárová, K.; Ergang, P.; Kunesə, J.; Vaneəcəková, I.; Zicha, J. Ontogenetic changes in contribution of calcium sensitization and calciumentry to blood pressure maintenance ofWistar-Kyoto and spontaneously hypertensive rats. J. Hypertens. 2015, 33, 2443–2454. [Google Scholar] [CrossRef]
- Pavelka, S. Development of radiometric assays for quantification of enzyme activities of the key enzymes of thyroid hormones metabolism. Physiol. Res. 2014, 63, S133–S140. [Google Scholar] [CrossRef]
- Rauchová, H.; Vokurková, M.; Pavelka, S.; Vaněčková, I.; Tribulová, N.; Soukup, T. Red palm oil supplementation does not increase blood glucose or serum lipids levels in wistar rats with different thyroid status. Physiol. Res. 2018, 67, 307–315. [Google Scholar] [CrossRef]
- Lojda, Z. Studies on dipeptidyl(amino)peptidase IV (glycyl-proline naphthylamidase). Histochemistry 1979, 59, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Bacova, B.S.; Viczenczova, C.; Andelova, K.; Sykora, M.; Chaudagar, K.; Barancik, M.; Adamcova, M.; Knezl, V.; Benova, T.E.; Weismann, P.; et al. Antiarrhythmic effects of melatonin and omega-3 are linked with protection of myocardial cx43 topology and suppression of fibrosis in catecholamine stressed normotensive and hypertensive rats. Antioxidants 2020, 9, 546. [Google Scholar] [CrossRef] [PubMed]
- Kesava Reddy, G.; Enwemeka, C.S. A simplified method for the analysis of hydroxyproline in biological tissues. Clin. Biochem. 1996, 29, 225–229. [Google Scholar] [CrossRef]
- Trease, A.J.; Capuccino, J.M.V.; Contreras, J.; Harris, A.L.; Sorgen, P.L. Intramolecular signaling in a cardiac connexin: Role of cytoplasmic domain dimerization. J. Mol. Cell. Cardiol. 2017, 111, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Chen, P.; Xu, Z.; Li, F.; Yi, X.P. Expression and redistribution of β-catenin in the cardiac myocytes of left ventricle of spontaneously hypertensive rat. J. Mol. Histol. 2013, 44, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Axelsen, L.N.; Calloe, K.; Holstein-Rathlou, N.-H.; Nielsen, M.S. Managing the complexity of communication: Regulation of gap junctions by post-translational modification. Front. Pharmacol. 2013, 4, 130. [Google Scholar] [CrossRef] [Green Version]
- Solan, J.L.; Lampe, P.D. Spatio-temporal regulation of connexin43 phosphorylation and gap junction dynamics. Biochim. Biophys. Acta-Biomembr. 2018, 1860, 83–90. [Google Scholar] [CrossRef]
- Dubois-Deruy, E.; Gelinas, R.; Beauloye, C.; Esfahani, H.; Michel, L.Y.M.; Dessy, C.; Bertrand, L.; Balligand, J.L. Beta 3 adrenoreceptors protect from hypertrophic remodelling through AMP-activated protein kinase and autophagy. ESC Heart Fail. 2020, 7, 920–932. [Google Scholar] [CrossRef]
- Hermida, N.; Michel, L.; Esfahani, H.; Dubois-Deruy, E.; Hammond, J.; Bouzin, C.; Markl, A.; Colin, H.; Van Steenbergen, A.; De Meester, C.; et al. Cardiac myocyte β3-adrenergic receptors prevent myocardial fibrosis by modulating oxidant stress-dependent paracrine signaling. Eur. Heart J. 2018, 39, 888–897. [Google Scholar] [CrossRef]
- Lambeir, A.M.; Durinx, C.; Scharpé, S.; De Meester, I. Dipeptidyl-peptidase IV from bench to bedside: An update on structural properties, functions, and clinical aspects of the enzyme DPP IV. Crit. Rev. Clin. Lab. Sci. 2003, 40, 209–294. [Google Scholar] [CrossRef]
- Wang, S.C.; Wang, X.Y.; Liu, C.T.; Chou, R.H.; Chen, Z.B.; Huang, P.H.; Lin, S.J. The Dipeptidyl Peptidase-4 Inhibitor Linagliptin Ameliorates Endothelial Inflammation and Microvascular Thrombosis in a Sepsis Mouse Model. Int. J. Mol. Sci. 2022, 23, 3065. [Google Scholar] [CrossRef] [PubMed]
- Andelova, K.; Bacova, B.S.; Sykora, M.; Hlivak, P.; Barancik, M.; Tribulova, N. Mechanisms Underlying Antiarrhythmic Properties of Cardioprotective Agents Impacting Inflammation and Oxidative Stress. Int. J. Mol. Sci. 2022, 23, 1416. [Google Scholar] [CrossRef] [PubMed]
- Suda, M.; Shimizu, I.; Yoshida, Y.; Hayashi, Y.; Ikegami, R.; Katsuumi, G.; Wakasugi, T.; Yoshida, Y.; Okuda, S.; Soga, T.; et al. Inhibition of dipeptidyl peptidase-4 ameliorates cardiac ischemia and systolic dysfunction by up-regulating the FGF-2/EGR-1 pathway. PLoS ONE 2017, 12, e182422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, B.S.; Li, X.; Baxan, N.; Roney, C.H.; Shchendrygina, A.; Mansfield, C.A.; Jabbour, R.J.; Pitcher, D.S.; Chowdhury, R.A.; Peters, N.S.; et al. Ventricular fibrillation mechanism and global fibrillatory organization are determined by gap junction coupling and fibrosis pattern. Cardiovasc. Res. 2021, 117, 1078–1090. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Reuss, L.; Altenberg, G.A. Regulation of Purified and Reconstituted Connexin 43 Hemichannels by Protein Kinase C-mediated Phosphorylation of Serine 368. J. Biol. Chem. 2004, 279, 20058–20066. [Google Scholar] [CrossRef] [Green Version]
- Swope, D.; Cheng, L.; Gao, E.; Li, J.; Radice, G.L. Loss of Cadherin-Binding Proteins β-Catenin and Plakoglobin in the Heart Leads to Gap Junction Remodeling and Arrhythmogenesis. Mol. Cell. Biol. 2012, 32, 1056–1067. [Google Scholar] [CrossRef] [Green Version]
- Egan Benova, T.; Viczenczova, C.; Szeiffova Bacova, B.; Zurmanova, J.; Knezl, V.; Andelova, K.; Tribulova, N. Omacor Protects Normotensive and Hypertensive Rats Exposed to Continuous Light from Increased Risk to Malignant Cardiac Arrhythmias. Mar. Drugs 2021, 19, 659. [Google Scholar] [CrossRef]
- Grohé, C.; Kahlert, S.; Löbbert, K.; Stimpel, M.; Karas, R.H.; Vetter, H.; Neyses, L. Cardiac myocytes and fibroblasts contain functional estrogen receptors. FEBS Lett. 1997, 416, 107–112. [Google Scholar] [CrossRef]
- Zheng, L.; Trease, A.J.; Katsurada, K.; Spagnol, G.; Li, H.; Shi, W.; Duan, B.; Patel, K.P.; Sorgen, P.L. Inhibition of Pyk2 and Src activity improves Cx43 gap junction intercellular communication. J. Mol. Cell. Cardiol. 2020, 149, 27–40. [Google Scholar] [CrossRef]
- Salameh, A.; Dhein, S. Adrenergic control of cardiac gap junction function and expression. Naunyn. Schmiedebergs. Arch. Pharmacol. 2011, 383, 331–346. [Google Scholar] [CrossRef]
- Tribulova, N.; Kurahara, L.H.; Hlivak, P.; Hirano, K.; Bacova, B.S. Pro-arrhythmic signaling of thyroid hormones and its relevance in subclinical hyperthyroidism. Int. J. Mol. Sci. 2020, 21, 2844. [Google Scholar] [CrossRef] [PubMed]
- Bačová, B.S.; Vinczenzová, C.; Žurmanová, J.; Kašparová, D.; Knezl, V.; Beňová, T.E.; Pavelka, S.; Soukup, T.; Tribulová, N. Altered thyroid status affects myocardial expression of connexin-43 and susceptibility of rat heart to malignant arrhythmias that can be partially normalized by red palm oil intake. Histochem. Cell Biol. 2017, 147, 63–73. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Dilution | Product Number | Manufacturer of Antibody |
|---|---|---|---|
| Cx43 | 1:5000 | C6219 | Sigma-Aldrich, Missouri, USA |
| pCx43368 | 1:1000 | sc-101660 | Santa Cruz Biotechnology, Texas, USA |
| pCx43279 | 1:1000 | sc-12900 | Santa Cruz Biotechnology, Texas, USA |
| β-catenin | 1:1000 | sc-7963 | Santa Cruz Biotechnology, Texas, USA |
| PKCε | 1:1000 | sc-214 | Santa Cruz Biotechnology, Texas, USA |
| TGF-β1 | 1:1000 | SAB4502954 | Sigma-Aldrich, Missouri, USA |
| SMAD2/3 | 1:1000 | #3102 | Cell Signaling Technology, Colorado, USA |
| Collagen-1 | 1:1000 | ab90395 | Abcam, United Kingdom |
| PKA | 1:1000 | sc-365615 | Santa Cruz Biotechnology, Texas, USA |
| PKG | 1:1000 | sc-25429 | Santa Cruz Biotechnology, Texas, USA |
| AktK | 1:1000 | sc-81436 | Santa Cruz Biotechnology, Texas, USA |
| MAPK42/44 | 1:1000 | 137F5 | Cell Signaling Technology, Colorado, USA |
| β1AR | 1:1000 | ab3442 | Abcam, Cambridge, United Kingdom |
| β2AR | 1:1000 | bs-0947R | Bioss, Woburn, Massachusetts, USA |
| β3AR | 1:1000 | bs-1063R | Bioss, Massachusetts, USA |
| GAPDH | 1:1000 | sc-25778 | Santa Cruz Biotechnology, Texas, USA |
| MALES—Variables | WKY | SHR | SHR TH | SHR HY | SHRM | SHRM TH | SHRM HY | |
| BW (g) | 410 ± 1.41 | 398.33 ± 21.08 | 286 ± 1.41 b | 383.5 ± 7.78 | 359 ± 42.76 | 307.33 ± 31.37 | 365.25 ± 38.46 | |
| HW (g) | 1.05 ± 0.01 | 1.53 ± 0.08 a | 1.43 ± 0.05 | 0.87 ± 0.21 b | 1.59 ± 0.13 a | 2.01 ± 0.14 e | 1.14 ± 0.25 e | |
| LVW (g) | 0.75 ± 0.01 | 1.19 ± 0.08 a | 1.06 ± 0.05 | 0.71 ± 0.08 b | 1.21 ± 0.13 a | 1.46 ± 0.11 | 0.89 ± 0.18 e | |
| TBARS (nmol/mg) | 1.75 ± 0.29 | 1.66 ± 0.45 | 2.06 ± 0.11 | 1.99 ± 0.21 | 2.00 ± 0.10 | 2.37 ± 0.45 | 1.75 ± 0.22 | |
| GSH (µmol/g) | 1.99 ± 1.13 | 1.97 ± 0.27 | 2.06 ± 0.33 | 2.43 ± 0.08 | 2.14 ± 0.15 | 2.27 ± 0.24 | 2.39 ± 0.11 | |
| T3 (nmol/L) | 0.84 ± 0.04 | 1.12 ± 0.02 a | 2.84 ± 0.11 b | 0.48 ± 0.06 b | 1.08 ± 0.03 a | 4.05 ± 0.16 e | 0.59 ± 0.01 e | |
| T4 (nmol/L) | 55.0 ± 0.8 | 44.0 ± 6.9 a | 62.3 ± 1.7 b | 12.4 ± 1.9 b | 46.2 ± 2.2 a | 69.0 ± 1.0 e | 16.1 ± 0.7 e | |
| TG (mmol/L) | 1.60 ± 0.20 | 0.66 ± 0.04 a | 0.96 ± 0.02 b | 0.43 ± 0.03 | 0.70 ± 0.13 a | 0.80 ± 0.06 | 0.51 ± 0.08 | |
| TC (mmol/L) | 1.88 ± 0.10 | 1.12 ± 0.02 a | 1.70 ± 0.24 b | 1.98 ± 0.07 b | 1.18 ± 0.25 a | 1.40 ± 0.06 | 2.42 ± 0.34 e | |
| HDL (mmol/L) | 1.33 ± 0.03 | 0.81 ± 0.04 a | 1.38 ± 0.23 b | 1.40 ± 0.05 b | 0.83 ± 0.14 a | 1.12 ± 0.06 | 1.77 ± 0.33 e | |
| LDL (mmol/L) | 0.24 ± 0.09 | 0.17 ± 0.03 | 0.15 ± 0.04 | 0.53 ± 0.10 | 0.20 ± 0.10 | 0.13 ± 0.04 | 0.58 ± 0.21 e | |
| TC/HDL (ratio) | 1.43 ± 0.10 | 1.39 ± 0.04 | 1.24 ± 0.03 | 1.40 ± 0.05 | 1.41 ± 0.09 | 1.26 ± 0.04 | 1.39 ± 0.20 | |
| FEMALES—Variables | WKY | SHR | SHR TH | SHR HY | SHRM | SHRM TH | SHRM HY | |
| BW (g) | 241.41 ± 2.83 | 201.50 ± 1.26 | 197.21 ± 29.70 | 200.67 ± 1.15 | 210.60 ± 13.67 a | 212.50 ± 9.19 | 185.50 ± 33.74 | |
| HW (g) | 0.752 ± 0.07 | 0.827 ± 0.06 a | 1.116 ± 0.25 b | 0.679 ± 0.05 | 0.917 ± 0.10 a | 1.101 ± 0.25 | 0.802 ± 0.14 | |
| LVW (g) | 0.544 ± 0.04 | 0.639 ± 0.02 a | 0.884 ± 0.22 b | 0.512 ± 0.01 b | 0.624 ± 0.23 a | 0.737 ± 0.12 | 0.540 ± 0.16 | |
| TBARS (nmol/mg) | 2.25 ± 0.27 | 2.33 ± 0.25 | 2.90 ± 0.07 | 2.85 ± 0.41 | 2.56 ± 0.43 | 3.51 ± 0.35 e | 3.85 ± 0.27 | |
| GSH (µmol/g) | 1.87 ± 0.07 | 1.98 ± 0.01 | 1.88 ± 0.18 | 2.21 ± 0.46 | 1.98 ± 0.15 | 1.97 ± 0.19 | 2.40 ± 0.31 | |
| T3 (nmol/L) | 0.92 ± 0.02 | 1.29 ± 0.02 a | 2.83 ± 0.11 | 0.38 ± 0.06 b | 1.08 ± 0.04 b | 2.75 ± 0.05 e | 0.51 ± 0.02 e | |
| T4 (nmol/L) | 47.6 ± 2.1 | 42.9 ± 1.8 a | 58.3 ± 1.0b | 14.8 ± 1.5 b | 43.8 ± 0.7 a | 62.7 ± 0.9 e | 18.2 ± 0.3 e | |
| TG (mmol/L) | 1.07 ± 0.26 | 1.28 ± 0.02 | 1.05 ± 0.34 | 0.56 ± 0.05 b | 1.03 ± 0.19 | 1.16 ± 0.14 | 0.39 ± 0.06 e | |
| TC (mmol/L) | 2.28 ± 0.32 | 1.61 ± 0.04 a | 1.86 ± 0.21 | 1.40 ± 0.10 | 1.37 ± 0.06 a | 1.59 ± 0.07 | 2.70 ± 0.07 e | |
| HDL (mmol/L) | 1.85 ± 0.31 | 1.19 ± 0.02 a | 1.47 ± 0.36 | 1.01 ± 0.16 | 1.07 ± 0.08 a | 1.31 ± 0.05 | 2.07 ± 0.17 e | |
| LDL (mmol/L) | 0.21 ± 0.07 | 0.17 ± 0.02 | 0.14 ± 0.04 | 0.28 ± 0.7 | 0.11 ± 0.10 | 0.11 ± 0.06 | 0.63 ± 0.04 e | |
| TC/HDL (ratio) | 1.26 ± 0.05 | 1.36 ± 0.02 | 1.28 ± 0.14 | 1.41 ± 0.13 | 1.29 ± 0.14 | 1.24 ± 0.09 | 1.36 ± 0.05 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andelova, K.; Szeiffova Bacova, B.; Sykora, M.; Pavelka, S.; Rauchova, H.; Tribulova, N. Cardiac Cx43 Signaling Is Enhanced and TGF-β1/SMAD2/3 Suppressed in Response to Cold Acclimation and Modulated by Thyroid Status in Hairless SHRM. Biomedicines 2022, 10, 1707. https://doi.org/10.3390/biomedicines10071707
Andelova K, Szeiffova Bacova B, Sykora M, Pavelka S, Rauchova H, Tribulova N. Cardiac Cx43 Signaling Is Enhanced and TGF-β1/SMAD2/3 Suppressed in Response to Cold Acclimation and Modulated by Thyroid Status in Hairless SHRM. Biomedicines. 2022; 10(7):1707. https://doi.org/10.3390/biomedicines10071707
Chicago/Turabian StyleAndelova, Katarina, Barbara Szeiffova Bacova, Matus Sykora, Stanislav Pavelka, Hana Rauchova, and Narcis Tribulova. 2022. "Cardiac Cx43 Signaling Is Enhanced and TGF-β1/SMAD2/3 Suppressed in Response to Cold Acclimation and Modulated by Thyroid Status in Hairless SHRM" Biomedicines 10, no. 7: 1707. https://doi.org/10.3390/biomedicines10071707