
Hypolipidemic Effects of Beetroot Juice in SHR-CRP and HHTg Rat Models of Metabolic Syndrome: Analysis of Hepatic Proteome
 , , , , and
, , , , and
Abstract
:1. Introduction
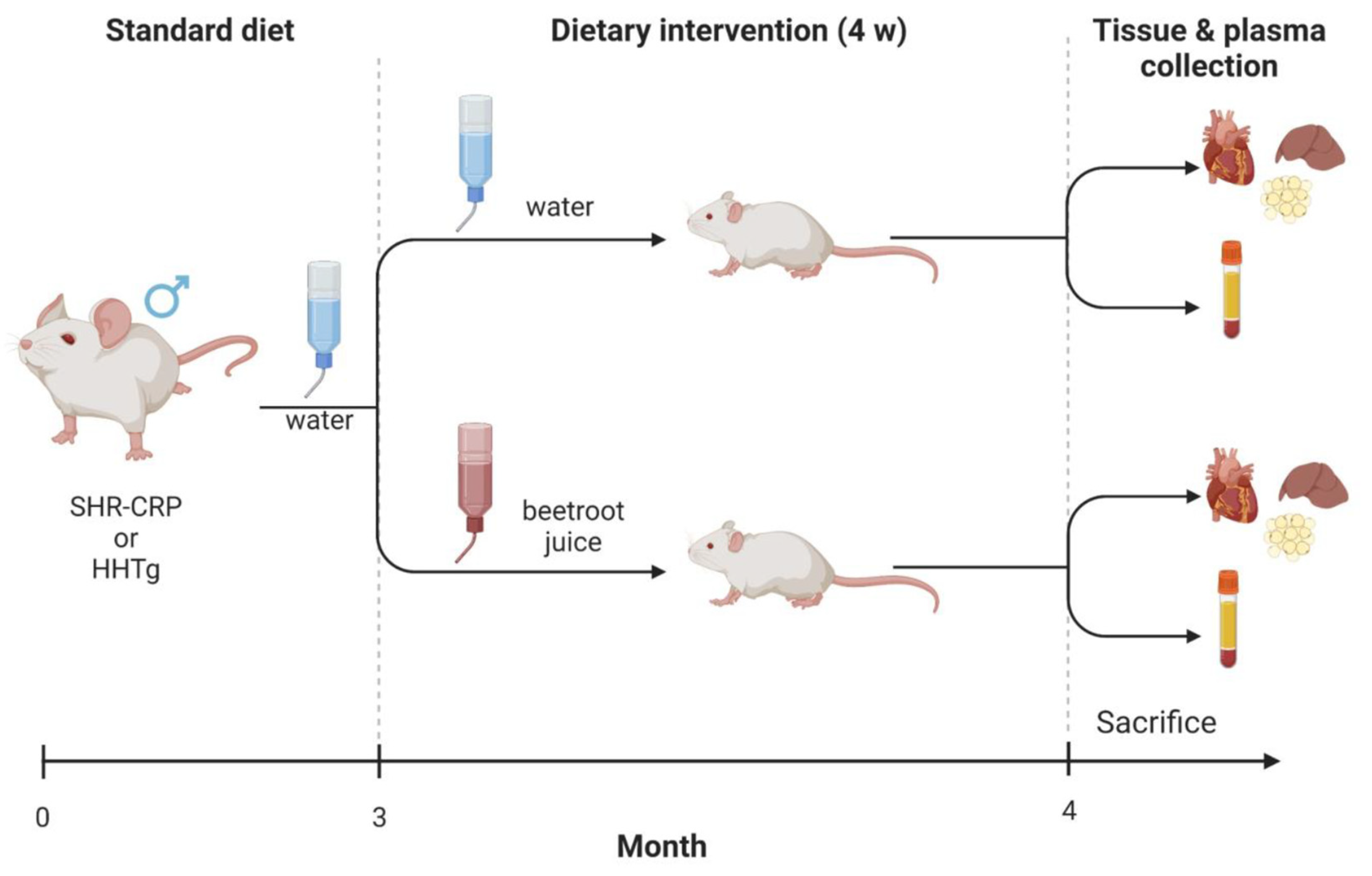
2. Experimental Design
2.1. Animals
2.2. Biochemical Analyses
2.3. Measurement of Plasma and Tissue cGMP Concentrations
2.4. Tissue Triglyceride and Cholesterol Measurements
2.5. Parameters of Insulin Sensitivity in Skeletal Muscle and Adipose Tissue
2.6. Parameters of Oxidative Stress
2.7. Proteomic Analysis
2.8. Statistical Analysis
3. Results
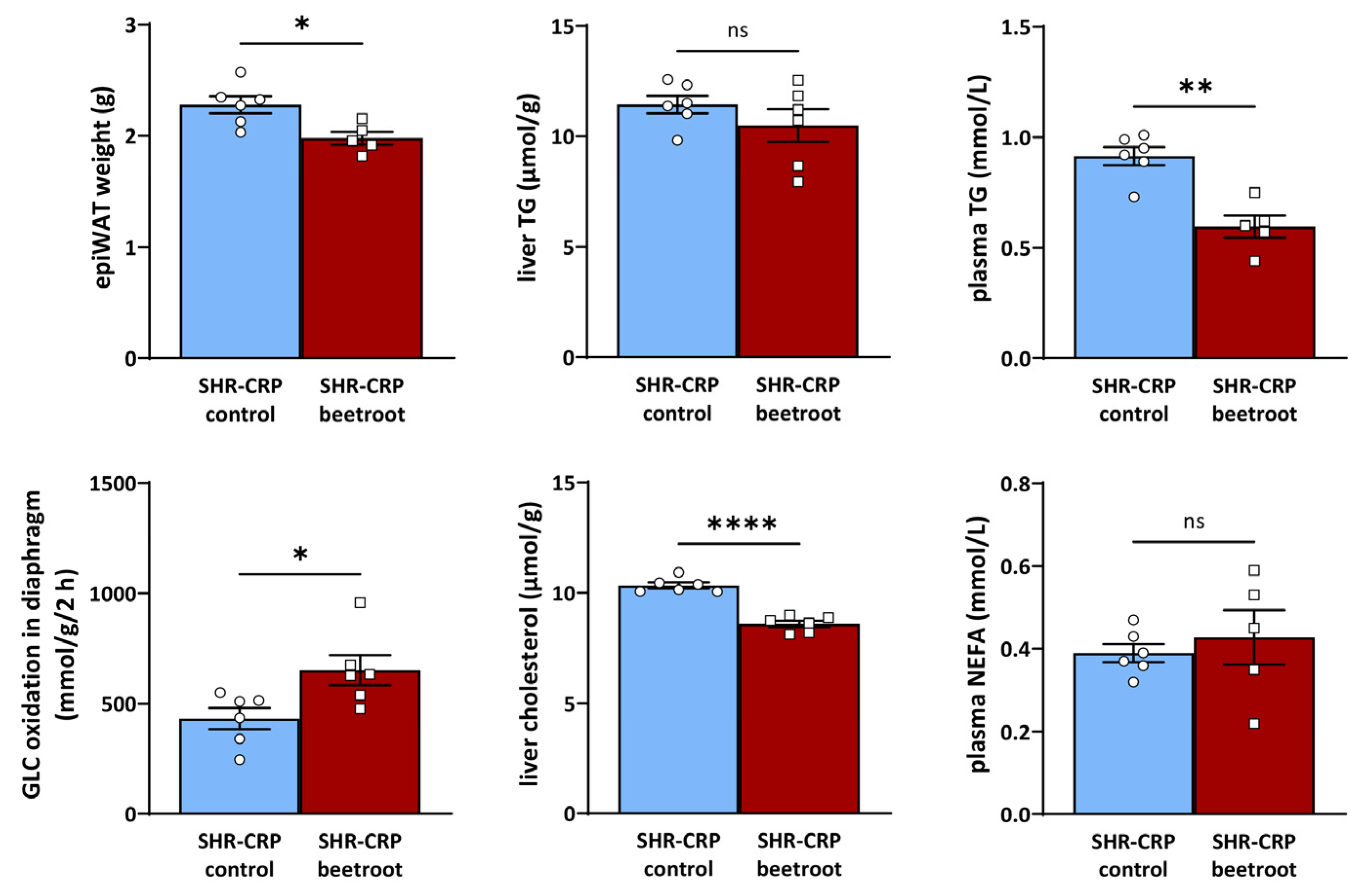
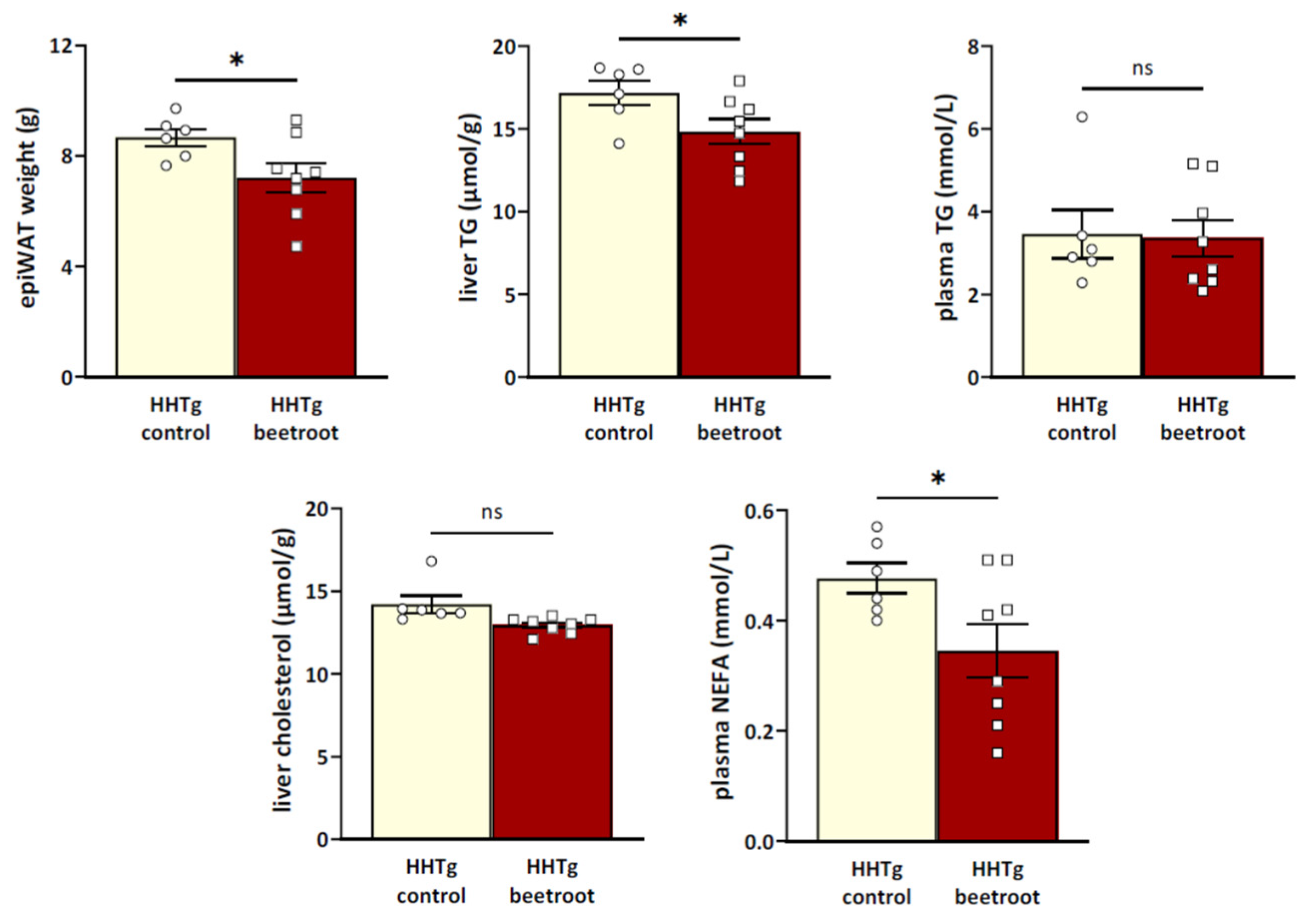
3.1. The Effects of Beetroot Juice on Parameters of Glucose and Lipid Metabolism
3.2. The Effects of Beetroot Juice on cGMP in Serum and Tissues
3.3. The Effects of Beetroot Juice on Inflammatory and Oxidative Stress Parameters
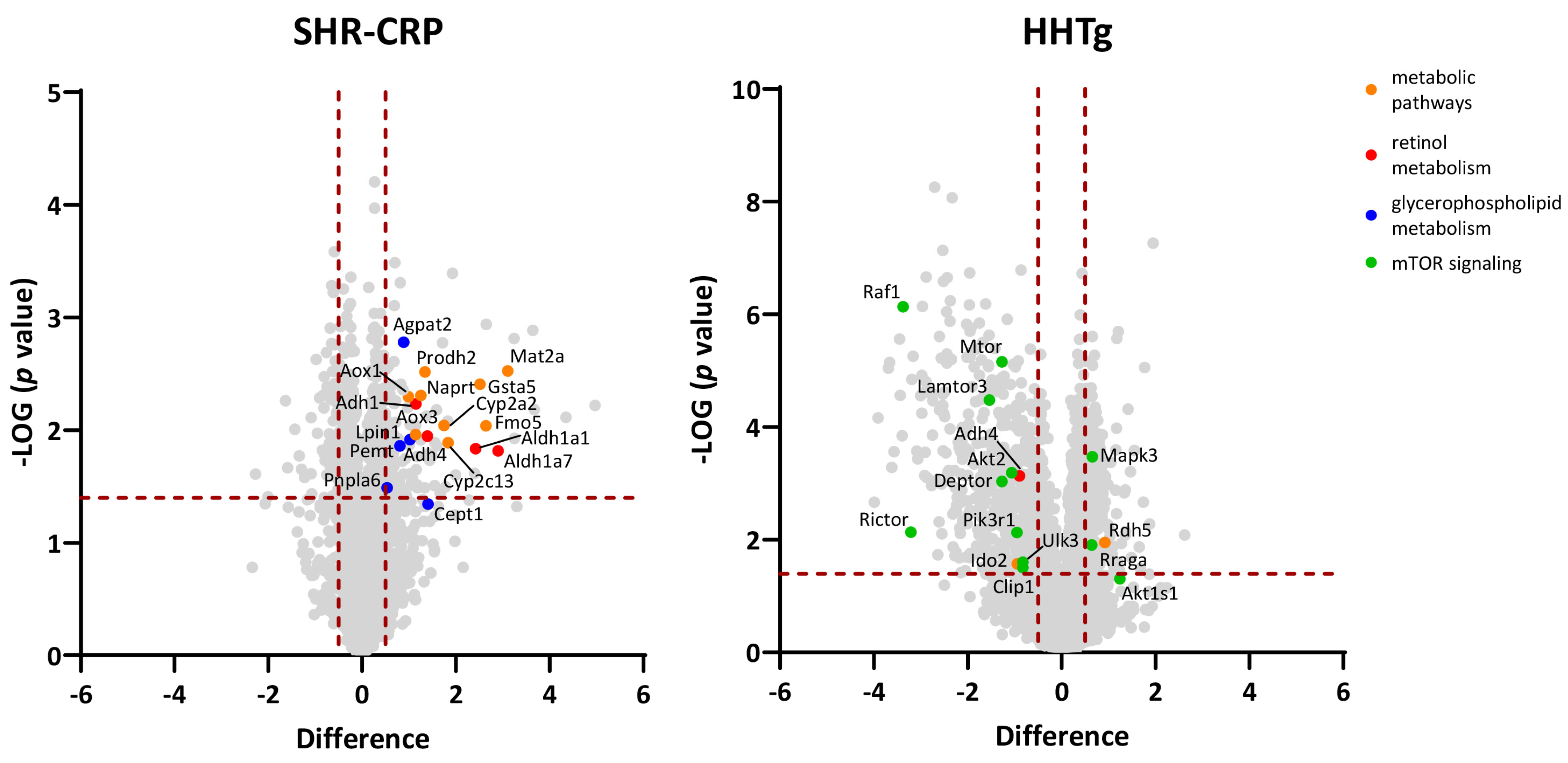
3.4. Analysis of Hepatic Proteome
4. Discussion
4.1. Increased Hepatic Expression of Enzymes Regulating Glycerophospholipid Metabolism in SHR-CRP Rats after Treatment with Beetroot
4.2. Increased Hepatic Expression of Enzymes Regulating S-Adenosylmethionine (SAM) Biosynthesis in SHR-CRP Rats after Treatment with Beetroot
4.3. Increased Hepatic Expression of Enzymes Regulating Nicotinamide Adenine Dinucleotide (NAD+) Metabolism in SHR-CRP Rats after Treatment with Beetroot
4.4. Increased Hepatic Expression of Cytochromes P450, Enzymes Regulating Glutathiome Metabolism and Retinol Metabolism in SHR-CRP Rats after Treatment with Beetroot
4.5. Reduced Expression of Proteins Involved in mTOR Signalling in HHTg Rats Treated with Beetroot
4.6. Reduced Expression of Proteins Involved in Cellular Trafficking in HHTg Rats Treated with Beetroot
4.7. Reduced Expression of Proteins Involved in Inflammation and Apoptosis in HHTg Rats Treated with Beetroot
4.8. Molecular Mechanisms of Beetroot Effects on Oxidative Stress, Inflammation, and Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versari, D.; Daghini, E.; Virdis, A.; Ghiadoni, L.; Taddei, S. Endothelial dysfunction as a target for prevention of cardiovascular disease. Diabetes Care 2009, 32 (Suppl. S2), S314–S321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadipour, E.; Taleghani, A.; Tayarani-Najaran, N.; Tayarani-Najaran, Z. Biological effects of red beetroot and betalains: A review. Phytother. Res. 2020, 34, 1847–1867. [Google Scholar] [CrossRef] [PubMed]
- Milton-Laskibar, I.; Martínez, J.A.; Portillo, M.P. Current knowledge on beetroot bioactive compounds: Role of nitrate and betalains in health and disease. Foods 2021, 10, 1314. [Google Scholar] [CrossRef]
- El Gamal, A.A.; AlSaid, M.S.; Raish, M.; Al-Sohaibani, M.; Al-Massarani, S.M.; Ahmad, A.; Hefnawy, M.; Al-Yahya, M.; Omer, A.; Basoudan, O.A.; et al. Beetroot (Beta vulgaris L.) extract ameliorates gentamicin-induced nephrotoxicity associated oxidative stress, inflammation, and apoptosis in rodent model. Mediat. Inflamm. 2014, 2014, 983952. [Google Scholar] [CrossRef] [Green Version]
- Vulic, J.J.; Cebovic, T.N.; Čanadanovic-Brunet, J.M.; Cetkovic, G.S.; Čanadanovic, V.M.; Djilas, S.M.; Šaponjac, V.T. In vivo and in vitro antioxidant effects of beetroot pomace extracts. J. Function. Foods 2014, 6, 168–175. [Google Scholar] [CrossRef]
- Albasher, G.; Almeer, R.; Al-Otibi, F.O.; Al-Kubaisi, N.; Mahmoud, A.M. Ameliorative effect of Beta vulgaris root extract on chlorpyrifos-induced oxidative stress, inflammation and liver injury in rats. Biomolecules 2019, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Raish, M.; Ahmad, A.; Ansari, M.A.; Alkharfy, K.M.; Ahad, A.; Khan, A.; Ali, N.; Ganaie, M.A.; Hamidaddin, M.A.A. Beetroot juice alleviates isoproterenol-induced myocardial damage by reducing oxidative stress, inflammation, and apoptosis in rats. 3 Biotech. 2019, 9, 147. [Google Scholar] [CrossRef]
- Morris, R.C., Jr.; Pravenec, M.; Šilhavý, J.; DiCarlo, S.E.; Kurtz, T.W. Small amounts of inorganic nitrate or beetroot provide substantial protection from salt-induced increases in blood pressure. Hypertension 2019, 73, 1042–1048. [Google Scholar] [CrossRef]
- Webb, A.J.; Patel, N.; Loukogeorgakis, S.; Okorie, M.; Aboud, Z.; Misra, S.; Rashid, R.; Miall, P.; Deanfield, J.; Benjamin, N.; et al. Acute blood pressure lowering, vasoprotective, and antiplatelet properties of dietary nitrate via bioconversion to nitrite. Hypertension 2008, 51, 784–790. [Google Scholar] [CrossRef]
- Coles, L.T.; Clifton, P.M. Effect of beetroot juice on lowering blood pressure in free-living, disease-free adults: A randomized, placebo-controlled trial. Nutr. J. 2012, 11, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wroblewska, M.; Juskiewicz, J.; Wiczkowski, W. Physiological properties of beetroot crisps applied in standard and dyslipidaemic diets of rats. Lipid. Health Dis. 2011, 10, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabeh, M.N.; Ibrahim, E.M. Antihypercholesterolemic effects of beet (Beta vulgaris L.) root waste extract on hypercholesterolemic rats and its antioxidant potential properties. Pak. J. Nutr. 2014, 13, 500. [Google Scholar] [CrossRef] [Green Version]
- Al-Dosari, M.; Alqasoumi, S.A.; Ahmed, M.; Al-Yahya, M.; Ansari, M.N.; Rafatullah, S. Effect of Beta vulgaris L. on cholesterol rich diet induced hypercholesterolemia in rats. Farmacia 2011, 59, 669–678. [Google Scholar]
- Singh, A.; Verma, S.K.; Singh, V.K.; Nanjappa, C.; Roopa, N.; Raju, P.S.; Singh, S.N. Beetroot juice supplementation increases high density lipoprotein-cholesterol and reduces oxidative stress in physically active individuals. J. Pharm. Nutr. Sci. 2015, 5, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Kerley, C.P.; Dolan, E.; Cormican, L. Nitrate-rich beetroot juice selectively lowers ambulatory pressures and LDL cholesterol in uncontrolled but not controlled hypertension: A pilot study. Ir. J. Med. Sci. 2017, 186, 895–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, T.; Sato, K. Effects of beet red inhibiting experimental diabetes in vivo-studies using ALS strain mice. Foods Food Ingred. J. Jpn. 2008, 213, 122. [Google Scholar]
- Shetty, A.K.; Rashmi, R.; Rajan, M.G.; Sambaiah, K.; Salimath, P.V. Antidiabetic influence of quercetin in streptozotocin-induced diabetic rats. Nutr. Res. 2004, 24, 373–381. [Google Scholar] [CrossRef]
- Lugo-Radillo, A.; Delgado-Enciso, I.; Peña-Beltrán, E. Betanidin significantly reduces blood glucose levels in BALB/c mice fed with an atherogenic diet. Nat. Prod. Bioprospect. 2012, 2, 154–155. [Google Scholar] [CrossRef] [Green Version]
- Pravenec, M.; Kajiya, T.; Zídek, V.; Landa, V.; Mlejnek, P.; Šimáková, M.; Šilhavý, J.; Malínská, H.; Oliyarnyk, O.; Kazdová, L.; et al. Effects of human C-reactive protein on pathogenesis of features of the metabolic syndrome. Hypertension 2011, 57, 731–737. [Google Scholar] [CrossRef] [Green Version]
- Zicha, J.; Pechánová, O.; Cacányiová, S.; Cebová, M.; Kristek, F.; Török, J.; Simko, F.; Dobešová, Z.; Kuneš, J. Hereditary hypertriglyceridemic rat: A suitable model of cardiovascular disease and metabolic syndrome? Physiol. Res. 2006, 55 (Suppl. S1), S49–S63. [Google Scholar] [CrossRef] [PubMed]
- Vrána, A.; Kazdová, L. The hereditary hypertriglyceridemic nonobese rat: An experimental model of human hypertriglyceridemia. Transplant. Proc. 1990, 22, 2579. [Google Scholar] [PubMed]
- Hüttl, M.; Marková, I.; Miklanková, D.; Zapletalová, I.; Poruba, M.; Haluzik, M.; Vaněčková, I.; Malinská, H. In a prediabetic model, empagliflozin improves hepatic lipid metabolism independently of obesity and before onset of hyperglycemia. Int. J. Mol. Sci. 2021, 22, 11513. [Google Scholar] [CrossRef] [PubMed]
- Malinská, H.; Hüttl, M.; Oliyarnyk, O.; Marková, I.; Poruba, M.; Racová, Z.; Kazdová, L.; Večeřa, R. Beneficial effects of troxerutin on metabolic disorders in non-obese model of metabolic syndrome. PLoS ONE 2019, 14, e0220377. [Google Scholar] [CrossRef] [Green Version]
- Johnston, H.E.; Yadav, K.; Kirkpatrick, J.M.; Biggs, G.S.; Oxley, D.; Kramer, H.B.; Samant, R.S. Solvent precipitation SP3 (SP4) enhances recovery for proteomics sample preparation without magnetic beads. Anal. Chem. 2022, 94, 10320–10328. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Method. 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Lutkewitte, A.J.; Finck, B.N. Regulation of signaling and metabolism by lipin-mediated phosphatidic acid phosphohydrolase activity. Biomolecules 2020, 10, 1386. [Google Scholar] [CrossRef]
- Cortés, V.A.; Curtis, D.E.; Sukumaran, S.; Shao, X.; Parameswara, V.; Rashid, S.; Smith, A.R.; Ren, J.; Esser, V.; Hammer, R.E.; et al. Molecular mechanisms of hepatic steatosis and insulin resistance in the AGPAT2-deficient mouse model of congenital generalized lipodystrophy. Cell Metab. 2009, 9, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Péterfy, M.; Phan, J.; Xu, P.; Reue, K. Lipodystrophy in the fld mouse results from mutation of a new gene encoding a nuclear protein, lipin. Nat. Genet. 2001, 27, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Rui, B.B.; Tang, L.Y.; Hu, C.M. Lipin family proteins--key regulators in lipid metabolism. Ann. Nutr. Metab. 2015, 66, 10–18. [Google Scholar] [CrossRef] [PubMed]
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef]
- Kim, G.-T.; Kim, S.-J.; Park, S.-H.; Lee, D.; Park, T.-S. Hepatic expression of the serine palmitoyltransferase subunit Sptlc2 reduces lipid droplets in the liver by activating VLDL secretion. J. Lipid Atheroscler. 2020, 9, 291–303. [Google Scholar] [CrossRef]
- Watkins, S.M.; Zhu, X.; Zeisel, S.H. Phosphatidylethanolamine-N-methyltransferase activity and dietary choline regulate liver-plasma lipid flux and essential fatty acid metabolism in mice. J. Nutr. 2003, 133, 3386–3391. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Jackson, P.; Gunter, C.; Wang, J.; Rock, C.O.; Jackowski, S. Placental thrombosis and spontaneous fetal death in mice deficient in ethanolamine kinase 2. J. Biol. Chem. 2006, 281, 28438–28449. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Guo, M.; Wang, Q.; Wang, Q.; Zuo, S.; Zhang, X.; Tong, H.; Chen, J.; Wang, H.; Chen, X.; et al. The patatin-like phospholipase domain containing protein 7 facilitates VLDL secretion by modulating ApoE stability. Hepatology 2020, 72, 1569–1585. [Google Scholar] [CrossRef]
- Mato, J.M.; Martínez-Chantar, M.L.; Lu, S.C. S-adenosylmethionine metabolism and liver disease. Ann. Hepatol. 2013, 12, 183–189. [Google Scholar] [CrossRef]
- Audrito, V.; Messana, V.G.; Deaglio, S. NAMPT and NAPRT: Two metabolic enzymes with key roles in inflammation. Front. Oncol. 2020, 10, 358. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, M.; Kanda, T.; Urai, H.; Kurokochi, A.; Kitahama, R.; Shigaki, S.; Ono, T.; Yukioka, H.; Hasegawa, K.; Tokuyama, H.; et al. NNMT activation can contribute to the development of fatty liver disease by modulating the NAD+ metabolism. Sci. Rep. 2018, 8, 8637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, C.J.; Otto, D.M.E.; Carrie, D.; Magnuson, M.A.; McLaren, A.W.; Rosewell, I.; Wolf, C.R. Inactivation of the hepatic cytochrome P450 system by conditional deletion of hepatic cytochrome P450 reductase. J. Biol. Chem. 2003, 278, 13480–13486. [Google Scholar] [CrossRef]
- Jin, Y.; Tan, Y.; Zhao, P.; Guo, Y.; Chen, S.; Wu, J.; Ren, Z. Glutathione S-transferase Mu 2 inhibits hepatic steatosis via ASK1 suppression. Commun. Biol. 2022, 5, 326. [Google Scholar] [CrossRef] [PubMed]
- Molotkov, A.; Fan, X.; Duester, G. Excessive vitamin A toxicity in mice genetically deficient in either alcohol dehydrogenase Adh1 or Adh3. Eur. J. Biochem. 2002, 269, 2607–2612. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, R.S.; Orena, S.J.; Rafidi, K.; Torchia, A.J.; Stock, J.L.; Hildebrandt, A.I.; Coskran, T.; Black, S.C.; Brees, D.J.; Wicks, J.R.; et al. Severe diabetes, age-dependent loss of adipose tissue, and mild growth deficiency in mice lacking Akt2/PKB beta. J. Clin. Investig. 2003, 112, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Y.; Sabatini, D.M. mTOR at the nexus of nutrition, growth, ageing and disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef]
- Ricoult, S.J.H.; Manning, B.D. The multifaceted role of mTORC1 in the control of lipid metabolism. EMBO Rep. 2013, 14, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Caron, A.; Mouchiroud, M.; Gautier, N.; Labbé, S.M.; Villot, R.; Turcotte, L.; Secco, B.; Lamoureux, G.; Shum, M.; Gélinas, Y.; et al. Loss of hepatic DEPTOR alters the metabolic transition to fasting. Mol. Metab. 2017, 6, 447–458. [Google Scholar] [CrossRef]
- Yuan, M.; Pino, E.; Wu, L.; Kacergis, M.; Soukas, A.S. Identification of Akt-independent regulation of hepatic lipogenesis by mammalian target of rapamycin (mTOR) complex 2. J. Biol. Chem. 2012, 287, 29579–29588. [Google Scholar] [CrossRef] [Green Version]
- Csaki, L.S.; Dwyer, J.R.; Li, X.; Nguyen, M.H.; Dewald, J.; Brindley, D.N.; Lusis, A.J.; Yoshinaga, Y.; de Jong, P.; Fong, L.; et al. Lipin-1 and lipin-3 together determine adiposity in vivo. Mol. Metab. 2013, 3, 145–154. [Google Scholar] [CrossRef]
- Rai, P.; Kumar, M.; Sharma, G.; Barak, P.; Das, S.; Kamat, S.S.; Mallika, R. Kinesin-dependent mechanism for controlling triglyceride secretion from the liver. Proc. Natl. Acad. Sci. USA 2017, 114, 12958–12963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.; Sanghavi, P.; Mallik, R. Microtubule motor driven interactions of lipid droplets: Specificities and opportunities. Front. Cell Dev. Biol. 2022, 10, 893375. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Akama, T.; Jiang, Y.; Chun, T.-H. The exocyst complex regulates free fatty acid uptake by adipocytes. PLoS ONE 2015, 10, e0120289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feitosa, M.F.; Wojczynski, M.K.; North, K.E.; Zhang, Q.; Province, M.A.; Carr, J.J.; Borecki, I.B. The ERLIN1-CHUK-CWF19L1 gene cluster influences liver fat deposition and hepatic inflammation in the NHLBI Family Heart Study. Atherosclerosis 2013, 228, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Al-Harbi, L.N.; Alshammari, G.M.; Al-Dossari, A.M.; Subash-Babu, P.; Binobead, M.A.; Alhussain, M.H.; AlSedairy, S.A.; Al-Nouri, D.M.; Shamlan, G. Beta vulgaris L. (Beetroot) methanolic extract prevents hepatic steatosis and liver damage in T2DM rats by hypoglycemic, insulin-sensitizing, antioxidant effects, and upregulation of PPARα. Biology 2021, 10, 1306. [Google Scholar] [CrossRef]
- Abedimanesh, N.; Asghari, S.; Mohammadnejad, K.; Daneshvar, Z.; Rahmani, S.; Shokoohi, S.; Farzaneh, A.H.; Hosseini, S.H.; Jafari Anarkooli, I.; Noubarani, M.; et al. The anti-diabetic effects of betanin in streptozotocin-induced diabetic rats through modulating AMPK/SIRT1/NF-κB signaling pathway. Nutr. Metab. 2021, 18, 92. [Google Scholar] [CrossRef]
- Romero, S.A.; Pavan, I.C.B.; Morelli, A.P.; Mancini, M.C.; da Silva, L.G.S.; Fagundes, I.; Silva, C.H.R.; Ponte, L.G.S.; Rostagno, M.A.; Bezerra, R.M.N.; et al. Anticancer effects of root and beet leaf extracts (Beta vulgaris L.) in cervical cancer cells (HeLa). Phytother. Res. 2021, 35, 6191–6203. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | SHR-CRP Control | SHR-CRP Beetroot | HHTG Control | HHTG Beetroot |
|---|---|---|---|---|
| Body weight (g) | 315 ± 6 | 306 ± 8 | 426 ± 6 | 427 ± 9 |
| Serum cholesterol (mmol/L) | 1.45 ± 0.06 | 1.47 ± 0.04 | 1.63 ± 0.10 | 1.58 ± 0.06 |
| Serum HDL cholesterol (mmol/L) | 1.22 ± 0.04 | 1.28 ± 0.02 | 0.785 ± 0.062 | 0.848 ± 0.039 |
| Non-fasting glucose (mmol/L) | 7.06 ± 0.10 | 6.95 ± 0.17 | 9.38 ± 0.41 | 9.09 ± 0.32 |
| Serum insulin (nmol/L) | 0.247 ± 0.011 | 0.237 ± 0.030 | 0.178 ± 0.022 | 0.219 ± 0.029 |
| M. gastrocnemius triglycerides (μmol/g) | 2.03 ± 0.21 | 1.98 ± 0.62 | 3.92 ± 0.67 | 3.70 ± 0.41 |
| Heart triglycerides (μmol/g) | 1.91 ± 0.07 | 1.40 ± 0.16 * | 3.52 ± 0.45 | 3.55 ± 0.38 |
| Basal lipogenesis (nmol gL./g/2 h) | 1009 ± 79 | 1726 ± 332 | 210 ± 15 | 230 ± 15 |
| Stimulated lipogenesis (nmol gL./g/2 h) | 1436 ± 148 | 2339 ± 507 | 241 ± 16 | 239 ± 20 |
| Basal glycogenesis (nmol gL./g/2 h) | 996 ± 182 | 1089 ± 174 | ND | ND |
| Stimulated glycogenesis (nmol gL./g/2 h) | 1548 ± 175 | 1358 ± 241 | ND | ND |
| Plasma cGMP (pmol/L) | 8.84 ± 0.64 | 13.41 ± 1.22 * | ND | ND |
| cGMP in aorta (pmol/g) | 40.66 ± 8.88 | 59.35 ± 27.57 | ND | ND |
| cGMP in myocardium (pmol/g) | 12.25 ± 1.08 | 23.37 ± 1.44 * | ND | ND |
| Trait | SHR-CRP Control | SHR-CRP Beetroot | HHTg Control | HHTg Beetroot |
|---|---|---|---|---|
| rat hsCRP (mg/mL) | 0.375 ± 0.018 | 0.329 ± 0.039 | 0.675 ± 0.030 | 0.729 ± 0.049 |
| human CRP (mg/L) | 163 ± 6 | 184 ± 19 | u.d. | u.d. |
| MCP-1 (pg/mL) | 6.96 ± 0.30 | 5.81 ± 0.36 | 3.29 ± 0.21 | 3.68 ± 0.16 |
| IL6 (pg/mL) | 39.13 ± 7.36 | 20.83 ± 6.17 | 44.27 ± 10.12 | 48.35 ± 13.79 |
| leptin (ng/mL) | 3.07 ± 0.22 | 5.17 ± 0.67 | 13.80 ± 1.00 | 11.56 ± 1.13 |
| TNFα (pg/mL) | 4.97 ± 0.32 | 4.76 ± 0.38 | 5.58 ± 0.53 | 6.36 ± 0.38 |
| Trait | SHR-CRP Control | SHR-CRP Beetroot | HHTg Control | HHTg Beetroot |
|---|---|---|---|---|
| SOD (U/mg protein) | 0.124 ± 0.011 | 0.127 ± 0.007 | 0.111 ± 0.005 | 0.130 ± 0.004 * |
| GSH-Px (μM NADPH/min/mg protein) | 271 ± 17 | 300 ± 22 | 211 ± 36 | 419 ± 48 ** |
| GR (μM NADPH/min/mg protein) | 123 ± 9 | 153 ± 15 | 73 ± 7 | 85 ± 5 |
| CAT (mM H2O2/min/mg protein) | 1603 ± 188 | 2274 ± 94 * | 1365 ± 23 | 1500 ± 50 * |
| GSH (μmol/mg protein) | 52.09 ± 3.60 | 58.09 ± 1.88 | 61.14 ± 1.62 | 61.12 ± 3.43 |
| GSSG(μmol/mg protein) | 4.08 ± 0.44 | 4.42 ± 0.45 | 3.01 ± 0.22 | 2.80 ± 0.19 |
| GSH/GSSG | 12.96 ± 0.41 | 13.33 ± 0.97 | 20.92 ± 1.83 | 22.33 ± 1.74 |
| Conjugated dienes (nM/mg protein) | 41.5 ± 3.3 | 30.2 ± 2.3 * | ||
| TBARS/MDA (nM/mg protein) | 1.73 ± 0.17 | 1.63 ± 0.13 | 1.82 ± 0.11 | 1.89 ± 0.12 |
| KEGG Pathway | Counts | p-Value | Benjamini |
|---|---|---|---|
| Metabolic pathways | 65 | 3.0 × 10−19 | 6.2 × 10−17 |
| ↑Aox1, ↑Aox3, ↑Akr1d1, ↑Amdhd1, ↑Aoc3, ↑Car14, ↑Cept1, ↑Cyp2a1, ↑Cyp2a2, ↑Cyp2c13, ↑Cyp2c7, ↑Cyp4a2, ↑Cox6a1, ↑Cox6b1, ↑Dpys, ↑Ephx2, ↑Etnk2, ↑Flad1, ↑Fmo5, ↑Fcsk, ↑Ggt5, ↑Gne, ↑Gsta2, ↑Gsta5, ↑Gstm3, ↑Gstm4, ↑Hykk, ↓Hsd17b2, ↓Ido2, ↓Inmt, ↓Impa2, ↑Lpin1, ↑Mat2a, ↓Mocs2, ↑Nnmt, ↑Naprt, ↑Pemt, ↑Pde8a, ↓Plod1, ↑Prodh2, ↑Rdh5, ↓Sptlc2, ↑Srd5a1, ↑Sdhd, ↑Tat, ↑Urad, ↑Vnn3 | |||
| Retinol metabolism | 14 | 7.1 × 10−11 | 7.4 × 10−9 |
| ↑Ugt1a7, ↑Adh1, ↑Adh4, ↑Aldh1a1, ↑Aldh1a7, ↑Aox1, ↑Aox3, ↑Cyp2a1, ↑Cyp2a2, ↑Cyp2c13, ↑Cyp2c7, ↑Cyp4a2 | |||
| Steroid hormone biosynthesis | 8 | 1.0 × 10−4 | 3.6 × 10−3 |
| ↑Ugt1a7, ↑Akr1d1, ↑Cyp2c13, ↑Cyp2c7, ↑Cyp2d3, ↓Hsd17b2, ↑Srd5a1 | |||
| Tyrosine metabolism | 6 | 1.3 × 10−4 | 3.8 × 10−3 |
| ↑Adh1, ↑Adh4, ↑Aox1, ↑Aox3, ↑Aoc3, ↑Tat | |||
| Glycerophospholipid metabolism | 7 | 1.8 × 10−3 | 4.2 × 10−2 |
| ↑Agpat2, ↑Cept1, ↑Lpin1, ↑Pnpla6, ↓Pnpla7, ↑Pemt | |||
| Glutathione metabolism | 6 | 2.4 × 10−3 | 4.9 × 10−2 |
| ↑Ggt5, ↑Gsta2, ↑Gsta5, ↑Gstm3, ↑Gstm4 | |||
| KEGG Pathway | Counts | p-Value | Benjamini |
|---|---|---|---|
| mTOR signalling | 12 | 2.3 × 10−4 | 4.7 × 10−2 |
| ↓Akt2, ↓Braf, ↓Clip1, ↓Deptor, ↓Rictor, ↓Raf1, ↓Stradb, ↓Chuk, ↓Lamtor3, ↓Lpin3, ↓Mtor, ↓Pik3r1 | |||
| Salmonella infection | 15 | 3.9 × 10−4 | 4.7 × 10−2 |
| ↓Akt2, ↓Raf1, ↓S100a10, ↓Actr1b, ↓Casp7, ↓Chuk, ↓Dctn6, ↓Exoc4, ↓Exoc5, ↓Ikbkg, ↓Irak1, ↓Klc1, ↓Pik3c3, ↓Tuba1a, ↓Tuba3a | |||
| Alzheimer disease | 19 | 5.3 × 10−4 | 4.7 × 10−2 |
| ↓Akt2, ↓Braf, ↓Ndufa1, ↓Ndufv3, ↓Raf1, ↓Casp7, ↓Chuk, ↓Cdk5, ↓Cox5a, ↓Ikbkg, ↓Klc1, ↓Mtor, ↓Mme, ↓Pik3c3, ↓Pik3r1, ↓Plcb1, ↓Slc39a4, ↓Tuba1a, ↓Tuba3a | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šilhavý, J.; Mlejnek, P.; Šimáková, M.; Malínská, H.; Marková, I.; Hüttl, M.; Miklánková, D.; Kazdová, L.; Vrbacký, M.; Pecinová, A.; et al. Hypolipidemic Effects of Beetroot Juice in SHR-CRP and HHTg Rat Models of Metabolic Syndrome: Analysis of Hepatic Proteome. Metabolites 2023, 13, 192. https://doi.org/10.3390/metabo13020192
Šilhavý J, Mlejnek P, Šimáková M, Malínská H, Marková I, Hüttl M, Miklánková D, Kazdová L, Vrbacký M, Pecinová A, et al. Hypolipidemic Effects of Beetroot Juice in SHR-CRP and HHTg Rat Models of Metabolic Syndrome: Analysis of Hepatic Proteome. Metabolites. 2023; 13(2):192. https://doi.org/10.3390/metabo13020192
Chicago/Turabian StyleŠilhavý, Jan, Petr Mlejnek, Miroslava Šimáková, Hana Malínská, Irena Marková, Martina Hüttl, Denisa Miklánková, Ludmila Kazdová, Marek Vrbacký, Alena Pecinová, and et al. 2023. "Hypolipidemic Effects of Beetroot Juice in SHR-CRP and HHTg Rat Models of Metabolic Syndrome: Analysis of Hepatic Proteome" Metabolites 13, no. 2: 192. https://doi.org/10.3390/metabo13020192