
An Interplay between Epigenetics and Translation in Oocyte Maturation and Embryo Development: Assisted Reproduction Perspective


Institute of Animal Physiology and Genetics, Czech Academy of Sciences, 277 21 Libechov, Czech Republic
*
Author to whom correspondence should be addressed.
Biomedicines 2022, 10(7), 1689; https://doi.org/10.3390/biomedicines10071689
Submission received: 27 May 2022
/
Revised: 25 June 2022
/
Accepted: 28 June 2022
/
Published: 13 July 2022
(This article belongs to the Special Issue Reproductive Medicine: Focus on Cell and Molecule)
Abstract
:Germ cell quality is a key prerequisite for successful fertilization and early embryo development. The quality is determined by the fine regulation of transcriptomic and proteomic profiles, which are prone to alteration by assisted reproduction technology (ART)-introduced in vitro methods. Gaining evidence shows the ART can influence preset epigenetic modifications within cultured oocytes or early embryos and affect their developmental competency. The aim of this review is to describe ART-determined epigenetic changes related to the oogenesis, early embryogenesis, and further in utero development. We confront the latest epigenetic, related epitranscriptomic, and translational regulation findings with the processes of meiotic maturation, fertilization, and early embryogenesis that impact the developmental competency and embryo quality. Post-ART embryo transfer, in utero implantation, and development (placentation, fetal development) are influenced by environmental and lifestyle factors. The review is emphasizing their epigenetic and ART contribution to fetal development. An epigenetic parallel among mouse, porcine, and bovine animal models and human ART is drawn to illustrate possible future mechanisms of infertility management as well as increase the awareness of the underlying mechanisms governing oocyte and embryo developmental complexity under ART conditions.
1. Introduction
The purpose of living organisms’ reproduction is to give rise to a new generation. According to the World Fertility Report from 2015, human fertility has halved in the past 65 years. In the 1950s, the norm was for one woman to give birth to 5 children, whereas in 2015 the average was only 2.5. If this pattern, along with demographic changes, continues, we would gradually face the need to support human fertility clinically by preserving the quality of germ cells, particularly oocytes.
Female fertility is influenced by genetic background, environmental factors, lifestyle, nutrition, psychosocial setting, and many other factors. The proper understanding of the molecular mechanisms underlying oocyte development is crucial for advancement in assisted reproduction technology. This review is focused on how oocyte and embryo development is affected by epigenetic changes, initially those originating from the in vitro methods of assisted reproduction technology (ART) followed by in utero changes caused by maternal diet, lifestyle, and environment. The epigenome is a complex of chemical compounds that modify or mark a genome but are not part of the DNA itself (Figure 1).
Epigenetic changes may cause temporary or heritable alterations of gene expression. Modifications to the epigenome are reversible and alter gene expression in different ways but do not interfere directly with the DNA genetic code. Epigenetic changes are categorized into DNA, RNA, histone modifications, and changes controlled by non-coding RNAs.
DNA methylation is the most studied epigenetic mechanism and plays a key role in transcriptional repression/activation (Figure 1 and Figure 2).
In vertebrates, DNA cytosine methylation is realized by the addition of a methyl group onto the fifth carbon of cytosine residue within cytosine-phosphate-guanin (CpG) sites, where a cytosine is followed by a guanine in the 5′ → 3′ direction of the DNA linear sequence. DNA methylation is functionally associated with gene silencing and is for the most part limited to CpG islands, i.e., areas rich in CpG dinucleotides that are typically located within and nearby sites of about 40% of mammalian gene promoters [4,5]. As the CpG dinucleotides are methylated symmetrically on both DNA strands, their methylation can be heritable during cell division [4].
DNA is methylated by methyltransferases (DNMTs), and the methyl group for methylation is provided by S-adenosylmethionine [6]. DNA methylation relies on the folate-methyl metabolic pathway that supplies the essential methyl groups [7]. There are two distinct types of DNA methylation. During cell division, DNA replication creates hemi-methylated DNA where CpG dinucleotides are methylated only on the original parent strand and methylation is absent from the newly synthesized daughter strand. Subsequently, the methylation maintenance methyltransferase DNMT1 methylates the unmethylated DNA strand [8]. In contrast, de novo methylation of un-methylated double stranded DNA is ensured by DNMT3A and DNMT3B along with their coactivator DNMT3L [9,10].
DNA cytosine methylation can affect DNA activity and when it occurs in a gene promoter, increased DNA methylation leads to a decrease in chromatin accessibility. Subsequently, gene transcription is repressed by recruiting gene-silencing repressive proteins to the methylated region, such as methyl-CpG-binding protein 2 (MeCP2) and methyl-binding-domain (MBD) proteins [11,12]. Methylated DNA also presents a spatial obstacle preventing binding of transcription factors and proteins to gene promoters [13,14]. However, methylation of CpGs over gene bodies is related to both active and repressed transcription according to the tissue in which it occurs [15,16]. Evidently, DNA methylation has dual roles, both inhibitory and permissive, depending on the genomic region.
Epigenetic modifications to RNA, the field of epitranscriptomics, have changed our perception on how epigenetics can directly modulate the translation and stability of mRNAs within oocytes and embryos. There are over 150 post-transcriptional mRNA transcript modifications in eukaryotes, but the most prevalent is the N6-methyladenosine (m6A) methylation (Figure 1) [17]. Generally, m6A transcript methylation is present ubiquitously in every cell and across multiple species. Translation-related m6A modification occurs within the 3′ UTR of mRNA. The interaction between the m6A reader and translation initiation factors is needed for cap-dependent translation [18]. m6A were also found to be capable of governing translation in a cap-independent manner by modifications within internal ribosome entry sites (IRES) [19] or 5′ UTR sequences [20]. A direct link between greater polysome occupancy and the presence of m6A methylation on mRNAs was demonstrated in yeast [21] and HeLa cells [18].
Epigenetic regulation by non-coding RNAs (ncRNAs) recently gained more attention in developmental biology literature. ncRNAs represent RNA molecules that do not code for proteins but have a regulatory role at pre- and post-transcriptional levels [22]. Generally, both short and long ncRNAs do exist. Among the smaller ones are small-interfering RNA (siRNA), microRNA (miRNA), piwi-interacting RNA (piRNA), followed by long ncRNA (lncRNA) (Figure 1). As ncRNAs are also implicated in histone modification and DNA methylation, it has been suggested that gene silencing and upregulation by ncRNAs is a part of the epigenetic mechanism [23,24]. The importance of ncRNAs in regulating primordial germ cell specification, spermatogenesis, and oogenesis has been highlighted (reviewed in [25,26]). ncRNAs are intensely expressed in the early human embryo and their role in early human development remains to be fully investigated. It has been suggested that future studies of the ncRNAs role could expand to the field of ART optimization [27,28].
2. Epigenetics of Germ Cells Development
2.1. Epigenetic Reprogramming
Two different waves of epigenetic reprogramming occur in gametes and the early embryo, the first one during gametogenesis and the second in the preimplantation embryos (Figure 2) [29,30]. During mammalian gametogenesis, pre-existing epigenetic marks are globally deleted in the primordial germ cells. During sex determination in the developing fetus, the global DNA methylation remains at very low levels in both male and female primordial germ cells [31]. However, de novo methylation establishment proceeds in different manners in male and female germlines. In the male gonocytes, a rapid increase of methylation is initiated during embryo development, and the methylome of male germ cells is fully established before the birth [32,33]. In female germ cells, DNA methylation is re-established during the postnatal oocyte growth phase and is dependent on the functional interaction of the DNA methyltransferase Dnmt3a and the Dnmt3-like protein (Dnmt3L) [34,35]. DNA de novo methylation is completed when oocytes reach the germinal vesicle stage [1,36]. After a period of intense activity during oocyte growth, transcription is ceased when the oocyte becomes arrested at the prophase of meiosis I, and this arrest is released after a preovulatory hormonal surge of gonadotropins [37]. It has been proposed that the phenomenon of meiotic arrest possibly protects oocytes from oxidative stress and DNA damage [38]. During oogenesis, the oocyte development and competence is not dependent on DNA methylation as oocytes with genetically ablated DNA methylation were successfully fertilized, and subsequent embryonic development progressed until the mid-gestation stage [39,40]. It has been suggested that methylation of CpG islands in gametes is not fully related to genomic imprinting but is an important factor in the regulation of gene expression in preimplantation embryos [34].
2.2. Histone Deacetylation in Oocyte Maturation and Energy Metabolism
Histones are positively charged proteins, which form the integral part of the chromatin core. The octamer core is composed of four histone variants—H2A, H2B, H3, and H4—with connecting histone H1 [41]. They are essential for packaging the DNA into superstructures, making it inaccessible for DNA-binding proteins to bind it and recruit further transcription or regulation machinery. Important modification regulating histone interactions are classified into methylations, acetylations, phosphorylations, ubiquitinations, and PolyADP ribosylations (Figure 1) [41]. Particularly, the regulation of histone acetylations is relevant for the final moments of oocyte maturation.
Histone deacetylases (HDACs) are enzymes that regulate a wide range of biological processes by removing acetyl groups from lysine ϵ-amino groups, not only on histones but also on many other proteins [42]. In particular, meiotic progression was discovered to be dependent on HDACs activity progression in mice. Histone acetylation of lysine residues was substantially reduced upon meiotic resumption in mouse oocytes [43]. Maintenance of genome integrity and chromatin structure is controlled by HDAC3. In meiosis, HDAC3 is located on the spindle. HDAC3 knockdown experiments on mice revealed defects in chromosome alignment, spindle structure, and microtubule-kinetochore attachment (MT-K) [44]. Suppression of HDAC3 activity in porcine oocytes led to similar phenotypes as in mice, spindle defects, chromosomal congression failure, and meiosis inhibition [45]. Disruption of oocyte maturation by the selective HDAC6 inhibitor tubastatin-A induced asymmetric division in maturing oocytes, failure to extrude the first polar body [46], increased α-tubulin acetylation, and incorrect MT-K attachment as seen in HDAC3 [47]. Recently, primordial follicle activation by mTOR signalling was associated with decreased HDAC6 activity. This finding may be of importance for the management of premature ovarian failure (POF) as an alternative approach to primordial follicle in vitro activation (IVA) [48]. HDAC8 has a similarly indispensable role as HDAC6 and is located on spindle poles. Its absence led to the defective recruitment of γ-tubulin and consequently caused aberrant spindle morphology and chromosome misalignment in mice [49] and pigs [50]. The activity of HDAC1/2 in embryos is required for proper DNA methylation, cell lineage development, and transformation from morula to blastocyst [51]. HDAC3 is closely connected with HDAC 11. Its inhibition increased the acetylation level of α-tubulin [45,52], significantly impaired the course of meiosis in mouse oocytes, and disrupted kinetochore–microtubule attachment and spindle assembly checkpoint [52], which resulted in abnormal spindle organization and chromosome misalignment.
The regulation of histone acetylation in oocyte meiosis is also reliant upon specific NAD+-dependent HDACs, silent information regulator 2 (Sir2) proteins (“sirtuins”) that belong to a seven-member family of deacetylases involved in the deacetylation of histones as well as nonhistone proteins. Moreover, sirtuins are employed in the regulation of metabolism, inflammation, and oxidative stress. [53]. SIRT1 plays a role in the activation of primordial follicles in a deacetylase-independent manner [54] and was reported to slow down the aging-related decrease of oocyte quality, which may under in vitro laboratory conditions impact oocyte maturation [55]. SIRT1 overexpression decreased H3 histone methylation and acetylation in post-ovulatory aged mouse oocytes as well as decreased aging related reactive oxygen species (ROS), spindle abnormalities, and mitochondrial dysfunction [56].
The SIRT2 impaired gap junctional communication during in vitro maturation of bovine oocytes by phosphorylation of connexin-43 [57] is another sirtuin activity. In human serum samples from IVF patients, a basal SIRT2 level was proposed as a pregnancy outcome predictor in combination with age, anti-Mullerian hormone (AMH), and antral follicle count (AFC) [58].
At the GV stage of mouse oocytes, SIRT7 is located within the nucleus, then upon meiotic resumption it is dispersed in the cytoplasm, with the highest SIRT7 concentration occurring around the chromosomes. SIRT7 knockdown compromised mitochondria function, significantly decreased ATP levels, and increased ROS [59]. SIRT4 exhibited similar phenotype to SIRT7 [60]. On the other hand, SIRT6 interacts with chromatin proteins and is employed in DNA double-strand break (DSB) repair mechanisms. Its knockdown in early mouse embryos shortened telomeres and caused an increase in DNA damage [61].
Sirtuins are also implicated in the regulation of energy metabolism and stress resistance; particularly, SIRT3, SIRT4, and SIRT5 mainly localize in mitochondria [62]. SIRT1 and SIRT3 have been revealed to play a crucial role in ensuring protection against oxidative stress in oocytes, granulosa cells, and early embryos [63]. It has been reported that the SIRT1 anti-oxidative stress effect in mouse oocytes is attenuated during aging [64]. Recently, a decrease of ovarian reserve in mice was linked to SIRT1-related changes in mitochondrial oxidative phosphorylation [65]. Additionally, a protective role of SIRT3 against oxidative stress was revealed in preimplantation mouse embryos [66]. A correlation between the decreased expression of SIRT3 and lower embryonic developmental competence was found in human in vitro cultured embryos [67].
2.3. Fertilization & Mitochondria
Mitochondria play a major role in providing each cell with energy by generating adenosine triphosphate (ATP) through electron transport-linked oxidative phosphorylation (OXPHOS) [68]. Since the aerobic respiratory pathway in eukaryotic cells is the only system that fully relies on mitochondria function [69], any mutations in mtDNA or nuclear-encoded mitochondrial genes can result in mitochondrial dysfunction that induces a variety of pathologies and contributes to an abnormal aging process [70,71]. Mitochondria are the prominent source energy for successful oocyte and sperm biogenesis and function. This dependence is due to the high energy demand for the support of proper chromosome segregation and the fertilization process [72,73]. In mammalian oocytes, a sufficient mtDNA copy number is essential to promote fertilization and early embryo development. Human oocytes with fewer than 100,000 copies of mtDNA evince a significantly lower fertilization rate than oocytes with more than 150,000 copies [74,75]. In vertebrates, inheritance of mitochondria is maternal as the paternal mitochondria of sperm origin are eliminated during early embryo development [76,77].
Mitochondrial activity is an indicator of oocyte developmental competence [78]. Oocyte maturation and early embryo development depend on ATP derived mainly from mitochondrial oxidative phosphorylation [79,80]. Events such as the formation and maintenance of the meiotic spindle are also dependent on mitochondrial function [81]. Deficiency of ATP and low mtDNA copy number are associated with poor oocyte quality, retarded embryo development, aneuploidy, and decreased implantation and placentation rates [82,83]. mtDNA has a considerably higher mutation rate than the nuclear genome, and it is assumed that mtDNA is prone to oxidative damage induced by reactive oxygen species [84]. Mitochondrial dysfunction and deficiency of mitochondria-derived ATP provoked by oxidative stress induces spindle disruption in MII mouse oocytes [85]. Mutations of mtDNA can cause a set of physical and cognitive disabilities, including pathologies of the nervous and muscular systems. However, progress has been made recently in the field of inherited mitochondrial disease and therapeutic approaches, such as the development of pre-implantation genetic screening and mitochondrial replacement therapy [86,87].
A decrease of mitochondrial number, function, and mtDNA quantity affect the viability of oocytes and female fertility [88,89]. Advanced maternal age is associated with a reduction of ATP production that leads to decreased metabolic activity and can negatively affect cell cycle regulation, meiotic spindle formation, chromosome segregation, fertilization, embryo development, and implantation [72,90,91]. An increased expression of the mitochondrial unfolded protein response gene Hspd1 in GV oocytes of PMSG-treated aged mice reflects the mitochondrial stress caused by advanced age [88]. In ovaries of aged mice, a decreased mRNA expression of mitochondrial antioxidants Prdx3 and Txn2 was reported [92]. An increased expression of the mitochondrial antioxidant TXN2 gene and mitochondrial transcription factor TFAM gene in cumulus cells of unstimulated aged cattle was detected [93]. To enhance the fertilization rate of aged oocytes, the technique of mitochondrial supplementation can be applied. For this purpose, it is possible to use the method of either partial or total cytoplasm transfer from donor to recipient oocyte [94,95].
2.4. Genomic Imprinting in Early Embryo
Genomic imprinting is defined as a monoallelic parent-of-origin-dependent gene expression in offspring and consists in differential methylation inherited from the gametes when one parental copy of the gene is silenced while the other parental allele is expressed [96]. DNA methylation is one of the epigenetic changes regulating the expression of imprinted genes during early development. It is an epigenetic process that involves DNA methylation and histone methylation without altering the genetic sequence. These epigenetic marks are established (“imprinted”) in the germline (sperm or egg cells) of the parents and are maintained through mitotic cell divisions in the somatic cells of an organism [14]. Although extensive nuclear reprogramming occurs in preimplantation embryos, the methylation of imprints acquired through gametogenesis escapes from this global epigenetic reprogramming and persists in preimplantation embryos (Figure 2) [9,97]. The methylation of the imprinted genes is thus preserved and then transmitted to the offspring. As of 2019, around 260 imprinted genes have been identified in mice and 230 in humans [98]. Imprinted genes are involved in the regulation of embryonic growth, placental function, postnatal growth, and neurobehavioral processes [99,100]. In humans, abnormal expression of some imprinted genes has been related to numerous diseases, developmental abnormalities, and malignant tumours [101,102,103]. Although DNA methylation is a key player in genomic imprinting through the establishment of imprinted marks on either paternal or maternal alleles (Figure 2), the genomic imprinting process is significantly influenced also by histone modifications and non-coding RNA [104,105,106].
2.5. Embryonic Genome Activation
During early embryogenesis between fertilization and implantation, parental DNA is subjected to rapid and extensive demethylation, and consequently epigenetic information inherited from the gametes is vastly reset in the preimplantation embryos [9,107]. In human embryos, a sharp decrease of paternal DNA methylation occurs between fertilization and the two-cell stage; however, the decrease of maternal DNA methylation is milder (Figure 2) [108]. The DNA methylation level is also decreased during zygotic activation in mice, bovine, and goat preimplantation embryos [109,110,111]. In mice, the most intense demethylation occurs in the zygotes, and gradual demethylation continues until the blastocyst stage [112]. The newly activated DNA demethylation that occurs during the early pronuclear stage precedes the increase of DNA replication indicating that DNA demethylation in the early zygote is independent of DNA replication [113].
The early human embryo consists of a large number of transposable elements (TE) that could be a potential cause of gene rearrangements, mutations, deletions, or duplications. Therefore, as a precaution, silencing of these evolutionary younger TE is ensured by DNA methylation or histone modifications [108].
Nearly all methyl groups are removed from the paternal-origin DNA immediately after fertilization [97]. Methylation of maternal-origin DNA is diluted with each round of replication and results in a substantial decrease of DNA methylation during the morula stage [114]. Global epigenetic reprogramming occurs in the early embryo when DNA demethylation is at the highest levels in the early blastocyst stage [115]. The genome-wide erasure of CpG methylation is more profound in early embryos from superovulated mice when compared to embryos from naturally mating control mice [116].
2.6. Intrauterine Epigenetic Inputs
Subsequent reinitiation of DNA methylation occurs in the blastocyst stage in the cells of the inner cell mass and establishment of new methylation marks continues during fetal development [97]. During post-implantation development the activity of DNMT3A and DNMT3B together with their coactivator DNMT3L are essential to establish the characteristic methylation profile in the developing embryo [117]. DNA methylation provides an epigenetic regulatory mechanism protecting the differentiating cells from regression to the undifferentiated state [115]. In post-implantation embryos, DNA methylation is an integral of epigenetic marks in the majority of embryonic tissues and persists in somatic tissues during the lifespan of adults [118]. Mice embryos with insufficient DNA methylation activity die at mid-gestation as a consequence of genome-wide demethylation [119].
3. Epigenetics within Translational Regulation: The Oocyte to Embryo Story
3.1. Active Transcription Fuels Maturing Oocyte
Primordial germ cells (PGCs) that are produced from female germ cells undergo mitosis, forming oogonia and during subsequent oogenesis, the oogonia become primary oocytes. Every oocyte originates from primordial germ cells (PGCs). PGCs migrate in utero into a future ovary and go through repeated mitotic cycles to form nests of germ cell syncytia [120]. All female mouse primordial germ cells are connected by intercellular bridges in the ovaries at embryonic day 11.5 to 17.5 and form synchronously dividing interconnected cysts or syncytia of up to 30 germ cells [121,122]. Following the homologous recombination and formation of cytoplasmic bridges, oocyte nuclei are arrested at the diplotene stage of meiotic prophase I. Following birth, germ cell nests are dispersed along with the invasion of pre-granulosa cells [121]. Individual primordial follicles made of primary oocytes enclosed by a basal layer of flattened granulosa cells are formed. However, a substantial amount of primordial follicles undergo atresia, and one of the proposed functions for follicular atresia is the selection of follicles containing oocytes of the highest developmental potential [123,124]. Activation of the primordial follicle that starts at prepubertal stage and extends throughout the reproductive life is cross-regulated by key transcriptional factors (FIGLA, LHX8, and SOHLH1) that cooperate on common downstream pathways in folliculogenesis [125]. If a primordial follicle is activated, it does so via the binding of Kit Ligand from granulosa cells (KL) onto a Kit receptor present on the oocyte and theca cells [126]. This Kit-KL system is connected downstream by a PI3K/Akt pathway [127]. A feedback loop is further secured by oocyte-secreted factors (OSFs) such as BMP-15 or GDF-9 [128,129]. As the oocyte grows further, macromolecules, proteins, and transcripts are rapidly accumulating within. Experiments on bovine oocytes have shown that a substantial amount of mRNA transcripts are capable of transport from granulosa cells into the oocyte via gap-junctions. These connections between the oocyte and cumulus cells were subsequently named as transzonal projections (TZPs) [130,131]. Once the oocyte reaches the fully grown germinal vesicle stage (GV), the transcription in the GV nucleus is ceased. Accumulated mRNAs are more stable than those present in somatic cells [132]. Further, during the MI/MII transition, the oocyte is dependent on the effective utilization of stored transcripts and proteins [133,134], which needs to be tightly regulated based on the metabolic requirement, nutrient availability, and presence of environmental stress. Fine tuning of oocyte meiosis and early embryo development is ensured by translational regulation.
3.2. Translational Regulation, the Key for Oocyte Success
The regulation of translation is orchestrated by many mechanisms, ranging from modulating polyA tail length, modifying mRNA post-transcriptionally, regulating interactions between proteins, degrading stored RNAs, clustering RNAs into ribonucleoproteins (RNPs), up to the 5′-terminal oligopyrimidine (TOP) motif mRNA regulation under stress from nutrient or oxygen deprivation [135]. The best known regulation mechanism of selective mRNA translation is the 3′ UTR polyadenylation, which works for about 70% of the oocyte’s mRNAs [136]. PolyA tail length positively correlates with ribosome occupancy. The ribosome loading itself is regulated by the CPE binding protein 1 (CPEB1) and deleted-in azoospermia such as (DAZL) binding to the cytoplasmic polyadenylation element (CPE) on the 3′ UTR of mRNA. Their absence compromises mouse oocyte meiotic maturation and MII oocyte development by dysregulating effector proteins, polyA binding protein 1 (PABP1), and polyA specific ribonuclease (PARN) responsible for mRNA polyadenylation and deadenylation respectively [137].
As oocyte meiosis is progressing, global translation decreases (Figure 2). However, translation of m7G capped mRNAs is mainly regulated by the mTOR/S6K1/4E-BP1 signalling (mTORC1 pathway) [2,138]. The main player here, the mammalian target of rapamycin (mTOR), a serine/threonine protein kinase, regulates diverse cellular functions (reviewed in [139]). It has been documented that mTOR activation and the protein synthesis initiation is influenced by the activity of cyclin-dependent kinase 1 [140]. mTOR is responsible for translational regulation of capped mRNAs containing a TOP motif by recruiting an eukaryotic initiation factor 4F (eIF4F) [2] and RNA-binding protein LARP1 [135]. Oocyte-specific conditional knockout of mTOR severely affected folliculogenesis. Deleting mTOR in meiotic maturation changed the oocyte’s proteome composition, caused spindle instability, aneuploidy, and failure to form a methaphase II equatorial plane [141].
Translatability of mRNAs is also indirectly regulated by the formation of superstructures with proteins into RNP cytosolic granules. In such a way, translation, degradation, and storage of transcripts can be controlled simply by regulating their physical availability to the translational or degradational machinery. RNP granules commonly found in oocytes and embryos are stress granules or p-bodies. Stress granules are formed upon nutritional deprivation, heat, or oxidative stress by liquid-liquid phase separation [142]. P- bodies, on the other hand, are engaged in the storage of mRNAs with regulatory functions, previously thought to play a role in RNA decay [143]. Continuous degradation of stored RNA expressed during early stages of oogenesis occurring in meiosis and during early embryo development is preventing them from being inherited by simple eradication of redundant RNAs [144].
3.3. Epitranscriptomics—Translational Regulation by mRNA Methylation
Not only oocyte translational regulation is key for meiotic maturation, but also the recently discovered regulation by the epitranscriptomic m6A mRNA methylation [145,146]. Earlier studies in Xenopus oocytes found m6A methylation to inhibit mRNA recruitment for translation. Key cell cycle and translation-related transcripts in Xenopus were demethylated in order to become translated [145]. However, a study on porcine oocytes showed rapid accumulation of m6A methylated transcripts inside the ooplasm as meiotic maturation progressed. Inhibition of m6A methylation by cycloleucine, a specific inhibitor of adenosyl-transferase, impaired oocyte maturation and further development [147]. A role of m6A mRNA methylation during meiotic maturation and maternal to zygotic transition has been confirmed in the mouse model [146], and proper regulation of m6A mRNA methylation was shown to be crucial for both preimplantation [148] as well as in utero [149] embryo development.
Recent evidence stressed the importance of m6A mRNA methylation in the development of fully matured and developmentally competent oocytes and early embryos. Epitranscriptomic m6A RNA methylation is an ubiquitous and reversible process orchestrated by methyltransferases (“writers”), binding proteins (“readers”) and demethylases (“erasers”) [150,151].
Three major epitranscriptomic writers exist: METTL3, METTL14, and METTL3 adapter—WTAP [51,152]. The methyltransferase complex METTL3 was studied in detail on mouse oocytes. Transient knockdown of METTL3 by RNAi led to a substantial decrease of mRNA translation efficiency, low oocyte maturation rate, problems with maternal to zygotic transition [146], and inability to form blastocysts [148]. METTL3 was found to play a role in folliculogenesis, ovulation, maintenance of DNA integrity, and preimplantation development [153]. The same enzyme is engaged in the angiogenesis of atherosclerotic mouse model embryos by upregulating vascular-endothelial growth factor (VEGF) [154] through m6A [155]. In zebrafish embryos, METTL3 regulated PHLPP2/mTOR-AKT signalling [156]. The recently published data have shown METTL3 direct interaction with the known p53 transcription factor that enhanced p53 stability and together cooperatively modified p53 targeted RNAs by m6A upon DNA damage [154]; METTL14 was found to be needed for embryonic post-implantation epiblast formation [149]. Further, data have shown the importance of methyltransferase KIAA1429 for folliculogenesis and oocyte development, as the KIAA1429 conditional knockout mice produced severe defects in oocyte growth, alterations in OSFs expression, and an inability to undergo nuclear envelope breakdown (NEBD) [157].
Many proteins can act as m6A readers. The most known are YTH domain containing family proteins (YTHDF1,2,3) [158] and IGF2BP1 protein regulating JAK2/STAT3 signalling [159], as well as many others. YTHDF1 promotes active translation in HeLa cells by interconnecting m6A mRNA transcripts with translation initiation factors, ribosomes, or stress granules [18]. YTHDF3 was shown in HeLa cells to enhance YTHDF1 upregulation of translation as well as the promotion of RNA decay via YTHDF2 [160]. The YTHDF2 reader in mice is responsible for the maintenance of correct gene dosage by utilizing RNA degradation machinery. This is coupled with the activation of CNOT7 deadenylase and DCP1A, DCP2 decapping enzymes. However, the most recent study on HeLa concluded that all YTHDF1,2,3 readers have common core sites and act together on selective m6A mRNA degradation via CCR4-NOT deadenylation complex [158]. Female YTHDF2 double knockout mice are infertile and show cytokinesis defects in early zygotic development. Nevertheless, YTHDF2 double knockout oocytes are capable of ovulation and fertilization [161]. Additional enzymes do exist that relay m6A regulation onto common physiologically important pathways. For example, recently discovered m6A reader activity in Fragile-X mental retardation protein (FMRP) is important for the maternal RNA decay in Drosophilla embryos. FMRP used m6A tagged mRNA transcripts for their sequestering into FMRP granules. [162]. FMRP was also shown to create granules at the onset of meiosis in human fetal ovaries, suggesting its importance in the translational regulation of oocyte maturation [163]. This was supported by the detection of FMRP in all stages of mouse oocyte meiotic maturation and its rapid decline in two cell embryos [26]. Therefore, m6A methylation could both directly and indirectly regulate the translation of certain mRNAs.
The removal of m6A methylation marks from transcripts is done by two main demethylases (“erasers”), fat mass- and obesity-associated (FTO), and α-ketoglutarate-dependent dioxygenase alkB homolog 5 (ALKBH5) [164]. A decrease of FTO expression with age was shown to increase m6A methylation in aged mouse ovaries and human granulosa cells of elderly patients [162]. The same decrease of FTO expression followed by an increase in m6A methylation was observed in ovarian tissues from premature ovarian failure (POF) patients and POF model mice [165]. Therefore, proper regulation of m6A methylation is one of the factors to ensure follicular developmental competence. Decreased m6A methylation in placental tissues of patients suffering from recurrent miscarriage caused by the upregulation of the second eraser, ALKB5H demethylase, revealed that in such endometrium the trophoblast is unable to nidate [166].
4. Translation of Epigenetics into ART
Assisted reproduction techniques (ARTs) are widely applied in the field of human reproduction (Figure 3) and animal breeding. Exposure to ART results in a decreased developmental competence of fertilized mouse oocytes, partially due to the induction of epigenetic changes [167] (Table 1).
In humans, no significant epigenetic changes were found between regular pregnancy and ART pregnancy in newborns, however only a few key imprinted genes were analyzed in a small cohort of patients [190]. Recently, epigenetic imprinting-related disorders were demonstrated on mouse models and also observed in ART newborns like Prader–Willi syndrome (PWS), Silver–Russell syndrome (SRS), Beckwith–Wiedemann syndrome (BWS), and Angelman syndrome (AS) [191]. The long-term epigenetic effects of ART still await evaluation. Needless to say, more extensive follow-up of children born from ART embryos should be carried out. Moreover, further investigations into the epigenetic impact of ART methodology on cultured oocytes and embryos should be done.
4.1. Hormonal Stimulation
Superovulation or the clinical term controlled ovarian stimulation (COS) is applied to statistically increase the chances of acquiring blastocyst stage embryos compared to normal ovulation and increase the likelihood of a successful pregnancy. The basis of COS is the stimulation by a recombinant follicle stimulating hormone (rFSH) and the ovulation trigger, human chorionic gonadotropin (hCG). A co-administration with gonadotropin-releasing hormone (GnRH) agonist or antagonist is needed to avoid premature ovulation.
Increased incidence of chromosomal aneuploidy in human COS oocytes was associated with changes in DNA methylation [168]. COS was also linked to embryo development retardation [192] and negative effects on child health [193,194,195] A loss of genomic imprinting, particularly associated with the overgrowth Beckwith–Wiedemann (BWS) syndrome was reported in bovine fetal tissues. Demethylation of imprinted genes was observed in other bovine genome loci [196].
Additionally, protein translation was affected using the mechanisms triggered by superovulation. The mRNA expression of two critical players in translational regulation of stored maternal mRNAs, the embryonic poly(A)-binding protein (ePAB), and the poly(A)-binding protein cytoplasmic 1 (PABPC1), was modified in oocytes and two-cell embryos [170]. After IVM oocyte culture in the presence of rFSH, a decreased global translation was observed in mouse, bovine, and porcine models including humans [197]. rFSH may compromise regulation of specific translatome essential for oocyte maturation and early embryo development. Therefore, the use and dosage of recombinant hormones in the conventional IVM should be thoroughly evaluated.
Superovulation in mouse oocytes and early embryos alters DNA methylation [198] and expression of methyltransferases [169] and impairs methylation of genes involved in glucose metabolism, nervous system development, cell cycle, cell proliferation, and mRNA processing [172]. Disrupted DNA methylation of imprinted loci in mouse blastocysts affected, for example, a body-weight-limiting H19 gene [173], and was more frequent at higher hormonal superovulation dosages [171]. Superovulation caused imprinting defects leading to embryonic abnormalities and higher mortality [174,199]. Detected epigenetic differences in DNA methylation between superovulated and naturally ovulated oocytes suggested that superovulation also recruits growing oocytes with incomplete epigenetic maturation [172]
4.2. Oocyte In Vitro Maturation
When hormonal stimulation does not produce a satisfactory number of matured MII oocytes, in vitro maturation (IVM) is considered. IVM is based on the retrieval of fully grown cumulus enclosed GV oocyte complexes (COC) from ovarian follicles. COCs are meiotically matured by the IVM in the presence of rFSH and LH (rLH) to produce MII oocytes. The conventional IVM technology is understandably inferior to standard COS with maturation in vivo.
As just mentioned, rFSH promoted a decrease of global proteosynthesis marker, the radioactively labelled 35S methionine, in mouse, bovine, porcine, and human IVM oocytes [197]. The use of recombinant hormones in the IVM media is one of the factors affecting proper translational regulation and proteosynthesis during maturation.
A hypothesis was proposed that the resumption of meiosis upon COC ovarian puncture (OPU) is premature and could also have a negative impact on IVM quality. In response to that, an alternative approach called capacitation IVM (CAPA-IVM) was devised [200]. This experimental ART maintains high cAMP concentration within the in vitro cultured human COC by stimulating cumulus cells with rFSH, insulin, estradiol, and C-natriuretic peptide, thus indirectly inhibiting the NEBD in oocytes for about 24 h. The recent clinical study resulted in a live birth rate after the first embryo transfer of 35.2% for CAPA-IVM compared to 43.2% for standard IVF control [200]. It can be hypothesized that CAPA-IVM has a direct downstream impact on translational regulation as it gives the oocytes more time to equilibrate before meiotic maturation. These promising results from CAPA-IVM deserve further investigation, particularly the clarification of underlying molecular mechanisms and epigenetic regulation.
IVM quality is indeed determined by epigenetic m6A transcript methylation or histone deacetylations. Numerous histone modifications are employed in oocyte meiotic maturation, ranging from most profound deacetylation to methylation, phosphorylation, ubiquitination, and SUMOylation as reviewed by He et al., 2021 [41]. General histone deacetylation was compromised throughout meiosis in aged human oocytes. The residual acetylation correlated with chromosome misalignment linked to aneuploidy [181]. Moreover, human IVM oocytes with reduced HDAC1 expression also exhibited MII spindle abnormalities. Comparatively, more HDAC1 transcripts were present in in vivo (IVO) matured oocytes than IVM [201].
Improved IVM technology could also utilize the discussed sirtuin deacetylase family as recently reviewed [53]. Authors of the review detected sirtuins application for the management of aging and stress related syndromes, PCOS, diabetes, or endometriosis, and concluded the beneficial improvement of the energy balance and ROS protection. Decreased SIRT3 expression for example, correlated with lower developmental competence in human in vitro cultured embryos, which was attributed to defective mitochondrial biogenesis [67]. Recently, basal human serum SIRT2 level was suggested as a novel biomarker of ART outcome [202]. This kind of IVM therapy is promising, however further investigation is needed to determine their best delivery, dosage, combination, and mode of action.
The IVM approach is most applied for the management of polycystic ovarian syndrome (PCOS) which is the most common endocrine disorder in 6–20% of women of reproductive age accompanied by oligoovulation and/or anovulation. PCOS is a multifactorial syndrome with strong epigenetic inheritance, where external environmental, lifestyle, and dietary factors play a significant role [203].
PCOS infertility is managed through IVF centres. Recent evidence suggests intergenerational epigenetic inheritance of this syndrome [204]. The puzzling question arises whether PCOS is trans-generationally inherited through offspring, who would again become clients of IVF clinics (Figure 4). Instead, could PCOS be cured by managing mentioned external epigenetic factors rather than just relayed onto the next generation by IVF?
4.3. Mitochondrial Therapy
Oocyte developmental competence is tightly connected with energy availability. Meiotic maturation, fertilization, and embryonic genome activation are all energetically demanding processes depending on essential mitochondrial ATP production through OXPHOS. Metabolic disorders and aging are coupled with oocytes of inferior quality, which is largely attributable to energy production in the form of ATP by mitochondria as reviewed by [207]. mtDNA copy number in human oocytes range from 20,000–600,000 [79,208] and was found to decline with increasing maternal age [209]. The ART solution to this trend in our population is to apply cutting-edge mitochondrial therapies and take advantage of enucleated donor oocytes in techniques such as Cytoplasmic Transfer (CT), Maternal Spindle Transfer (MST), first Polar Body Transfer (PB1T), or in case of zygotes, Pronuclear Transfer (PNT). Nuclear transfer in GVs has been described, but due to complete zygote arrest of such maturated and fertilized mouse model oocytes, is not further considered for clinical applications (reviewed by [210]).
CT of about 10–15% of donor oocyte cytoplasm into one of the two sibling oocytes of an aged patient increased their developmental competence. Newborns from such treated oocytes were considered being healthy [95].
The MST method is based on the removal of the MII spindle from a healthy donor oocyte. In the same way, MII spindle is transferred from the patient’s affected oocyte into a healthy enucleated donor oocyte [211,212]. The first MST in human oocytes had low zygote survival ability, but with comparable blastocyst rates [211]. In 2017, MST was used for the first time therapeutically in the management of Leigh syndrome, which affects mtDNA and mitochondria [213]. Following a live birth, the load of mutated mtDNA was estimated to be below 10%. Concerns about MST introducing new epigenetic changes in donor oocytes were negligible as revealed by analysis of gene expression, which found no significant differences [214].
Whole genome RNA-sequencing of PNT derived blastocysts also found no major epigenetic differences [215]. An inferior method to MST, the PB1T method successfully reconstituted spindle in about 67% of enucleated donor oocytes but following fertilization only about half of the produced viable zygotes were capable of blastocyst development [216,217]. First polar bodies (PB1) were shown to mirror the methylomes of the oocytes they originated from, and therefore PB1s could have more of a diagnostic, rather than therapeutic, role [218].
Another non-invasive strategy to enhance oocyte and early embryo competence is to support mitochondrial functions in in vitro cultured oocytes by supplementing the culture media with compounds upregulating the functions of histone deacetylases sirtuins. SIRT1 for instance stimulates mitochondrial activity, enhances its biosynthesis and regulates degradation of mitochondria [219].
4.4. IVF & ICSI
New evidence from genome-wide sequencing of neonatal cord blood has shown that epigenomes of ART newborns exhibit the loss of CpG methylation compared to those of natural conception (NC). A total of 176 genes were differentially methylated including genes employed in growth and neurodevelopment [195]. Similar findings were obtained from 7–9 week old fetal human tissue after elective termination of pregnancy. A total of 164 differentially methylated genes were detected and associated with the development of the skeletal system, body size, lipid, and steroid synthesis [220]. Moreover, a study on histone modifications between IVF and ICSI found significant differences in global H3K4me3. ICSI placentas had lower H3K4me3 levels than IVF placentas in line with its lower transcription activity [221]. It is important to bear in mind that ART vs NC comparison will always accompany epigenetic changes acquired during the in vitro embryo culture so no clear epigenetic input of IVF and ICSI can be drawn.
4.5. Embryo In Vitro Culture
In the female reproductive tract, particularly the oviducts, developing early embryos are under the influence of hormones, nutrients, growth factors, and cytokines [222]. The epigenome of in vitro cultured mammalian embryos is vulnerable to exposed culture conditions as aberrant DNA demethylation kinetics was detected in in vitro grown embryos compared to embryos of in vivo origin [183].
During in vitro culture, early embryos are exposed to limited nutrient availability and furthermore, they are influenced by the end products of their metabolism [223]. A stress response can be induced through the manipulation of embryos by pipetting, exposure to thermal stress, and/or detrimental change in pH [224,225]. It is evident that embryo in vitro culture conditions contribute to its epigenetic status. Embryo exposure to suboptimal culture conditions or toxic substances in the medium can result in altered DNA methylation, genetic reprogramming, developmental disruption, and consequently early embryo loss as demonstrated in the mouse, rat, and rabbit [183,226].
In vivo early embryonic development takes place in the oviducts, but in vitro embryo culture conditions are far from the oviductal environment, respective to nutrients, oxygen concentration, or epigenetic messengers. One example for both, oviductal extracellular vesicles (oEV) were shown to harbour miRNA that can downregulate specific mRNAs or modify gene expression is other ways [185]. These oviduct–embryo interactions are missing in vitro and would most likely play much more important roles in the regulation of the embryonic epigenome that await their discovery [227,228,229,230].
Adequate oxygen supply is another crucial condition for the success of in vitro culture. Concentrations of 5% oxygen and 20% oxygen are commonly used during in vitro culture of oocytes and early embryos. DNA methylation and gene transcription of mouse oocytes grown in in vitro conditions under 20% oxygen correlated more with the reality of in vivo conditions, indicating that higher oxygen concentration is beneficial for mouse oocytes matured in vitro [231]. In contrast, exposure of in vitro cultured bovine preimplantation embryos to 20% oxygen was associated with an increase in global DNA methylation indicating that in this case oxidative stress can alter the embryonic epigenome [178,179]. These findings suggest variations in the optimal oxygen concentration among different species. In order to optimize the oxygen in in vitro culture of mammalian embryos, it was suggested to reduce the concentration up to the physiological oxygen tension as the median oxygen rate in the mammalian oviduct is around 8% [232,233]. The importance of oxygen tension reduction during in vitro embryo culture was confirmed by systematic review and meta-analysis of published human ART studies, revealing an increase in pregnancy and live birth rates of embryos cultured at 5% oxygen concentration [234].
At 20% oxygen concentration in bovine embryo blastocyst culture, elevated ROS were detected [178]. Although the ROS are important signalling molecules in certain biological processes and are normal products of oocyte metabolism, they can interact with biological molecules such as lipids, proteins, and nucleic acids, cause oxidative stress and cellular damage leading to the impairment of oocyte quality [235]. Oxidative stress can also increase the risk of aberrant DNA methylation in in vitro cultured preimplantation embryos [236,237].
In order to compensate for the adverse conditions of in vitro culture and maintain DNA methylation, a supply of methyl donors is needed, e.g., folates, which are often absent in culture media [237]. Methionine, an important intermediate metabolite of the one-carbon metabolism pathway (OCM) (Figure 5), contributes to epigenetic regulation by providing methyl groups for DNA methylation via S-adenosyl methionine (SAM) [238]. The methionine level is an important factor influencing the quality of the early embryo epigenome. Elevated concentrations of the methionine product homocysteine in the oocyte and embryo environment is, however, harmful due to its toxicity and has to be converted back to methionine. Limited remethylation of homocysteine to methionine leads to a decrease of SAM causing DNA hypomethylation [238].
A high level of homocysteine was also detected in the serum of PCOS patients and in the follicular fluid of polycystic ovaries and was linked with poor oocyte quality [242,243]. The aging process may be an indirect factor contributing to the decrease of embryo quality and oocyte maturation through increasing homocysteine levels in follicular fluid; hence, a decrease of homocysteine level in follicular fluid can significantly improve the oocyte maturation rate and embryo quality [244]. Elevated homocysteine levels in follicular fluid is also associated with hypermethylation of mitochondrial DNA accompanied with a mitochondrial malfunction in oocytes retrieved from porcine polycystic ovaries [245]. In porcine in vitro cultured oocytes, exposure to homocysteine significantly reduced the survival rate, polar body extrusion rate, and cleavage rate; however, DNA methyltransferase inhibitor 5-AZA rescued the homocysteine-induced mitochondrial dysfunction and improved the quality of oocytes and developmental competence [245]
In vitro oxidative stress can also affect mitochondrial function. Treatment of in vitro cultured bovine oocytes with palmitic acid induced upregulation of peroxiredoxin 3, which is a mitochondrion-specific H2O2-scavenging enzyme, and elevation of the mitochondrial HADHB, UQCRB, and cytochrome C proteins suggesting that oxidative stress increased electron transport in bovine oocytes [180]. Exposure of MII mouse oocytes to H2O2 led to a decrease in mitochondria-derived ATP and disassembly of meiotic spindles [85].
4.6. Cryopreservation
Two main methods of germ cell cryopreservation exist: slow freezing and vitrification, both with the goal to eliminate ice crystal formation inside the cells. The slow freezing method is based on the continuous and steady decrease of temperature by 1–2 °C/min in the presence of cryoprotective agents and is conventionally applied for the preservation of human fertilized zygotes [246]. The second approach, called vitrification, is now the most widely used method for cryopreservation. It combines a rapid increase in media viscosity and solute concentration with snap freezing in liquid nitrogen. Vitrification has outperformed slow freezing in conventional ART human embryo cryopreservation by both higher clinical pregnancy and live births [247].
However, from the epigenetics perspective cryopreservation is still questionable [186]. Vitrifcation significantly reduced the ATP content in oocytes of various mammalian species, including humans [248,249]. Particularly changes in global DNA methylation, histone modifications and genetic imprinting are to be scrutinized. Research performed on vitrified mouse embryos found a common pattern of decreased global [250] and imprinted DNA [251] methylation. Vitrification of bovine oocytes was associated with a significant reduction in the expression profile of three epigenetic-related genes DNMT1, DNMT3B, and HDAC1 [252]. In vitrified porcine embryos, a greatly reduced expression of epigenetically associated key genes SMYD3, TET2, and HDAC8 led to altered epigenetic reprogramming and decreased blastocyst rates [253]. The cryoprotective agents present in IVF vitrification media might negatively affect the epigenetic profile in embryos, as dimethylsulfoxide (DMSO) was found to be responsible for the disruption of global DNA methylation and significant decrease of ATP content in vitrified human cardiac tissue [254]. A detailed analysis of molecular changes occurring in cryopreserved germ cells and embryos is necessary to distinguish possible molecular targets that could contribute to improve the cryopreservation procedures.
5. In Utero Epigenetics, beyond ART?
Exposure to unfavorable conditions before pregnancy and during intrauterine development lead inevitably to epigenetic alterations of a newborn and can evolve into pathogenesis of metabolic, cardiovascular, endocrine, and malignant disorders in adulthood [255]. As every ART-produced embryo has to be eventually planted into a mother’s womb, these in vivo epigenetic factors deserve a closer look in order to understand the full story.
5.1. Endometrial Receptivity & Placentation
Following ART embryo transfer, each embryo faces the selective process of nidation and implantation into the endometrial tissue of the uterine wall. This in utero process itself is highly complex and involves the cooperation of many signalling pathways. Many epigenetical mechanisms are involved as well, such as DNA methylation [256], m6A methylation [166], or interaction with ncRNAs secreted from exosomes [257]. It has even been suggested that DNA methylation profiles of cervical secretion could serve in the future as an alternative way for diagnosing endometrial receptivity [258]. Decidualization, the process of endometrial preparation for blastocyst implantation, is suppressed in human endometrial stromal cells by an ncRNA, the miR-542-3p. Overexpression of this miRNA also downregulates VEGF, cyclooxygenase-2 (COX-2), and matrix metalloproteinase (MMP-9), all linked with angiogenesis [259]. Most ncRNAs are able to exert their effects by transportation as cargos in lipophilic exosomes. In this way, ncRNAs influence the embryonic development in the oviduct and uterus [257]. So far, little is known about the scope of the influence of these ncRNAs on the epigenome of a developing individual.
Following successful implantation, the embryo turns to cardiac and neural development. These processes are energy-dependent and highly susceptible to proper dietary intake and environmental conditions. Defective placentation is associated with impaired mitochondrial function and associated low ATP production [82,83]. Incorrect DNA methylation on imprinted genomic regions dysregulates placental function [99,100]. Nutritional and environmental status can influence the uterine and fetal epigenome to a large extent, and therefore their effects have to be taken seriously into account.
5.2. Nutritional Epigenetics
Numerous studies describe the effects of nutrition on the epigenome during embryonic development (reviewed in [238,260,261]). Particularly, the nutritional source of dietary methyl donors in early development can influence the DNA methylation process [239]. Methyl groups or so called one-carbon groups are produced through OCM, which integrates folate and methionine cycles and is a source for epigenetic DNA methylations, biosynthesis of DNA, proteins, and lipids. Amino acids such as methionine, glycine, and serine and appropriate levels of especially B class vitamins (B2, B6, B12) and folic acid (B9) are integral inputs for the successful functioning of one-carbon metabolism (Figure 4). Epigenetic changes in humans associated with OCM modifications affect pathways related to growth, metabolic functions, neural development, and stress response [239].
The modulation of epigenetic modifications, including DNA methylation, is done by the mTOR signalling through the OCM [262]. mTOR signalling is able to affect OCM by increasing the de novo synthesis of serine, one of the major single carbon donors [263]. Elevation of placental mTOR observed in obese women increased birth weight [264]. As one of the main functions of OCM is to produce S-adenosylmethionine (SAM) to ensure methyl-group transfer reactions, mTOR signalling, by influencing the OCM, is able to influence epigenetic modifications, including DNA methylation [262]. The epigenetic regulator methyltransferase DNMT1 is one of the downstream targets of the mTOR pathway [265]. The experimental inhibition of mTOR induced the suppression of DNA methyltransferase DNMT1 [260].
Microelements in the diet, e.g., Cu, Mn, Se, and Zn are often required for proper enzymatic function, neutralizing ROS, and are also involved in epigenetic regulation. For example, zinc is implicated in the correct functioning of methyltransferases and methyl-binding proteins and its deficiency has been suggested to affect the activities of zinc-dependent epigenetic enzymes, which are essential for DNA methylation [261]. Prolonged dietary Se supplementation in rats affected global and specific DNA methylation in liver and colon tissue [266]. Mn tends to accumulate in the placenta, and its supplementation experiments in chick embryos counteracted hyperthermic stress effects by modulating DNA methylation and histone acetylation [267]. A recent US study found a similar mode of action in Cu metabolism, which may be employed in DNA methylation and the regulation of human placentation [268].
Malnutrition can epigenetically induce in utero obesity in offspring, which usually manifests in adulthood. Blood analysis of human adults revealed that periconceptional exposure to famine altered the DNA methylation of genes implicated in growth and metabolic regulation. Prenatal famine exposure resulted in changes of DNA methylation patterns in genes associated with cell growth, metabolic health, mitochondrial function, adipogenesis, and its deposition [269,270]. A preovulatory protein restriction diet in rats induced abnormal mitochondrial ultrastructure in oocytes and negatively affected gene expression related to mitochondrial biogenesis [271].
This demonstrates epigenome vulnerability by famine in the early stages of development. There is strong evidence that maternal nutrition influences the development and future health of offspring. Studies done on mice [272] and cattle [273] confirmed maternal diet effects on oocyte DNA methylation. Postpartum cows exposed to negative energy balance and metabolic stress had a number of maternally imprinted genes in their oocytes hypomethylated [274].
Maternal obesity is another risk factor affecting offspring health epigenetically and has been shown when together with excessive nutrition intake to have a positive correlation with offspring obesity [275]. Causes of fetal overgrowth have been explored on the mouse model. It was documented that obesity in pregnancy is linked to stimulation of placental insulin/IGF-1/mTOR and leptin signalling pathways [163]. The obesity mouse model resembles the changes in placental mTOR signalling and amino acid transporters activity observed in obese women giving birth to large babies [276]. DNA hypermethylation in the placentas of obese pregnant women was associated with reduced expression of the ten-eleven translocation (TET) methylcytosine dioxygenases, enzymes involved in DNA demethylation [58]. Increased gestational weight in early pregnancy is related to the enhanced CpG methylation of MMP7, KCNK4, TRPM5, and NFKB1 genes in offspring cord blood DNA [277].
Maternal obesity and overnutrition affect mitochondria function and induce epigenetic changes of mtDNA. The mtDNA copy number, elevated expression of nuclear genes encoding mtDNA transcription factors Tfam and Nrf1 were detected in oocytes of obese mice [278]. A maternal obesogenic diet (high fat/high sugar) was associated with elevated mtDNA content and increased expression of mtDNA biogenesis regulators Tfam and Pgc-1α, enhanced mitochondrial antioxidant defence, increased lipoxygenase expression, enhanced expression of transcriptional regulator NF-κB, and depletion of ovarian follicular reserve in young adult female mouse offspring [279]. The analysis of the newborn umbilical cord indicated that promoter methylation of the mitochondrial biogenesis regulator PPARGC1A in babies was positively correlated with maternal BMI [280]. It is evident that maternal obesity may affect the offspring metabolism through epigenetic regulation of specific genes.
It is common knowledge that consuming alcohol in pregnancy affects embryo development and can induce a variety of birth defects and neuronal disorders in offspring [281]. Alcohol intake interferes with normal folate metabolism (Figure 5) and decreases folate bioavailability for methyl donors by inhibiting methionine synthase and methionine adenosyl transferase [282].
It has been shown that alcohol metabolites, such as acetaldehyde, modify DNA methylation by inhibiting DNA methyltransferases [283].
The effect of alcohol abuse on the methylation of specific genes resulted in alterations of gene expression and neural development as reported in numerous studies (reviewed in [284]). In in vitro cultured fetal mouse neurons, the alcohol exposure induced a decrease of DNA methylation detected in the vicinity of the NMDA receptor subunit NR2B gene, which plays an important role in neural development and in learning and memory [285,286].
In human oocytes, alcohol-associated epigenetic changes were detected already in the growth phase when genomic imprints are established and could possibly affect the health of the child [287].The in vivo exposure of mice embryos to ethanol resulted in retardation of embryo development and was accompanied by epigenetic alteration of the H19/Igf2 methylation in the placenta; the paternal allele of H19/Igf2 was less methylated while the methylation of the maternal allele was elevated [288]. In mouse embryos exposed to alcohol, in vitro changes in methylation on chromosomes 7, 10, and X related to neural tube defects were detected [289].
6. Fetal Epigenetics Dependence on Maternal Lifestyle and Environment
In the present day, people live in a highly stressful world in compromised living environments. As we have already mentioned, both undernutrition and overnutrition can impact one’s epigenome irrespective to ART. Here, we emphasize that postponing parenthood or living in physically and psychologically toxic environments can induce changes to the new generation’s epigenome (Table 2).
6.1. Lifestyle
Advanced maternal age negatively influences oocyte maturation, meiotic divisions, and embryonic development [306,307]. Increasing maternal age raises the chances of miscarriage and adverse health issues in offspring, mainly due to chromosomal aneuploidies such as Down’s syndrome [308]. Age-related decrease of ooplasm quality, mitochondrial defects, and abnormalities in meiotic maturation mechanisms are possible causes of the advanced maternal age-related decline of oocyte competence [168,309,310] A higher incidence of chromosomal abnormalities was reported in mammalian oocytes acquired from aged females (reviewed in [311]), and an elevated occurrence of aneuploidy related diseases was observed in babies born to mothers over 35 years of age [309]. The meiosis-specific cohesin subunits, REC8 and SMC1B, were found to be decreased in oocytes of women aged 40 and over, suggesting that age-related decrease of meiotic cohesin subunits impair sister chromatid cohesion and results in increased segregation errors [295]. The effect of maternal age on oocyte quality and associated epigenetic changes have been well-documented and extensively reviewed in humans [310].
Oocytes of older women (41–44 years old) were more prone to oxidative damage by the attenuated expression of cytochrome c oxidases (COX gene family) involved in oxidative phosphorylation and energy production and down-regulation of members of the peroxiredoxin gene family [296]. Similarly, gene expression analysis of ovaries in aged mice revealed a decrease in mRNA expression of mitochondrial antioxidant genes, peroxiredoxin 3 (Prdx3), and thioredoxin 2 (Txn2) [92]. Gene transcriptome analysis of human oocytes retrieved from patients older than 40 years revealed a decreased expression of spindle checkpoint genes, DNA damage checkpoint-related genes, ADP ribosylation factors involved in protein trafficking, and mRNA coding for the EIF4ENIF1 protein, which mediates the nuclear import of eIF4E [294]. Closer analysis of MII aged mouse oocytes gene expression (42–45 weeks old) revealed a downregulation of genes involved in mitochondrial functions, antiapoptotic mechanisms, and those involved in the ubiquitin-proteasome degradation pathway. Moreover, expression was reduced in transcripts related to microtubule cytoskeleton, chromosome segregation, and maintenance of the DNA methyltransferases Dnmt1o and Dnmt1s [298]. Hence, reduced DNA methylation in MII oocytes and early embryos of aged mice together with a low abundance of DNA methyltransferases clearly points to a lower reproductive potential [297,312]. More recently, it was suggested that decreased DNA methylation related to advanced maternal age may partially induce significant changes to gene expression and alter developmental fitness (reviewed in [167,313]).
Maternal smoking in pregnancy remains a serious issue that gravely affects child health. Fetal exposure to maternal smoking during pregnancy induces changes in DNA methylation of different tissues. The impact of prenatal exposure to tobacco smoke on DNA methylation was mostly analyzed in the cord blood and placenta of newborns—reviewed in [314]. It was found that DNA methylation patterns associated with smoking are relayed to a low birthweight [315] and schizophrenia induction in adulthood [316]. Moreover, the meta-analysis mapping association between maternal smoking in pregnancy and newborn blood DNA methylation revealed that smoking whilst pregnant causes changes in the CpGs methylation of numerous genes including those involved in teeth and neurologic development as well as cancer induction [293]. AHRR and CYP1A1, genes of aryl hydrocarbon receptor signalling, which is engaged in detoxification, were also found to be differentially methylated in the cord blood of newborns exposed to maternal smoking [317].
Lifestyle can also be a factor determining the occurrence of gestational diabetes mellitus (GDM) in women. GDM is associated with an increased risk of cardiometabolic diseases and diabetes in the offspring [301]. DNA collected from venous blood of GDM women offspring detected differentially methylated CpGs in genes associated with type 2 diabetes, diabetic nephropathy, obesity, and coronary heart disease [301]. It has been shown that pregestational hyperglycemia renders the offspring more vulnerable to glucose intolerance. The expression of TET3 dioxygenase, responsible for 5-methylcytosine oxidation and DNA demethylation in the zygote, is decreased in oocytes from a mouse model of hyperglycaemia (HG mice) and in people with diabetes [318].
6.2. Environment
Endocrine Disrupting Chemicals (EDCs) cause serious defects in human health. EDCs are chemicals of natural or man-made origin that interfere with the endocrine system. Humans are exposed to EDCs from many sources, including diet, thermal receipt papers, cosmetics, cleaning products, pesticides etc. [319]. Exposure to EDCs during development can induce permanent alterations of physiology and increase predisposition to health issues such as obesity, asthma, and cancer [320].
Bisphenol A (BPA) is a ubiquitous plasticizer, EDC with probable estrogen-like activity. Newborn cord blood DNA studies revealed that prenatal exposure to BPA (but not bisphenol F and bisphenol S) induces hypomethylation of gene promoters related to adipogenesis, growth and metabolism [321]. Hypomethylation of the obesity-associated mesoderm-specific-transcript (MEST) gene promoter enhanced MEST expression, which resulted in a significant increase of body mass index (BMI) in children [303]. Exposure to BPA during the late stages of oocyte development and the early stages of embryonic development disrupted the expression of imprinted genes in mouse embryos and placentas [304]. Mouse BPA exposure at 9–16 day of pregnancy led to decreased methylation and enhanced expression of the homeobox gene Hoxa10 in offspring, a key regulator of in utero organ development [322]. Additionally, polystyrene nanoparticles inhibit meiotic maturation by negatively affecting spindle assembly and chromosome alignment in mice oocytes; moreover, exposure to polystyrene nanoparticles increased oxidative stress and mitochondrial aggregation during meiotic maturation [305]
Stress is a widespread environmental factor affecting human reproduction. Preconception and pregnancy time exposed to stress is associated with developmental problems and new-born physical and psychological health. Maternal stress has been linked to infant mortality, premature weight and low birth weight [323]. Increased DNA methylation of the glucocorticoid receptor NR3C1 gene promoter, which is related to maternal stress via controlling hypothalamic-pituitary-adrenal axis (HPA) has been reported in human cord and new-born blood [324]. Chronic maternal distress in pregnancy was accompanied by altered CpG methylation on glucocorticoid pathway genes in human placentas, which suggests that the placenta can be the main mediator between maternal and fetal stress. [299]. Similarly, differential DNA methylation of the HPA axis genes CRH and NR3C1 was detected in cord blood of new-borns and CRH, CRHBP, NR3C1, and FKBP5 in their placentas [325].
7. Conclusions
Over the past years, an increasing research interest has been focussed on understanding the regulation of the animal and human epigenomes. Many publications have demonstrated that DNA and RNA methylation, histone modifications, and non-coding RNA regulation are integral to the normal embryo development and future health of a newborn. However, our current understanding of these mechanisms is unsatisfactory. Here, we endeavoured to emphasize the connection between assisted reproduction technology and its epigenetic implications for oocyte and embryo development. We did not want to overlook the fact that following an embryo transfer, the in utero development is epigenetically affected by the actual fitness and age of the reproductive system as well as by external stimuli such as diet or nutrition. We want to show a complex picture of what is behind ART live-birth rates and the resulting health implications from the epigenetic point of view.
ART is of enormous importance to infertile couples and our society in general. Therefore, it is much needed to focus on improving ART and minimizing unnecessary negative impacts by discussing and bringing in the latest knowledge of epigenetic mechanisms involved in clinical infertility treatments. Many of the questions regarding epigenetics influencing in vitro oocyte and embryo culture protocols remain unanswered. Future studies should search for epigenetic key points in the concerned developmental pathways and investigate their clinical relevance as biomarkers or new treatments.
Author Contributions
Writing—original draft preparation, M.D.; writing and editing, L.N.; conceptualization, writing—review and editing, J.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the grant CZ.02.1.01/0.0/0.0/15_003/0000460 from the Operational Programme Research, Development and Education, by Institutional Research Concept RVO67985904, by GACR (22-27301S) and by GAUK 373621 for M.D.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ivanova, E.; Canovas, S.; Garcia-Martínez, S.; Romar, R.; Lopes, J.S.; Rizos, D.; Sanchez-Calabuig, M.J.; Krueger, F.; Andrews, S.; Perez-Sanz, F.; et al. DNA methylation changes during preimplantation development reveal inter-species differences and reprogramming events at imprinted genes. Clin. Epigenet. 2020, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Susor, A.; Jansova, D.; Cerna, R.; Danylevska, A.; Anger, M.; Toralova, T.; Malik, R.; Supolikova, J.; Cook, M.S.; Oh, J.S.; et al. Temporal and spatial regulation of translation in the mammalian oocyte via the mTOR–eIF4F pathway. Nat. Commun. 2015, 6, 6078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asami, M.; Lam, B.Y.H.; Ma, M.K.; Rainbow, K.; Braun, S.; VerMilyea, M.D.; Yeo, G.S.H.; Perry, A.C.F. Human embryonic genome activation initiates at the one-cell stage. Cell Stem Cell 2022, 29, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.; Taggart, M.; Frommer, M.; Miller, O.J.; Macleod, D. A fraction of the mouse genome that is derived from islands of nonmethylated, CpG-rich DNA. Cell 1985, 40, 91–99. [Google Scholar] [CrossRef]
- Illingworth, R.S.; Gruenewald-Schneider, U.; Webb, S.; Kerr, A.R.W.; James, K.D.; Turner, D.J.; Smith, C.; Harrison, D.J.; Andrews, R.; Bird, A.P. Orphan CpG Islands Identify numerous conserved promoters in the mammalian genome. PLoS Genet. 2010, 6, e1001134. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, M.; Wang, R.Y.H. 5-Methylcytosine in eukaryotic DNA. Science 1981, 212, 1350–1357. [Google Scholar] [CrossRef]
- Friso, S.; Choi, S.-W.; Girelli, D.; Mason, J.B.; Dolnikowski, G.G.; Bagley, P.J.; Olivieri, O.; Jacques, P.F.; Rosenberg, I.H.; Corrocher, R.; et al. A common mutation in the 5,10-methylenetetrahydrofolate reductase gene affects genomic DNA methylation through an interaction with folate status. Proc. Natl. Acad. Sci. USA 2002, 99, 5606–5611. [Google Scholar] [CrossRef] [Green Version]
- Hermann, A.; Goyal, R.; Jeltsch, A. The Dnmt1 DNA-(cytosine-C5)-methyltransferase methylates DNA processively with high preference for hemimethylated target sites. J. Biol. Chem. 2004, 279, 48350–48359. [Google Scholar] [CrossRef] [Green Version]
- Kaneda, M.; Okano, M.; Hata, K.; Sado, T.; Tsujimoto, N.; Li, E.; Sasaki, H. Essential role for de novo DNA methyltransferase Dnmt3a in paternal and maternal imprinting. Nature 2004, 429, 900–903. [Google Scholar] [CrossRef]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Cedar, H.; Bergman, Y. Linking DNA methylation and histone modification: Patterns and paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Hackett, J.A.; Sengupta, R.; Zylicz, J.J.; Murakami, K.; Lee, C.; Down, T.A.; Surani, M.A. Germline DNA Demethylation Dynamics and Imprint Erasure Through 5-Hydroxymethylcytosine. Science 2013, 339, 448–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyes, J.; Bird, A. DNA methylation inhibits transcription indirectly via a methyl-CpG binding protein. Cell 1991, 64, 1123–1134. [Google Scholar] [CrossRef]
- Hark, A.T.; Schoenherr, C.J.; Katz, D.J.; Ingram, R.S.; Levorse, J.M.; Tilghman, S.M. CTCF mediates methylation-sensitive enhancer-blocking activity at the H19/Igf2 locus. Nature 2000, 405, 486–489. [Google Scholar] [CrossRef]
- Arechederra, M.; Daian, F.; Yim, A.; Bazai, S.K.; Richelme, S.; Dono, R.; Saurin, A.J.; Habermann, B.H.; Maina, F. Hypermethylation of gene body CpG islands predicts high dosage of functional oncogenes in liver cancer. Nat. Commun. 2018, 9, 3164. [Google Scholar] [CrossRef]
- Christopher, M.A.; Kyle, S.M.; Katz, D.J. Neuroepigenetic mechanisms in disease. Epigenet. Chromatin 2017, 10, 47. [Google Scholar] [CrossRef]
- Rottman, F.; Shatkin, A.J.; Perry, R.P. Sequences containing methylated nucleotides at the 5’ termini of messenger RNAs: Possible implications for processing. Cell 1974, 3, 197–199. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N6-methyladenosine Modulates Messenger RNA Translation Efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.-B.; Jaffrey, S.R. 5′ UTR m6A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Bodi, Z.; Bottley, A.; Archer, N.; May, S.T.; Fray, R.G. Yeast m6A Methylated mRNAs Are Enriched on Translating Ribosomes during Meiosis, and under Rapamycin Treatment. PLoS ONE 2015, 10, e0132090. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.-W.; Huang, K.; Yang, C.; Kang, C.-S. Non-coding RNAs as regulators in epigenetics. Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peschansky, V.J.; Wahlestedt, C. Non-coding RNAs as direct and indirect modulators of epigenetic regulation. Epigenetics 2014, 9, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Robles, V.; Valcarce, D.G.; Riesco, M.F. Non-coding RNA regulation in reproduction: Their potential use as biomarkers. Non-coding RNA Res. 2019, 4, 54–62. [Google Scholar] [CrossRef]
- Aleshkina, D.; Iyyappan, R.; Lin, C.J.; Masek, T.; Pospisek, M.; Susor, A. ncRNA BC1 influences translation in the oocyte. RNA Biol. 2021, 18, 1893–1904. [Google Scholar] [CrossRef]
- Kalish, J.M.; Jiang, C.; Bartolomei, M.S. Epigenetics and imprinting in human disease. Int. J. Dev. Biol. 2014, 58, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Bouckenheimer, J.; Assou, S.; Riquier, S.; Hou, C.; Philippe, N.; Sansac, C.; Lavabre-Bertrand, T.; Commes, T.; Lemaître, J.-M.; Boureux, A.; et al. Long non-coding RNAs in human early embryonic development and their potential in ART. Hum. Reprod. Update 2016, 23, 19–40. [Google Scholar] [CrossRef]
- Seisenberger, S.; Andrews, S.; Krueger, F.; Arand, J.; Walter, J.; Santos, F.; Popp, C.; Thienpont, B.; Dean, W.; Reik, W. The dynamics of genome-wide DNA methylation reprogramming in mouse primordial germ cells. Mol. Cell 2012, 48, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Saadeh, H.; Schulz, R. Protection of CpG islands against de novo DNA methylation during oogenesis is associated with the recognition site of E2f1 and E2f2. Epigenet. Chromatin 2014, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Tomizawa, S.-I.I.; Nowacka-Woszuk, J.; Kelsey, G. DNA methylation establishment during oocyte growth: Mechanisms and significance. Int. J. Dev. Biol. 2012, 56, 867–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamudio, N.M.; Chong, S.; O’Bryan, M.K. Epigenetic regulation in male germ cells. Reproduction 2008, 136, 131–146. [Google Scholar] [CrossRef] [Green Version]
- Langenstroth-Röwer, D.; Gromoll, J.; Wistuba, J.; Tröndle, I.; Laurentino, S.; Schlatt, S.; Neuhaus, N. De novo methylation in male germ cells of the common marmoset monkey occurs during postnatal development and is maintained in vitro. Epigenetics 2017, 12, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, S.A.; Kelsey, G. De novo DNA methylation: A germ cell perspective. Trends Genet. 2012, 28, 33–42. [Google Scholar] [CrossRef] [PubMed]
- von Meyenn, F.; Berrens, R.V.; Andrews, S.; Santos, F.; Collier, A.J.; Krueger, F.; Osorno, R.; Dean, W.; Rugg-Gunn, P.J.; Reik, W. Comparative Principles of DNA Methylation Reprogramming during Human and Mouse In Vitro Primordial Germ Cell Specification. Dev. Cell 2016, 39, 104–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, K.R.; Veselovska, L.; Kelsey, G. Establishment and functions of DNA methylation in the germline. Epigenomics 2016, 8, 1399–1413. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, R.; Eppig, J.J. Transcriptional Activity of the Mouse Oocyte Genome: Companion Granulosa Cells Modulate Transcription and Chromatin Remodeling. Dev. Biol. 2001, 229, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Mira, A. Why is Meiosis Arrested? J. Theor. Biol. 1998, 194, 275–287. [Google Scholar] [CrossRef]
- Kaneda, M.; Hirasawa, R.; Chiba, H.; Okano, M.; Li, E.; Sasaki, H. Genetic evidence for Dnmt3a-dependent imprinting during oocyte growth obtained by conditional knockout with Zp3-Cre and complete exclusion of Dnmt3b by chimera formation. Genes Cells 2010, 15, 169–179. [Google Scholar] [CrossRef]
- Branco, M.R.; King, M.; Perez-Garcia, V.; Bogutz, A.B.; Caley, M.; Fineberg, E.; Lefebvre, L.; Cook, S.J.; Dean, W.; Hemberger, M.; et al. Maternal DNA Methylation Regulates Early Trophoblast Development. Dev. Cell 2016, 36, 152–163. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Zhang, T.; Yang, Y.; Wang, C. Mechanisms of Oocyte Maturation and Related Epigenetic Regulation. Front. Cell Dev. Biol. 2021, 9, 654028. [Google Scholar] [CrossRef] [PubMed]
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The many roles of histone deacetylases in development and physiology: Implications for disease and therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Liu, H.; Tazaki, M.; Nagata, M.; Aoki, F. Changes in histone acetylation during mouse oocyte meiosis. J. Cell Biol. 2003, 162, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, X.; Gao, M.; Han, L.; Qiu, D.; Wang, H.; Xiong, B.; Sun, S.C.; Liu, H.; Gu, L. HDAC3 promotes meiotic apparatus assembly in mouse oocytes by modulating tubulin acetylation. Development 2017, 144, 3789–3797. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Li, X.; Liu, X.; Gao, M.; He, Y.; Xiong, B.; Liu, H. HDAC3 inhibition disrupts the assembly of meiotic apparatus during porcine oocyte maturation. J. Cell. Physiol. 2019, 234, 10178–10183. [Google Scholar] [CrossRef]
- Zhou, D.; Choi, Y.J.; Kim, J.H. Histone deacetylase 6 (HDAC6) is an essential factor for oocyte maturation and asymmetric division in mice. Sci. Rep. 2017, 7, 8131. [Google Scholar] [CrossRef] [Green Version]
- Ling, L.; Hu, F.; Ying, X.; Ge, J.; Wang, Q. HDAC6 inhibition disrupts maturational progression and meiotic apparatus assembly in mouse oocytes. Cell Cycle 2018, 17, 550–556. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; He, M.; Zhao, L.; Qin, S.; Zhu, Z.; Du, X.; Zhou, B.; Yang, Y.; Liu, X.; Xia, G.; et al. HDAC6 regulates primordial follicle activation through mTOR signaling pathway. Cell Death Dis. 2021, 12, 559. [Google Scholar] [CrossRef]
- Zhang, K.; Lu, Y.; Jiang, C.; Liu, W.; Shu, J.; Chen, X.; Shi, Y.; Wang, E.; Wang, L.; Hu, Q.; et al. HDAC8 functions in spindle assembly during mouse oocyte meiosis. Oncotarget 2017, 8, 20092–20102. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Pan, C.; Lu, Y.; Miao, Y.; Xiong, B. HDAC8 drives spindle organization during meiotic maturation of porcine oocytes. Cell Prolif. 2021, 54, e13119. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, H.; Wang, H.; Dang, Y.; Luo, L.; Li, S.; Shi, Y.; Wang, L.; Wang, S.; Mager, J.; et al. Essential roles of HDAC1 and 2 in lineage development and genome-wide DNA methylation during mouse preimplantation development. Epigenetics 2020, 15, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Sui, L.; Zhang, S.; Huang, R.; Li, Z. HDAC11 promotes meiotic apparatus assembly during mouse oocyte maturation via decreasing H4K16 and α-tubulin acetylation. Cell Cycle 2020, 19, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; di Emidio, G.; Barbonetti, A.; Carta, G.; Luciano, A.M.; Falone, S.; Amicarelli, F. Sirtuins in gamete biology and reproductive physiology: Emerging roles and therapeutic potential in female and male infertility. Hum. Reprod. Update 2018, 24, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Du, X.; Zhao, L.; He, M.; Lin, L.; Guo, C.; Zhang, X.; Han, J.; Yan, H.; Huang, K.; et al. SIRT1 facilitates primordial follicle recruitment independent of deacetylase activity through directly modulating Akt1 and mTOR transcription. FASEB J. 2019, 33, 14703–14716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iljas, J.D.; Wei, Z.; Homer, H.A. Sirt1 sustains female fertility by slowing age-related decline in oocyte quality required for post-fertilization embryo development. Aging Cell 2020, 19, e13204. [Google Scholar] [CrossRef]
- Xing, X.; Zhang, J.; Wu, T.; Zhang, J.; Wang, Y.; Su, J.; Zhang, Y. SIRT1 reduces epigenetic and non-epigenetic changes to maintain the quality of postovulatory aged oocytes in mice. Exp. Cell Res. 2021, 399, 112421. [Google Scholar] [CrossRef]
- Xu, D.; He, H.; Liu, D.; Geng, G.; Li, Q. A novel role of SIRT2 in regulating gap junction communications via connexin-43 in bovine cumulus-oocyte complexes. J. Cell. Physiol. 2020, 235, 7332–7343. [Google Scholar] [CrossRef]
- Shen, W.-B.; Ni, J.; Yao, R.; Goetzinger, K.R.; Harman, C.; Reece, E.A.; Wang, B.; Yang, P. Maternal obesity increases DNA methylation and decreases RNA methylation in the human placenta. Reprod. Toxicol. 2022, 107, 90–96. [Google Scholar] [CrossRef]
- Gao, M.; Li, X.; He, Y.; Han, L.; Qiu, D.; Ling, L.; Liu, H.; Liu, J.; Gu, L. SIRT7 functions in redox homeostasis and cytoskeletal organization during oocyte maturation. FASEB J. 2018, 32, 6228–6238. [Google Scholar] [CrossRef]
- Zeng, J.; Jiang, M.; Wu, X.; Diao, F.; Qiu, D.; Hou, X.; Wang, H.; Li, L.; Li, C.; Ge, J.; et al. SIRT4 is essential for metabolic control and meiotic structure during mouse oocyte maturation. Aging Cell 2018, 17, e12789. [Google Scholar] [CrossRef]
- Ge, J.; Li, C.; Li, C.; Huang, Z.; Zeng, J.; Han, L.; Wang, Q. SIRT6 participates in the quality control of aged oocytes via modulating telomere function. Aging 2019, 11, 1965–1976. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Lombard, D.B. Mitochondrial Sirtuins and Their Relationships with Metabolic Disease and Cancer. Antioxid. Redox Signal. 2015, 22, 1060–1077. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; Di Emidio, G.; Vitti, M.; Di Carlo, M.; Santini, S.; D’Alessandro, A.M.; Falone, S.; Amicarelli, F. Sirtuin Functions in Female Fertility: Possible Role in Oxidative Stress and Aging. Oxid. Med. Cell. Longev. 2015, 2015, 659687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Emidio, G.; Falone, S.; Vitti, M.; D’Alessandro, A.M.; Vento, M.; Di Pietro, C.; Amicarelli, F.; Tatone, C. SIRT1 signalling protects mouse oocytes against oxidative stress and is deregulated during aging. Hum. Reprod. 2014, 29, 2006–2017. [Google Scholar] [CrossRef]
- Guo, L.; Liu, X.; Chen, H.; Wang, W.; Gu, C.; Li, B. Decrease in ovarian reserve through the inhibition of SIRT1-mediated oxidative phosphorylation. Aging 2022, 14, 2335–2347. [Google Scholar] [CrossRef]
- Kawamura, Y.; Uchijima, Y.; Horike, N.; Tonami, K.; Nishiyama, K.; Amano, T.; Asano, T.; Kurihara, Y.; Kurihara, H. Sirt3 protects in vitro–fertilized mouse preimplantation embryos against oxidative stress–induced p53-mediated developmental arrest. J. Clin. Investig. 2010, 120, 2817–2828. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.-C.; Ding, T.; Ren, Y.; Li, T.-J.; Li, R.; Fan, Y.; Yan, J.; Zhao, Y.; Li, M.; Yu, Y.; et al. Role of Sirt3 in mitochondrial biogenesis and developmental competence of human in vitro matured oocytes. Hum. Reprod. 2016, 31, 607–622. [Google Scholar] [CrossRef] [Green Version]
- Hatefi, Y. The mitochondrial electron transport and oxidative phosphorylation system. Annu. Rev. Biochem. 1985, 54, 1015–1069. [Google Scholar] [CrossRef]
- McBride, H.M.; Neuspiel, M.; Wasiak, S. Mitochondria: More Than Just a Powerhouse. Curr. Biol. 2006, 16, R551–R560. [Google Scholar] [CrossRef] [Green Version]
- Shutt, T.E.; Shadel, G.S. A compendium of human mitochondrial gene expression machinery with links to disease. Environ. Mol. Mutagen. 2010, 51, 360–379. [Google Scholar] [CrossRef] [Green Version]
- Wallace, D.C. A Mitochondrial Paradigm of Metabolic and Degenerative Diseases, Aging, and Cancer: A Dawn for Evolutionary Medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Blerkom, J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 2011, 11, 797–813. [Google Scholar] [CrossRef] [PubMed]
- Chappel, S. The Role of Mitochondria from Mature Oocyte to Viable Blastocyst. Obstet. Gynecol. Int. 2013, 2013, 183024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynier, P.; May-Panloup, P.; Chretien, M.-F.; Morgan, C.J.; Jean, M.; Savagner, F.; Barriere, P.; Malthiery, Y. Mitochondrial DNA content affects the fertilizability of human oocytes. Mol. Hum. Reprod. 2001, 7, 425–429. [Google Scholar] [CrossRef] [Green Version]
- Santos, T.A.; El Shourbagy, S.; John, J.C.S. Mitochondrial content reflects oocyte variability and fertilization outcome. Fertil. Steril. 2006, 85, 584–591. [Google Scholar] [CrossRef] [Green Version]
- Al Rawi, S.; Louvet-Vallée, S.; Djeddi, A.; Sachse, M.; Culetto, E.; Hajjar, C.; Boyd, L.; Legouis, R.; Galy, V. Postfertilization Autophagy of Sperm Organelles Prevents Paternal Mitochondrial DNA Transmission. Science 2011, 334, 1144–1147. [Google Scholar] [CrossRef]
- Cummins, J.M.; Wakayama, T.; Yanagimachi, R. Fate of microinjected sperm components in the mouse oocyte and embryo. Zygote 1997, 5, 301–308. [Google Scholar] [CrossRef]
- Perry, J.R.B.; Murray, A.; Day, F.R.; Ong, K.K. Molecular insights into the aetiology of female reproductive ageing. Nat. Rev. Endocrinol. 2015, 11, 725–734. [Google Scholar] [CrossRef]
- Ben-Meir, A.; Burstein, E.; Borrego-Alvarez, A.; Chong, J.; Wong, E.; Yavorska, T.; Naranian, T.; Chi, M.; Wang, Y.; Bentov, Y.; et al. Coenzyme Q10 restores oocyte mitochondrial function and fertility during reproductive aging. Aging Cell 2015, 14, 887–895. [Google Scholar] [CrossRef]
- Harris, S.E.; Adriaens, I.; Leese, H.J.; Gosden, R.G.; Picton, H.M. Carbohydrate metabolism by murine ovarian follicles and oocytes grown in vitro. Reproduction 2007, 134, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Benkhalifa, M.; Ferreira, Y.J.; Chahine, H.; Louanjli, N.; Miron, P.; Merviel, P.; Copin, H. Mitochondria: Participation to infertility as source of energy and cause of senescence. Int. J. Biochem. Cell Biol. 2014, 55, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, S.L.; Lane, M.; Mitchell, M. Impaired Mitochondrial Function in the Preimplantation Embryo Perturbs Fetal and Placental Development in the Mouse1. Biol. Reprod. 2011, 84, 572–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wai, T.; Ao, A.; Zhang, X.; Cyr, D.; Dufort, D.; Shoubridge, E.A. The Role of Mitochondrial DNA Copy Number in Mammalian Fertility1. Biol. Reprod. 2010, 83, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Guillaumet-Adkins, A.; Yañez, Y.; Peris-Diaz, M.D.; Calabria, I.; Palanca-Ballester, C.; Sandoval, J. Epigenetics and Oxidative Stress in Aging. Oxid. Med. Cell. Longev. 2017, 2017, 9175806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wu, X.Q.; Lu, S.; Guo, Y.L.; Ma, X. Deficit of mitochondria-derived ATP during oxidative stress impairs mouse MII oocyte spindles. Cell Res. 2006, 16, 841–850. [Google Scholar] [CrossRef] [Green Version]
- Smeets, H.J.M.; Sallevelt, S.C.E.H.; Dreesen, J.C.F.M.; de Die-Smulders, C.E.M.; de Coo, I.F.M. Preventing the transmission of mitochondrial DNA disorders using prenatal or preimplantation genetic diagnosis. Ann. N. Y. Acad. Sci. 2015, 1350, 29–36. [Google Scholar] [CrossRef]
- Wolf, D.P.; Mitalipov, N.; Mitalipov, S. Mitochondrial replacement therapy in reproductive medicine. Trends Mol. Med. 2015, 21, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Babayev, E.; Wang, T.; Szigeti-Buck, K.; Lowther, K.; Taylor, H.S.; Horvath, T.; Seli, E. Reproductive aging is associated with changes in oocyte mitochondrial dynamics, function, and mtDNA quantity. Maturitas 2016, 93, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Spikings, E.C.; Alderson, J.; John, J.C.S. Regulated Mitochondrial DNA Replication During Oocyte Maturation Is Essential for Successful Porcine Embryonic Development. Biol. Reprod. 2007, 76, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Rambags, B.P.B.; van Boxtel, D.C.J.; Tharasanit, T.; Lenstra, J.A.; Colenbrander, B.; Stout, T.A.E. Advancing maternal age predisposes to mitochondrial damage and loss during maturation of equine oocytes in vitro. Theriogenology 2014, 81, 959–965. [Google Scholar] [CrossRef]
- Igarashi, H.; Takahashi, T.; Takahashi, E.; Tezuka, N.; Nakahara, K.; Takahashi, K.; Kurachi, H. Aged Mouse Oocytes Fail to Readjust Intracellular Adenosine Triphosphates at Fertilization1. Biol. Reprod. 2005, 72, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Luderer, U. Oxidative Damage Increases and Antioxidant Gene Expression Decreases with Aging in the Mouse Ovary. Biol. Reprod. 2011, 84, 775–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cree, L.M.; Hammond, E.R.; Shelling, A.N.; Berg, M.C.; Peek, J.C.; Green, M.P. Maternal age and ovarian stimulation independently affect oocyte mtDNA copy number and cumulus cell gene expression in bovine clones. Hum. Reprod. 2015, 30, 1410–1420. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.F.; Soares, M.; Almeida Reis, S.; Ramalho-Santos, J.; Sousa, A.P.; Almeida-Santos, T. Does supplementation with mitochondria improve oocyte competence? A systematic review. Reproduction 2021, 161, 269–287. [Google Scholar] [CrossRef]
- Sobek, A.; Tkadlec, E.; Klaskova, E.; Prochazka, M. Cytoplasmic Transfer Improves Human Egg Fertilization and Embryo Quality: An Evaluation of Sibling Oocytes in Women with Low Oocyte Quality. Reprod. Sci. 2021, 28, 1362–1369. [Google Scholar] [CrossRef]
- Ferguson-Smith, A.C. Genomic imprinting: The emergence of an epigenetic paradigm. Nat. Rev. Genet. 2011, 12, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Hendrich, B.; Reik, W.; Dean, W. Dynamic Reprogramming of DNA Methylation in the Early Mouse Embryo. Dev. Biol. 2002, 241, 172–182. [Google Scholar] [CrossRef] [Green Version]
- Tucci, V.; Isles, A.R.; Kelsey, G.; Ferguson-Smith, A.C.; Tucci, V.; Bartolomei, M.S.; Benvenisty, N.; Bourc’his, D.; Charalambous, M.; Dulac, C.; et al. Genomic Imprinting and Physiological Processes in Mammals. Cell 2019, 176, 952–965. [Google Scholar] [CrossRef] [Green Version]
- McGrath, J.; Solter, D. Completion of mouse embryogenesis requires both the maternal and paternal genomes. Cell 1984, 37, 179–183. [Google Scholar] [CrossRef]
- Surani, M.A. Imprinting and the Initiation of Gene Silencing in the Germ Line. Cell 1998, 93, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Goovaerts, T.; Steyaert, S.; Vandenbussche, C.A.; Galle, J.; Thas, O.; Van Criekinge, W.; De Meyer, T. A comprehensive overview of genomic imprinting in breast and its deregulation in cancer. Nat. Commun. 2018, 9, 4120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, N.; Kamataki, A.; Kurotaki, H. Methylation profile of imprinted genes provides evidence for teratomatous origin of a subset of mucinous ovarian tumours. J. Pathol. 2021, 254, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, M.; Ferguson-Smith, A.C. DNA methylation in genomic imprinting, development, and disease. J. Pathol. 2001, 195, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Elhamamsy, A.R. Role of DNA methylation in imprinting disorders: An updated review. J. Assist. Reprod. Genet. 2017, 34, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, H.; Sakurai, T.; Imai, M.; Takahashi, N.; Fukuda, A.; Yayoi, O.; Sato, S.; Nakabayashi, K.; Hata, K.; Sotomaru, Y.; et al. Contribution of Intragenic DNA Methylation in Mouse Gametic DNA Methylomes to Establish Oocyte-Specific Heritable Marks. PLoS Genet. 2012, 8, e1002440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paczkowski, M.; Schoolcraft, W.B.; Krisher, R.L. Dysregulation of methylation and expression of imprinted genes in oocytes and reproductive tissues in mice of advanced maternal age. J. Assist. Reprod. Genet. 2015, 32, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Duranthon, V.; Watson, A.J.; Lonergan, P. Preimplantation embryo programming: Transcription, epigenetics, and culture environment. Reproduction 2008, 135, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zhu, P.; Yan, L.; Li, R.; Hu, B.; Lian, Y.; Yan, J.; Ren, X.; Lin, S.; Li, J.; et al. The DNA methylation landscape of human early embryos. Nature 2014, 511, 606–610. [Google Scholar] [CrossRef]
- Smith, Z.D.; Chan, M.M.; Mikkelsen, T.S.; Gu, H.; Gnirke, A.; Regev, A.; Meissner, A. A unique regulatory phase of DNA methylation in the early mammalian embryo. Nature 2012, 484, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.E.; Jiang, Z.C.; Alqahtani, F.; Mandoiu, I.; Dong, H.; Zheng, X.; Marjani, S.L.; Chen, J.; Tian, X.C. Methylome Dynamics of Bovine Gametes and in vivo Early Embryos. Front. Genet. 2019, 10, 512. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.; Zhang, G.; Cai, Y.; Liu, Z.; Zhang, Y.; Meng, F.; Wang, F.; Wan, Y. DNA methylation dynamics during zygotic genome activation in goat. Theriogenology 2020, 156, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Hanna, C.W.; Demond, H.; Kelsey, G. Epigenetic regulation in development: Is the mouse a good model for the human? Hum. Reprod. Update 2018, 24, 556–576. [Google Scholar] [CrossRef] [PubMed]
- Amouroux, R.; Nashun, B.; Shirane, K.; Nakagawa, S.; Hill, P.W.S.; D’Souza, Z.; Nakayama, M.; Matsuda, M.; Turp, A.; Ndjetehe, E.; et al. De novo DNA methylation drives 5hmC accumulation in mouse zygotes. Nat. Cell Biol. 2016, 18, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haaf, T. The battle of the sexes after fertilization: Behaviour of paternal and maternal chromosomes in the early mammalian embryo. Chromosome Res. 2001, 9, 263–271. [Google Scholar] [CrossRef]
- Messerschmidt, D.M.; Knowles, B.B.; Solter, D. DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos. Genes Dev. 2014, 28, 812–828. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Jayavelu, N.D.; Battle, S.L.; Mar, J.C.; Schimmel, T.; Cohen, J.; Hawkins, R.D. Single-cell analysis of transcriptome and DNA methylome in human oocyte maturation. PLoS ONE 2020, 15, e0241698. [Google Scholar] [CrossRef]
- Zeng, Y.; Ren, R.; Kaur, G.; Hardikar, S.; Ying, Z.; Babcock, L.; Gupta, E.; Zhang, X.; Chen, T.; Cheng, X. The inactive Dnmt3b3 isoform preferentially enhances Dnmt3b-mediated DNA methylation. Genes Dev. 2020, 34, 1546–1558. [Google Scholar] [CrossRef]
- Sabag, O.; Zamir, A.; Keshet, I.; Hecht, M.; Ludwig, G.; Tabib, A.; Moss, J.; Cedar, H. Establishment of methylation patterns in ES cells. Nat. Struct. Mol. Biol. 2014, 21, 110–112. [Google Scholar] [CrossRef]
- Li, E.; Bestor, T.H.; Jaenisch, R. Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell 1992, 69, 915–926. [Google Scholar] [CrossRef]
- Ginsburg, M.; Snow, M.H.L.; McLaren, A. Primordial germ cells in the mouse embryo during gastrulation. Development 1990, 110, 521–528. [Google Scholar] [CrossRef]
- Pepling, M.E.; Spradling, A.C. Mouse Ovarian Germ Cell Cysts Undergo Programmed Breakdown to Form Primordial Follicles. Dev. Biol. 2001, 234, 339–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepling, M.E.; Spradling, A.C. Female mouse germ cells form synchronously dividing cysts. Development 1998, 125, 3323–3328. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K. Regulation of primordial follicle assembly and development. Hum. Reprod. Update 2005, 11, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- HSUEH, A.J.W.; BILLIG, H.; TSAFRIRI, A. Ovarian Follicle Atresia: A Hormonally Controlled Apoptotic Process*. Endocr. Rev. 1994, 15, 707–724. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, C.Y.; Zhao, Y.; Dean, J. FIGLA, LHX8 and SOHLH1 transcription factor networks regulate mouse oocyte growth and differentiation. Nucleic Acids Res. 2020, 48, 3525–3541. [Google Scholar] [CrossRef]
- Driancourt, M.A.; Reynaud, K.; Cortvrindt, R.; Smitz, J. Roles of KIT and KIT LIGAND in ovarian function. Rev. Reprod. 2000, 5, 143–152. [Google Scholar] [CrossRef]
- Jin, X.; Han, C.S.; Yu, F.Q.; Wei, P.; Hu, Z.Y.; Liu, Y.X. Anti-apoptotic action of stem cell factor on oocytes in primordial follicles and its signal transduction. Mol. Reprod. Dev. 2005, 70, 82–90. [Google Scholar] [CrossRef]
- Eppig, J.J. Oocyte control of ovarian follicular development and function in mammals. Reproduction 2001, 122, 829–838. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Ritter, L.J.; Myllymaa, S.; Kaivo-Oja, N.; Dragovic, R.A.; Hickey, T.E.; Ritvos, O.; Mottershead, D.G. Molecular basis of oocyte-paracrine signalling that promotes granulosa cell proliferation. J. Cell Sci. 2006, 119, 3811–3821. [Google Scholar] [CrossRef] [Green Version]
- Macaulay, A.D.; Gilbert, I.; Caballero, J.; Barreto, R.; Fournier, E.; Tossou, P.; Sirard, M.A.; Clarke, H.J.; Khandjian, É.W.; Richard, F.J.; et al. The gametic synapse: RNA transfer to the bovine oocyte. Biol. Reprod. 2014, 91, 1–12. [Google Scholar] [CrossRef]
- Macaulay, A.D.; Gilbert, I.; Scantland, S.; Fournier, E.; Ashkar, F.; Bastien, A.; Shojaei Saadi, H.A.; Gagné, D.; Sirard, M.A.; Khandjian, É.W.; et al. Cumulus cell transcripts transit to the bovine oocyte in preparation for maturation. Biol. Reprod. 2016, 94, 16–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternlicht, A.L.; Schultz, R.M. Biochemical studies of mammalian oogenesis: Kinetics of accumulation of total and poly(A)-containing RNA during growth of the mouse oocyte. J. Exp. Zool. 1981, 215, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, F.; Smitz, J. Molecular control of oogenesis. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 1896–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Gonzalez, R.; Moreira, P.N.; Pérez-Crespo, M.; Sánchez-Martín, M.; Ramirez, M.A.; Pericuesta, E.; Bilbao, A.; Bermejo-Alvarez, P.; Hourcade, J.D.; Fonseca, F.R.; et al. Long-Term Effects of Mouse Intracytoplasmic Sperm Injection with DNA-Fragmented Sperm on Health and Behavior of Adult Offspring1. Biol. Reprod. 2008, 78, 761–772. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.-J.; Lahr, R.M.; Solgaard, M.T.; Moraes, B.J.; Pointet, R.; Yang, A.-D.; Celucci, G.; Graber, T.E.; Hoang, H.-D.; Niklaus, M.R.; et al. mTORC1 promotes TOP mRNA translation through site-specific phosphorylation of LARP1. Nucleic Acids Res. 2021, 49, 3461–3489. [Google Scholar] [CrossRef] [PubMed]
- Clegg, K.B.; Pikó, L. Quantitative aspects of RNA synthesis and polyadenylation in 1-cell and 2-cell mouse embryos. J. Embryol. Exp. Morphol. 1983, 74, 169–182. [Google Scholar] [CrossRef]
- Martins, J.P.S.; Liu, X.; Oke, A.; Arora, R.; Franciosi, F.; Viville, S.; Laird, D.J.; Fung, J.C.; Conti, M. DAZL and CPEB1 regulate mRNA translation synergistically during oocyte maturation. J. Cell Sci. 2016, 129, 1271–1282. [Google Scholar] [CrossRef] [Green Version]
- Jansova, D.; Koncicka, M.; Tetkova, A.; Cerna, R.; Malik, R.; del Llano, E.; Kubelka, M.; Susor, A. Regulation of 4E-BP1 activity in the mammalian oocyte. Cell Cycle 2017, 16, 927–939. [Google Scholar] [CrossRef]
- Kim, J.; Guan, K.-L.L. mTOR as a central hub of nutrient signalling and cell growth. Nat. Cell Biol. 2019, 21, 63–71. [Google Scholar] [CrossRef]
- Kalous, J.; Jansová, D.; Šušor, A. Role of Cyclin-Dependent Kinase 1 in Translational Regulation in the M-Phase. Cells 2020, 9, 1568. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, T.; Guo, Y.; Sun, T.; Li, H.; Zhang, X.; Yin, H.; Cao, G.; Yin, Y.; Wang, H.; et al. Oocyte stage-specific effects of MTOR determine granulosa cell fate and oocyte quality in mice. Proc. Natl. Acad. Sci. USA 2018, 115, E5326–E5333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schisa, J.A. Germ Cell Responses to Stress: The Role of RNP Granules. Front. Cell Dev. Biol. 2019, 7, 220. [Google Scholar] [CrossRef] [PubMed]
- Standart, N.; Weil, D. P-Bodies: Cytosolic Droplets for Coordinated mRNA Storage. Trends Genet. 2018, 34, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Blatt, P.; Wong-Deyrup, S.W.; McCarthy, A.; Breznak, S.; Hurton, M.D.; Upadhyay, M.; Bennink, B.; Camacho, J.; Lee, M.T.; Rangan, P. RNA degradation is required for the germ-cell to maternal transition in Drosophila. Curr. Biol. 2021, 31, 2984–2994. [Google Scholar] [CrossRef]
- Qi, S.T.; Ma, J.Y.; Wang, Z.B.; Guo, L.; Hou, Y.; Sun, Q.Y. N6-Methyladenosine Sequencing Highlights the Involvement of mRNA Methylation in Oocyte Meiotic Maturation and Embryo Development by Regulating Translation in Xenopus laevis. J. Biol. Chem. 2016, 291, 23020–23026. [Google Scholar] [CrossRef] [Green Version]
- Sui, X.; Hu, Y.; Ren, C.; Cao, Q.; Zhou, S.; Cao, Y.; Li, M.; Shu, W.; Huo, R. METTL3-mediated m6A is required for murine oocyte maturation and maternal-to-zygotic transition. Cell Cycle 2020, 19, 391–404. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, S.; Zhai, Y.; Han, Y.; Huang, R.; An, X.; Dai, X.; Li, Z. Cycloleucine negatively regulates porcine oocyte maturation and embryo development by modulating N6-methyladenosine and histone modifications. Theriogenology 2022, 179, 128–140. [Google Scholar] [CrossRef]
- Kwon, J.; Jo, Y.J.; Namgoong, S.; Kim, N.H. Functional roles of hnRNPA2/B1 regulated by METTL3 in mammalian embryonic development. Sci. Rep. 2019, 9, 8640. [Google Scholar] [CrossRef] [Green Version]
- Meng, T.G.; Lu, X.; Guo, L.; Hou, G.M.; Ma, X.S.; Li, Q.N.; Huang, L.; Fan, L.H.; Zhao, Z.H.; Ou, X.H.; et al. Mettl14 is required for mouse postimplantation development by facilitating epiblast maturation. FASEB J. 2019, 33, 1179–1187. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, X.; Weng, Y.-L.; Lu, Z.; Liu, Y.; Lu, Z.; Li, J.; Hao, P.; Zhang, Y.; Zhang, F.; et al. m6A facilitates hippocampus-dependent learning and memory through YTHDF1. Nature 2018, 563, 249–253. [Google Scholar] [CrossRef]
- Yang, Y.; Hsu, P.J.; Chen, Y.-S.; Yang, Y.-G. Dynamic transcriptomic m6A decoration: Writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Ping, X.-L.; Sun, B.-F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.-J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.-S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, H.; Zhang, T.; Yang, Y.; Zhang, D.; Gao, J.; Li, J.; Yue, L.; Gao, D.; Shi, B.; Han, Y.; et al. METTL3-mediated mRNA N6-methyladenosine is required for oocyte and follicle development in mice. Cell Death Dis. 2021, 12, 989. [Google Scholar] [CrossRef] [PubMed]
- Raj, N.; Wang, M.; Seoane, J.A.; Zhao, R.L.; Kaiser, A.M.; Moonie, N.A.; Demeter, J.; Boutelle, A.M.; Kerr, C.H.; Mulligan, A.S.; et al. The Mettl3 epitranscriptomic writer amplifies p53 stress responses. Mol. Cell 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Wu, Y.; Liu, Y.; Weng, H.; Huang, H. N 6 -methyladenosine Steers RNA Metabolism and Regulation in Cancer. Cancer Commun. 2021, 41, 538–559. [Google Scholar] [CrossRef]
- Parial, R.; Li, H.; Li, J.; Archacki, S.; Yang, Z.; Wang, I.Z.; Chen, Q.; Xu, C.; Wang, Q.K. Role of epigenetic m 6 A RNA methylation in vascular development: Mettl3 regulates vascular development through PHLPP2/mTOR-AKT. signaling. FASEB J. 2021, 35, e21465. [Google Scholar] [CrossRef]
- Fan, L.H.; Wang, Z.B.; Li, Q.N.; Meng, T.G.; Dong, M.Z.; Hou, Y.; Ouyang, Y.C.; Schatten, H.; Sun, Q.Y. Absence of mitochondrial DNA methylation in mouse oocyte maturation, aging and early embryo development. Biochem. Biophys. Res. Commun. 2019, 513, 912–918. [Google Scholar] [CrossRef]
- Zaccara, S.; Jaffrey, S.R. A Unified Model for the Function of YTHDF Proteins in Regulating m6A-Modified mRNA. Cell 2020, 181, 1582–1595. [Google Scholar] [CrossRef]
- Dong, G.; Yu, J.; Shan, G.; Su, L.; Yu, N.; Yang, S. N6-Methyladenosine Methyltransferase METTL3 Promotes Angiogenesis and Atherosclerosis by Upregulating the JAK2/STAT3 Pathway via m6A Reader IGF2BP1. Front. Cell Dev. Biol. 2021, 9, 731810. [Google Scholar] [CrossRef]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 facilitates translation and decay of N6-methyladenosine-modified RNA. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, I.; Much, C.; Di Giacomo, M.; Azzi, C.; Morgan, M.; Moreira, P.N.; Monahan, J.; Carrieri, C.; Enright, A.J.; O’Carroll, D. The RNA m 6 A Reader YTHDF2 Is Essential for the Post-transcriptional Regulation of the Maternal Transcriptome and Oocyte Competence. Mol. Cell 2017, 67, 1059–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Sun, J.; Bu, S.; Li, B.; Zhang, Q.; Wang, Q.; Lai, D. Melatonin protects against chronic stress-induced oxidative meiotic defects in mice MII oocytes by regulating SIRT1. Cell Cycle 2020, 19, 1677–1695. [Google Scholar] [CrossRef]
- Rosario, F.J.; Powell, T.L.; Jansson, T. Activation of placental insulin and mTOR signaling in a mouse model of maternal obesity associated with fetal overgrowth. Am. J. Physiol. Integr. Comp. Physiol. 2016, 310, R87–R93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Wei, J.; He, C. Where, When, and How: Context-Dependent Functions of RNA Methylation Writers, Readers, and Erasers. Mol. Cell 2019, 74, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yan, G.; Wang, B.; Xu, L.; Gu, Y.; Ru, T.; Cui, X.; Lei, L.; Liu, J.; Sheng, X.; et al. Transplantation of UC-MSCs on collagen scaffold activates follicles in dormant ovaries of POF patients with long history of infertility. Sci. China Life Sci. 2018, 61, 1554–1565. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Li, X.; Lü, M.; Wang, J.; Tang, J.; Luo, S.; Qian, Y. ALKBH5 suppresses migration and invasion of human trophoblast cells by inhibiting epithelial-mesenchymal transition. Nan Fang Yi Ke Da Xue Xue Bao 2020, 40, 1720–1725. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.L.; Rivera, R.M. The effects of superovulation and reproductive aging on the epigenome of the oocyte and embryo. Mol. Reprod. Dev. 2018, 85, 90–105. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-W.; Peng, Y.-T.; Wang, B.; Zeng, Y.-H.; Zhuang, G.-L.; Zhou, C.-Q. High follicle-stimulating hormone increases aneuploidy in human oocytes matured in vitro. Fertil. Steril. 2011, 95, 99–104. [Google Scholar] [CrossRef]
- Uysal, F.; Ozturk, S.; Akkoyunlu, G. Superovulation alters DNA methyltransferase protein expression in mouse oocytes and early embryos. J. Assist. Reprod. Genet. 2018, 35, 503–513. [Google Scholar] [CrossRef]
- Ozturk, S.; Yaba-Ucar, A.; Sozen, B.; Mutlu, D.; Demir, N. Superovulation alters embryonic poly(A)-binding protein (Epab) and poly(A)-binding protein, cytoplasmic 1 (Pabpc1) gene expression in mouse oocytes and early embryos. Reprod. Fertil. Dev. 2016, 28, 375. [Google Scholar] [CrossRef]
- Market-Velker, B.A.; Zhang, L.; Magri, L.S.; Bonvissuto, A.C.; Mann, M.R.W. Dual effects of superovulation: Loss of maternal and paternal imprinted methylation in a dose-dependent manner. Hum. Mol. Genet. 2010, 19, 36–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Y.; Yan, Z.Q.; Yuan, P.; Qin, M.; Kuo, Y.; Li, R.; Yan, L.Y.; Feng, H.L.; Qiao, J. Single-cell DNA methylation sequencing reveals epigenetic alterations in mouse oocytes superovulated with different dosages of gonadotropins. Clin. Epigenet. 2020, 12, 75. [Google Scholar] [CrossRef] [PubMed]
- Fauque, P.; Jouannet, P.; Lesaffre, C.; Ripoche, M.-A.; Dandolo, L.; Vaiman, D.; Jammes, H. Assisted Reproductive Technology affects developmental kinetics, H19 Imprinting Control Region methylation and H19 gene expression in individual mouse embryos. BMC Dev. Biol. 2007, 7, 116. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-Y.; Wang, N.; Le, F.; Li, L.; Lou, H.-Y.; Liu, X.-Z.; Zheng, Y.-M.; Qian, Y.-Q.; Chen, Y.-L.; Jiang, X.-H.; et al. Superovulation Induced Changes of Lipid Metabolism in Ovaries and Embryos and Its Probable Mechanism. PLoS ONE 2015, 10, e0132638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.-K.; Wang, Q.; Zhang, T.-T.; Yin, S.; Zhang, C.-L.; Ge, Z.-J. Repeated superovulation may affect mitochondrial functions of cumulus cells in mice. Sci. Rep. 2016, 6, 31368. [Google Scholar] [CrossRef] [PubMed]
- Kalthur, G.; Salian, S.R.; Nair, R.; Mathew, J.; Adiga, S.K.; Kalthur, S.G.; Zeegers, D.; Hande, M.P. Distribution pattern of cytoplasmic organelles, spindle integrity, oxidative stress, octamer-binding transcription factor 4 (Oct4) expression and developmental potential of oocytes following multiple superovulation. Reprod. Fertil. Dev. 2016, 28, 2027. [Google Scholar] [CrossRef]
- Lee, M.; Ahn, J.I.; Lee, A.R.; Ko, D.W.; Yang, W.S.; Lee, G.; Ahn, J.Y.; Lim, J.M. Adverse Effect of Superovulation Treatment on Maturation, Function and Ultrastructural Integrity of Murine Oocytes. Mol. Cells 2017, 40, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Bui, A.D.; Sharma, R.; Henkel, R.; Agarwal, A. Reactive oxygen species impact on sperm DNA and its role in male infertility. Andrologia 2018, 50, e13012. [Google Scholar] [CrossRef]
- Bomfim, M.M.; Andrade, G.M.; del Collado, M.; Sangalli, J.R.; Fontes, P.K.; Nogueira, M.F.G.; Meirelles, F.V.; da Silveira, J.C.; Perecin, F. Antioxidant responses and deregulation of epigenetic writers and erasers link oxidative stress and DNA methylation in bovine blastocysts. Mol. Reprod. Dev. 2017, 84, 1296–1305. [Google Scholar] [CrossRef]
- Li, W.; Goossens, K.; Van Poucke, M.; Forier, K.; Braeckmans, K.; Van Soom, A.; Peelman, L.J. High oxygen tension increases global methylation in bovine 4-cell embryos and blastocysts but does not affect general retrotransposon expression. Reprod. Fertil. Dev. 2016, 28, 948. [Google Scholar] [CrossRef]
- Marei, W.F.A.; Van den Bosch, L.; Pintelon, I.; Mohey-Elsaeed, O.; Bols, P.E.J.; Leroy, J.L.M.R. Mitochondria-targeted therapy rescues development and quality of embryos derived from oocytes matured under oxidative stress conditions: A bovine in vitro model. Hum. Reprod. 2019, 34, 1984–1998. [Google Scholar] [CrossRef]
- van den Berg, M.M.J.; van Maarle, M.C.; van Wely, M.; Goddijn, M. Genetics of early miscarriage. Biochim. Biophys. Acta. Mol. Basis Dis. 2012, 1822, 1951–1959. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Wu, L.; Jiang, X.; Yang, L.; Cheng, J.; Chen, H.; Hua, R.; Geng, G.; Yang, L.; Li, Q. SIRT2 Inhibition Results in Meiotic Arrest, Mitochondrial Dysfunction, and Disturbance of Redox Homeostasis during Bovine Oocyte Maturation. Int. J. Mol. Sci. 2019, 20, 1365. [Google Scholar] [CrossRef] [Green Version]
- Reis e Silva, A.R.; Bruno, C.; Fleurot, R.; Daniel, N.; Archilla, C.; Peynot, N.; Lucci, C.M.; Beaujean, N.; Duranthon, V. Alteration of DNA demethylation dynamics by in vitro culture conditions in rabbit pre-implantation embryos. Epigenetics 2012, 7, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Estill, M.S.; Bolnick, J.M.; Waterland, R.A.; Bolnick, A.D.; Diamond, M.P.; Krawetz, S.A. Assisted reproductive technology alters deoxyribonucleic acid methylation profiles in bloodspots of newborn infants. Fertil. Steril. 2016, 106, 629–639. [Google Scholar] [CrossRef]
- Bauersachs, S.; Mermillod, P.; Almiñana, C. The Oviductal Extracellular Vesicles’ RNA Cargo Regulates the Bovine Embryonic Transcriptome. Int. J. Mol. Sci. 2020, 21, 1303. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, J.; Coutifaris, C.; Sapienza, C.; Mainigi, M. Global DNA methylation levels are altered by modifiable clinical manipulations in assisted reproductive technologies. Clin. Epigenet. 2017, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- El Hajj, N.; Haaf, T. Epigenetic disturbances in in vitro cultured gametes and embryos: Implications for human assisted reproduction. Fertil. Steril. 2013, 99, 632–641. [Google Scholar] [CrossRef]
- de Waal, E.; Vrooman, L.A.; Fischer, E.; Ord, T.; Mainigi, M.A.; Coutifaris, C.; Schultz, R.M.; Bartolomei, M.S. The cumulative effect of assisted reproduction procedures on placental development and epigenetic perturbations in a mouse model. Hum. Mol. Genet. 2015, 24, 6975–6985. [Google Scholar] [CrossRef] [Green Version]
- Pliushch, G.; Schneider, E.; Schneider, T.; El Hajj, N.; Rösner, S.; Strowitzki, T.; Haaf, T. In vitro maturation of oocytes is not associated with altered deoxyribonucleic acid methylation patterns in children from in vitro fertilization or intracytoplasmic sperm injection. Fertil. Steril. 2015, 103, 720–727. [Google Scholar] [CrossRef] [Green Version]
- Horánszky, A.; Becker, J.L.; Zana, M.; Ferguson-Smith, A.C.; Dinnyés, A. Epigenetic Mechanisms of ART-Related Imprinting Disorders: Lessons From iPSC and Mouse Models. Genes 2021, 12, 1704. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Wang, Y.; Lin, J.; Xu, J.; Ding, G.; Huang, H. Genetic and epigenetic risks of assisted reproduction. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 44, 90–104. [Google Scholar] [CrossRef] [PubMed]
- La Bastide-Van Gemert, S.; Seggers, J.; Haadsma, M.L.; Heineman, M.J.; Middelburg, K.J.; Roseboom, T.J.; Schendelaar, P.; Hadders-Algra, M.; Van den Heuvel, E.R. Is ovarian hyperstimulation associated with higher blood pressure in 4-year-old IVF offspring? Part II: An explorative causal inference approach. Hum. Reprod. 2014, 29, 510–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceelen, M.; van Weissenbruch, M.M.; Vermeiden, J.P.W.; van Leeuwen, F.E.; Delemarre-van de Waal, H.A. Growth and development of children born after in vitro fertilization. Fertil. Steril. 2008, 90, 1662–1673. [Google Scholar] [CrossRef] [PubMed]
- Håberg, S.E.; Page, C.M.; Lee, Y.; Nustad, H.E.; Magnus, M.C.; Haftorn, K.L.; Carlsen, E.Ø.; Denault, W.R.P.; Bohlin, J.; Jugessur, A.; et al. DNA methylation in newborns conceived by assisted reproductive technology. Nat. Commun. 2022, 13, 1896. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Hagen, D.E.; Elsik, C.G.; Ji, T.; Morris, C.J.; Moon, L.E.; Rivera, R.M. Characterization of global loss of imprinting in fetal overgrowth syndrome induced by assisted reproduction. Proc. Natl. Acad. Sci. USA 2015, 112, 4618–4623. [Google Scholar] [CrossRef] [Green Version]
- Tetkova, A.; Susor, A.; Kubelka, M.; Nemcova, L.; Jansova, D.; Dvoran, M.; Del Llano, E.; Holubcova, Z.; Kalous, J. Follicle-stimulating hormone administration affects amino acid metabolism in mammalian oocytes. Biol. Reprod. 2019, 101, 719–732. [Google Scholar] [CrossRef]
- Saenz-De-Juano, M.D.; Ivanova, E.; Romero, S.; Lolicato, F.; Sánchez, F.; Van Ranst, H.; Krueger, F.; Segonds-Pichon, A.; De Vos, M.; Andrews, S.; et al. DNA methylation and mRNA expression of imprinted genes in blastocysts derived from an improved in vitro maturation method for oocytes from small antral follicles in polycystic ovary syndrome patients. Hum. Reprod. 2019, 34, 1640–1649. [Google Scholar] [CrossRef]
- Fortier, A.L.; Lopes, F.L.; Darricarrère, N.; Martel, J.; Trasler, J.M. Superovulation alters the expression of imprinted genes in the midgestation mouse placenta. Hum. Mol. Genet. 2008, 17, 1653–1665. [Google Scholar] [CrossRef] [Green Version]
- Vuong, L.N.; Le, A.H.; Ho, V.N.A.; Pham, T.D.; Sanchez, F.; Romero, S.; De Vos, M.; Ho, T.M.; Gilchrist, R.B.; Smitz, J. Live births after oocyte in vitro maturation with a prematuration step in women with polycystic ovary syndrome. J. Assist. Reprod. Genet. 2020, 37, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Li, T.; Ding, C.-H.; Brosens, J.; Zhou, C.-Q.; Wang, H.-H.; Xu, Y.-W. Insufficient histone-3 lysine-9 deacetylation in human oocytes matured in vitro is associated with aberrant meiosis. Fertil. Steril. 2012, 97, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.-N.; Zhang, T.-F.; Lin, W.-Q.; Jiang, N.; Cao, H.-F.; Li, H.; Qian, J.-H. Value of serum and follicular fluid sirtuin (SIRT)1 and SIRT2 protein levels in predicting the outcome of assisted reproduction. Ann. Transl. Med. 2021, 9, 343. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Morreale, H.F. Polycystic ovary syndrome: Definition, aetiology, diagnosis and treatment. Nat. Rev. Endocrinol. 2018, 14, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Stener-Victorin, E.; Deng, Q. Epigenetic inheritance of polycystic ovary syndrome—challenges and opportunities for treatment. Nat. Rev. Endocrinol. 2021, 17, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Bruni, V.; Capozzi, A.; Lello, S. The Role of Genetics, Epigenetics and Lifestyle in Polycystic Ovary Syndrome Development: The State of the Art. Reprod. Sci. 2022, 29, 668–679. [Google Scholar] [CrossRef]
- Mimouni, N.E.H.; Paiva, I.; Barbotin, A.-L.; Timzoura, F.E.; Plassard, D.; Le Gras, S.; Ternier, G.; Pigny, P.; Catteau-Jonard, S.; Simon, V.; et al. Polycystic ovary syndrome is transmitted via a transgenerational epigenetic process. Cell Metab. 2021, 33, 513–530. [Google Scholar] [CrossRef]
- Schatten, H.; Sun, Q.-Y.; Prather, R. The impact of mitochondrial function/dysfunction on IVF and new treatment possibilities for infertility. Reprod. Biol. Endocrinol. 2014, 12, 111. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Moley, K.H. Maternal diabetes and oocyte quality. Mitochondrion 2010, 10, 403–410. [Google Scholar] [CrossRef] [Green Version]
- May-Panloup, P.; Boucret, L.; Chao de la Barca, J.-M.; Desquiret-Dumas, V.; Ferré-L’Hotellier, V.; Morinière, C.; Descamps, P.; Procaccio, V.; Reynier, P. Ovarian ageing: The role of mitochondria in oocytes and follicles. Hum. Reprod. Update 2016, 22, 725–743. [Google Scholar] [CrossRef] [Green Version]
- Craven, L.; Tang, M.-X.; Gorman, G.S.; De Sutter, P.; Heindryckx, B. Novel reproductive technologies to prevent mitochondrial disease. Hum. Reprod. Update 2017, 23, 501–519. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, M.; Amato, P.; Sparman, M.; Woodward, J.; Sanchis, D.M.; Ma, H.; Gutierrez, N.M.; Tippner-Hedges, R.; Kang, E.; Lee, H.-S.; et al. Towards germline gene therapy of inherited mitochondrial diseases. Nature 2013, 493, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, E.; Wu, J.; Gutierrez, N.M.; Koski, A.; Tippner-Hedges, R.; Agaronyan, K.; Platero-Luengo, A.; Martinez-Redondo, P.; Ma, H.; Lee, Y.; et al. Mitochondrial replacement in human oocytes carrying pathogenic mitochondrial DNA mutations. Nature 2016, 540, 270–275. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Luo, S.; Lu, Z.; Chávez-Badiola, A.; Liu, Z.; Yang, M.; Merhi, Z.; Silber, S.J.; Munné, S.; et al. Live birth derived from oocyte spindle transfer to prevent mitochondrial disease. Reprod. Biomed. Online 2017, 34, 361–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Paula, W.B.M.; Lucas, C.H.; Agip, A.-N.A.; Vizcay-Barrena, G.; Allen, J.F. Energy, ageing, fidelity and sex: Oocyte mitochondrial DNA as a protected genetic template. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyslop, L.A.; Blakeley, P.; Craven, L.; Richardson, J.; Fogarty, N.M.E.; Fragouli, E.; Lamb, M.; Wamaitha, S.E.; Prathalingam, N.; Zhang, Q.; et al. Towards clinical application of pronuclear transfer to prevent mitochondrial DNA disease. Nature 2016, 534, 383–386. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; O’Neil, R.C.; Marti Gutierrez, N.; Hariharan, M.; Zhang, Z.Z.; He, Y.; Cinnioglu, C.; Kayali, R.; Kang, E.; Lee, Y.; et al. Functional Human Oocytes Generated by Transfer of Polar Body Genomes. Cell Stem Cell 2017, 20, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.-P.; Lu, C.-F.; Gong, F.; Xie, P.-Y.; Hu, L.; Zhang, S.-J.; Lu, G.-X.; Lin, G. Polar body transfer restores the developmental potential of oocytes to blastocyst stage in a case of repeated embryo fragmentation. J. Assist. Reprod. Genet. 2017, 34, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Guo, Q.; Guo, H.; Lian, Y.; Zhai, F.; Yan, Z.; Long, C.; Zhu, P.; Tang, F.; Qiao, J.; et al. The methylome of a human polar body reflects that of its sibling oocyte and its aberrance may indicate poor embryo development. Hum. Reprod. 2021, 36, 318–330. [Google Scholar] [CrossRef]
- Takeo, S.; Sato, D.; Kimura, K.; Monji, Y.; Kuwayama, T.; Kawahara-miki, R.; Iwata, H. Resveratrol Improves the Mitochondrial Function and Fertilization Outcome of Bovine Oocytes. J. Reprod. Dev. 2014, 60, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, X.; Chen, S.; Wang, L.; Tan, Y.; Li, X.; Tang, L.; Zhang, J.; Wu, D.; Wu, Y.; et al. Comparison of Genome-Wide DNA Methylation Profiles of Human Fetal Tissues Conceived by in vitro Fertilization and Natural Conception. Front. Cell Dev. Biol. 2021, 9, 694769. [Google Scholar] [CrossRef]
- Yang, H.; Ma, Z.; Peng, L.; Kuhn, C.; Rahmeh, M.; Mahner, S.; Jeschke, U.; von Schönfeldt, V. Comparison of Histone H3K4me3 between IVF and ICSI Technologies and between Boy and Girl Offspring. Int. J. Mol. Sci. 2021, 22, 8574. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Lane, M.; Stevens, J.; Schoolcraft, W.B. Noninvasive assessment of human embryo nutrient consumption as a measure of developmental potential. Fertil. Steril. 2001, 76, 1175–1180. [Google Scholar] [CrossRef]
- Bolton, V.N.; Cutting, R.; Clarke, H.; Brison, D.R. ACE consensus meeting report: Culture systems. Hum. Fertil. 2014, 17, 239–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pöhland, R.; Souza-Cácares, M.B.; Datta, T.K.; Vanselow, J.; Martins, M.I.M.; da Silva, W.A.L.; Cardoso, C.J.T.; de Andrade Melo-Sterza, A. Influence of long-term thermal stress on the in vitro maturation on embryo development and Heat Shock Protein abundance in zebu cattle. Anim. Reprod. 2020, 17, e20190085. [Google Scholar] [CrossRef] [PubMed]
- Swain, J.E. Is there an optimal pH for culture media used in clinical IVF? Hum. Reprod. Update 2012, 18, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.; Haaf, T. Aberrant methylation patterns at the two-cell stage as an indicator of early developmental failure. Mol. Reprod. Dev. 2002, 63, 329–334. [Google Scholar] [CrossRef]
- Takamura, M.; Zhou, W.; Rombauts, L.; Dimitriadis, E. The long noncoding RNA PTENP1 regulates human endometrial epithelial adhesive capacity in vitro: Implications in infertility. Biol. Reprod. 2020, 102, 53–62. [Google Scholar] [CrossRef]
- Ibrahim, S.; Salilew-Wondim, D.; Rings, F.; Hoelker, M.; Neuhoff, C.; Tholen, E.; Looft, C.; Schellander, K.; Tesfaye, D. Expression Pattern of Inflammatory Response Genes and Their Regulatory MicroRNAs in Bovine Oviductal Cells in Response to Lipopolysaccharide: Implication for Early Embryonic Development. PLoS ONE 2015, 10, e0119388. [Google Scholar] [CrossRef] [Green Version]
- Barrera, A.D.; García, E.V.; Hamdi, M.; Sánchez-Calabuig, M.J.; López-Cardona, Á.P.; Balvís, N.F.; Rizos, D.; Gutiérrez-Adán, A. Embryo culture in presence of oviductal fluid induces DNA methylation changes in bovine blastocysts. Reproduction 2017, 154, 1–12. [Google Scholar] [CrossRef]
- Capalbo, A.; Ubaldi, F.M.; Cimadomo, D.; Noli, L.; Khalaf, Y.; Farcomeni, A.; Ilic, D.; Rienzi, L. MicroRNAs in spent blastocyst culture medium are derived from trophectoderm cells and can be explored for human embryo reproductive competence assessment. Fertil. Steril. 2016, 105, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Naillat, F.; Saadeh, H.; Nowacka-Woszuk, J.; Gahurova, L.; Santos, F.; Tomizawa, S.; Kelsey, G. Oxygen concentration affects de novo DNA methylation and transcription in in vitro cultured oocytes. Clin. Epigenet. 2021, 13, 132. [Google Scholar] [CrossRef] [PubMed]
- Fischer, B.; Bavister, B.D. Oxygen tension in the oviduct and uterus of rhesus monkeys, hamsters and rabbits. Reproduction 1993, 99, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Kasterstein, E.; Strassburger, D.; Komarovsky, D.; Bern, O.; Komsky, A.; Raziel, A.; Friedler, S.; Ron-El, R. The effect of two distinct levels of oxygen concentration on embryo development in a sibling oocyte study. J. Assist. Reprod. Genet. 2013, 30, 1073–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nastri, C.O.; Nóbrega, B.N.; Teixeira, D.M.; Amorim, J.; Diniz, L.M.M.; Barbosa, M.W.P.; Giorgi, V.S.I.; Pileggi, V.N.; Martins, W.P. Low versus atmospheric oxygen tension for embryo culture in assisted reproduction: A systematic review and meta-analysis. Fertil. Steril. 2016, 106, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Varela, C.; Labarta, E. Clinical Application of Antioxidants to Improve Human Oocyte Mitochondrial Function: A Review. Antioxidants 2020, 9, 1197. [Google Scholar] [CrossRef] [PubMed]
- Martín-Romero, F.J.; Miguel-Lasobras, E.M.; Domínguez-Arroyo, J.A.; González-Carrera, E.; Álvarez, I.S. Contribution of culture media to oxidative stress and its effect on human oocytes. Reprod. Biomed. Online 2008, 17, 652–661. [Google Scholar] [CrossRef]
- Menezo, Y.; Clément, P.; Dale, B. DNA methylation patterns in the early human embryo and the epigenetic/imprinting problems: A plea for a more careful approach to human assisted reproductive technology (ART). Int. J. Mol. Sci. 2019, 20, 1342. [Google Scholar] [CrossRef] [Green Version]
- Menezo, Y.; Clement, P.; Dale, B.; Elder, K. Modulating oxidative stress and epigenetic homeostasis in preimplantation IVF embryos. Zygote 2022, 30, 149–158. [Google Scholar] [CrossRef]
- Korsmo, H.W.; Jiang, X. One carbon metabolism and early development: A diet-dependent destiny. Trends Endocrinol. Metab. 2021, 32, 579–593. [Google Scholar] [CrossRef]
- Burgess, K.; Bennett, C.; Mosnier, H.; Kwatra, N.; Bethel, F.; Jadavji, N.M. The Antioxidant Role of One-Carbon Metabolism on Stroke. Antioxidants 2020, 9, 1141. [Google Scholar] [CrossRef]
- Clare, C.E.; Brassington, A.H.; Kwong, W.Y.; Sinclair, K.D. One-Carbon Metabolism: Linking Nutritional Biochemistry to Epigenetic Programming of Long-Term Development. Annu. Rev. Anim. Biosci. 2019, 7, 263–287. [Google Scholar] [CrossRef] [PubMed]
- Schachter, M. Insulin resistance in patients with polycystic ovary syndrome is associated with elevated plasma homocysteine. Hum. Reprod. 2003, 18, 721–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berker, B.; Kaya, C.; Aytac, R.; Satiroglu, H. Homocysteine concentrations in follicular fluid are associated with poor oocyte and embryo qualities in polycystic ovary syndrome patients undergoing assisted reproduction. Hum. Reprod. 2009, 24, 2293–2302. [Google Scholar] [CrossRef] [Green Version]
- Razi, Y.; Eftekhar, M.; Fesahat, F.; Dehghani Firouzabadi, R.; Razi, N.; Sabour, M.; Razi, M.H. Concentrations of homocysteine in follicular fluid and embryo quality and oocyte maturity in infertile women: A prospective cohort. J. Obstet. Gynaecol. 2021, 41, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Zeng, Y.; Hu, Y.; Liu, J.; Yin, C.; Niu, Y.; Wang, C.; Li, J.; Jia, Y.; Hong, J.; et al. Homocysteine impairs porcine oocyte quality via deregulation of one-carbon metabolism and hypermethylation of mitochondrial DNA. Biol. Reprod. 2019, 100, 907–916. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.A.; Ledger, W.; Edgar, D.H.; Sullivan, E.A. Clinical outcomes following cryopreservation of blastocysts by vitrification or slow freezing: A population-based cohort study. Hum. Reprod. 2014, 29, 2794–2801. [Google Scholar] [CrossRef] [Green Version]
- Fasano, G.; Fontenelle, N.; Vannin, A.-S.; Biramane, J.; Devreker, F.; Englert, Y.; Delbaere, A. A randomized controlled trial comparing two vitrification methods versus slow-freezing for cryopreservation of human cleavage stage embryos. J. Assist. Reprod. Genet. 2014, 31, 241–247. [Google Scholar] [CrossRef]
- Zhao, X.-M.; Du, W.-H.; Wang, D.; Hao, H.-S.; Liu, Y.; Qin, T.; Zhu, H.-B. Effect of cyclosporine pretreatment on mitochondrial function in vitrified bovine mature oocytes. Fertil. Steril. 2011, 95, 2786–2788. [Google Scholar] [CrossRef]
- Wang, N.; Hao, H.-S.; Li, C.-Y.; Zhao, Y.-H.; Wang, H.-Y.; Yan, C.-L.; Du, W.-H.; Wang, D.; Liu, Y.; Pang, Y.-W.; et al. Calcium ion regulation by BAPTA-AM and ruthenium red improved the fertilisation capacity and developmental ability of vitrified bovine oocytes. Sci. Rep. 2017, 7, 10652. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Geng, L.; Huang, R.; Peng, W.; Chen, X.; Jiang, X.; Yu, M.; Li, M.; Huang, Y.; Yang, X. Effect of vitrification on in vitro development and imprinted gene Grb10 in mouse embryos. Reproduction 2017, 154, 197–205. [Google Scholar] [CrossRef]
- Saenz-de-Juano, M.D.; Billooye, K.; Smitz, J.; Anckaert, E. The loss of imprinted DNA methylation in mouse blastocysts is inflicted to a similar extent by in vitro follicle culture and ovulation induction. Mol. Hum. Reprod. 2016, 22, 427–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi, A.; Naderi, M.M.; Hassanpour, H.; Heidari, M.; Borjian, S.; Sarvari, A.; Akhondi, M.M. The effect of ovine oocyte vitrification on expression of subset of genes involved in epigenetic modifications during oocyte maturation and early embryo development. Theriogenology 2016, 86, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Liu, C.; Zhang, M.; Wang, X.; Yan, Y.; Liu, Q.; Ma, Y.; Yu, T.; Sathanawongs, A.; Jiao, J.; et al. Vitrification of Pronuclear Zygotes Perturbs Porcine Zygotic Genome Activation. Animals 2022, 12, 610. [Google Scholar] [CrossRef] [PubMed]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO induces drastic changes in human cellular processes and epigenetic landscape in vitro. Sci. Rep. 2019, 9, 4641. [Google Scholar] [CrossRef] [Green Version]
- Sales, V.M.; Ferguson-Smith, A.C.; Patti, M.-E. Epigenetic Mechanisms of Transmission of Metabolic Disease across Generations. Cell Metab. 2017, 25, 559–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathare, A.D.S.; Hinduja, I. Aberrant DNA methylation profiling affecting the endometrial receptivity in recurrent implantation failure patients undergoing in vitro fertilization. Am. J. Reprod. Immunol. 2020, 83, e13196. [Google Scholar] [CrossRef]
- Jiang, N.-X.; Li, X.-L. The Complicated Effects of Extracellular Vesicles and Their Cargos on Embryo Implantation. Front. Endocrinol. 2021, 12, 681266. [Google Scholar] [CrossRef]
- Chen, C.-W.; Huang, R.-L.; Do, A.Q.; Wang, H.-C.; Lee, Y.-X.; Wang, C.-W.; Hsieh, C.-C.; Tzeng, C.-R.; Hu, Y.-M.; Chen, C.-H.; et al. Genome-wide analysis of cervical secretions obtained during embryo transfer reveals the association between deoxyribonucleic acid methylation and pregnancy outcomes. F&S Sci. 2022, 3, 74–83. [Google Scholar] [CrossRef]
- Qu, X.; Fang, Y.; Zhuang, S.; Zhang, Y. Micro-RNA miR-542-3p suppresses decidualization by targeting ILK pathways in human endometrial stromal cells. Sci. Rep. 2021, 11, 7186. [Google Scholar] [CrossRef]
- Zhang, Y.-P.; Huang, Y.-T.; Huang, T.-S.; Pang, W.; Zhu, J.-J.; Liu, Y.-F.; Tang, R.-Z.; Zhao, C.-R.; Yao, W.-J.; Li, Y.-S.; et al. The Mammalian Target of Rapamycin and DNA methyltransferase 1 axis mediates vascular endothelial dysfunction in response to disturbed flow. Sci. Rep. 2017, 7, 14996. [Google Scholar] [CrossRef] [Green Version]
- Jing, M.; Rech, L.; Wu, Y.; Goltz, D.; Taylor, C.G.; House, J.D. Effects of zinc deficiency and zinc supplementation on homocysteine levels and related enzyme expression in rats. J. Trace Elem. Med. Biol. 2015, 30, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Friso, S.; Udali, S.; De Santis, D.; Choi, S.-W. One-carbon metabolism and epigenetics. Mol. Aspects Med. 2017, 54, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ben-Sahra, I.; Hoxhaj, G.; Ricoult, S.J.H.; Asara, J.M.; Manning, B.D. mTORC1 induces purine synthesis through control of the mitochondrial tetrahydrofolate cycle. Science 2016, 351, 728–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansson, N.; Rosario, F.J.; Gaccioli, F.; Lager, S.; Jones, H.N.; Roos, S.; Jansson, T.; Powell, T.L. Activation of Placental mTOR Signaling and Amino Acid Transporters in Obese Women Giving Birth to Large Babies. J. Clin. Endocrinol. Metab. 2013, 98, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Lv, X.; Zhang, C.; Du, W.; Liu, Y.; Zhu, L.; Hao, J. DNMT1, a Novel Regulator Mediating mTORC1/mTORC2 Pathway-Induced NGF Expression in Schwann Cells. Neurochem. Res. 2018, 43, 2141–2154. [Google Scholar] [CrossRef]
- Zeng, H.; Yan, L.; Cheng, W.-H.; Uthus, E.O. Dietary Selenomethionine Increases Exon-Specific DNA Methylation of the p53 Gene in Rat Liver and Colon Mucosa. J. Nutr. 2011, 141, 1464–1468. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Lu, L.; Liao, X.; Li, W.; Zhang, L.; Ji, C.; Lin, X.; Liu, H.-C.; Odle, J.; Luo, X. Maternal dietary manganese protects chick embryos against maternal heat stress via epigenetic-activated antioxidant and anti-apoptotic abilities. Oncotarget 2017, 8, 89665–89680. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, E.; Everson, T.M.; Punshon, T.; Jackson, B.P.; Hao, K.; Lambertini, L.; Chen, J.; Karagas, M.R.; Marsit, C.J. Copper associates with differential methylation in placentae from two US birth cohorts. Epigenetics 2020, 15, 215–230. [Google Scholar] [CrossRef]
- Tobi, E.W.; Slieker, R.C.; Luijk, R.; Dekkers, K.F.; Stein, A.D.; Xu, K.M.; Slagboom, P.E.; van Zwet, E.W.; Lumey, L.H.; Heijmans, B.T. DNA methylation as a mediator of the association between prenatal adversity and risk factors for metabolic disease in adulthood. Sci. Adv. 2018, 4, eaao4364. [Google Scholar] [CrossRef] [Green Version]
- Tobi, E.W.; Lumey, L.H.; Talens, R.P.; Kremer, D.; Putter, H.; Stein, A.D.; Slagboom, P.E.; Heijmans, B.T. DNA methylation differences after exposure to prenatal famine are common and timing- and sex-specific. Hum. Mol. Genet. 2009, 18, 4046–4053. [Google Scholar] [CrossRef]
- Schutt, A.K.; Blesson, C.S.; Hsu, J.W.; Valdes, C.T.; Gibbons, W.E.; Jahoor, F.; Yallampalli, C. Preovulatory exposure to a protein-restricted diet disrupts amino acid kinetics and alters mitochondrial structure and function in the rat oocyte and is partially rescued by folic acid. Reprod. Biol. Endocrinol. 2019, 17, 12. [Google Scholar] [CrossRef]
- Dahlhoff, M.; Pfister, S.; Blutke, A.; Rozman, J.; Klingenspor, M.; Deutsch, M.J.; Rathkolb, B.; Fink, B.; Gimpfl, M.; Hrabě de Angelis, M.; et al. Peri-conceptional obesogenic exposure induces sex-specific programming of disease susceptibilities in adult mouse offspring. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 304–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Doherty, A.M.; O’Gorman, A.; al Naib, A.; Brennan, L.; Daly, E.; Duffy, P.; Fair, T. Negative energy balance affects imprint stability in oocytes recovered from postpartum dairy cows. Genomics 2014, 104, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Fair, T. DNA methylation dynamics during oocyte and embryo development and its association with environmental induced alterations. Anim. Reprod. 2016, 13, 250–256. [Google Scholar] [CrossRef]
- Desai, M.; Jellyman, J.K.; Han, G.; Beall, M.; Lane, R.H.; Ross, M.G. Maternal obesity and high-fat diet program offspring metabolic syndrome. Am. J. Obstet. Gynecol. 2014, 211, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansson, N.; Nilsfelt, A.; Gellerstedt, M.; Wennergren, M.; Rossander-Hultheén, L.; Powell, T.L.; Jansson, T. Maternal hormones linking maternal body mass index and dietary intake to birth weight. Am. J. Clin. Nutr. 2008, 87, 1743–1749. [Google Scholar] [CrossRef] [PubMed]
- Morales, E.; Groom, A.; Lawlor, D.A.; Relton, C.L. DNA methylation signatures in cord blood associated with maternal gestational weight gain: Results from the ALSPAC cohort. BMC Res. Notes 2014, 7, 278. [Google Scholar] [CrossRef] [Green Version]
- Igosheva, N.; Abramov, A.Y.; Poston, L.; Eckert, J.J.; Fleming, T.P.; Duchen, M.R.; McConnell, J. Maternal Diet-Induced Obesity Alters Mitochondrial Activity and Redox Status in Mouse Oocytes and Zygotes. PLoS ONE 2010, 5, e10074. [Google Scholar] [CrossRef] [Green Version]
- Aiken, C.E.; Tarry-Adkins, J.L.; Penfold, N.C.; Dearden, L.; Ozanne, S.E. Decreased ovarian reserve, dysregulation of mitochondrial biogenesis, and increased lipid peroxidation in female mouse offspring exposed to an obesogenic maternal diet. FASEB J. 2016, 30, 1548–1556. [Google Scholar] [CrossRef] [Green Version]
- Gemma, C.; Sookoian, S.; Alvariñas, J.; García, S.I.; Quintana, L.; Kanevsky, D.; González, C.D.; Pirola, C.J. Maternal Pregestational BMI Is Associated With Methylation of the PPARGC1A Promoter in Newborns. Obesity 2009, 17, 1032–1039. [Google Scholar] [CrossRef]
- Hoyme, H.E.; May, P.A.; Kalberg, W.O.; Kodituwakku, P.; Gossage, J.P.; Trujillo, P.M.; Buckley, D.G.; Miller, J.H.; Aragon, A.S.; Khaole, N.; et al. A Practical Clinical Approach to Diagnosis of Fetal Alcohol Spectrum Disorders: Clarification of the 1996 Institute of Medicine Criteria. Pediatrics 2005, 115, 39–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halsted, C.H.; Medici, V. Aberrant Hepatic Methionine Metabolism and Gene Methylation in the Pathogenesis and Treatment of Alcoholic Steatohepatitis. Int. J. Hepatol. 2012, 2012, 959746. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Pan, C.-H.; Chen, C.-C.; Huang, M.-C. Increased Oxidative DNA Damage in Patients With Alcohol Dependence and Its Correlation With Alcohol Withdrawal Severity. Alcohol. Clin. Exp. Res. 2011, 35, 338–344. [Google Scholar] [CrossRef]
- Ungerer, M.; Knezovich, J.; Ramsay, M. In utero alcohol exposure, epigenetic changes, and their consequences. Alcohol Res. 2013, 35, 37–46. [Google Scholar] [PubMed]
- Wang, D.; Jacobs, S.A.; Tsien, J.Z. Targeting the NMDA receptor subunit NR2B for treating or preventing age-related memory decline. Expert Opin. Ther. Targets 2014, 18, 1121–1130. [Google Scholar] [CrossRef]
- Qiang, M.; Denny, A.; Chen, J.; Ticku, M.K.; Yan, B.; Henderson, G. The site specific demethylation in the 5’-regulatory area of NMDA receptor 2B subunit gene associated with CIE-induced up-regulation of transcription. PLoS ONE 2010, 5, e8798. [Google Scholar] [CrossRef]
- Carpenter, B.L.; Remba, T.K.; Thomas, S.L.; Madaj, Z.; Brink, L.; Tiedemann, R.L.; Odendaal, H.J.; Jones, P.A. Oocyte age and preconceptual alcohol use are highly correlated with epigenetic imprinting of a noncoding RNA (nc886). Proc. Natl. Acad. Sci. USA 2021, 118, e2026580118. [Google Scholar] [CrossRef]
- Haycock, P.C.; Ramsay, M. Exposure of Mouse Embryos to Ethanol During Preimplantation Development: Effect on DNA Methylation in the H19 Imprinting Control Region1. Biol. Reprod. 2009, 81, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Balaraman, Y.; Wang, G.; Nephew, K.P.; Zhou, F.C. Alcohol exposure alters DNA methylation profiles in mouse embryos at early neurulation. Epigenetics 2009, 4, 500–511. [Google Scholar] [CrossRef] [Green Version]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [Green Version]
- Geraghty, A.A.; Lindsay, K.L.; Alberdi, G.; McAuliffe, F.M.; Gibney, E.R. Nutrition during Pregnancy Impacts Offspring’s Epigenetic Status—Evidence from Human and Animal Studies. Nutr. Metab. Insights 2015, 8, S29527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jousse, C.; Parry, L.; Lambert-Langlais, S.; Maurin, A.; Averous, J.; Bruhat, A.; Carraro, V.; Tost, J.; Letteron, P.; Chen, P.; et al. Perinatal undernutrition affects the methylation and expression of the leptin gene in adults: Implication for the understanding of metabolic syndrome. FASEB J. 2011, 25, 3271–3278. [Google Scholar] [CrossRef] [PubMed]
- Joubert, B.R.; Felix, J.F.; Yousefi, P.; Bakulski, K.M.; Just, A.C.; Breton, C.; Reese, S.E.; Markunas, C.A.; Richmond, R.C.; Xu, C.-J.; et al. DNA Methylation in Newborns and Maternal Smoking in Pregnancy: Genome-wide Consortium Meta-analysis. Am. J. Hum. Genet. 2016, 98, 680–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steuerwald, N.M.; Bermúdez, M.G.; Wells, D.; Munné, S.; Cohen, J. Maternal age-related differential global expression profiles observed in human oocytes. Reprod. Biomed. Online 2007, 14, 700–708. [Google Scholar] [CrossRef]
- Tsutsumi, M.; Fujiwara, R.; Nishizawa, H.; Ito, M.; Kogo, H.; Inagaki, H.; Ohye, T.; Kato, T.; Fujii, T.; Kurahashi, H. Age-Related Decrease of Meiotic Cohesins in Human Oocytes. PLoS ONE 2014, 9, e96710. [Google Scholar] [CrossRef] [Green Version]
- Ntostis, P.; Iles, D.; Kokkali, G.; Vaxevanoglou, T.; Kanavakis, E.; Pantou, A.; Huntriss, J.; Pantos, K.; Picton, H.M. The impact of maternal age on gene expression during the GV to MII transition in euploid human oocytes. Hum. Reprod. 2021, 37, 80–92. [Google Scholar] [CrossRef]
- Battaglia, R.; Vento, M.E.; Ragusa, M.; Barbagallo, D.; La Ferlita, A.; Di Emidio, G.; Borzi, P.; Artini, P.G.; Scollo, P.; Tatone, C.; et al. MicroRNAs Are Stored in Human MII Oocyte and Their Expression Profile Changes in Reproductive Aging. Biol. Reprod. 2016, 95, 131. [Google Scholar] [CrossRef]
- Hamatani, T.; Falco, G.; Carter, M.G.; Akutsu, H.; Stagg, C.A.; Sharov, A.A.; Dudekula, D.B.; VanBuren, V.; Ko, M.S.H. Age-associated alteration of gene expression patterns in mouse oocytes. Hum. Mol. Genet. 2004, 13, 2263–2278. [Google Scholar] [CrossRef] [Green Version]
- Monk, C.; Feng, T.; Lee, S.; Krupska, I.; Champagne, F.A.; Tycko, B. Distress During Pregnancy: Epigenetic Regulation of Placenta Glucocorticoid-Related Genes and Fetal Neurobehavior. Am. J. Psychiatry 2016, 173, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Mulligan, C.; D’Errico, N.; Stees, J.; Hughes, D. Methylation changes at NR3C1 in newborns associate with maternal prenatal stress exposure and newborn birth weight. Epigenetics 2012, 7, 853–857. [Google Scholar] [CrossRef] [Green Version]
- Hjort, L.; Martino, D.; Grunnet, L.G.; Naeem, H.; Maksimovic, J.; Olsson, A.H.; Zhang, C.; Ling, C.; Olsen, S.F.; Saffery, R.; et al. Gestational diabetes and maternal obesity are associated with epigenome-wide methylation changes in children. JCI Insight 2018, 3, e122572. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Li, J.; He, B.; Jia, Y.; Niu, Y.; Wang, C.; Zhao, R. Abnormally activated one-carbon metabolic pathway is associated with mtDNA hypermethylation and mitochondrial malfunction in the oocytes of polycystic gilt ovaries. Sci. Rep. 2016, 6, 19436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junge, K.M.; Leppert, B.; Jahreis, S.; Wissenbach, D.K.; Feltens, R.; Grützmann, K.; Thürmann, L.; Bauer, T.; Ishaque, N.; Schick, M.; et al. MEST mediates the impact of prenatal bisphenol A exposure on long-term body weight development. Clin. Epigenet. 2018, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Susiarjo, M.; Sasson, I.; Mesaros, C.; Bartolomei, M.S. Bisphenol A Exposure Disrupts Genomic Imprinting in the Mouse. PLoS Genet. 2013, 9, e1003401. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Jeon, H.-J.; Choi, D.Y.; Oh, J.S. Polystyrene nanoparticles incorporate into the endoplasmic reticulum and disturb translation during meiotic maturation in mouse oocytes. Toxicol. In Vitro 2022, 82, 105380. [Google Scholar] [CrossRef] [PubMed]
- Llonch, S.; Barragán, M.; Nieto, P.; Mallol, A.; Elosua-Bayes, M.; Lorden, P.; Ruiz, S.; Zambelli, F.; Heyn, H.; Vassena, R.; et al. Single human oocyte transcriptome analysis reveals distinct maturation stage-dependent pathways impacted by age. Aging Cell 2021, 20, e13360. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, O.; Michaeli, M.; Aslih, N.; Poltov, D.; Estrada, D.; Atzmon, Y.; Shalom-Paz, E. Embryonic Development in Relation to Maternal Age and Conception Probability. Reprod. Sci. 2021, 28, 2292–2300. [Google Scholar] [CrossRef]
- Hassold, T.; Hunt, P. Maternal age and chromosomally abnormal pregnancies: What we know and what we wish we knew. Curr. Opin. Pediatr. 2009, 21, 703–708. [Google Scholar] [CrossRef] [Green Version]
- Sanders, K.D.; Silvestri, G.; Gordon, T.; Griffin, D.K. Analysis of IVF live birth outcomes with and without preimplantation genetic testing for aneuploidy (PGT-A): UK Human Fertilisation and Embryology Authority data collection 2016–2018. J. Assist. Reprod. Genet. 2021, 38, 3277–3285. [Google Scholar] [CrossRef]
- Ge, Z.-J.J.; Schatten, H.; Zhang, C.-L.L.; Sun, Q.-Y.Y. Oocyte ageing and epigenetics. Reproduction 2015, 149, R103–R114. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.-Y.; Li, S.; Chen, L.-N.; Schatten, H.; Ou, X.-H.; Sun, Q.-Y. Why is oocyte aneuploidy increased with maternal aging? J. Genet. Genom. 2020, 47, 659–671. [Google Scholar] [CrossRef]
- Yue, M.X.; Fu, X.W.; Zhou, G.B.; Hou, Y.P.; Du, M.; Wang, L.; Zhu, S.E. Abnormal DNA methylation in oocytes could be associated with a decrease in reproductive potential in old mice. J. Assist. Reprod. Genet. 2012, 29, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimadomo, D.; Fabozzi, G.; Vaiarelli, A.; Ubaldi, N.; Ubaldi, F.M.; Rienzi, L. Impact of Maternal Age on Oocyte and Embryo Competence. Front. Endocrinol. 2018, 9, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, A.; François, O.; Lepeule, J. Epigenetic Alterations of Maternal Tobacco Smoking during Pregnancy: A Narrative Review. Int. J. Environ. Res. Public Health 2021, 18, 5083. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.K.; Adigun, A.; Huang, Z.; Overcash, F.; Wang, F.; Jirtle, R.L.; Schildkraut, J.M.; Murtha, A.P.; Iversen, E.S.; Hoyo, C. Gender-specific methylation differences in relation to prenatal exposure to cigarette smoke. Gene 2012, 494, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Wiklund, P.; Karhunen, V.; Richmond, R.C.; Parmar, P.; Rodriguez, A.; De Silva, M.; Wielscher, M.; Rezwan, F.I.; Richardson, T.G.; Veijola, J.; et al. DNA methylation links prenatal smoking exposure to later life health outcomes in offspring. Clin. Epigenet. 2019, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Joubert, B.R.; Håberg, S.E.; Nilsen, R.M.; Wang, X.; Vollset, S.E.; Murphy, S.K.; Huang, Z.; Hoyo, C.; Midttun, Ø.; Cupul-Uicab, L.A.; et al. 450K Epigenome-Wide Scan Identifies Differential DNA Methylation in Newborns Related to Maternal Smoking during Pregnancy. Environ. Health Perspect. 2012, 120, 1425–1431. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Du, Y.-R.; Zhu, H.; Sun, M.-L.; Wang, C.; Cheng, Y.; Pang, H.; Ding, G.; Gao, J.; Tan, Y.; et al. Maternal inheritance of glucose intolerance via oocyte TET3 insufficiency. Nature 2022, 605, 761–766. [Google Scholar] [CrossRef]
- Gore, A.C. Endocrine-Disrupting Chemicals. JAMA Intern. Med. 2016, 176, 1705. [Google Scholar] [CrossRef]
- Prusinski, L.; Al-Hendy, A.; Yang, Q. Developmental Exposure to Endocrine Disrupting Chemicals Alters the Epigenome: Identification of Reprogrammed Targets. Gynecol. Obstet. Res. Open J. 2016, 3, 1–6. [Google Scholar] [CrossRef]
- McCabe, C.F.; Padmanabhan, V.; Dolinoy, D.C.; Domino, S.E.; Jones, T.R.; Bakulski, K.M.; Goodrich, J.M. Maternal environmental exposure to bisphenols and epigenome-wide DNA methylation in infant cord blood. Environ. Epigenet. 2020, 6, dvaa021. [Google Scholar] [CrossRef]
- Bromer, J.G.; Zhou, Y.; Taylor, M.B.; Doherty, L.; Taylor, H.S. Bisphenol-A exposure in utero leads to epigenetic alterations in the developmental programming of uterine estrogen response. FASEB J. 2010, 24, 2273–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobel, C.J.; Goldstein, A.; Barrett, E.S. Psychosocial Stress and Pregnancy Outcome. Clin. Obstet. Gynecol. 2008, 51, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Hompes, T.; Izzi, B.; Gellens, E.; Morreels, M.; Fieuws, S.; Pexsters, A.; Schops, G.; Dom, M.; Van Bree, R.; Freson, K.; et al. Investigating the influence of maternal cortisol and emotional state during pregnancy on the DNA methylation status of the glucocorticoid receptor gene (NR3C1) promoter region in cord blood. J. Psychiatr. Res. 2013, 47, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Kertes, D.A.; Kamin, H.S.; Hughes, D.A.; Rodney, N.C.; Bhatt, S.; Mulligan, C.J. Prenatal Maternal Stress Predicts Methylation of Genes Regulating the Hypothalamic–Pituitary–Adrenocortical System in Mothers and Newborns in the Democratic Republic of Congo. Child. Dev. 2016, 87, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Epigenetic modifications: This diagram shows the complexity of epigenetic processes, which are divided into the following subgroups: DNA modifications—5-methylcytosine (5mC) DNA methylation, 5-hydroxymethylcytosine (5hmC) DNA methylation; Histone modifications—Acetylation, Methylation, Phosphorylation, Poly-ADP ribosylation, Ubiquitination; Non-coding RNA interactions—piwi RNA (piRNA), small interfering RNA (siRNA), long non-coding RNA (lncRNA), micro RNA (miRNA); RNA modifications—6-methyladenosine (6mA) RNA methylation, 5-methylcytosine (5mC) RNA methylation, 7-methylguanosine (7mG) RNA methylation, mRNA CAP, 5-hydroxymethylcytosine (5hmC) RNA methylation. The image was created with BioRender.com.
Figure 1.
Epigenetic modifications: This diagram shows the complexity of epigenetic processes, which are divided into the following subgroups: DNA modifications—5-methylcytosine (5mC) DNA methylation, 5-hydroxymethylcytosine (5hmC) DNA methylation; Histone modifications—Acetylation, Methylation, Phosphorylation, Poly-ADP ribosylation, Ubiquitination; Non-coding RNA interactions—piwi RNA (piRNA), small interfering RNA (siRNA), long non-coding RNA (lncRNA), micro RNA (miRNA); RNA modifications—6-methyladenosine (6mA) RNA methylation, 5-methylcytosine (5mC) RNA methylation, 7-methylguanosine (7mG) RNA methylation, mRNA CAP, 5-hydroxymethylcytosine (5hmC) RNA methylation. The image was created with BioRender.com.

Figure 2.
Dynamic DNA methylation and protein translation changes in human oogenesis and early embryogenesis: (A) Prenatal DNA demethylation in primordial germ cells (PGC) is followed by de novo DNA methylation, which occurs earlier in males than females. Genomic imprinting loci consisting of DMR (differentially methylated regions) maintain their methylation status despite the important genome-wide DNA demethylation in pre-implantation embryos [1]. (B) Oocytes are transcriptionally active during oocyte growth with rapid decline and silencing throughout meiotic maturation [2]. According to the new evidence human embryonic genome activation is initiated at the zygotic stage, but the transcriptional activity remains low until the 8-cell stage [3]. Correct regulation of CAP-dependent translation is a key process in meiotic maturation despite continuous decrease in global translational activity from oocytes to zygotes. The image was created with BioRender.com.
Figure 2.
Dynamic DNA methylation and protein translation changes in human oogenesis and early embryogenesis: (A) Prenatal DNA demethylation in primordial germ cells (PGC) is followed by de novo DNA methylation, which occurs earlier in males than females. Genomic imprinting loci consisting of DMR (differentially methylated regions) maintain their methylation status despite the important genome-wide DNA demethylation in pre-implantation embryos [1]. (B) Oocytes are transcriptionally active during oocyte growth with rapid decline and silencing throughout meiotic maturation [2]. According to the new evidence human embryonic genome activation is initiated at the zygotic stage, but the transcriptional activity remains low until the 8-cell stage [3]. Correct regulation of CAP-dependent translation is a key process in meiotic maturation despite continuous decrease in global translational activity from oocytes to zygotes. The image was created with BioRender.com.

Figure 3.
Assisted reproduction technology (ART) and external factors involved in epigenetic alterations: visualization of ART procedures employed in the process of oocyte meiotic maturation and early embryo development with proven (red arrows) or insignificant (pale red arrows) impact on the epigenome. External factors influencing epigenetics of post-implantation in utero embryo and fetal development (yellow arrows) are divided among nutritional (alcohol, diet) and lifestyle factors (advanced age, smoking, living environment, endocrine-disrupting chemicals (EDCs)). The image was created with BioRender.com.
Figure 3.
Assisted reproduction technology (ART) and external factors involved in epigenetic alterations: visualization of ART procedures employed in the process of oocyte meiotic maturation and early embryo development with proven (red arrows) or insignificant (pale red arrows) impact on the epigenome. External factors influencing epigenetics of post-implantation in utero embryo and fetal development (yellow arrows) are divided among nutritional (alcohol, diet) and lifestyle factors (advanced age, smoking, living environment, endocrine-disrupting chemicals (EDCs)). The image was created with BioRender.com.

Figure 4.
Transgenerational epigenetic inheritance and the therapy (the polycystic ovary syndrome (PCOS) case: Recent evidence showed that acquired epigenetic changes can be inherited between generations. One such example is the PCOS, which presents the main cause of infertility in humans. PCOS patients are usually managed by assisted reproduction techniques (ART), but there is high probability of transmission from mother to daughter. DNA hypomethylation in PCOS is also influenced by external factors such as nutrition, living environment, or lifestyle [205]. The nutritional supplementation with one carbon metabolism (OCM) compound, the methyl donor S-adenosylmethionine (SAM) could possibly have a therapeutic potential to mitigate or alleviate PCOS in humans [206]. The image was created with BioRender.com.
Figure 4.
Transgenerational epigenetic inheritance and the therapy (the polycystic ovary syndrome (PCOS) case: Recent evidence showed that acquired epigenetic changes can be inherited between generations. One such example is the PCOS, which presents the main cause of infertility in humans. PCOS patients are usually managed by assisted reproduction techniques (ART), but there is high probability of transmission from mother to daughter. DNA hypomethylation in PCOS is also influenced by external factors such as nutrition, living environment, or lifestyle [205]. The nutritional supplementation with one carbon metabolism (OCM) compound, the methyl donor S-adenosylmethionine (SAM) could possibly have a therapeutic potential to mitigate or alleviate PCOS in humans [206]. The image was created with BioRender.com.

Figure 5.
One-carbon metabolism (OCM)-associated pathways in nutrition and epigenetics: Many OCM compounds are nutritional supplements (green) such as B vitamins; Riboflavin (B2), Pyridoxine (B6), Folate (B9), or Cobalamin (B12). 5-methyl-tetrahydrofolate (5-mTHF) or methionine and betaine are another currently introduced nutritional supplements. OCM pathways produce molecules for epigenetic modifications [239], antioxidant production [240], and building compounds of nucleic acids, amino acids, or phospholipids (orange) [241], which are all together influencing in utero fetal development (purple). Abbreviations: 5,10-mTHF, 5,10-methenyl-tetrahydrofolate; 10-fTHF, 10-formyl-tetrahydrofolate; BHMT, betaine-homocysteine S-methyltransferase; CBS, cystathionine β-synthase; DHF, dihydrofolate; DHFR, dihydrofolate reductase; DMG, dimethylglycine; DNMTs, de novo and maintenance DNA methyltransferases; dTMP, deoxythymidine monophosphate; dUMP, deoxyuridine monophosphate; FA, fatty acids; GSH, reduced glutathione; MTHFR, 5,10-methylenetetrahydrofolate reductase; SAH, S-adenosylhomocysteine; SAHH, S-adenosylhomocysteine hydrolase; SAM, S-adenosylmethionine; SHMT, serine hydroxymethyltransferase; THF, tetrahydrofolate; VLDL, very-low density lipoprotein. The image was created with BioRender.com.
Figure 5.
One-carbon metabolism (OCM)-associated pathways in nutrition and epigenetics: Many OCM compounds are nutritional supplements (green) such as B vitamins; Riboflavin (B2), Pyridoxine (B6), Folate (B9), or Cobalamin (B12). 5-methyl-tetrahydrofolate (5-mTHF) or methionine and betaine are another currently introduced nutritional supplements. OCM pathways produce molecules for epigenetic modifications [239], antioxidant production [240], and building compounds of nucleic acids, amino acids, or phospholipids (orange) [241], which are all together influencing in utero fetal development (purple). Abbreviations: 5,10-mTHF, 5,10-methenyl-tetrahydrofolate; 10-fTHF, 10-formyl-tetrahydrofolate; BHMT, betaine-homocysteine S-methyltransferase; CBS, cystathionine β-synthase; DHF, dihydrofolate; DHFR, dihydrofolate reductase; DMG, dimethylglycine; DNMTs, de novo and maintenance DNA methyltransferases; dTMP, deoxythymidine monophosphate; dUMP, deoxyuridine monophosphate; FA, fatty acids; GSH, reduced glutathione; MTHFR, 5,10-methylenetetrahydrofolate reductase; SAH, S-adenosylhomocysteine; SAHH, S-adenosylhomocysteine hydrolase; SAM, S-adenosylmethionine; SHMT, serine hydroxymethyltransferase; THF, tetrahydrofolate; VLDL, very-low density lipoprotein. The image was created with BioRender.com.


{kind=link}
{kind=link}
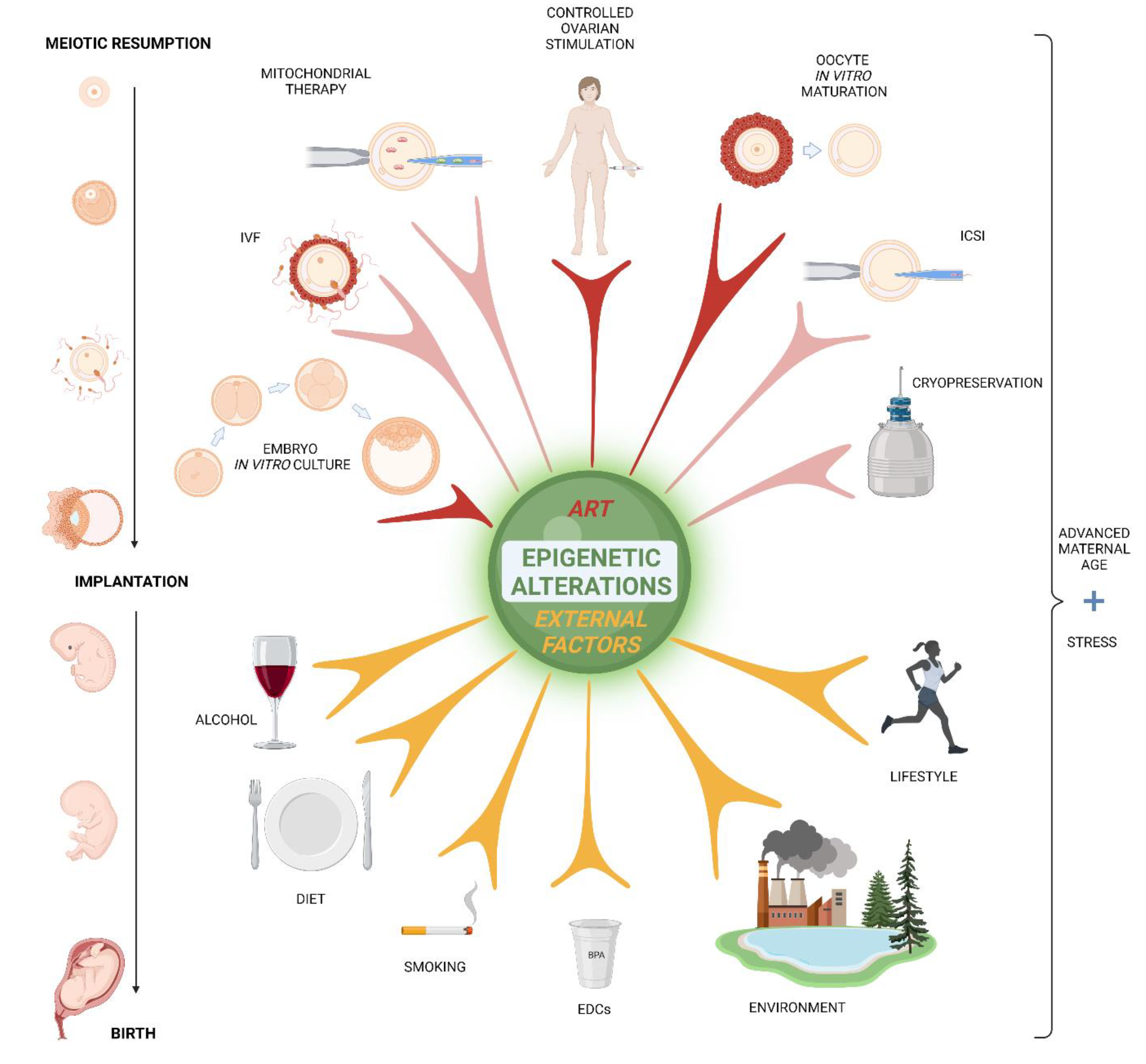{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of effects induced by ART on epigenetic changes in oocytes, early embryos and their influence on offspring. “NE” indicates specific gene effects were not evaluated.
Table 1.
Summary of effects induced by ART on epigenetic changes in oocytes, early embryos and their influence on offspring. “NE” indicates specific gene effects were not evaluated.
| Stressor | Species | Genes Affected | Main Findings | Reference |
|---|---|---|---|---|
| Ovarian Stimulation | ||||
| Controlled ovarian stimulation | human | NE | Chromosomal aneuploidy | [168] |
| Superovulation | mouse | Dnmt1, Dnmt3A, Dnmt3B | Affected expressin of methyltransferases in GV, MII oocytes, in one-cell and two-cell embryos | [169] |
| Superovulation | mouse | Epab, Pabc1 | Altered expression of translational regulators mRNA in mouse GV and MII oocytes and in zygots | [170] |
| Superovulation | mouse | Snrpn, Peg3, Kcnq1ot1, H19 | Disrupted methylation of imprinted genes in blastocysts | [171] |
| Superovulation | mouse | Gfod2, Foxi3, Celf4, Syf2 | In oocytes, altered methylation of genes involved in glucose metabolism, nervous system development, cell cycle, cell proliferation, and mRNA processing | [172] |
| Superovulation | mouse | H19 | Altered H19 methylation in mouse blastocysts after in vivo fertilization | [173] |
| Superovulation | mouse | Fasn, Dgat1, Dgat2 | Decreased fatty acid content in mice 2-cell embryos by reducing the Fasn and increasing the Dgat1 and Dgat2 expression. | [174] |
| Repeated superovulation | mouse | Cox1, Cytb, Nd2, Nd4 | Altered expression of mitochondrial genes in mouse cumulus cells | [175] |
| Repeated superovulation | mouse | NE | Abnormalities in mitochondrial structure and distribution in mouse oocytes | [176] |
| Superovulation | mouse | NE | Decrease of mitochondrial activity and ATP production in mouse oocytes | [177] |
| Superovulation | bovine | TXN2, PDX3 | Decline of mtDNA copy number in bovine oocytes., decreased expression of antioxidant genes in bovine cumulus cells | [93] |
| Oxidative stress | ||||
| Presence of reactive oxygen species | human | NE | Sperm originated changes to epigenetic regulation of human embryo development | [178] |
| Culture under 20% of oxygen | bovine | CAT, GLRX2, HSP90AA1. KEAP1, NFR2, PRDX1, PRDX3, SOD1, TXN, TXNRD1, H2AFZ, H3F3B | Increase of transcript of genes associated with epigenetic remodelling, oxidative stress and cellular stress response in blastocysts | [179] |
| Culture under 20% of oxygen | bovine | DNMT3A | Elevated DNMT3A expresiion and increase of global DNA methylation in 4-cell embryos and blastocysts | [180] |
| Oxidative stress (palmitic acid) | bovine | PRDX3, HADHB, UQCRB, CYCS | Upregulation of PRDX3 protein. Elevation of the mitochondrial HADHB, UQCRB and CYCS proteins in oocytes | [181] |
| Oxidative stress (H2O2) | mouse | NE | Decrease in mitochondria-derived ATP and disassembly of spindles in in vitro cultured MII oocytes | [85] |
| In vitro techniques | ||||
| Oocyte in vitro maturation | human | HDAC1 | Compromised deacetylation in oocytes. Residual acetylation linked to aneuploidy | [182] |
| Oocyte in vitro maturation | bovine | SIRT2 | Faulty mitochondria | [183] |
| Cytoplasmic transfer | human | Not tested yet | 10–15% cytoplasm transfer into aged oocytes prroduced healthy offspring | [95] |
| Suboptimal culture media | rabbit | NE | Alteration of DNA methylation reprogramming in paternal pronuclei of zygotes | [184] |
| In vitro fertilization & ICSI | human | H19 | ART caused demethylation resulted in the changes of genomic imprinting | [185] |
| Embryo in vitro culture | human | NE | miRNAS detected in spent culture medium downregulate embryonic mRNAs | [186] |
| Cryopreservation | human | LINE1 | Differently methylated placental DNA between fresh and frozen embryotransfers | [187] |
| Suboptimal culture media | mouse | NE | Higher methylation disturbances in embryos from superovulated females and IVF | [188] |
| Intracytoplasmic sperm injection | mouse | H19, Snrpn, Peg3, Igf2 | Imprinting defects in somatic tissues | [189] |
Table 2.
Summary of lifestyle, diet, and environment effect on epigenetic changes in oocytes, early embryos, and their impact on offspring. “NE” indicates specific gene effects were not evaluated.
Table 2.
Summary of lifestyle, diet, and environment effect on epigenetic changes in oocytes, early embryos, and their impact on offspring. “NE” indicates specific gene effects were not evaluated.
| Stressor | Species | Genes Affected | Main Findings | Reference |
|---|---|---|---|---|
| Undernutrition | ||||
| Periconceptional exposure to famine | human | IGF2 | IGF2 hypomethylation in adults 60 years later | [290] |
| Periconceptional exposure to famine | human | IL10, INSIGF, LEP, MEG3, ABCA1, GNASAS | Altered DNA methylation of genes implicated in growth and metabolic regulation | [270] |
| Prenatal exposure to famine | human | ABCG1, PFKFB3, METTL8 | Altered DNA methylation of genes associated with lipid metabolism, glycolysis, and adipogenesis in adults | [269] |
| Low levels of dietary methyl donors during embryonic development | human | NE | Affected DNA methylation process and impact on postnatal long-term health | [291] |
| Protein restriction during pregnancy | mouse | Lep | Increased Lep promoter methylation and decreased leptin expression in offspring | [292] |
| Preovulatory protein restriction | rat | Drp1, Opa1, Mfn1/2, Parl, Ndufb6, Hk2 | Altered expression of genes involved in mitochondrial biogenesis in superovulated oocytes | [271] |
| Negative energy balance and metabolic stress | bovine | NE | Hypomethylation of maternally inherited imprinted genes in oocytes of postpartum cows | [274] |
| Obesity | ||||
| Obesity in pregnancy | human | TET1, TET2. TET3 | DNA hypermethylation and reduced expression of methylcytosine dioxygenases in placenta | [58] |
| Gestational weight gain | human | MMP7,KCNK4,TRPM5 and NFKB1 | Increased DNA methylation in offspring. | [277] |
| Obesity in pregnancy | mouse | IGF-1, mTOR, LEP | Stimulation of placental insulin/IGF-1/mTOR and leptin signalling pathways | [163] |
| Alcohol, smoking | ||||
| Alcohol | mouse, human | NE | Birth defects and neuronal disorders in offspring | [281,284] |
| Alcohol | mouse | Cyp4f13, Nlgn3, Elavl2, Sox21, Sim1, Igf2r, Hist1h3d | Decreased methylation of genes associated with development, imprinting and chromatin in embryos exposed to ethanol in vitro | [289] |
| Alcohol | mouse | H19/Igf2 | Retardation of embryo development in vivo and alteration of the H19/Igf2 methylation in placenta | [288] |
| Maternal smoking | human | BMP4, BMHT2, DLGAP2, PRDM8, NRP2, ESR1, IL32, HOXB2 | In newborns, changes in CpGs methylation of genes involved in tooth and neuronal development and in cancer induction | [293] |
| Maternal age | ||||
| Advanced maternal age (more than 40 years) | human | BUB1B, BUB3, MAD3, BUB1, REC8, ATR, CHEK1, NBS1, RAD17, EIF4ENIF1 | In oocytes, reduced expression of spindle checkpoint and DNA damage checkpoint-related genes, lowered mRNA expression of the nuclear import mediator of eIF4E | [294] |
| Advanced maternal age (more than 40 years) | human | REC8, SMC1B | Decreased expression of the meiosis-specific cohesins components, REC8 proteins, and SMC1B in oocytes | [295] |
| Advanced maternal age (41–44 years) | human | PRDX1, PRDX2, PRDX4, PRDX6 COX5A, COX7B, COX8A, COX8C, COX11, COX14, COX17 | Down-regulation of the peroxiredoxin gene family members and attenuated expression of the cytochrome c oxidases in oocytes | [296] |
| Advanced age | mouse | Dnmt3a, Dnmt3b, Tfam | Downregulation of maintenance DNA methyltransferases and mitochondrial transcription factor in oocytes | [297] |
| Advanced age | mouse | Hook1, Tuba1, Tubd1, Dncic2, Kif3, Rnf19/Dorfin, Pcnt2, Nin, Smc4l1, Dnmt1o, Dmap1, Dnmt3L | Decrease of transcripts related to microtubule cytoskeleton and chromosome segregation, downregulation of methyltransferases in oocytes | [298] |
| Other causes | ||||
| Maternal stress during pregnancy | human | HSD11B2, NR3C1, FKBP5 | Increased methylation and expression of glucocorticoid pathway-related genes in placenta and children blood. | [299,300] |
| Maternal gestational diabetes | human | PDE6A, PRKCZ, PVT1, GALNT2, MS4A3, IL1RN, BTD | In children, differentially methylated genes associated with type 2 diabetes, obesity, diabetic nephropathy, and coronary heart disease. | [301] |
| Eleveted homocysteine level | porcine | mtDNA (12S, 16S rRNA and ND4) ND1, ND4L, ND5, COX1, CYTB mRNA | Hypermethylation of mtDNA in oocytes from PCOS ovaries | [302] |
| Pollutants | ||||
| Bisphenol A | human | MEST | Hypomethylation of the obesity-associated mesoderm-specific-transcript (MEST) gene promoter and enhanced MEST expression in children | [303] |
| Bisphenol A | mouse | Snrpn, Ube3a, Igf2, Kcnq1ot1, Cdkn1c, Ascl2 | Disruption of imprinted gene expression in embryos and placentas. | [304] |
| Polystyrene | mouse | NE | Negative effect on oocyte spindle assembly and chromosome alignment, increased oxidative stress, and mitochondrial agregation | [305] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dvoran, M.; Nemcova, L.; Kalous, J. An Interplay between Epigenetics and Translation in Oocyte Maturation and Embryo Development: Assisted Reproduction Perspective. Biomedicines 2022, 10, 1689. https://doi.org/10.3390/biomedicines10071689
AMA Style
Dvoran M, Nemcova L, Kalous J. An Interplay between Epigenetics and Translation in Oocyte Maturation and Embryo Development: Assisted Reproduction Perspective. Biomedicines. 2022; 10(7):1689. https://doi.org/10.3390/biomedicines10071689
Chicago/Turabian StyleDvoran, Michal, Lucie Nemcova, and Jaroslav Kalous. 2022. "An Interplay between Epigenetics and Translation in Oocyte Maturation and Embryo Development: Assisted Reproduction Perspective" Biomedicines 10, no. 7: 1689. https://doi.org/10.3390/biomedicines10071689
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.