
Identification and Characterization of a Novel Umbra-like Virus, Strawberry Virus A, Infecting Strawberry Plants


Abstract
:1. Introduction
2. Results and Discussion
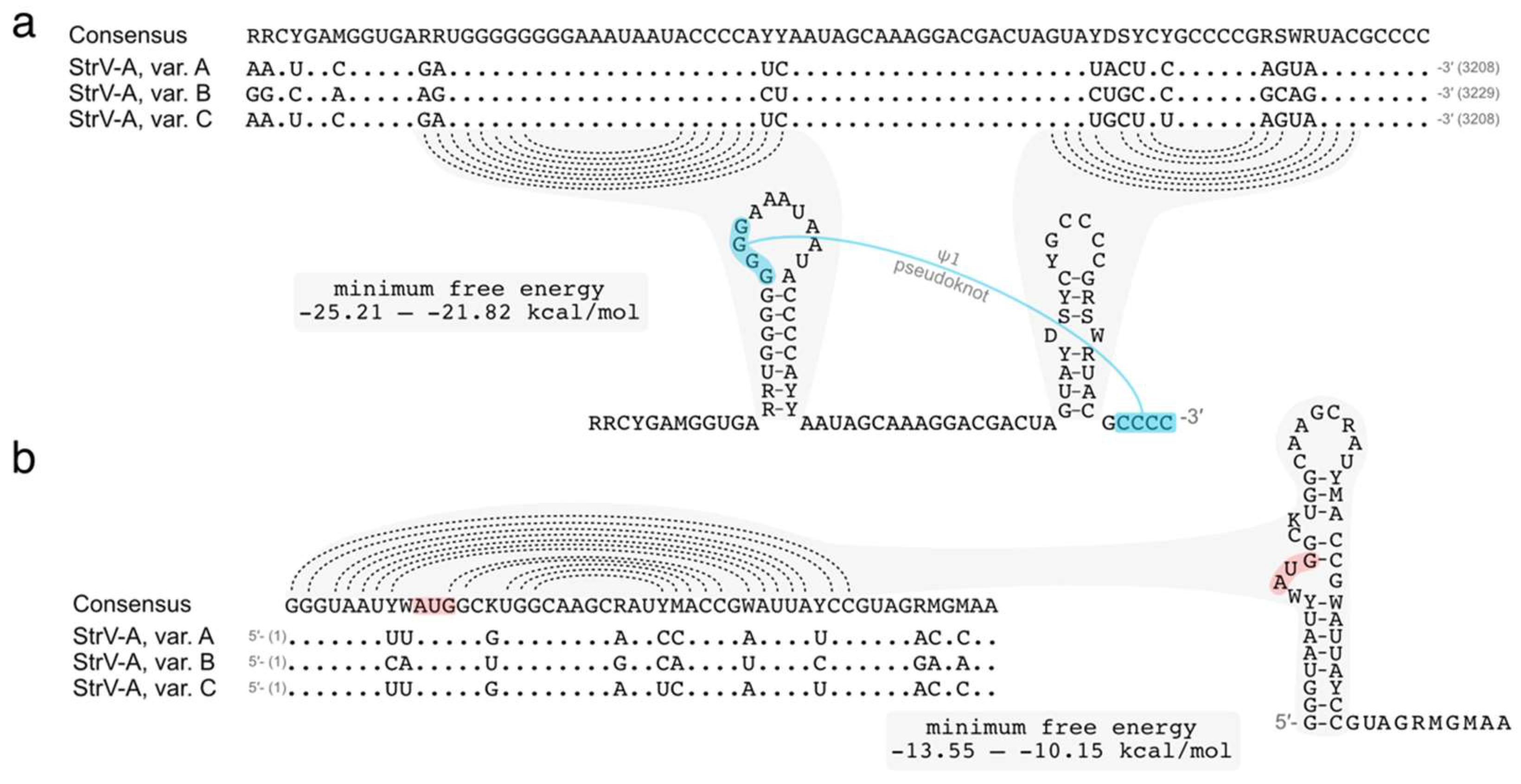
2.1. StrVA Genome Sequencing and Characterization
2.2. Real-Time PCR for StrVA Detection
2.3. StrVA Presence in Strawberry Samples
2.4. Single-Host Infection with Multiple Genetic Variants of StrVA
2.5. Detection of StrVA in Non-Strawberry Hosts
3. Conclusions
4. Materials and Methods
4.1. Plant Materials
4.2. Sampling, Real-Time Reverse Transcription PCR and Reverse Transcription PCR Detection of Viruses
4.3. Sequence Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taliansky, M.E.; Robinson, D.J. Molecular Biology of Umbraviruses: Phantom Warriors. J. Gen. Virol. 2003, 84, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Science: Burlington, MA, USA, 2011; ISBN 978-0-12-384685-3. [Google Scholar]
- Syller, J. Molecular and Biological Features of Umbraviruses, the Unusual Plant Viruses Lacking Genetic Information for a Capsid Protein. Physiol. Mol. Plant Pathol. 2003, 63, 35–46. [Google Scholar] [CrossRef]
- Campbell, A.J.; Erickson, A.; Pellerin, E.; Salem, N.; Mo, X.; Falk, B.W.; Ferriol, I. Phylogenetic Classification of a Group of Self-Replicating RNAs That Are Common in Co-Infections with Poleroviruses. Virus Res. 2020, 276, 197831. [Google Scholar] [CrossRef] [PubMed]
- Yoo, R.H.; Lee, S.-W.; Lim, S.; Zhao, F.; Igori, D.; Baek, D.; Hong, J.-S.; Lee, S.-H.; Moon, J.S. Complete Genome Analysis of a Novel Umbravirus-Polerovirus Combination Isolated from Ixeridium Dentatum. Arch. Virol. 2017, 162, 3893–3897. [Google Scholar] [CrossRef] [PubMed]
- Quito-Avila, D.F.; Alvarez, R.A.; Ibarra, M.A.; Martin, R.R. Detection and Partial Genome Sequence of a New Umbra-like Virus of Papaya Discovered in Ecuador. Eur. J. Plant Pathol. 2015, 143, 199–204. [Google Scholar] [CrossRef]
- Sá Antunes, T.F.; Amaral, R.J.V.; Ventura, J.A.; Godinho, M.T.; Amaral, J.G.; Souza, F.O.; Zerbini, P.A.; Zerbini, F.M.; Fernandes, P.M.B. The DsRNA Virus Papaya Meleira Virus and an SsRNA Virus Are Associated with Papaya Sticky Disease. PLoS ONE 2016, 11, e0155240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornejo-Franco, J.F.; Flores, F.; Mollov, D.; Quito-Avila, D.F. An Umbra-Related Virus Found in Babaco (Vasconcellea × Heilbornii). Arch. Virol. 2021, 166, 2321–2324. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.-J.; Bodaghi, S.; Dang, T.; Gadhave, K.R.; Ho, T.; Osman, F.; Al Rwahnih, M.; Tzanetakis, I.E.; Simon, A.E.; Vidalakis, G. Complete Nucleotide Sequence, Genome Organization, and Comparative Genomic Analyses of Citrus Yellow-Vein Associated Virus (CYVaV). Front. Microbiol. 2021, 12, 683130. [Google Scholar] [CrossRef] [PubMed]
- Redila, C.D.; Prakash, V.; Nouri, S. Metagenomics Analysis of the Wheat Virome Identifies Novel Plant and Fungal-Associated Viral Sequences. Viruses 2021, 13, 2457. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Olmedo-Velarde, A.; Larrea-Sarmiento, A.; Simon, A.E.; Kong, A.; Borth, W.; Suzuki, J.Y.; Wall, M.M.; Hu, J.; Melzer, M. Genome Characterization of Fig Umbra-like Virus. Virus Genes 2021, 57, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Fránová, J.; Přibylová, J.; Koloniuk, I. Molecular and Biological Characterization of a New Strawberry Cytorhabdovirus. Viruses 2019, 11, 982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fránová, J.; Lenz, O.; Přibylová, J.; Čmejla, R.; Valentová, L.; Koloniuk, I. High Incidence of Strawberry Polerovirus 1 in the Czech Republic and Its Vectors, Genetic Variability and Recombination. Viruses 2021, 13, 2487. [Google Scholar] [CrossRef] [PubMed]
- McCormack, J.C.; Yuan, X.; Yingling, Y.G.; Kasprzak, W.; Zamora, R.E.; Shapiro, B.A.; Simon, A.E. Structural Domains within the 3′ Untranslated Region of Turnip Crinkle Virus. J. Virol. 2008, 82, 8706–8720. [Google Scholar] [CrossRef] [Green Version]
- Guan, H.; Carpenter, C.D.; Simon, A.E. Analysis of Cis-Acting Sequences Involved in Plus-Strand Synthesis of a Turnip Crinkle Virus-Associated Satellite RNA Identifies a New Carmovirus Replication Element. Virology 2000, 268, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimešová, J.; Kociánová, A.; Martínková, J. Weeds That Can Do Both Tricks: Vegetative versus Generative Regeneration of the Short-Lived Root-Sprouting Herbs Rorippa Palustris and Barbarea Vulgaris. Weed Res. 2008, 48, 131–135. [Google Scholar] [CrossRef]
- Thompson, J.R.; Wetzel, S.; Klerks, M.M.; Vašková, D.; Schoen, C.D.; Špak, J.; Jelkmann, W. Multiplex RT-PCR Detection of Four Aphid-Borne Strawberry Viruses in Fragaria Spp. in Combination with a Plant MRNA Specific Internal Control. J. Virol. Methods 2003, 111, 85–93. [Google Scholar] [CrossRef]
- Thekke-Veetil, T.; Tzanetakis, I.E. First Report of Strawberry Polerovirus-1 in Strawberry in the United States. Plant Dis. 2016, 100, 867. [Google Scholar] [CrossRef]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A Computer Program for Analyzing Recombination in, and Removing Signals of Recombination from, Nucleotide Sequence Datasets. Virus Evol. 2021, 7, veaa087. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Samples (n, %) | StrVA Positivity (n, %) | Mixed Infections (n, %) |
|---|---|---|---|
| Commercial plantings | 234 (35.9%) | 147 (62.8%) | 62 (42.2%) |
| Gardens | 42 (6.5%) | 9 (21.4%) | 8 (88.9%) |
| SE 1-certified propagation material | 80 (12.3%) | 3 (3.8%) | 2 (66.7%) |
| CAC 2-certified propagation material | 186 (28.6%) | 25 (13.4%) | 0 |
| Hobby markets | 109 (16.7%) | 0 | 0 |
| Total | 651 | 184 (28.3%) | 72 (39.1%) |
| Virus | StrVA, var. A | StrVA, var. B | StrVA, var. C | |
| StrVA, var. A | - | 69 (70) | 89 (91) | ORF3 (p43) |
| StrVA, var. B | 80 (87) | - | 71 (70) | |
| StrVA, var. C | 92 (97) | 92 (87) | - | |
| ORF1+ORF2 (p21 and p59.4) | - | |||
| Primer/Probe | Sequence (5′ to 3′ Direction) | Final Concentration (μM) | Usage |
|---|---|---|---|
| StrVA | |||
| Forward | ACCTGGCCTTGTCYCGGC | 0.5 | sequencing of virus isolates |
| Reverse | WGGTGGWGGGGACGTACAAC | 0.5 | |
| 1750 Forward | GATGTCTGGTGATATGGACA | 0.5 | RT-PCR assay |
| 1751 Reverse | ACCAATTCTCTACATCGTGT | 0.5 | |
| Forward | ATCTGGGTGAAGRCGGGGT | 0.5 | qPCR |
| Reverse | CAAAACCCTCTCATAAGGTTRTCCA | 0.5 | |
| Probe | ACTGTGGATATCCCCGACTGTATTGT | 0.2 | |
| StrV-1 | |||
| Forward | AACGGATATTGTGGCGCRAA | 0.5 | qPCR |
| Reverse | CCTGATGTTGTTKATATARCTGAGRTC | 0.5 | |
| Probe 01 | AAACCTCTTACCATCATCTCGTAAA | 0.175 | |
| Probe 02 | AARCCCCTCACCATCATYTCGTAA | 0.175 | |
| SMYEV | |||
| Forward | CCCTCCTGACGTACACAACAACTG | 0.5 | qPCR |
| Reverse 01 | CCGTGAGGGAGGAGAATACGC | 0.25 | |
| Reverse 02 | CCGTGAGGGAGGAGAATACAC | 0.25 | |
| Probe | TACTCTAGTYGCCATCGAGGTACAGTGC | 0.2 | |
| SCV | |||
| Forward 01 | ACAGTRTGCGCTTTAGAGGTTGTT | 0.5 | qPCR |
| Forward 02 | ACAGTGTGCGCTTTAGAGGTTATTC | 0.5 | |
| Forward 03 | ACAGTGTGCGCTTTAGAAGTTGTTC | 0.25 | |
| Reverse 01 | ACCTGATTATCTCCCATYCCCATT | 0.5 | |
| Reverse 02 | ACTTGATTATCCCCCATCCCCA | 0.5 | |
| Probe 01 | TCTCAATAYGATTGTACATACCGCAT | 0.15 | |
| Probe 02 | TCTCAATACGATTGCACATATCGCAT | 0.15 | |
| SVBV | |||
| Forward | AATATCTGTCTTTACTTGATSATGAACTTG | 0.5 | qPCR |
| Reverse 01 | CGTCTTCGCTGCTGCTGTAGTC | 0.25 | |
| Reverse 02 | CATCTTCACTGCTGCTGCTGTAG | 0.25 | |
| Probe | AGTTACAGGTACTTGTAGCAAAAGARATGA | 0.2 | |
| SMoV | |||
| Forward | GTAGGACACCGGCTCTTGGYAGT | 0.5 | qPCR |
| Reverse | TTGGRTCGTCACCTGAYCTCG | 0.5 | |
| Probe | ACAGGWGGCACTGTTTACAGTGTTCC | 0.25 | |
| SPV1 | |||
| Forward | CAACTGGGGTCGTACACTCGC | 0.5 | qPCR |
| Reverse | GGCCAGCCGAATCCTTTGAC | 0.5 | |
| Probe | ACCTACCCTGAACTCGCCGAACA | 0.2 | |
| Nad5 | |||
| Forward | GATGCTTCTTGGGGCTTCTTGTT | 0.25 | qPCR |
| Reverse | ACATAAATCGAGGGCTATGCGG | 0.25 | |
| Probe | CCACAATTAACATCACTACGGTCGGGCTA | 0.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koloniuk, I.; Přibylová, J.; Čmejla, R.; Valentová, L.; Fránová, J. Identification and Characterization of a Novel Umbra-like Virus, Strawberry Virus A, Infecting Strawberry Plants. Plants 2022, 11, 643. https://doi.org/10.3390/plants11050643
Koloniuk I, Přibylová J, Čmejla R, Valentová L, Fránová J. Identification and Characterization of a Novel Umbra-like Virus, Strawberry Virus A, Infecting Strawberry Plants. Plants. 2022; 11(5):643. https://doi.org/10.3390/plants11050643
Chicago/Turabian StyleKoloniuk, Igor, Jaroslava Přibylová, Radek Čmejla, Lucie Valentová, and Jana Fránová. 2022. "Identification and Characterization of a Novel Umbra-like Virus, Strawberry Virus A, Infecting Strawberry Plants" Plants 11, no. 5: 643. https://doi.org/10.3390/plants11050643