
Memantine and Riluzole Exacerbate, Rather Than Ameliorate Behavioral Deficits Induced by 8-OH-DPAT Sensitization in a Spatial Task

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs and Design
2.3. Apparatus and Behavioral Procedures
2.3.1. Active Place Avoidance Task in Carousel Maze
2.3.2. 8-OH-DPAT Sensitization and Habituation to the Arena
2.3.3. Acquisition Testing
2.4. Data Analysis and Statistics
3. Results
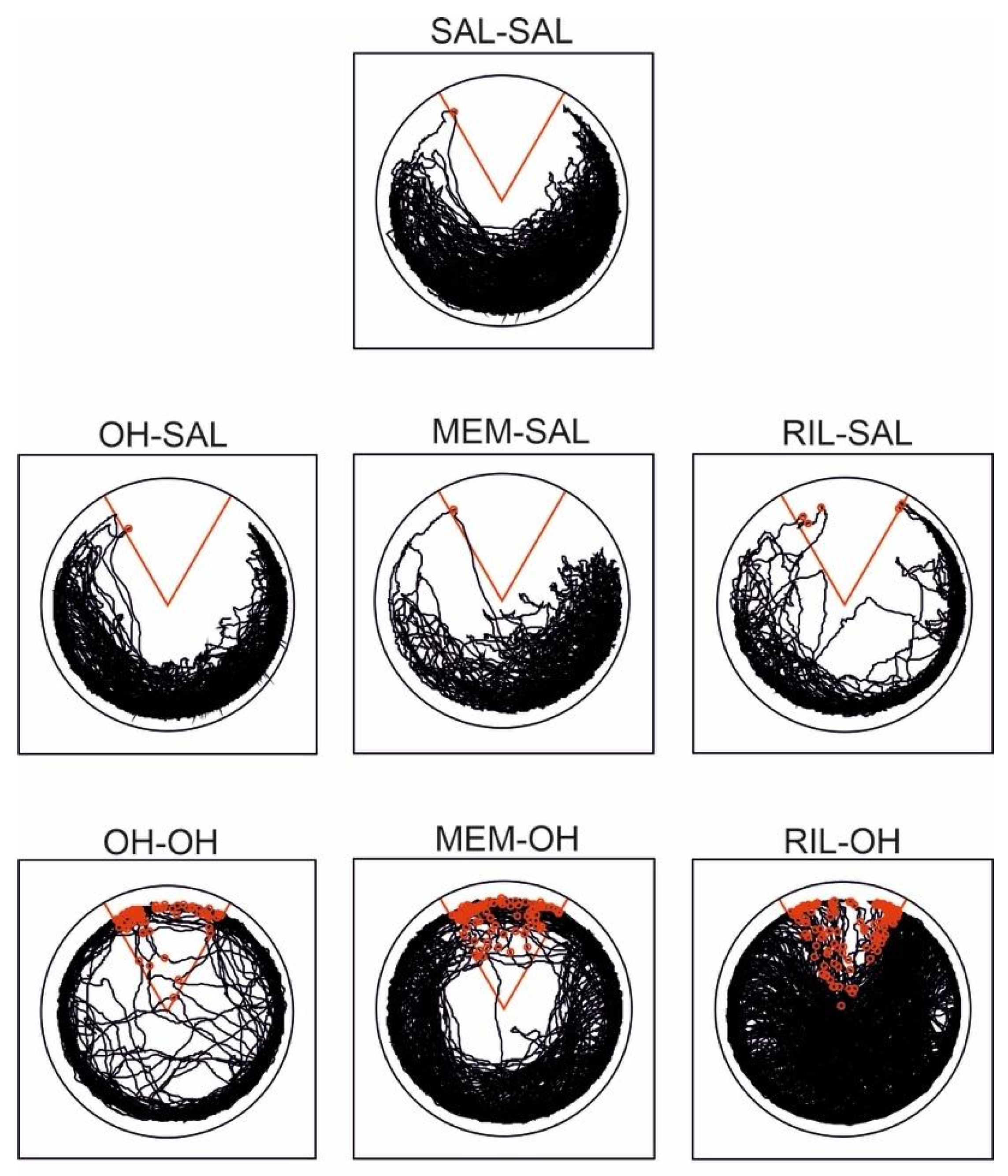
3.1. Habituation
3.2. Acquisition
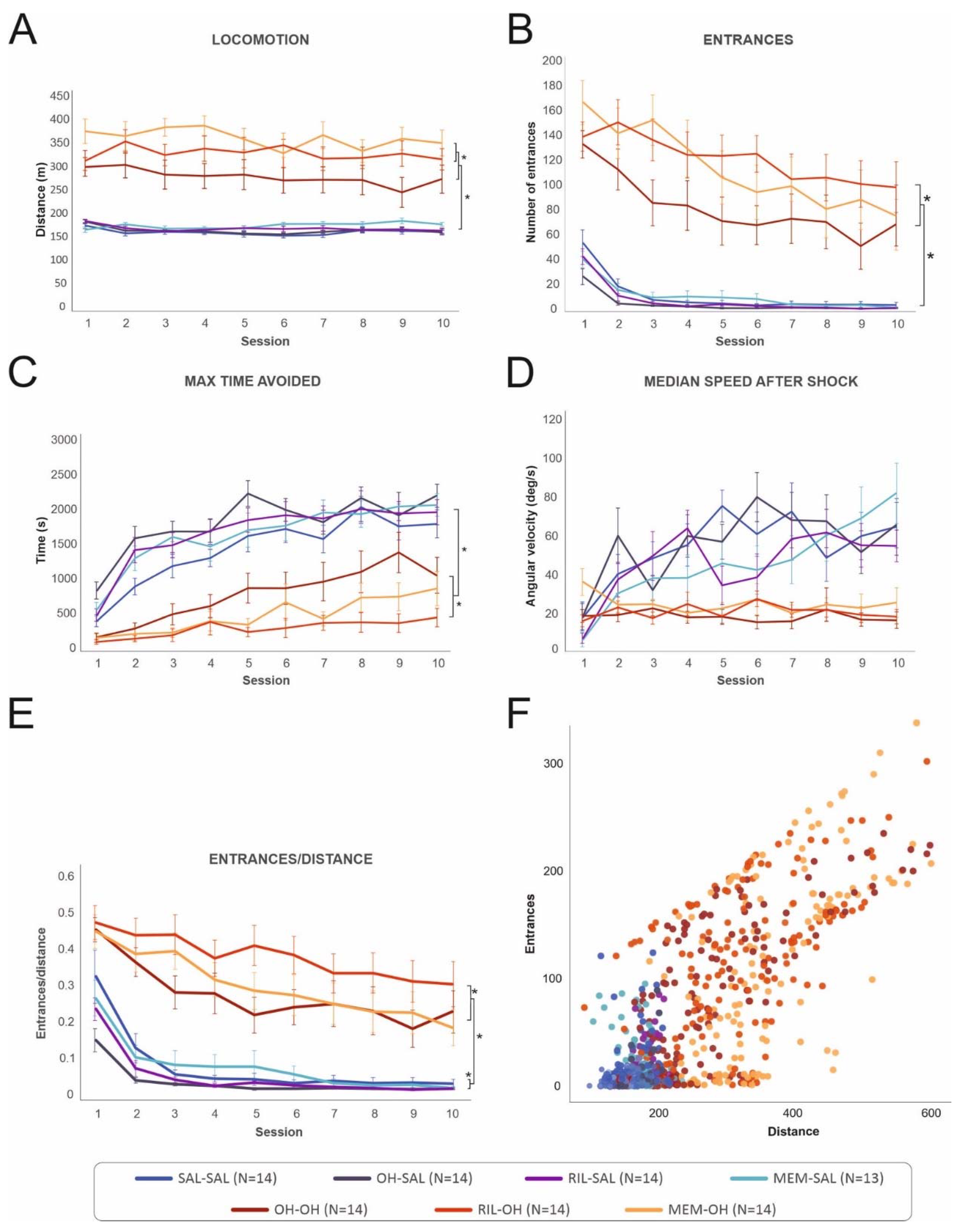
3.2.1. Locomotion
3.2.2. Entrances to the Shock Sector
3.2.3. Maximum Time Avoided
3.2.4. Median Speed after Shock
3.2.5. Entrances/Distance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stein, D.J.; Denys, D.; Gloster, A.T.; Hollander, E.; Leckman, J.F.; Rauch, S.L.; Phillips, K.A. Obsessive-compulsive Disorder: Diagnostic and Treatment Issues. Psychiatr. Clin. N. Am. 2009, 32, 665–685. [Google Scholar] [CrossRef]
- Jenike, M.A. Obsessive—Compulsive Disorder. N. Engl. J. Med. 2004, 203, 259–265. [Google Scholar] [CrossRef]
- Shin, N.Y.; Lee, T.Y.; Kim, E.; Kwon, J.S. Cognitive functioning in obsessive compulsive disorder: A meta-analysis. Psychol. Med. 2004, 44, 1121–1130. [Google Scholar] [CrossRef]
- Snyder, H.R.; Kaiser, R.H.; Warren, S.L.; Heller, W. Obsessive-compulsive disorder is associated with broad impairments in executive function: A metaanalysis. Clin. Psychol. Sci. J. Assoc. Psychol. Sci. 2015, 3, 301–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.; Young Kim, I.; Kim, J.-J. Assessment of cognitive flexibility in real life using virtual reality: A comparison of healthy individuals and schizophrenia patients. Comput. Biol. Med. 2015, 42, 841–847. [Google Scholar] [CrossRef] [PubMed]
- La Paglia, F.; la cascia, C.; Rizzo, R.; Riva, G.; La Barbera, D. Assessment of Executive Functions in Patients with Obsessive Compulsive Disorder by NeuroVR. Stud. Health Technol. Inform. 2015, 181, 98–102. [Google Scholar] [CrossRef]
- Delahaye, M.; Lemoine, P.; Cartwright, S.; Deuring, G.; Beck, J.; Pflueger, M.; Graf, M.; Hachtel, H. Learning aptitude, spatial orientation and cognitive flexibility tested in a virtual labyrinth after virtual stress induction. BMC Psychol. 2015, 3, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, W.K.; Grice, D.E.; Lapidus, K.A.B.; Coffey, B.J. Obsessive-compulsive disorder. Psychiatr. Clin. N. Am. 2014, 37, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Derksen, M.; Feenstra, M.; Willuhn, I.; Denys, D. The serotonergic system in obsessive-compulsive disorder. Handb. Behav. Neurosci. 2020, 31, 865–891. [Google Scholar] [CrossRef]
- Marazziti, D.; Hollander, E.; Lensi, P.; Ravagli, S.; Cassano, G.B. Peripheral markers of serotonin and dopamine function in obsessive-compulsive disorder. Psychiatry Res. 1992, 42, 41–51. [Google Scholar] [CrossRef]
- Erzegovesi, S.; Ronchi, P.; Smeraldi, E. 5HT-2 receptor and fluvoxamine effect in obsessive-compulsive disorder. Hum. Psychopharmacol. Clin. Exp. 1992, 7, 287–289. [Google Scholar] [CrossRef]
- Moreno, F.A.; Wiegand, C.B.; Taitano, E.K.; Delgado, P.L. Safety, tolerability, and efficacy of psilocybin in 9 patients with obsessive-compulsive disorder. J. Clin. Psychiatry 2006, 67, 1735–1740. [Google Scholar] [CrossRef] [Green Version]
- Kotapati, V.P.; Khan, A.M.; Dar, S.; Begum, G.; Bachu, R.; Adnan, M.; Zubair, A.; Ahmed, R.A. The Effectiveness of Selective Serotonin Reuptake Inhibitors for Treatment of Obsessive-Compulsive Disorder in Adolescents and Children: A Systematic Review and Meta-Analysis. Front. Psychiatry 2019, 10. [Google Scholar] [CrossRef]
- Rotge, J.Y.; Aouizerate, B.; Tignol, J.; Bioulac, B.; Burbaud, P.; Guehl, D. The glutamate-based genetic immune hypothesis in obsessive-compulsive disorder. An integrative approach from genes to symptoms. Neuroscience 2010, 165, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, K.; Bhattacharyya, S.; Christopher, R.; Khanna, S. Glutamatergic dysfunction in OCD. Neuropsychopharmacology 2005, 30, 1735–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteside, S.P.; Port, J.D.; Deacon, B.J.; Abramowitz, J.S. A magnetic resonance spectroscopy investigation of obsessive-compulsive disorder and anxiety. Psychiatry Res. Neuroimaging 2006, 146, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, D.R.; Macmaster, F.P.; Keshavan, M.S.; Fitzgerald, K.D.; Stewart, C.M.; Moore, G.J. Decrease in caudate glutamatergic concentrations in pediatric obsessive-compulsive disorder patients taking paroxetine. J. Am. Acad. Child Adolesc. Psychiatry 2000, 39, 1096–1103. [Google Scholar] [CrossRef]
- Kushner, M.G.; Kim, S.W.; Donahue, C.; Thuras, P.; Adson, D.; Kotlyar, M.; McCabe, J.; Peterson, J.; Foa, E.B. D-Cycloserine Augmented Exposure Therapy for Obsessive-Compulsive Disorder. Biol. Psychiatry 2007, 62, 835–838. [Google Scholar] [CrossRef]
- Norberg, M.M.; Krystal, J.H.; Tolin, D.F. A Meta-Analysis of D-Cycloserine and the Facilitation of Fear Extinction and Exposure Therapy. Biol. Psychiatry 2008, 63, 1118–1126. [Google Scholar] [CrossRef]
- Rodriguez, C.I.; Kegeles, L.S.; Levinson, A.; Feng, T.; Marcus, S.M.; Vermes, D.; Flood, P.; Simpson, H.B. Randomized Controlled Crossover Trial of Ketamine in Obsessive-Compulsive Disorder: Proof-of-Concept. Neuropsychopharmacology 2013, 38, 2475–2483. [Google Scholar] [CrossRef] [Green Version]
- Grant, P.; Song, J.Y.; Swedo, S.E. Review of the use of the glutamate antagonist riluzole in psychiatric disorders and a description of recent use in childhood obsessive-compulsive disorder. J. Child. Adolesc. Psychopharmacol. 2010, 20, 309–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emamzadehfard, S.; Kamaloo, A.; Paydary, K.; Ahmadipour, A.; Zeinoddini, A.; Ghaleiha, A.; Mohammadinejad, P.; Zeinoddini, A.; Akhondzadeh, S. Riluzole in augmentation of fluvoxamine for moderate to severe obsessive-compulsive disorder: Randomized, double-blind, placebo-controlled study. Psychiatry Clin. Neurosci. 2010, 70, 332–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquini, M.; Biondi, M. Memantine augmentation for refractory obsessive-compulsive disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2006, 30, 1173–1175. [Google Scholar] [CrossRef]
- Poyurovsky, M.; Weizman, R.; Weizman, A.; Koran, L. Memantine for treatment-resistant OCD. Am. J. Psychiatry 2005, 162, 2191–2192. [Google Scholar] [CrossRef]
- Kishi, T.; Matsuda, Y.; Iwata, N. Combination Therapy of Serotonin Reuptake Inhibitors and Memantine for Obsessive-Compulsive Disorder: A Meta-Analysis of Double-Blind, Randomized, Placebo-Controlled Trials. J. Alzheimer’s Dis. 2018, 64, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Modarresi, A.; Sayyah, M.; Razooghi, S.; Eslami, K.; Javadi, M.; Kouti, L. Memantine Augmentation Improves Symptoms in Serotonin Reuptake Inhibitor-Refractory Obsessive-Compulsive Disorder: A Randomized Controlled Trial. Pharmacopsychiatry 2018, 51, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Alkhatib, A.H.; Dvorkin-Gheva, A.; Szechtman, H. Quinpirole and 8-OH-DPAT induce compulsive checking behavior in male rats by acting on different functional parts of an OCD neurocircuit. Behav. Pharmacol. 2013, 24, 65–73. [Google Scholar] [CrossRef]
- Monteiro, P.; Feng, G. Learning from animal models of obsessive-compulsive disorder. In Biol. Psychiatry 2016, 79, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Odland, A.U.; Jessen, L.; Fitzpatrick, C.M.; Andreasen, J.T. 8-OH-DPAT Induces Compulsive-like Deficit in Spontaneous Alternation Behavior: Reversal by MDMA but Not Citalopram. ACS Chem. Neurosci. 2019, 10, 3094–3100. [Google Scholar] [CrossRef]
- Yadin, E.; Friedman, E.; Bridger, W.H. Spontaneous alternation behavior: An animal model for obsessive-compulsive disorder? Pharmacol. Biochem. Behav. 1991, 40, 311–315. [Google Scholar] [CrossRef]
- Bures, J.; Fenton, A.A.; Kaminsky, Y.; Zinyuk, L. Place cells and place navigation. Proc. Natl. Acad. Sci. USA 1997, 94, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Bures, J.; Fenton, A.A.; Kaminsky, Y.; Wesierska, M.; Zahalka, A. Rodent navigation after dissociation of the allocentric and idiothetic representations of space. Neuropharmacology 1998, 37, 689–699. [Google Scholar] [CrossRef]
- Stuchlík, A.; Petrásek, T.; Prokopová, I.; Holubová, K.; Hatalová, H.; Valeš, K.; Kubík, Š.; Dockery, C.; Wesierska, M. Place Avoidance Tasks as Tools in the Behavioral Neuroscience of Learning and Memory Origin of place avoidance tasks. Physiol. Res. 2013, 62, 1–19. [Google Scholar] [CrossRef]
- Janikova, M.; Brozka, H.; Radostova, D.; Svoboda, J.; Stuchlik, A. No effect of riluzole and memantine on learning deficit following quinpirole sensitization—An animal model of obsessive-compulsive disorder. Physiol. Behav. 2019, 204, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, A.; Saitoh, A.; Iwai, T.; Takahashi, K.; Yamada, M.; Sasaki-Hamada, S.; Oka, J.; Inagaki, M.; Yamada, M. Riluzole produces distinct anxiolytic-like effects in rats without the adverse effects associated with benzodiazepines. Neuropharmacology 2012, 62, 2489–2498. [Google Scholar] [CrossRef]
- Beconi, M.G.; Howland, D.; Park, L.; Lyons, K.; Giuliano, J.; Dominguez, C.; Munoz-Sanjuan, I.; Pacifici, R. Pharmacokinetics of memantine in rats and mice. PLoS Curr. Huntingt. Dis. 2012, 1, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Willis, E.F.; Bartlett, P.F.; Vukovic, J. Protocol for Short- and Longer-term Spatial Learning and Memory in Mice. Front. Behav. Neurosci. 2017, 11, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahnik, S. Carousel Maze Manager. Version 0.4.0. 2014. Available online: https://github.com/bahniks/CM_Manager_0_4_0 (accessed on 21 September 2020).
- Johnson, E.F.; Szechtman, H. A dose-response study of separate and combined effects of the serotonin agonist 8-OH-DPAT and the dopamine agonist quinpirole on locomotor sensitization, /cross-sensitization, and conditioned activity. Behav. Pharmacol. 2016, 27, 439–450. [Google Scholar] [CrossRef]
- Réus, G.Z.; Valvassori, S.S.; Machado, R.A.; Martins, M.R.; Gavioli, E.C.; Quevedo, J. Acute treatment with low doses of memantine does not impair aversive, non-associative and recognition memory in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2008, 376, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Sukhanov, I.M.; Zakharova, E.S.; Danysz, W.; Bespalov, A.Y. Effects of NMDA receptor channel blockers, MK-801 and memantine, on locomotor activity and tolerance to delay of reward in Wistar-Kyoto and spontaneously hypertensive rats. Behav. Pharmacol. 2004, 15, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, B.D.; Kratzer, U.; Schmidt, W.J. Riluzole, a glutamate release inhibitor, and motor behavior. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1998, 358, 181–190. [Google Scholar] [CrossRef]
- Wald, R.; Dodman, N.; Shuster, L. The Combined Effects of Memantine and Fluoxetine on an Animal Model of Obsessive Compulsive Disorder. Exp. Clin. Psychopharmacol. 2009, 17, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Egashira, N.; Okuno, R.; Harada, S.; Matsushita, M.; Mishima, K.; Iwasaki, K.; Fujiwara, M. Effects of glutamate-related drugs on marble-burying behavior in mice: Implications for obsessive-compulsive disorder. Eur. J. Pharmacol. 2008, 586, 164–170. [Google Scholar] [CrossRef]
- Costa, L.; Trovato, C.; Musumeci, S.A.; Catania, M.V.; Ciranna, L. 5-HT1A and 5-HT7 receptors differently modulate AMPA receptor-mediated hippocampal synaptic transmission. Hippocampus 2012, 22, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, D.; Gloveli, T.; Empson, R.M.; Draguhn, A.; Heinemann, U. Serotonin reduces synaptic excitation in the superficial medial entorhinal cortex of the rat via a presynaptic mechanism. J. Physiol. 1998, 508, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Ciranna, L. Serotonin as a Modulator of Glutamate- and GABA-Mediated Neurotransmission: Implications in Physiological Functions and in Pathology. Curr. Neuropharmacol. 2006, 4, 101–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboujaoude, E.; Barry, J.J.; Gamel, N. Memantine augmentation in treatment-resistant obsessive-compulsive disorder: An open-label trial. J. Clin. Psychopharmacol. 2009, 29, 51–55. [Google Scholar] [CrossRef]
- Pittenger, C.; Kelmendi, B.; Wasylink, S.; Bloch, M.H.; Coric, V. Riluzole augmentation in treatment-refractory obsessive-compulsive disorder: A series of 13 cases, with long-term follow-up. J. Clin. Psychopharmacol. 2008, 28, 363–367. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habituation | N | Acquisition | N | Group | Description | |
|---|---|---|---|---|---|---|
| 30 min before the Test | Before the Test | |||||
| Saline | 14 | Saline | Saline | 14 | SAL-SAL a | Global controls |
| 8-OH-DPAT | 84 | Saline | Saline | 14 | OH-SAL a | Sensitized/undrugged/untreated |
| 8-OH-DPAT | Memantine | Saline | 14 | MEM-SAL a | Sensitized/undrugged/memantine treated | |
| 8-OH-DPAT | Riluzole | Saline | 14 | RIL-SAL a | Sensitized/undrugged/riluzole treated | |
| 8-OH-DPAT | Saline | 8-OH-DPAT | 14 | OH-OH b | Sensitized/drugged/untreated | |
| 8-OH-DPAT | Memantine | 8-OH-DPAT | 14 | MEM-OH b | Sensitized/drugged/memantine treated | |
| 8-OH-DPAT | Riluzole | 8-OH-DPAT | 14 | RIL-OH b | Sensitized/drugged/riluzole treated | |
| Parameter | Treatment Group Differences | df | t | p | 95% CI |
|---|---|---|---|---|---|
| Locomotion | SAL-SAL * OH-SAL | 90 | 0.164 | 0.870 | −37.08, 43.860 |
| SAL-SAL * MEM-SAL | 90 | 0.664 | 0.509 | −27.27, 55.206 | |
| SAL-SAL * RIL-SAL | 90 | 0.368 | 0.714 | −32.86, 48,073 | |
| OH-SAL * MEM-SAL | 90 | 0.503 | 0.616 | −30.66, 51.814 | |
| OH-SAL * RIL-SAL | 90 | 0.204 | 0.839 | −36.255, 44.681 | |
| MEM-SAL * RIL-SAL | 90 | 0.302 | 0.764 | −32.86, 48.073 | |
| OH-OH * SAL-SAL | 90 | −5.819 | <0.001 | −160.63, 79.687 | |
| OH-OH * OH-SAL | 90 | −5.655 | <0.001 | −157.24, −76.295 | |
| OH-OH * MEM-OH | 90.1 | 3.919 | <0.001 | 40.45, 121.408 | |
| OH-OH * RIL-OH | 90 | 2.363 | 0.020 | 8.33, 89.271 | |
| MEM-OH * RIL-OH | 90 | −1.556 | 0.123 | −72.61, 8.341 |
| Parameter | Treatment Group Differences | df | t | p | 95% CI |
|---|---|---|---|---|---|
| Entrances | SAL-SAL * OH-SAL | 90 | −0.839 | 0.404 | −2.635, 1.06 |
| SAL-SAL * MEM-SAL | 90 | −0.009 | 0.992 | −1.890, 1.87 | |
| SAL-SAL * RIL-SAL | 90 | −0.413 | 0.681 | −2.234, 1.46 | |
| OH-SAL * MEM-SAL | 90 | −0.944 | 0.348 | −2.734, 0.957 | |
| OH-SAL * RIL-SAL | 90 | 0.814 | 0.418 | −1.099, 2.661 | |
| MEM-SAL * RIL-SAL | 90 | 0.395 | 0.694 | −1.501, 2.260 | |
| OH-OH * SAL-SAL | 90 | 5.529 | <0.001 | 3.360, 7.05 | |
| OH-OH * OH-SAL | 90 | −6.368 | <0.001 | −7.84, 4.150 | |
| OH-OH * MEM-OH | 90 | −1.82 | 0.072 | −3.56, 0.13 | |
| OH-OH * RIL-OH | 90 | 2.767 | 0.007 | 0.76, 4.45 | |
| MEM-OH * RIL-OH | 90 | 0.944 | 0.348 | −2.734, 0.957 |
| Parameter | Treatment Group Differences | df | t | p | 95% CI |
|---|---|---|---|---|---|
| Max time avoided | SAL-SAL * OH-SAL | 90 | 1.858 | 0.067 | −0.307, 11.430 |
| SAL-SAL * MEM-SAL | 90 | 0.93 | 0.355 | −3.14, 8.82 | |
| SAL-SAL * RIL-SAL | 90 | 1.155 | 0.251 | −2.411, 9.326 | |
| OH-SAL * MEM-SAL | 90 | −0.892 | 0.375 | −8.702, 3.258 | |
| OH-SAL * RIL-SAL | 90 | −0.703 | 0.484 | −7.97, 3.765 | |
| MEM-SAL * RIL-SAL | 90 | −0.203 | 0.840 | −6.598, 5.362 | |
| OH-OH * SAL-SAL | 90 | −3.696 | <0.001 | −995.5, −305.5 | |
| OH-OH * OH-SAL | 90 | 5.926 | <0.001 | 11.875, 23.61 | |
| OH-OH * MEM-OH | 90 | 1.955 | 0.054 | −0.0014, 11.725 | |
| OH-OH * RIL-OH | 90 | −3.11 | 0.002 | −15.184, −3.45 | |
| MEM-OH * RIL-OH | 90 | 1.155 | 0.251 | −2.41, 9.330 |
| Parameter | Treatment Group Differences | df | t | p | 95% CI |
|---|---|---|---|---|---|
| Median speed after | SAL-SAL * OH-SAL | 90 | 0.119 | 0.906 | −1.019, 1.150 |
| shock | SAL-SAL * MEM-SAL | 90 | −1.3996 | 0.165 | −1.897, 0.316 |
| SAL-SAL * RIL-SAL | 90 | −1.365 | 0.176 | −1.838, 0.329 | |
| OH-SAL * MEM-SAL | 90 | 0.3999 | 0.689 | −1.987, 3.006 | |
| OH-SAL * RIL-SAL | 90 | −0.021 | 0.983 | −2.476, 2.424 | |
| MEM-SAL * RIL-SAL | 90 | −0.4204 | 0.674 | −3.032, 1.961 | |
| OH-OH * SAL-SAL | 90 | 4.954 | <0.001 | 1.653, 3.817 | |
| OH-OH * OH-SAL | 90 | 5.0849 | <0.001 | 1.721, 3.880 | |
| OH-OH * MEM-OH | 90 | 1.089 | 0.279 | −0.478, 1.673 | |
| OH-OH * RIL-OH | 90 | 0.554 | 0.581 | −0.770, 1.378 | |
| MEM-OH * RIL-OH | 90 | −0.537 | 0.593 | −1.366, 0.778 |
| Parameter | Treatment Group Differences | df | t | p | 95% CI |
|---|---|---|---|---|---|
| Entrances/distance | SAL-SAL * OH-SAL | 346.8 | 3.36 | <0.001 | 0.034, 0.1141 |
| SAL-SAL * MEM-SAL | 606.9 | −0.4557 | 0.649 | −0.048, 0.0299 | |
| SAL-SAL * RIL-SAL | 443.6 | 2.463 | 0.014 | 0.0105, 0.0920 | |
| OH-SAL * MEM-SAL | 550.3 | 3.297 | 0.001 | 0.0264, 0.1037 | |
| OH-SAL * RIL-SAL | 529.5 | 1.151 | 0.250 | −0.0161, 0.0618 | |
| MEM-SAL * RIL-SAL | 819.5 | 2.1692 | 0.030 | 0.0041, 0.0803 | |
| OH-OH * SAL-SAL | 440.3 | −9.714 | <0.001 | −0.2381, −0.1581 | |
| OH-OH * OH-SAL | 405.3 | −13.552 | <0.001 | −0.312, −0.233 | |
| OH-OH * MEM-OH | 777.2 | 1.1267 | 0.260 | −0.0157, 0.0581 | |
| OH-OH * RIL-OH | 782.3 | 4.321 | <0.001 | 0.0437, 0.1164 | |
| MEM-OH * RIL-OH | 731.5 | 2.995 | 0.003 | 0.0203, 0.0974 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janikova, M.; Mainerova, K.; Vojtechova, I.; Petrasek, T.; Svoboda, J.; Stuchlik, A. Memantine and Riluzole Exacerbate, Rather Than Ameliorate Behavioral Deficits Induced by 8-OH-DPAT Sensitization in a Spatial Task. Biomolecules 2021, 11, 1007. https://doi.org/10.3390/biom11071007
Janikova M, Mainerova K, Vojtechova I, Petrasek T, Svoboda J, Stuchlik A. Memantine and Riluzole Exacerbate, Rather Than Ameliorate Behavioral Deficits Induced by 8-OH-DPAT Sensitization in a Spatial Task. Biomolecules. 2021; 11(7):1007. https://doi.org/10.3390/biom11071007
Chicago/Turabian StyleJanikova, Martina, Karolina Mainerova, Iveta Vojtechova, Tomas Petrasek, Jan Svoboda, and Ales Stuchlik. 2021. "Memantine and Riluzole Exacerbate, Rather Than Ameliorate Behavioral Deficits Induced by 8-OH-DPAT Sensitization in a Spatial Task" Biomolecules 11, no. 7: 1007. https://doi.org/10.3390/biom11071007