
Metabolome-Wide, Phylogenetically Controlled Comparison Indicates Higher Phenolic Diversity in Tropical Tree Species

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Species Selection and Sample Processing
2.2. Data Analysis
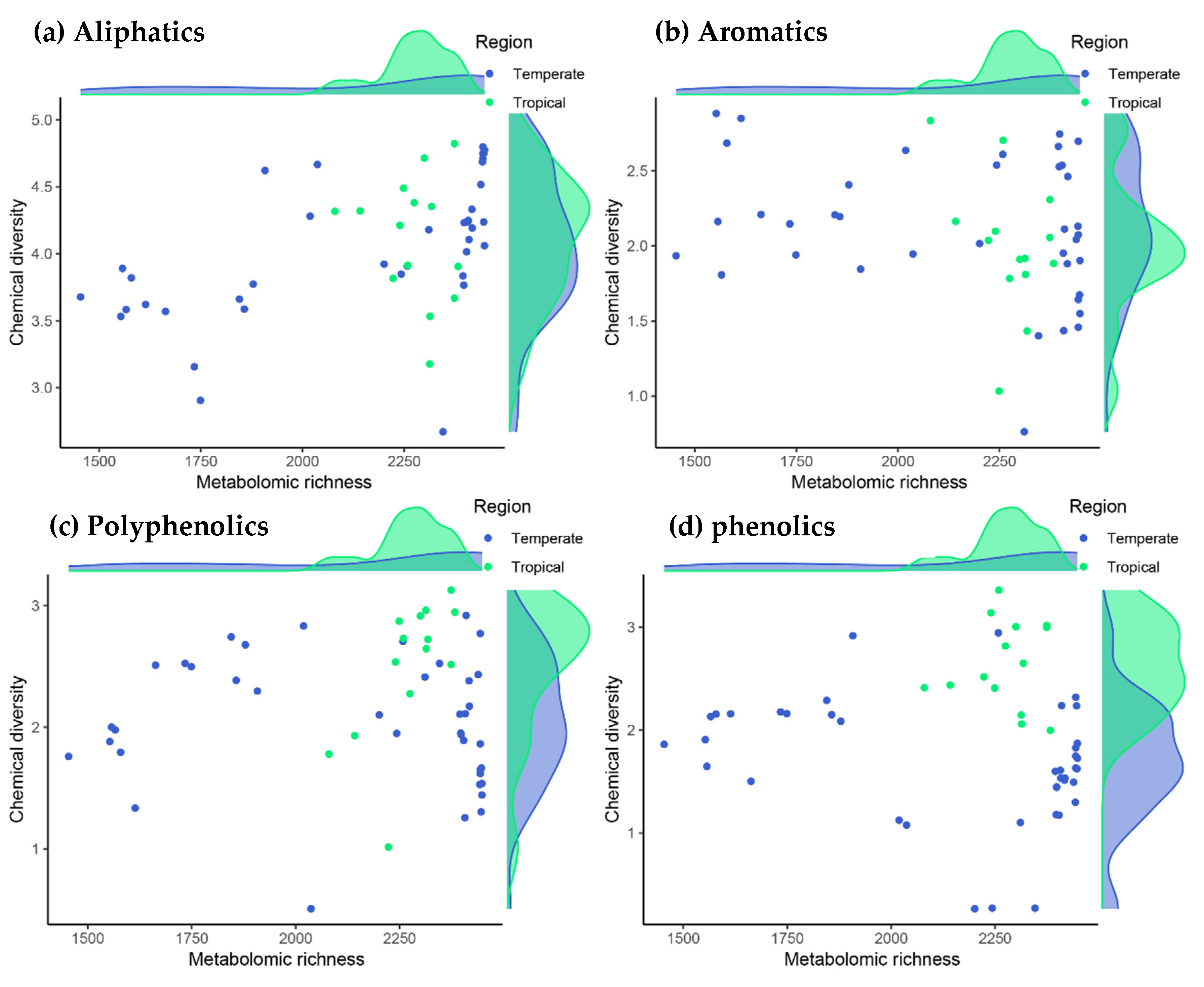
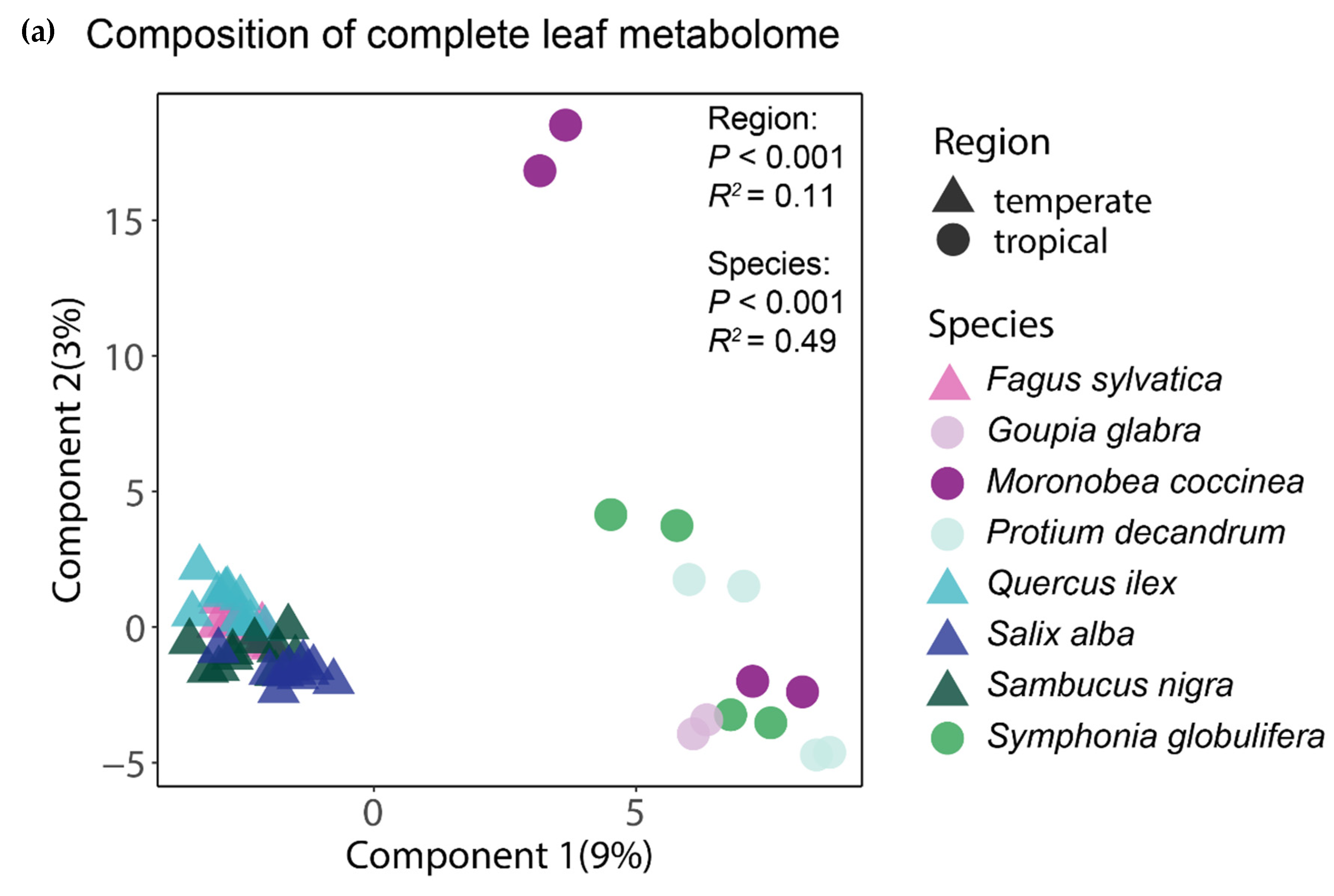
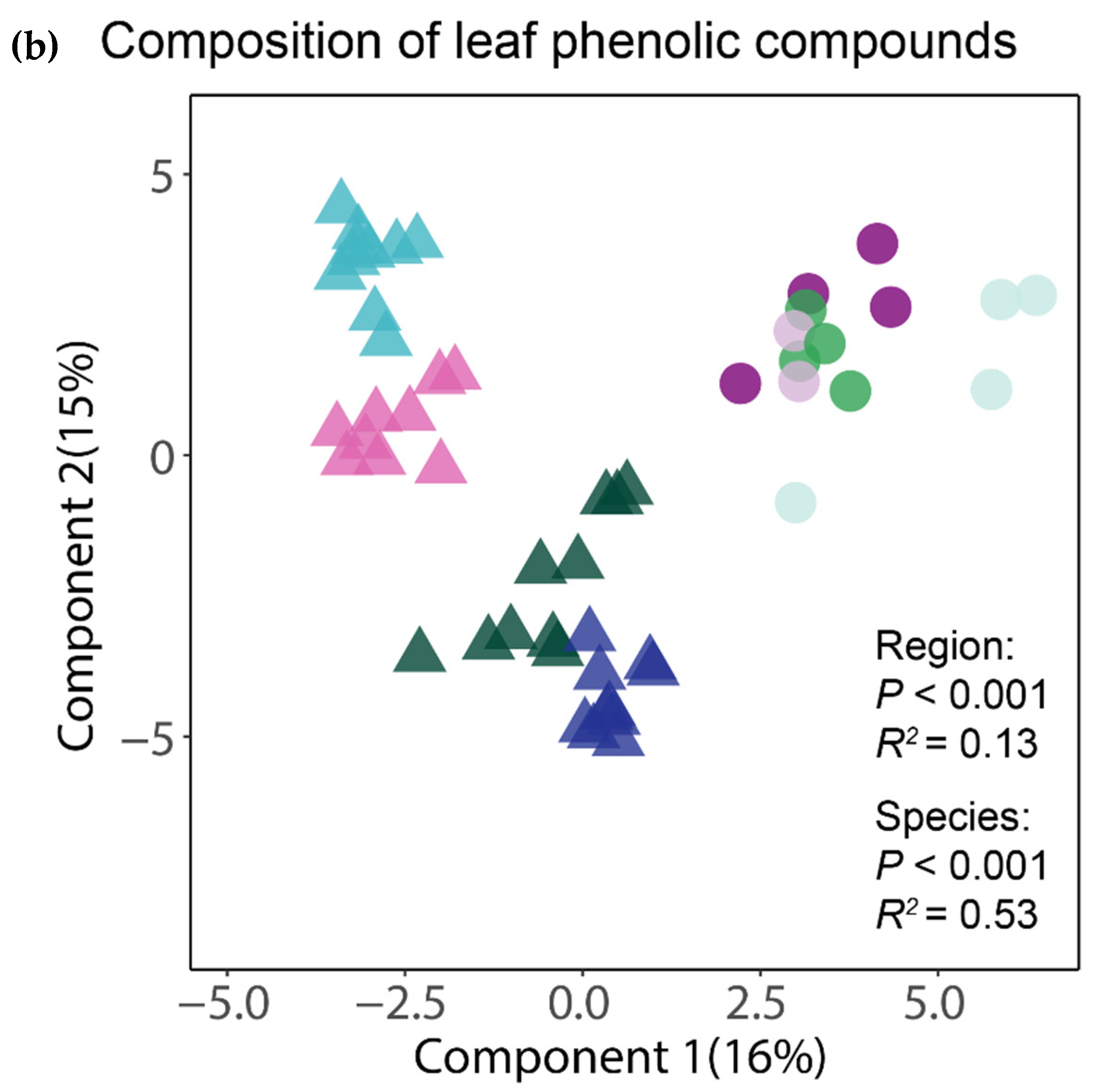
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, J.H. Why are there so many species in the tropics? J. Biogeogr. 2014, 41, 8–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schemske, D.W.; Mittelbach, G.G.; Cornell, H.V.; Sobel, J.M.; Roy, K. Is there a latitudinal gradient in the importance of biotic interactions? Annu. Rev. Ecol. Evol. Syst. 2009, 40, 245–269. [Google Scholar] [CrossRef] [Green Version]
- Dobzhansky, T. Evolution in the tropics. Am. Sci. 1950, 38, 209–221. [Google Scholar]
- Ehrlich, P.R.; Raven, P.H. Butterflies and Plants: A Study in Coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- MacArthur, R.H. Geographical Ecology: Patterns in The Distribution of Species; Princeton University Press: Princeton, NJ, USA, 1972. [Google Scholar]
- Mittelbach, G.G.; Schemske, D.W.; Cornell, H.V.; Allen, A.P.; Brown, J.M.; Bush, M.B.; Harrison, S.P.; Hurlbert, A.H.; Knowlton, N.; Lessios, H.A.; et al. Evolution and the latitudinal diversity gradient: Speciation, extinction and biogeography. Ecol. Lett. 2007, 10, 315–331. [Google Scholar] [CrossRef]
- Coley, P.D.; Barone, J.A. Herbivory and Plant Defenses in Tropical Forests. Annu. Rev. Ecol. Syst. 1996, 27, 305–335. [Google Scholar] [CrossRef]
- Levin, D.A. The Chemical Defenses of Plants to Pathogens and Herbivores. Annu. Rev. Ecol. Syst. 1976, 7, 121–159. [Google Scholar] [CrossRef]
- Bagchi, R.; Gallery, R.E.; Gripenberg, S.; Gurr, S.J.; Narayan, L.; Addis, C.E.; Freckleton, R.P.; Lewis, O.T. Pathogens and insect herbivores drive rainforest plant diversity and composition. Nature 2014, 506, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Salazar, D.; Marquis, R.J. Herbivore pressure increases toward the equator. Proc. Natl. Acad. Sci. USA 2012, 109, 12616–12620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kursar, T.A.; Dexter, K.G.; Lokvam, J.; Pennington, R.T.; Richardson, J.E.; Weber, M.G.; Murakami, E.T.; Drake, C.; McGregor, R.; Coley, P.D. The evolution of antiherbivore defenses and their contribution to species coexistence in the tropical tree genus Inga. Proc. Natl. Acad. Sci. USA 2009, 106, 18073–18078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, J.X. On the factors that promote the diversity of herbivorous insects and plants in tropical forests. Proc. Natl. Acad. Sci. USA 2015, 112, 6098–6103. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.Y.; Fine, P.V.A.; Mittelbach, G.G. Assessing the latitudinal gradient in herbivory. Glob. Ecol. Biogeogr. 2015, 24, 1106–1112. [Google Scholar] [CrossRef] [Green Version]
- Roslin, T.; Hardwick, B.; Novotny, V.; Petry, W.K.; Andrew, N.R.; Asmus, A.; Barrio, I.C.; Basset, Y.; Boesing, A.L.; Bonebrake, T.C.; et al. Higher predation risk for insect prey at low latitudes and elevations. Science 2017, 356, 742–744. [Google Scholar] [CrossRef] [Green Version]
- Fine, P.V.A.; Mesones, I.; Coley, P.D. Herbivores Promote Habitat Specialization by Trees in Amazonian Forests. Science 2004, 305, 663–665. [Google Scholar] [CrossRef] [Green Version]
- Mangan, S.A.; Schnitzer, S.A.; Herre, E.A.; Mac, K.K.M.L.; Valencia, M.C.; Sanchez, E.I.; Bever, J.D. Negative plant-soil feedback predicts tree-species relative abundance in a tropical forest. Nature 2010, 466, 752–755. [Google Scholar] [CrossRef]
- Moles, A.T.; Bonser, S.P.; Poore, A.G.B.; Wallis, I.R.; Foley, W.J. Assessing the evidence for latitudinal gradients in plant defence and herbivory. Funct. Ecol. 2011, 25, 380–388. [Google Scholar] [CrossRef]
- Moles, A.T.; Wallis, I.R.; Foley, W.J.; Warton, D.I.; Stegen, J.C.; Bisigato, A.J.; Pizarro, L.C.; Clark, C.J.; Cohen, P.S.; Cornwell, W.K.; et al. Putting plant resistance traits on the map: A test of the idea that plants are better defended at lower latitudes. New Phytol. 2011, 191, 777–788. [Google Scholar] [CrossRef]
- Anstett, D.N.; Nunes, K.A.; Baskett, C.; Kotanen, P.M. Sources of Controversy Surrounding Latitudinal Patterns in Herbivory and Defense. Trends Ecol. Evol. 2016, 31, 789–802. [Google Scholar] [CrossRef]
- Sedio, B.E. Recent breakthroughs in metabolomics promise to reveal the cryptic chemical traits that mediate plant community composition, character evolution and lineage diversification. New Phytol. 2017, 214, 952–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, A.A. Macroevolution of plant defense strategies. Trends Ecol. Evol. 2006, 22, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Hall, R.D. Plant metabolomics: From holistic hope, to hype, to hot topic. New Phytol. 2005, 169, 468–543. [Google Scholar] [CrossRef]
- Hilker, M. New Synthesis: Parallels Between Biodiversity and Chemodiversity. J. Chem. Ecol. 2014, 40, 225–226. [Google Scholar] [CrossRef]
- Dyer, L.A.; Philbin, C.S.; Ochsenrider, K.M.; Richards, L.A.; Massad, T.J.; Smilanich, A.M.; Forister, M.L.; Parchman, T.L.; Galland, L.M.; Hurtado, P.J.; et al. Modern approaches to study plant–insect interactions in chemical ecology. Nat. Rev. Chem. 2018, 2, 50–64. [Google Scholar] [CrossRef]
- Hadfield, J.D.; Nakagawa, S. General quantitative genetic methods for comparative biology: Phylogenies, taxonomies and multi-trait models for continuous and categorical characters. J. Evol. Biol. 2010, 23, 494–508. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Liu, Y.; Bianchi, T.S.; Tolić, N.; Jansson, C.; Paša-Tolić, L. Moving beyond the van Krevelen Diagram: A New Stoichiometric Approach for Compound Classification in Organisms. Anal. Chem. 2018, 90, 6152–6160. [Google Scholar] [CrossRef] [PubMed]
- Volf, M.; Segar, S.T.; Miller, S.E.; Isua, B.; Sisol, M.; Aubona, G.; Šimek, P.; Moos, M.; Laitila, J.; Kim, J.; et al. Community structure of insect herbivores is driven by conservatism, escalation and divergence of defensive traits in Ficus. Ecol. Lett. 2018, 21, 83–92. [Google Scholar] [CrossRef]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.K.; Eastman, J.M.; Smith, S.A.; FitzJohn, R.G.; McGlinn, D.J.; O’Meara, B.C.; Moles, A.T.; Reich, P.B.; et al. Three keys to the radiation of angiosperms into freezing environments. Nature 2014, 506, 89–92. [Google Scholar] [CrossRef]
- Kellerman, A.M.; Dittmar, T.; Kothawala, D.N.; Tranvik, L.J. Chemodiversity of dissolved organic matter in lakes driven by climate and hydrology. Nat. Commun. 2014, 5, 3804. [Google Scholar] [CrossRef] [Green Version]
- Bürkner, P.C. Brms: An R package for Bayesian multilevel models using Stan. J. Stat. Softw. 2017, 80. [Google Scholar] [CrossRef] [Green Version]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Qian, H.; Jin, Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. J. Plant Ecol. 2016, 9, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Rohart, F.; Gautier, B.; Singh, A.; Cao, K.A.L. MixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Friendly, M.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.4-3. Vienna: R Foundation for Statistical Computing. 2016. Available online: https://www.researchgate.net/publication/323265822_vegan_Community_Ecology_Package_R_package_version_24-3_2017_accessed_2016_Jan_1 (accessed on 13 March 2021).
- Salazar, D.; Lokvam, J.; Mesones, I.; Pilco, M.V.; Zuñiga, J.M.A.; de Valpine, P.; Fine, P.V.A. Origin and maintenance of chemical diversity in a species-rich tropical tree lineage. Nat. Ecol. Evol. 2018, 2, 983–990. [Google Scholar] [CrossRef]
- Forister, M.L.; Novotny, V.; Panorska, A.K.; Baje, L.; Basset, Y.; Butterill, P.T.; Cizek, L.; Coley, P.D.; Dem, F.; Diniz, I.R.; et al. The global distribution of diet breadth in insect herbivores. Proc. Natl. Acad. Sci. USA 2015, 112, 442–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peguero, G.; Bonal, R.; Sol, D.; Muñoz, A.; Sork, V.L.; Espelta, J.M. Tropical insect diversity: Evidence of greater host specialization in seed-feeding weevils. Ecology 2017, 98, 2180–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, L.A.; Dyer, L.A.; Forister, M.L.; Smilanich, A.M.; Dodson, C.D.; Leonard, M.D.; Jeffrey, C.S. Phytochemical diversity drives plant–insect community diversity. Proc. Natl. Acad. Sci. USA 2015, 112, 10973–10978. [Google Scholar] [CrossRef] [Green Version]
- Sedio, B.E.; Parker, J.D.; McMahon, S.M.; Wright, S.J. Comparative foliar metabolomics of a tropical and a temperate forest community. Ecology 2018, 99, 2647–2653. [Google Scholar] [CrossRef] [Green Version]
- Sedio, B.E.; Echeverri, J.C.R.; Boya, C.A.; Wright, S.J. Sources of variation in foliar secondary chemistry in a tropical forest tree community. Ecology 2017, 98, 616–623. [Google Scholar] [CrossRef] [Green Version]
- Garriga, A.G.; Sardans, J.; Granda, V.; Llusià, J.; Peguero, G.; Asensio, D.; Ogaya, R.; Urbina, I.; van Langenhove, L.; Verryckt, L.T.; et al. Different “metabolomic niches” of the highly diverse tree species of the French Guiana rainforests. Sci. Rep. 2020, 10, 6937. [Google Scholar] [CrossRef] [PubMed]
- Sedio, B.E.; Archibold, A.D.; Echeverri, J.C.R.; Debyser, C.; Cristopher, A.B.P.; Wright, S.J. A comparison of inducible, ontogenetic, and interspecific sources of variation in the foliar metabolome in tropical trees. PeerJ 2019, 7, e7536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas-Ubach, A.; Pérez-Trujillo, M.; Sardans, J.; Gargallo-Garriga, A.; Parella, T.; Peñuelas, J. Ecometabolomics: Optimized NMR-based method. Methods Ecol. Evol. 2013, 4, 464–473. [Google Scholar] [CrossRef]
- Koch, B.P.; Dittmar, T.; Witt, M.; Kattner, G. Fundamentals of molecular formula assignment to ultrahigh resolution mass data of natural organic matter. Anal. Chem. 2007, 79, 1758–1763. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Region | Species Richness | Phylodiversity | Mean Pairwise Distance | Mean Neighbor Taxon Distance |
|---|---|---|---|---|
| Tropical | 4 | 377 | 200 | 146 |
| Temperate | 4 | 392 | 208 | 157 |
| Species Names and Family | Sampled Leaves | Individuals |
|---|---|---|
| Fagus sylvatica L. (Fagaceae) | 9 | 9 |
| Goupia glabra Aubl. (Goupiaceae) | 2 | 1 |
| Moronobea coccinea Aubl. (Clusiaceae) | 4 | 1 |
| Protium decandrum Aubl. (Burseraceae) | 4 | 1 |
| Quercus ilex L. (Fagaceae) | 10 | 10 |
| Salix alba L. (Salicaceae) | 10 | 4 |
| Sambucus nigra L. (Adoxaceae) | 10 | 10 |
| Symphonia globulifera L.f. (Clusiaceae) | 4 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peguero, G.; Gargallo-Garriga, A.; Maspons, J.; Klem, K.; Urban, O.; Sardans, J.; Peñuelas, J. Metabolome-Wide, Phylogenetically Controlled Comparison Indicates Higher Phenolic Diversity in Tropical Tree Species. Plants 2021, 10, 554. https://doi.org/10.3390/plants10030554
Peguero G, Gargallo-Garriga A, Maspons J, Klem K, Urban O, Sardans J, Peñuelas J. Metabolome-Wide, Phylogenetically Controlled Comparison Indicates Higher Phenolic Diversity in Tropical Tree Species. Plants. 2021; 10(3):554. https://doi.org/10.3390/plants10030554
Chicago/Turabian StylePeguero, Guille, Albert Gargallo-Garriga, Joan Maspons, Karel Klem, Otmar Urban, Jordi Sardans, and Josep Peñuelas. 2021. "Metabolome-Wide, Phylogenetically Controlled Comparison Indicates Higher Phenolic Diversity in Tropical Tree Species" Plants 10, no. 3: 554. https://doi.org/10.3390/plants10030554