Raman Microspectroscopic Analysis of Selenium Bioaccumulation by Green Alga Chlorella vulgaris

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Se-Enriched C. vulgaris
2.2. Analysis of Se Accumulation Mechanism in Living and Dead Cells
2.3. Sample Collection and Preparation for ICP-MS and RS
2.4. Inductively Coupled Plasma Mass Spectrometry (ICP-MS) Analysis
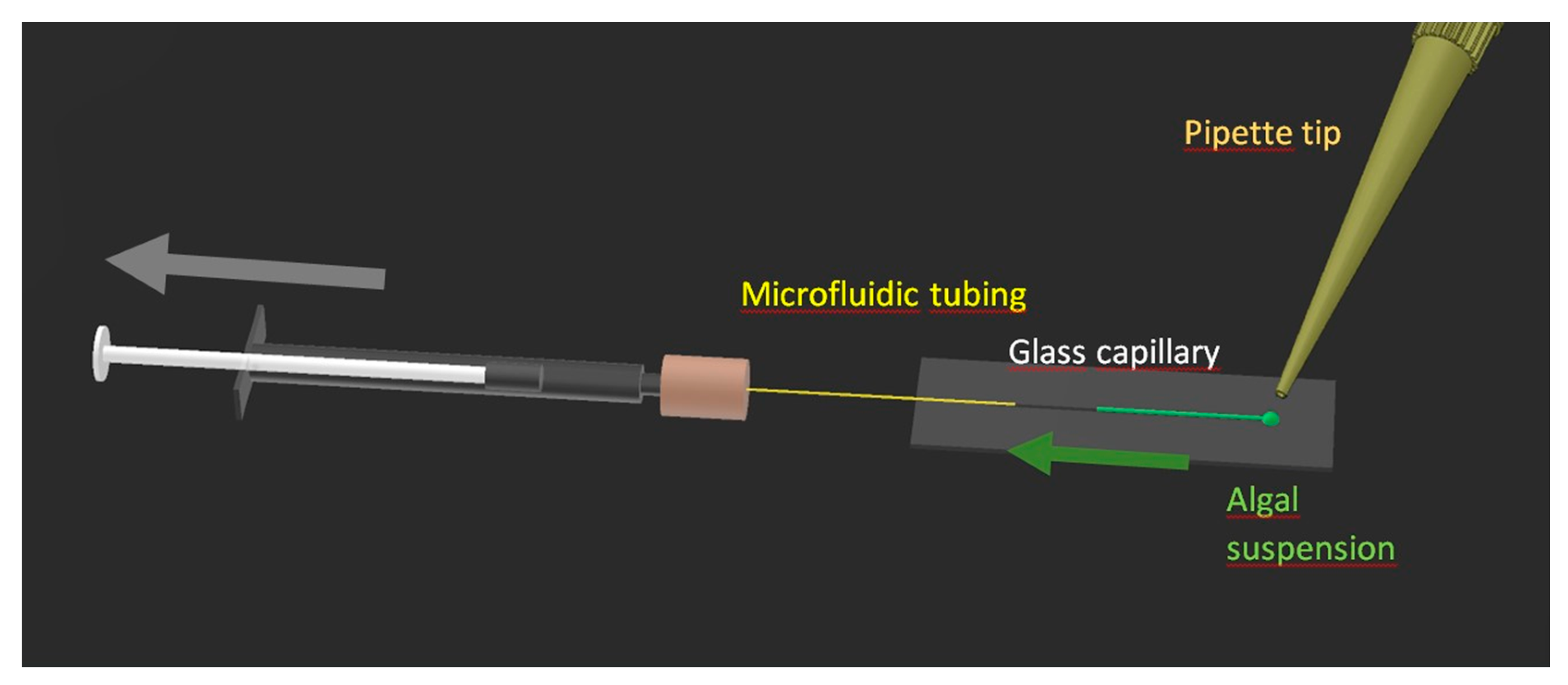
2.5. Sample Handling Device
2.6. Raman Microspectroscopy
2.7. Scanning Electron Microscopy (SEM) and EDX Analyses
3. Results and Discussion
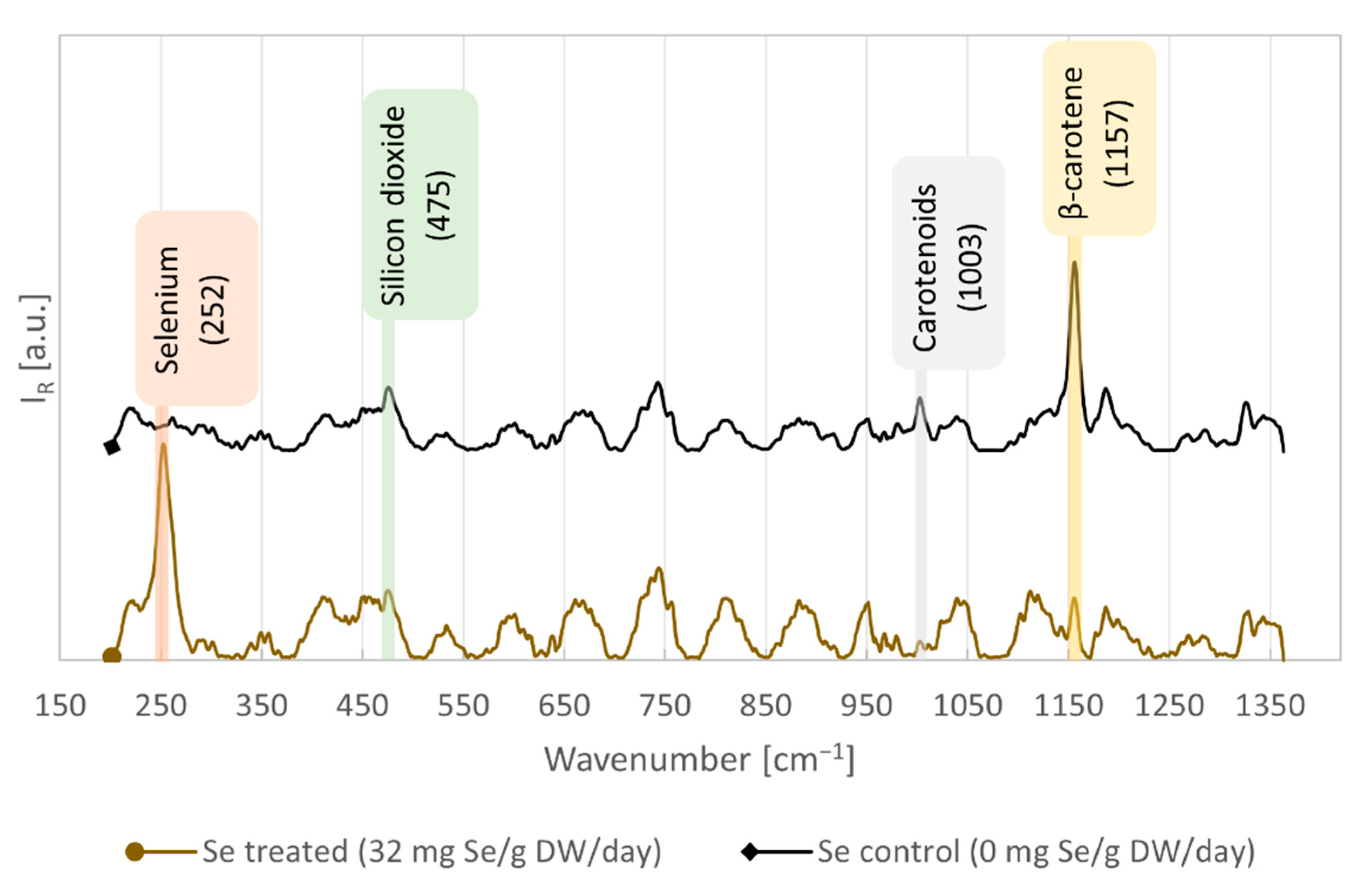
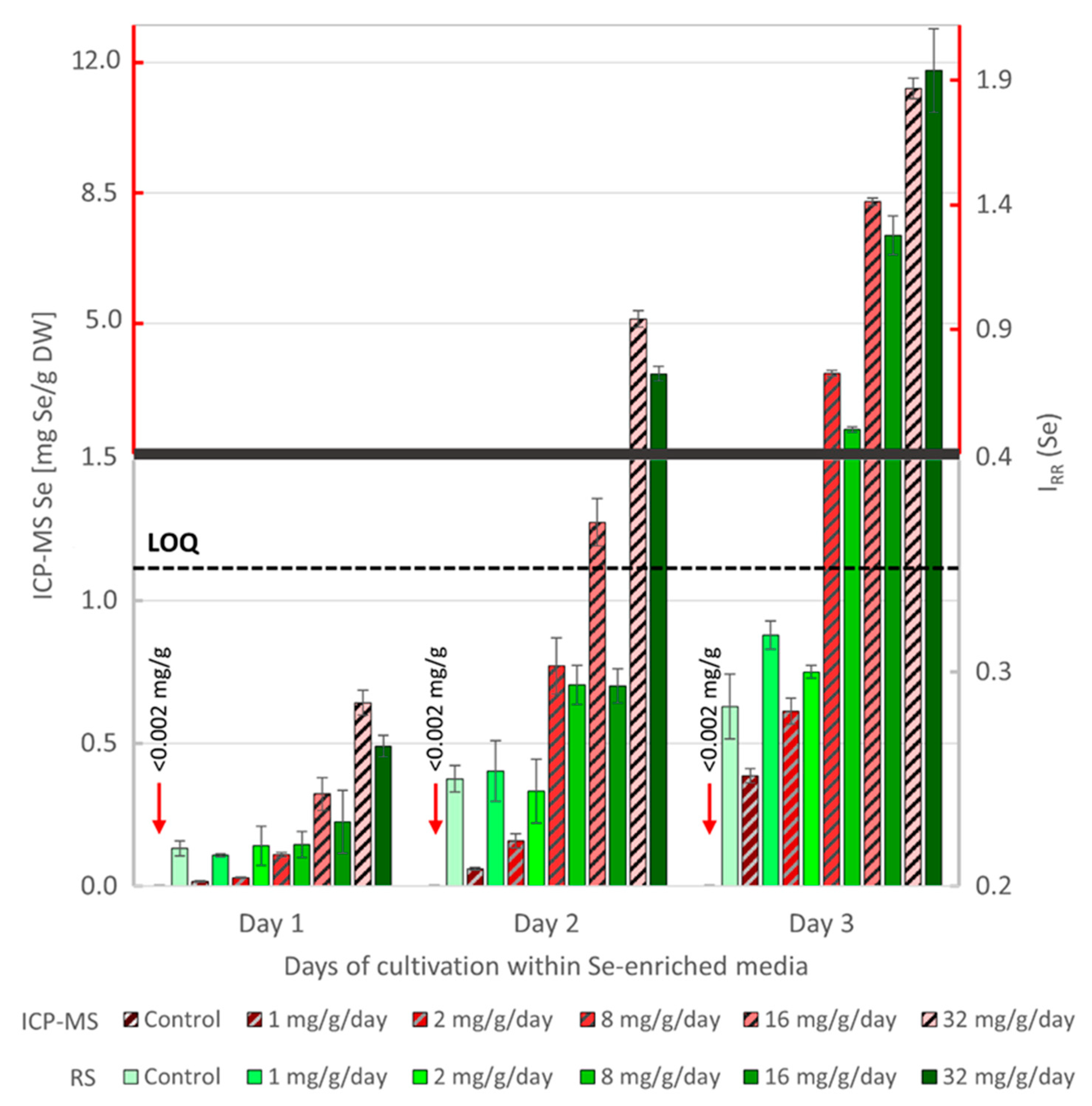
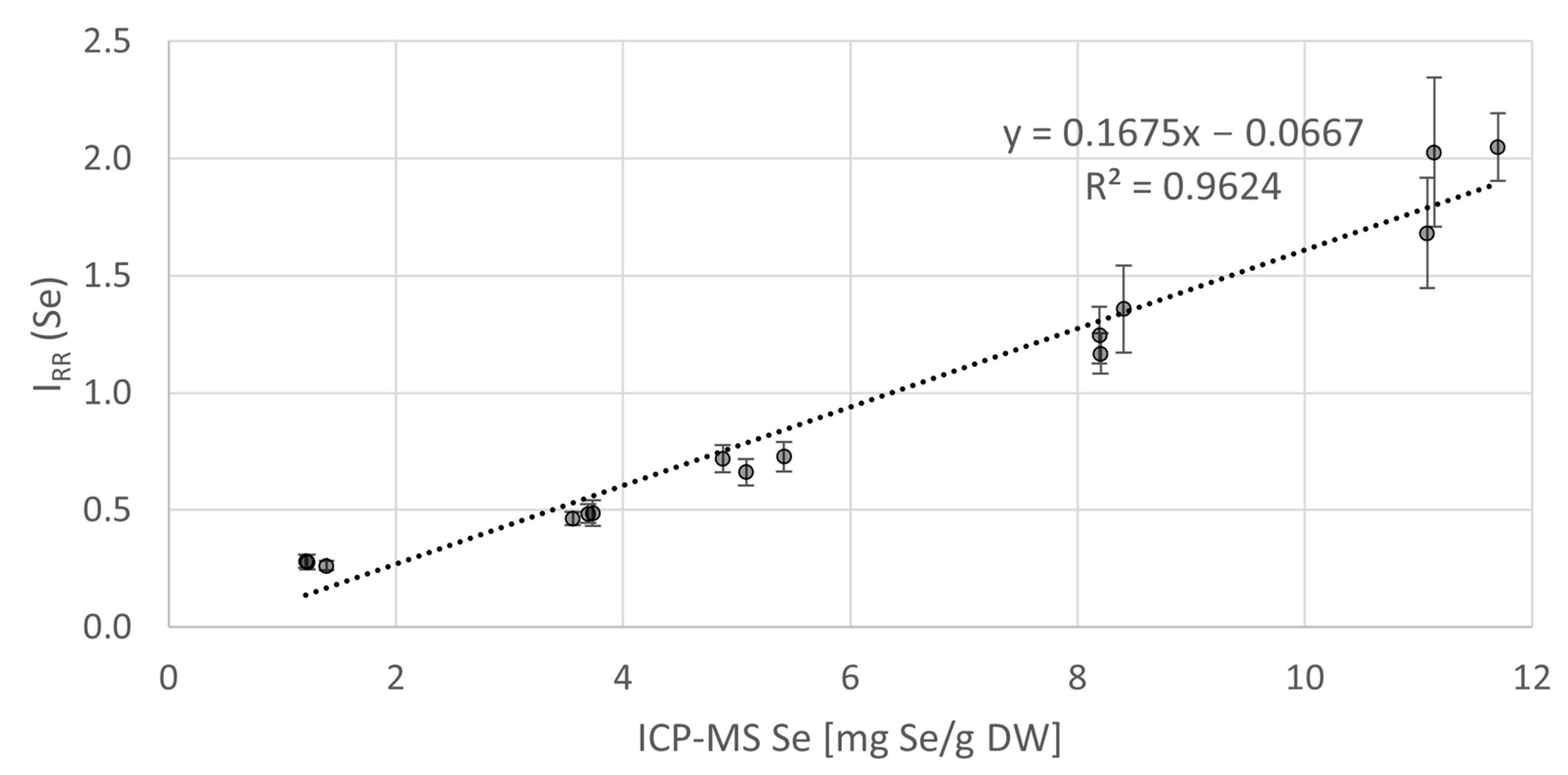
3.1. RS Quantification of Se in Algal Cells and Validation by ICP-MS
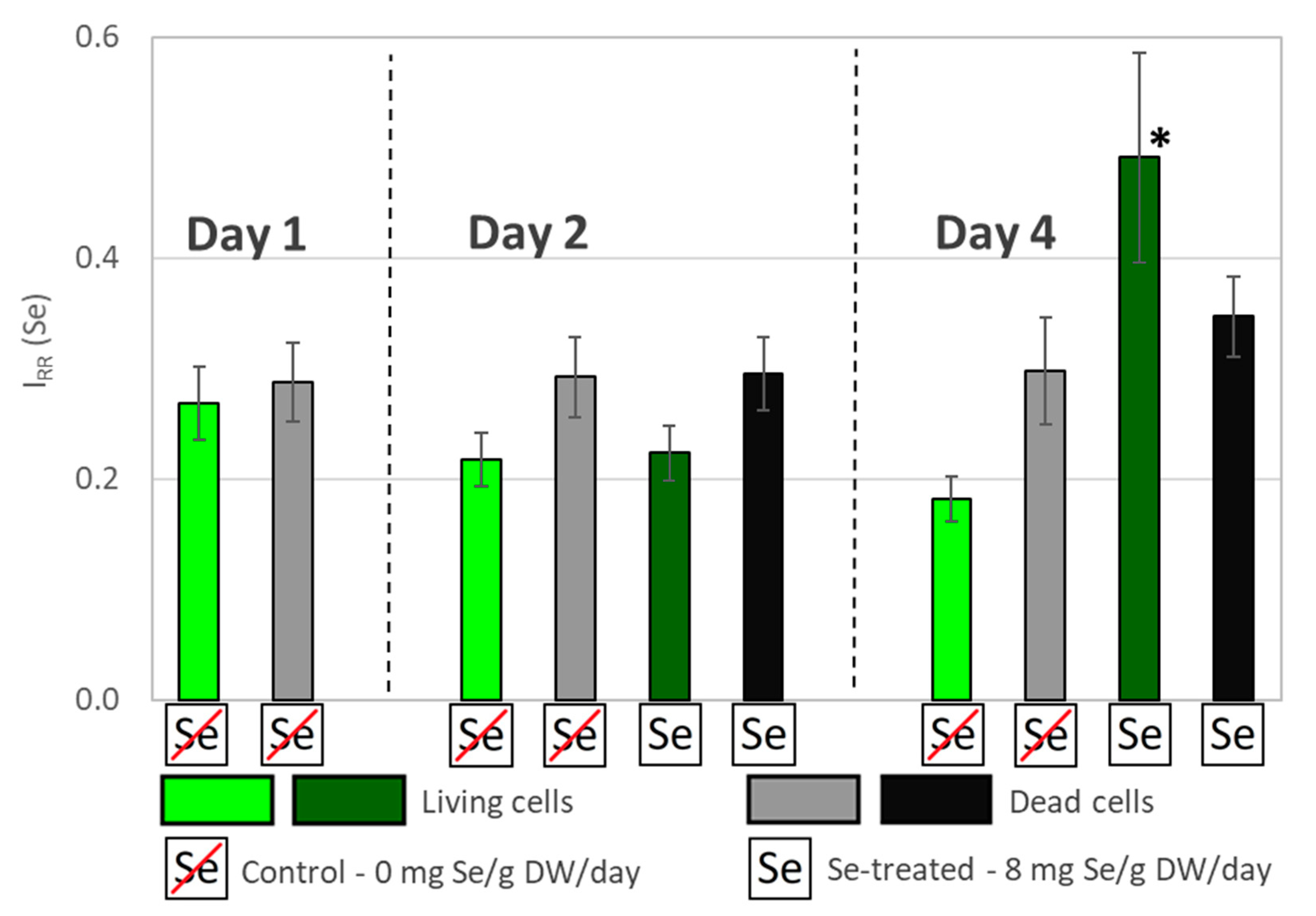
3.2. Comparative RS Analysis of Se Uptake Dynamics by Living and Dead Algal Cells
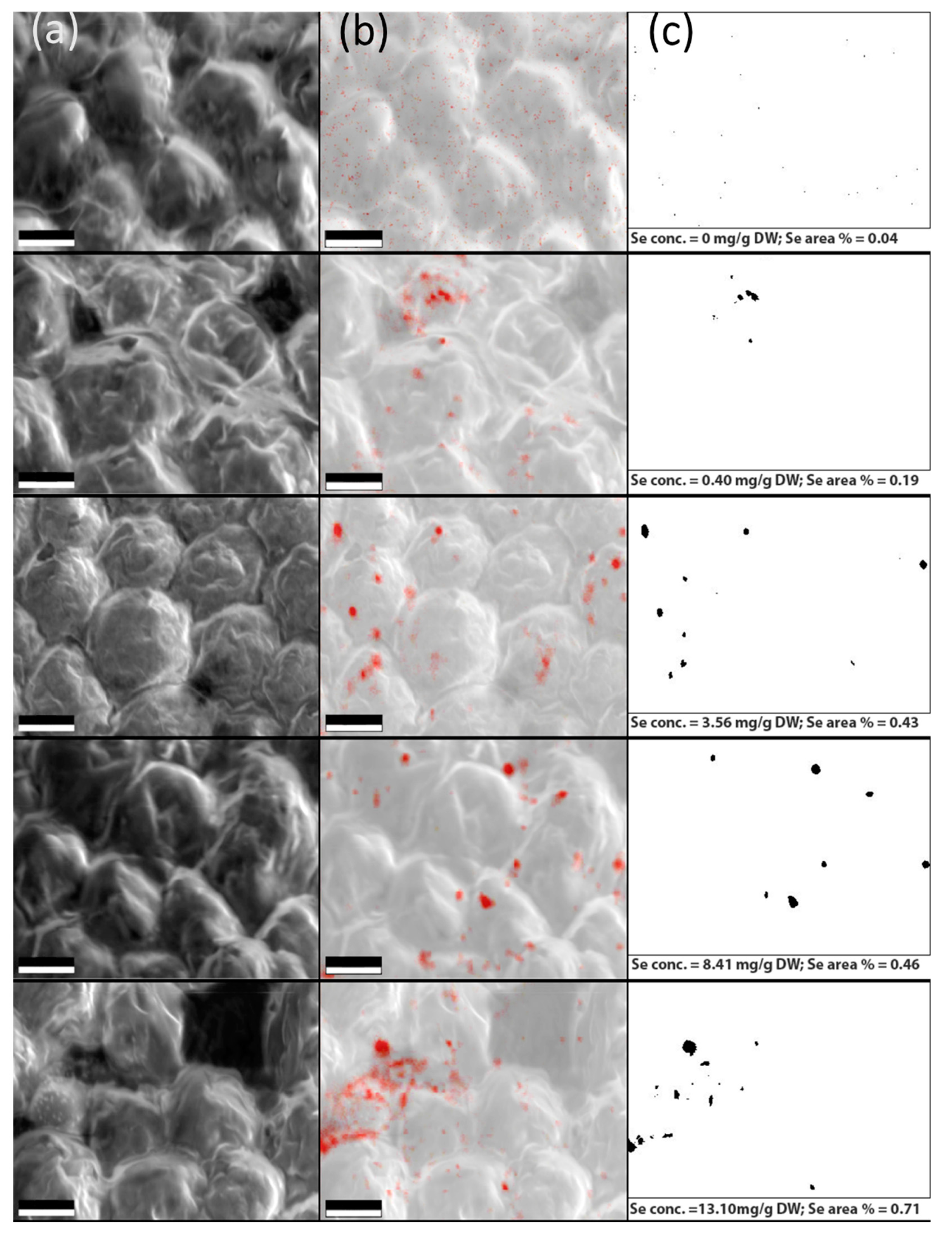
3.3. Verification of the Elemental Se Deposition in Algal Cells by SEM and EDX Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dobbs, M.G.; Cherry, D.S.; Cairns, J. Toxicity and bioaccumulation of selenium to a three-trophic level food chain. Environ. Toxicol. Chem. 1996, 15, 340–347. [Google Scholar] [CrossRef]
- Schiavon, M.; Ertani, A.; Parrasia, S.; Vecchia, F.D. Selenium accumulation and metabolism in algae. Aquat. Toxicol. 2017, 189, 1–8. [Google Scholar] [CrossRef]
- Tinggi, U. Selenium: Its role as antioxidant in human health. Environ. Health Prev. Med. 2008, 13, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gojkovic, Ž.; Garbayo, I.; Ariza, J.L.G.; Marova, I.; Vílchez, C. Selenium bioaccumulation and toxicity in cultures of green microalgae. Algal Res. 2015, 7, 106–116. [Google Scholar] [CrossRef]
- Kretzschmar, J.; Jordan, N.; Brendler, E.; Tsushima, S.; Franzen, C.; Foerstendorf, H.; Stockmann, M.; Heim, K. Spectroscopic evidence for selenium(iv) dimerization in aqueous solution. Dalton Trans. 2015, 44, 10508–10515. [Google Scholar] [CrossRef] [Green Version]
- Mylenko, M.; Vu, D.L.; Kuta, J.; Ranglová, K.; Kubáč, D.; Lakatos, G.; Grivalský, T.; Caporgno, M.P.; Manoel, J.A.D.C.; Kopecký, J.; et al. Selenium Incorporation to Amino Acids in Chlorella Cultures Grown in Phototrophic and Heterotrophic Regimes. J. Agric. Food Chem. 2020, 68, 1654–1665. [Google Scholar] [CrossRef]
- Schiavon, M.; Vecchia, F.D. Selenium and Algae: Accumulation, Tolerance Mechanisms and Dietary Perspectives. In Selenium in Plants; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Chen, T.-F.; Zheng, W.-J.; Luo, Y.; Yang, F.; Bai, Y.; Tu, F. Effects of selenium stress on photosynthetic pigment contents and growth of Chlorella vulgaris. J. Plant Physiol. Mol. Boil. 2005, 31, 369–373. [Google Scholar]
- Vu, D.L.; Saurav, K.; Mylenko, M.; Ranglová, K.; Kuta, J.; Ewe, D.; Masojídek, J.; Hrouzek, P. In vitro bioaccessibility of selenoamino acids from selenium (Se)-enriched Chlorella vulgaris biomass in comparison to selenized yeast; a Se-enriched food supplement; and Se-rich foods. Food Chem. 2019, 279, 12–19. [Google Scholar] [CrossRef]
- Doucha, J.; Lívanský, K.; Kotrbáček, V.; Zachleder, V. Production of Chlorella biomass enriched by selenium and its use in animal nutrition: A review. Appl. Microbiol. Biotechnol. 2009, 83, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Umysová, D.; Vítová, M.; Doušková, I.; Bišová, K.; Hlavová, M.; Čížková, M.; Machát, J.; Doucha, J.; Zachleder, V. Bioaccumulation and toxicity of selenium compounds in the green alga Scenedesmus quadricauda. BMC Plant Biol. 2009, 9, 58. [Google Scholar] [CrossRef] [Green Version]
- Vriens, B.; Behra, R.; Voegelin, A.; Zupanic, A.; Winkel, L.H.E. Selenium Uptake and Methylation by the Microalga Chlamydomonas reinhardtii. Environ. Sci. Technol. 2016, 50, 711–720. [Google Scholar] [CrossRef]
- Zhong, Y.; Cheng, J.J. Effects of Selenite on Unicellular Green MicroalgaChlorella pyrenoidosa: Bioaccumulation of Selenium, Enhancement of Photosynthetic Pigments, and Amino Acid Production. J. Agric. Food Chem. 2017, 65, 10875–10883. [Google Scholar] [CrossRef]
- Simmons, D.B.D.; Emery, R.J.N. Phytochelatin induction by selenate in Chlorella vulgaris, and regulation of effect by sulfate levels. Environ. Toxicol. Chem. 2011, 30, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Gupta, S. An Overview of Selenium Uptake, Metabolism, and Toxicity in Plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P.M.; De Souza, M.P.; Pickering, I.J.; Terry, N. Rapid microalgal metabolism of selenate to volatile dimethylselenide. Plant Cell Environ. 2003, 26, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Oremland, R.S.; Herbel, M.J.; Blum, J.S.; Langley, S.; Beveridge, T.J.; Ajayan, P.M.; Sutto, T.; Ellis, A.V.; Curran, S. Structural and Spectral Features of Selenium Nanospheres Produced by Se-Respiring Bacteria. Appl. Environ. Microbiol. 2004, 70, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Bodnar, M.; Szczyglowska, M.; Konieczka, P.; Namiesnik, J. Methods of Selenium Supplementation: Bioavailability and Determination of Selenium Compounds. Crit. Rev. Food Sci. Nutr. 2016, 56, 36–55. [Google Scholar] [CrossRef]
- Gerrard, T.L.; Telford, J.N.; Williams, H.H. Detection of Selenium Deposits in Escherichia coli by Electron Microscopy. J. Bacteriol. 1974, 119, 1057–1060. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Lamana, J.; Abad-Álvaro, I.; Bierla, K.; Laborda, F.; Szpunar, J.; Lobinski, R. Detection and characterization of biogenic selenium nanoparticles in selenium-rich yeast by single particle ICPMS. J. Anal. At. Spectrom. 2018, 33, 452–460. [Google Scholar] [CrossRef]
- Montes-Bayón, M.; LeDuc, D.L.; Terry, N.; Caruso, J.A. Selenium speciation in wild-type and genetically modified Se accumulating plants with HPLC separation and ICP-MS/ES-MS detection. J. Anal. At. Spectrom. 2002, 17, 872–879. [Google Scholar] [CrossRef]
- McSheehy, S.; Yang, W.; Pannier, F.; Szpunar, J.; Łobiński, R.; Auger, J.; Potin-Gautier, M. Speciation analysis of selenium in garlic by two-dimensional high-performance liquid chromatography with parallel inductively coupled plasma mass spectrometric and electrospray tandem mass spectrometric detection. Anal. Chim. Acta 2000, 421, 147–153. [Google Scholar] [CrossRef]
- Sun, X.; Zhong, Y.; Huang, Z.; Yang, Y. Selenium Accumulation in Unicellular Green Alga Chlorella vulgaris and Its Effects on Antioxidant Enzymes and Content of Photosynthetic Pigments. PLoS ONE 2014, 9, e112270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponton, D.E.; Fortin, C.; Hare, L. Organic selenium, selenate, and selenite accumulation by lake plankton and the algaChlamydomonas reinhardtiiat different pH and sulfate concentrations. Environ. Toxicol. Chem. 2018, 37, 2112–2122. [Google Scholar] [CrossRef]
- Vu, D.L.; Ranglová, K.; Hájek, J.; Hrouzek, P. Quantification of methionine and selenomethionine in biological samples using multiple reaction monitoring high performance liquid chromatography tandem mass spectrometry (MRM-HPLC-MS/MS). J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1084, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu-Aruxandei, D.; Frîncu, R.M.; Capră, L.; Oancea, F. Selenium Analysis and Speciation in Dietary Supplements Based on Next-Generation Selenium Ingredients. Nutrients 2018, 10, 1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilát, Z.; Bernatová, S.; Ježek, J.; Šery, M.; Samek, O.; Zemánek, P.; Nedbal, L.; Trtílek, M. Raman microspectroscopy of algal lipid bodies: β-carotene as a volume sensor. In Proceedings of the Photonics, Devices, and Systems V, Prague, Czech Republic, 24–26 August 2011; Volume 8306, p. 83060L. [Google Scholar] [CrossRef]
- Samek, O.; Jonáš, A.; Pilát, Z.; Zemánek, P.; Nedbal, L.; Tříska, J.; Kotas, P.; Trtílek, M. Raman spectroscopy for the characterization of algal cells. In Proceedings of the 17th Slovak-Czech-Polish Optical Conference on Wave and Quantum Aspects of Contemporary Optics, Liptovsky Jan, Slovakia, 6–10 September 2010; Volume 7746, p. 77460X. [Google Scholar] [CrossRef]
- Parab, N.D.T.; Tomar, V. Raman spectroscopy of algae: A review. J. Nanomed. Nanotechnol. 2012, 3. [Google Scholar] [CrossRef]
- Samek, O.; Obruča, S.; Šiler, M.; Sedláček, P.; Benešová, P.; Kučera, D.; Márova, I.; Ježek, J.; Bernatová, S.; Zemánek, P. Quantitative Raman Spectroscopy Analysis of Polyhydroxyalkanoates Produced by Cupriavidus necator H16. Sensors 2016, 16, 1808. [Google Scholar] [CrossRef] [Green Version]
- Serra-Maia, R.; Bernard, O.; Gonçalves, A.; Bensalem, S.; Lopes, F. Influence of temperature on Chlorella vulgaris growth and mortality rates in a photobioreactor. Algal Res. 2016, 18, 352–359. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with imageJ. Biophotonics Int. 2004, 11, 36–41. [Google Scholar] [CrossRef]
- Jadhav, A.A.; Khanna, P.K. Impact of microwave irradiation on cyclo-octeno-1,2,3-selenadiazole: Formation of selenium nanoparticles and their polymorphs. RSC Adv. 2015, 5, 44756–44763. [Google Scholar] [CrossRef]
- Goldan, A.H.; Li, C.; Pennycook, S.J.; Schneider, J.; Blom, A.; Zhao, W. Molecular structure of vapor-deposited amorphous selenium. J. Appl. Phys. 2016, 120, 14072–14081. [Google Scholar] [CrossRef]
- Takaichi, S. Distributions, biosyntheses and functions of carotenoids in algae. Agro Food Ind. Hi Tech 2013, 24, 55–58. [Google Scholar]
- Jablonski, P.P.; Anderson, J.W. Light-dependent reduction of selenite by sonicated pea chloroplasts. Phytochemistry 1982, 21, 2179–2184. [Google Scholar] [CrossRef]
- Casano, L.M.; Gómez, L.D.; Trippi, V.S. Oxygen- and Light-Induced Proteolysis in Isolated Oat Chloroplasts. Plant Cell Physiol. 1990, 31, 377–382. [Google Scholar] [CrossRef]
- Savchenko, D.; Vorlíček, V.; Prokhorov, A.; Kalabukhova, E.; Lančok, J.; Jelínek, M. Raman and EPR spectroscopic studies of chromium-doped diamond-like carbon films. Diam. Relat. Mater. 2018, 83, 30–37. [Google Scholar] [CrossRef]
- Fereche, E.; Tombo, I.; Ntwampe SK, O.; Baptist, J.; Mudumbi, N. Leaching of Cyanogens and Mycotoxins from Cultivated Cassava into Agricultural Soil: Effects on Groundwater Quality World’s Largest Science; Instrument Society of America: Albuquerque, NM, USA, 2017. [Google Scholar] [CrossRef] [Green Version]
- Brazhe, N.A.; Erokhova, L.A.; Churin, A.A.; Maksimov, G.V. The Relation of Different-Scale Membrane Processes Under Nitric Oxide Influence. J. Biol. Phys. 2005, 31, 533–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Li, Z.; Lu, L.; Xiao, Y.; Liu, J.; Guo, J.; Fang, F. Effects of stepwise nitrogen depletion on carotenoid content, fluorescence parameters and the cellular stoichiometry of Chlorella vulgaris. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 181, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Wenzl, T.; Haedrich, J.; Schaechtele, A.; Robouch, P.; Stroka, J. Guidance Document on the Estimation of LOD and LOQ for Measurements in the Field of Contaminants in Feed and Food; European Union Reference Laboratory, Publications Office of the European Union: Luxembourg, 2016; Volume EUR 28099, ISBN 978-92-79-61768-3. [Google Scholar] [CrossRef]
- Shoeibi, S.; Mozdziak, P.; Golkar-Narenji, A. Biogenesis of Selenium Nanoparticles Using Green Chemistry. Top. Curr. Chem. 2017, 375, 88. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Wavenumber [cm−1] | Assignment | Reference |
|---|---|---|
| 252 | Selenium Se-Se stretching in monoclinic Se8 rings | [33] |
| 475 | silicon dioxide Si-O-Si rocking and bending | [38] |
| 1003 | Carotenoids CH3 in-plane rocking | [39,40] |
| 1157 | β-carotene C-C stretching, C-H in-plane bending | [39,40,41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kizovský, M.; Pilát, Z.; Mylenko, M.; Hrouzek, P.; Kuta, J.; Skoupý, R.; Krzyžánek, V.; Hrubanová, K.; Adamczyk, O.; Ježek, J.; et al. Raman Microspectroscopic Analysis of Selenium Bioaccumulation by Green Alga Chlorella vulgaris. Biosensors 2021, 11, 115. https://doi.org/10.3390/bios11040115
Kizovský M, Pilát Z, Mylenko M, Hrouzek P, Kuta J, Skoupý R, Krzyžánek V, Hrubanová K, Adamczyk O, Ježek J, et al. Raman Microspectroscopic Analysis of Selenium Bioaccumulation by Green Alga Chlorella vulgaris. Biosensors. 2021; 11(4):115. https://doi.org/10.3390/bios11040115
Chicago/Turabian StyleKizovský, Martin, Zdeněk Pilát, Mykola Mylenko, Pavel Hrouzek, Jan Kuta, Radim Skoupý, Vladislav Krzyžánek, Kamila Hrubanová, Olga Adamczyk, Jan Ježek, and et al. 2021. "Raman Microspectroscopic Analysis of Selenium Bioaccumulation by Green Alga Chlorella vulgaris" Biosensors 11, no. 4: 115. https://doi.org/10.3390/bios11040115