
Fungal Endophytes of Vitis vinifera—Plant Growth Promotion Factors

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Fungal Endophytes Isolation and Cultivation
2.3. Molecular Genetic Identification of Endophytes
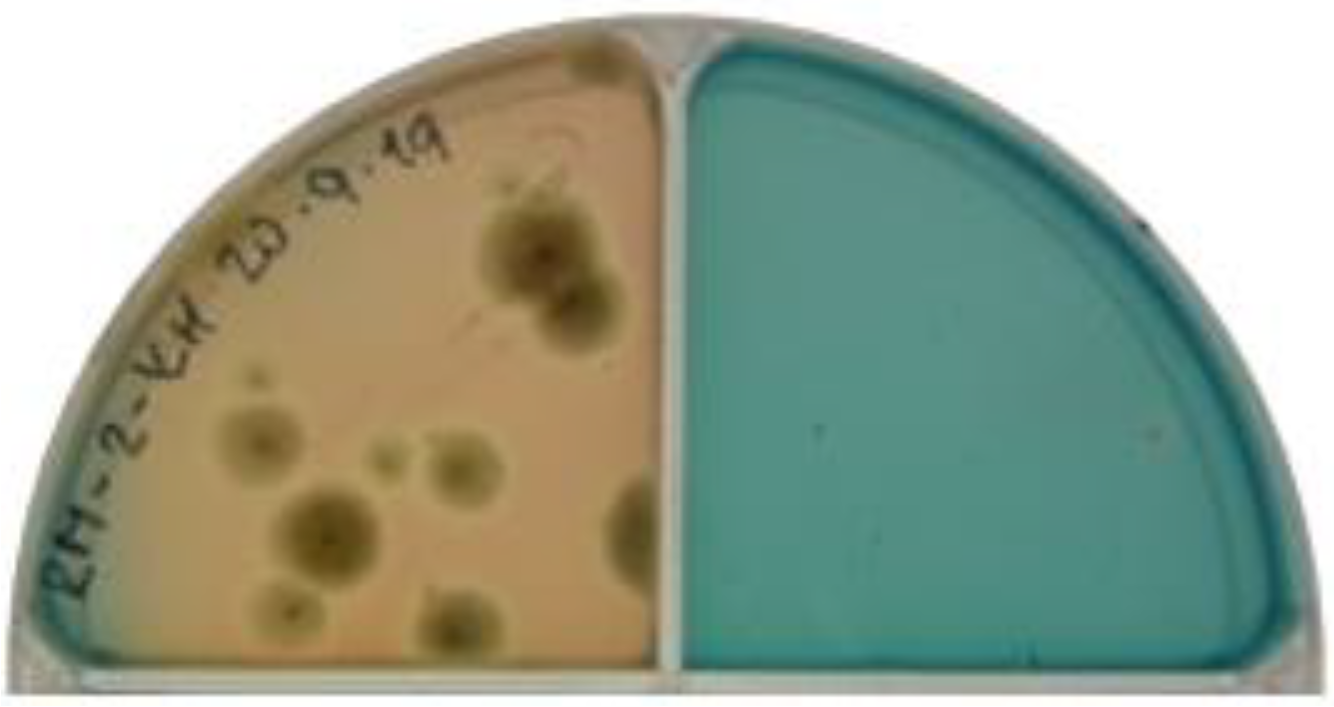
2.4. Determination of Siderophores Production of the Isolates
2.5. Determination of Antioxidant Activity of the Isolates
2.6. Determination of Antifungal Activity of the Isolates
2.7. Statistical Analysis
3. Results
3.1. Fungal Endophytes Characterization and Molecular Genetic Identification
3.2. Production of Siderophores
3.3. Antioxidant Activity
3.4. Antifungal Activity
4. Discussion
4.1. Fungal Endophytes Characterization and Molecular Genetic Identification
4.2. Production of Siderophores
4.3. Antioxidant Activity
4.4. Antifungal Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kolouchová, I.; Melzoch, K.; Šmidrkal, J.; Filip, V. The content of resveratrol in vegetables and fruit. Chem. Listy 2005, 99, 492–495. [Google Scholar]
- Schulz, B. Mutualistic interactions with fungal root endophytes. In Microbial Root Endophytes; Springer: Berlin/Heidelberg, Germany, 2006; pp. 261–279. [Google Scholar]
- Maheshwari, D.K.; Maheshwari, R.; Rinaudo. Endophytes: Biology and Biotechnology; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.N.; Singh, S.; Mishra, S.; Gupta, A. Recent Advancement in White Biotechnology through Fungi; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- González, V.; Tello, M.L. The endophytic mycota associated with V itis vinifera in central Spain. Fungal Divers. 2011, 47, 29–42. [Google Scholar] [CrossRef]
- Toghueo, R.M.K.; Boyom, F.F. Endophytic Penicillium species and their agricultural, biotechnological, and pharmaceutical applications. 3 Biotech 2020, 10, 107. [Google Scholar] [CrossRef]
- Gupta, S.; Chaturvedi, P.; Kulkarni, M.G.; Van Staden, J. A critical review on exploiting the pharmaceutical potential of plant endophytic fungi. Biotechnol. Adv. 2020, 39, 107462. [Google Scholar] [CrossRef]
- Dina, B.; Mamtaj, S. Endophytic microorganisms: Colonization, plant-microbe interaction, diversity and their Bioprospecting. Res. J. Biotechnol. 2020, 15, 151–179. [Google Scholar]
- Zimowska, B.; Bielecka, M.; Abramczyk, B.; Nicoletti, R. Bioactive products from endophytic fungi of sages (Salvia spp.). Agriculture 2020, 10, 543. [Google Scholar] [CrossRef]
- Yadav, G.; Meena, M. Bioprospecting of endophytes in medicinal plants of Thar Desert: An attractive resource for biopharmaceuticals. Biotechnol. Rep. 2021, 30, e00629. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.; Gupta, S.; Dhar, M.K.; Kaul, S. Endophytic Fungi-Mediated Biocatalysis and Biotransformations Paving the Way Toward Green Chemistry. Front. Bioeng. Biotechnol. 2021, 9, 419. [Google Scholar] [CrossRef]
- Ntemafack, A.; Kapoor, N.; Ali, S.; Jamwal, V.L.; Hassan, Q.P.; Gandhi, S.G. Comprehensive Review of Endophytic Flora from African Medicinal Plants. Curr. Microbiol. 2021, 78, 2860–2898. [Google Scholar] [CrossRef]
- Ancheeva, E.; Daletos, G.; Proksch, P. Bioactive secondary metabolites from endophytic fungi. Curr. Med. Chem. 2020, 27, 1836–1854. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef]
- Górska, A.; Sloderbach, A.; Marszałł, M.P. Siderophore–drug complexes: Potential medicinal applications of the ‘Trojan horse’strategy. Trends Pharmacol. Sci. 2014, 35, 442–449. [Google Scholar] [CrossRef] [PubMed]
- El-Tarabily, K.A.; Sivasithamparam, K. Potential of yeasts as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Mycoscience 2006, 47, 25–35. [Google Scholar] [CrossRef]
- Schena, L.; Ippolito, A.; Zahavi, T.; Cohen, L.; Nigro, F.; Droby, S. Genetic diversity and biocontrol activity of Aureobasidium pullulans isolates against postharvest rots. Postharvest Biol. Technol. 1999, 17, 189–199. [Google Scholar] [CrossRef]
- Schena, L.; Nigro, F.; Pentimone, I.; Ligorio, A.; Ippolito, A. Control of postharvest rots of sweet cherries and table grapes with endophytic isolates of Aureobasidium pullulans. Postharvest Biol. Technol. 2003, 30, 209–220. [Google Scholar] [CrossRef]
- Sullivan, R.F.; White, J.F., Jr. Phoma glomerata as a mycoparasite of powdery mildew. Appl. Environ. Microbiol. 2000, 66, 425–427. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Hu, Z.; Hou, D.; Xu, H.; Song, P. Biodiversity and antifungal potential of endophytic fungi from the medicinal plant Cornus officinalis. Symbiosis 2020, 81, 223–233. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Suprun, A.R.; Nityagovsky, N.N.; Dubrovina, A.S.; Kiselev, K.V. The Influence of the Grapevine Bacterial and Fungal Endophytes on Biomass Accumulation and Stilbene Production by the In Vitro Cultivated Cells of Vitis amurensis Rupr. Plants 2021, 10, 1276. [Google Scholar] [CrossRef] [PubMed]
- Mondello, V.; Spagnolo, A.; Larignon, P.; Clement, C.; Fontaine, F. Phytoprotection potential of Fusarium proliferatum for control of Botryosphaeria dieback pathogens in grapevine. Phytopathol. Mediterr. 2019, 58, 293–306. [Google Scholar]
- Shi, J.; Zeng, Q.; Liu, Y.; Pan, Z. Alternaria sp. MG1, a resveratrol-producing fungus: Isolation, identification, and optimal cultivation conditions for resveratrol production. Appl. Microbiol. Biotechnol. 2012, 95, 369–379. [Google Scholar] [CrossRef]
- Yang, M.-Z.; Ma, M.-D.; Yuan, M.-Q.; Huang, Z.-Y.; Yang, W.-X.; Zhang, H.-B.; Huang, L.-H.; Ren, A.-Y.; Shan, H. Fungal Endophytes as a Metabolic Fine-Tuning Regulator for Wine Grape. PLoS ONE 2016, 11, e0163186. [Google Scholar] [CrossRef] [PubMed]
- Kolařík, M.; Spakowicz, D.J.; Gazis, R.; Shaw, J.; Kubátová, A.; Nováková, A.; Chudíčková, M.; Forcina, G.C.; Kang, K.W.; Kelnarová, I. Biatriospora (Ascomycota: Pleosporales) is an ecologically diverse genus including facultative marine fungi and endophytes with biotechnological potential. Plant Syst. Evol. 2017, 303, 35–50. [Google Scholar] [CrossRef]
- Píchová, K.; Pažoutová, S.; Kostovčík, M.; Chudíčková, M.; Stodůlková, E.; Novák, P.; Flieger, M.; van der Linde, E.; Kolařík, M. Evolutionary history of ergot with a new infrageneric classification (Hypocreales: Clavicipitaceae: Claviceps). Mol. Phylogenet. Evol. 2018, 123, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.P.; Pires, C.; Moreira, H.; Rangel, A.O.; Castro, P.M. Assessment of the plant growth promotion abilities of six bacterial isolates using Zea mays as indicator plant. Soil Biol. Biochem. 2010, 42, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Fidler, M.; Kolářová, L. Analýza antioxidantů v chmelu a pivu. Chem. Listy 2009, 103, 232–235. [Google Scholar]
- Bell, D.; Wells, H.; Markham, C. In vitro antagonism of Trichoderma species against six fungal plant pathogens. Phytopathology 1982, 72, 379–382. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Proksch, P. Fungal endophytes: Unique plant inhabitants with great promises. Appl. Microbiol. Biotechnol. 2011, 90, 1829–1845. [Google Scholar] [CrossRef]
- Núñez-Trujillo, G.; Cabrera, R.; Burgos-Reyes, R.; Silva, E.d.; Giménez, C.; Cosoveanu, A.; Brito, N. Endophytic fungi from Vitis vinifera L. isolated in Canary Islands and Azores as potential biocontrol agents of Botrytis cinerea Pers.: Fr. J. Hortic. For. Biotechnol. 2012, 16, 1–6. [Google Scholar]
- Muvea, A.M.; Subramanian, S.; Maniania, N.K.; Poehling, H.-M.; Ekesi, S.; Meyhöfer, R. Endophytic colonization of onions induces resistance against viruliferous thrips and virus replication. Front. Plant Sci. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Varanda, C.M.R.; Oliveira, M.; Materatski, P.; Landum, M.; Clara, M.I.E.; do Rosário Félix, M. Fungal endophytic communities associated to the phyllosphere of grapevine cultivars under different types of management. Fungal Biol. 2016, 120, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.S.; Leite, L.F.; Santos, V.B.; Carneiro, R.F. Soil microbial activity in conventional and organic agricultural systems. Sustainability 2009, 1, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Sahib, M.R.; Amna, A.; Opiyo, S.O.; Zhao, Z.; Gao, Y.G. Culturable endophytic fungal communities associated with plants in organic and conventional farming systems and their effects on plant growth. Sci. Rep. 2019, 9, 1–10. [Google Scholar]
- Geissen, V.; Silva, V.; Lwanga, E.H.; Beriot, N.; Oostindie, K.; Bin, Z.; Pyne, E.; Busink, S.; Zomer, P.; Mol, H. Cocktails of pesticide residues in conventional and organic farming systems in Europe–Legacy of the past and turning point for the future. Environ. Pollut. 2021, 278, 116827. [Google Scholar] [CrossRef]
- Burns, K.N.; Kluepfel, D.A.; Strauss, S.L.; Bokulich, N.A.; Cantu, D.; Steenwerth, K.L. Vineyard soil bacterial diversity and composition revealed by 16S rRNA genes: Differentiation by geographic features. Soil Biol. Biochem. 2015, 91, 232–247. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Suryanarayanan, T.; Murali, T.; Venkatesan, G. Occurrence and distribution of fungal endophytes in tropical forests across a rainfall gradient. Can. J. Bot. 2002, 80, 818–826. [Google Scholar] [CrossRef]
- Bacon, C.W.; White, J. Microbial Endophytes; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Sarsaiya, S.; Jain, A.; Jia, Q.; Fan, X.; Fuxing, S.; Chen, Z.; Zhou, Q.; Shi, J.; Chen, J. Molecular Identification of Endophytic Fungi and Their Pathogenicity Evaluation Against Dendrobium nobile and Dendrobium officinale. Int. J. Mol. Sci. 2020, 21, 316. [Google Scholar] [CrossRef] [Green Version]
- Wijekoon, C.; Quill, Z. Fungal endophyte diversity in table grapes. Can. J. Microbiol. 2021, 67, 29–36. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Purahong, W.; Zhang, W.; Wubet, T.; Li, X.; Liu, M.; Zhao, W.; Hyde, K.D.; Liu, J.; Yan, J. Biodiversity of fungi on Vitis vinifera L. revealed by traditional and high-resolution culture-independent approaches. Fungal Divers. 2018, 90, 1–84. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, H.; Akiyama, M.; Kobayashi, K.; Yamaji, K. Fe and P solubilization under limiting conditions by bacteria isolated from Carex kobomugi roots at the Hasaki coast. Curr. Microbiol. 2013, 66, 314–321. [Google Scholar] [CrossRef]
- Haas, H. Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat. Prod. Rep. 2014, 31, 1266–1276. [Google Scholar] [CrossRef] [Green Version]
- Sharma, H.; Rai, A.K.; Chettri, R.; Nigam, P.S. Bioactivites of Penicillium citrinum isolated from a medicinal plant Swertia chirayita. Arch. Microbiol. 2021, 203, 5173–5182. [Google Scholar] [CrossRef]
- Chowdappa, S.; Jagannath, S.; Konappa, N.; Udayashankar, A.C.; Jogaiah, S. Detection and Characterization of Antibacterial Siderophores Secreted by Endophytic Fungi from Cymbidium aloifolium. Biomolecules 2020, 10, 1412. [Google Scholar] [CrossRef]
- Huang, W.-Y.; Cai, Y.-Z.; Xing, J.; Corke, H.; Sun, M. A potential antioxidant resource: Endophytic fungi from medicinal plants. Econ. Bot. 2007, 61, 14–30. [Google Scholar] [CrossRef]
- Nischitha, R.; Shivanna, M. Metabolite fingerprinting, in vitro antimicrobial and antioxidant activities and in-silico docking in Alloteropsis cimicina and its endophytic fungus Penicillium pinophilum. Mol. Biol. Rep. 2021, 48, 4021–4037. [Google Scholar] [CrossRef]
- Arora, D.S.; Chandra, P. Antioxidant activity of Aspergillus fumigatus. Int. Sch. Res. Not. 2011, 2011, 619395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharaf, M.H.; Abdelaziz, A.M.; Kalaba, M.H.; Radwan, A.A.; Hashem, A.H. Antimicrobial, Antioxidant, Cytotoxic Activities and Phytochemical Analysis of Fungal Endophytes Isolated from Ocimum Basilicum. Appl. Biochem. Biotechnol. 2021, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Vellingiri, M.M.; Ashwin, J.K.M.; Soundari, A.J.P.G.; Sathiskumar, S.; Priyadharshini, U.; Paramasivam, D.; Liu, W.-C.; Balasubramanian, B. Mycofabrication of AgONPs derived from Aspergillus terreus FC36AY1 and its potent antimicrobial, antioxidant, and anti-angiogenesis activities. Mol. Biol. Rep. 2021, 48, 7933–7946. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Singh, A.; Singh, D.K.; Sharma, V.K.; Kumar, J.; Gupta, V.K.; Bhattacharya, S.; Kharwar, R. Isolation and purification of bioactive metabolites from an endophytic fungus Penicillium citrinum of Azadirachta indica. S. Afr. J. Bot. 2021, 139, 449–457. [Google Scholar] [CrossRef]
- Devi, N.; Prabakaran, J. Bioactive metabolites from an endophytic fungus Penicillium sp. isolated from Centella asiatica. Curr. Res. Environ. Appl. Mycol. 2014, 4, 34–43. [Google Scholar] [CrossRef]
- Kaur, R.; Kaur, J.; Kaur, M.; Kalotra, V.; Chadha, P.; Kaur, A.; Kaur, A. An endophytic Penicillium oxalicum isolated from Citrus limon possesses antioxidant and genoprotective potential. J. Appl. Microbiol. 2020, 128, 1400–1413. [Google Scholar] [CrossRef]
- Nasrin, M.; Afroz, F.; Begum, M.N.; Rony, S.R.; Sharmin, S.; Moni, F.; Rana, M.S.; Sohrab, M.H. Isolation and Bioactivity Screening of Endophytic Fungi from Commelina diffusa. Indian J. Pharm. Educ. Res. 2021, 55, 829–836. [Google Scholar] [CrossRef]
- Kumar, V.; Prasher, I. Phytochemical analysis and antimicrobial potential of Nigrospora sphaerica (Berk. & Broome) Petch, a fungal endophyte isolated from Dillenia indica L. Adv. Tradit. Med. 2021, 1–13. [Google Scholar] [CrossRef]
- Acuña-Rodríguez, I.S.; Newsham, K.K.; Gundel, P.E.; Torres-Díaz, C.; Molina-Montenegro, M.A. Functional roles of microbial symbionts in plant cold tolerance. Ecol. Lett. 2020, 23, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Monggoot, S.; Popluechai, S.; Gentekaki, E.; Pripdeevech, P. Fungal endophytes: An alternative source for production of volatile compounds from agarwood oil of Aquilaria subintegra. Microb. Ecol. 2017, 74, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.E.; Bauerle, T.L. A new currency for mutualism? Fungal endophytes alter antioxidant activity in hosts responding to drought. Fungal Divers. 2012, 54, 39–49. [Google Scholar] [CrossRef]
- Gams, W.; Anderson, T.-H. Compendium of Soil Fungi; Academic Press: Cambridge, MA, USA, 1980. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Springer: Berlin/Heidelberg, Germany, 2009; Volume 519. [Google Scholar]
- Mao, Z.; Xue, M.; Gu, G.; Wang, W.; Li, D.; Lai, D.; Zhou, L. Lophiostomin A–D: New 3, 4-dihydroisocoumarin derivatives from the endophytic fungus Lophiostoma sp. Sigrf10. RSC Adv. 2020, 10, 6985–6991. [Google Scholar] [CrossRef] [Green Version]
- Shushni, M.A.M.; Azam, F.; Lindequist, U. Oxasetin from Lophiostoma sp. of the Baltic Sea: Identification, in silico binding mode prediction and antibacterial evaluation against fish pathogenic bacteria. Nat. Prod. Commun. 2013, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Motaal, F.F.; Nassar, M.S.; El-Zayat, S.A.; El-Sayed, M.A.; Ito, S.I. Antifungal activity of endophytic fungi isolated from Egyptian henbane (Hyoscyamus muticus L.). Pak. J. Bot. 2010, 42, 2883–2894. [Google Scholar]
- Premjanu, N.; Jaynthy, C.; Diviya, S. Antifungal activity of endophytic fungi isolated from lannea coromandelica–an insilico approach. Int. J. Pharm. Pharm. Sci. 2016, 8, 207–210. [Google Scholar]
- Rouissi, W.; Ugolini, L.; Martini, C.; Lazzeri, L.; Mari, M. Control of Postharvest Fungal Pathogens by Antifungal Compounds from Penicillium expansum. J. Food Prot. 2013, 76, 1879–1886. [Google Scholar] [CrossRef]
- Li, H.; Wei, J.; Pan, S.-Y.; Gao, J.-M.; Tian, J.-M. Antifungal, phytotoxic and toxic metabolites produced by Penicillium purpurogenum. Nat. Prod. Res. 2014, 28, 2358–2361. [Google Scholar] [CrossRef] [PubMed]
- Korejo, F.; Ali, S.A.; Shafique, H.A.; Sultana, V.; Ara, J.; Ehteshamul-Haque, S. Antifungal and antibacterial activity of endophytic Penicillium species isolated from Salvadora species. Pak. J. Bot. 2014, 46, 2313–2318. [Google Scholar]
- Urooj, F.; Farhat, H.; Ali, S.A.; Ahmed, M.; Sultana, V.; Shams, Z.I.; Ara, J.; Ehteshamul-Haque, S. Role of endophytic Penicillium species in suppressing the root rotting fungi of sunflower. Pak. J. Bot. 2018, 50, 1621–1628. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sample Code | Sampling Period | Grapevine Variety | Plant Part | Growing Locality (Farming System) |
|---|---|---|---|---|
| Z-MT-G-S6 | January 2019 | Muller Thurgau | canes | Prague (conventional) |
| Z-RR-G-S | January 2019 | Riesling Rheinhessen | canes | Prague (conventional) |
| Z-MT-KH-S1 | January 2019 | Muller Thurgau | canes | Kutna Hora (organic) |
| Z-MT-KH-S2 | January 2019 | Muller Thurgau | canes | Kutna Hora (organic) |
| Z-RM-KH-S1 | January 2019 | Pinot Noir | canes | Kutna Hora (organic) |
| Z-RM-KH-S2 | January 2019 | Pinot Noir | canes | Kutna Hora (organic) |
| J-MT-G-L2 | May 2019 | Muller Thurgau | leaves | Prague (conventional) |
| J-RR-G-S2 | May 2019 | Riesling Rheinhessen | canes | Prague (conventional) |
| J-RS-KH-L1 | May 2019 | Pinot Gris | leaves | Kutna Hora (organic) |
| J-RS-KH-L2 | May 2019 | Pinot Gris | leaves | Kutna Hora (organic) |
| J-MT-KH-S4 | May 2019 | Muller Thurgau | canes | Kutna Hora (organic) |
| J-RM-KH-S3 | May 2019 | Pinot Noir | canes | Kutna Hora (organic) |
| J-RM-KH-S4 | May 2019 | Pinot Noir | canes | Kutna Hora (organic) |
| J-RM-KH-S5 | May 2019 | Pinot Noir | canes | Kutna Hora (organic) |
| J-RR-KH-S2 | May 2019 | Riesling Rheinhessen | canes | Kutna Hora (organic) |
| J-RR-KH-S3 | May 2019 | Riesling Rheinhessen | canes | Kutna Hora (organic) |
| L-MT-KH-L5 | August 2019 | Muller Thurgau | leaves | Kutna Hora (organic) |
| L-RS-KH-L4 | August 2019 | Pinot Gris | leaves | Kutna Hora (organic) |
| L-RR-KH-L4 | August 2019 | Riesling Rheinhessen | leaves | Kutna Hora (organic) |
| L-RM-KH-S6 | August 2019 | Pinot Noir | canes | Kutna Hora (organic) |
| P-RM-G-L1 | October 2019 | Pinot Noir | leaves | Prague (conventional) |
| P-RS-G-S2 | October 2019 | Pinot Gris | canes | Prague (conventional) |
| P-MT-KH-L7 | October 2019 | Muller Thurgau | leaves | Kutna Hora (organic) |
| P-RM-KH-L7 | October 2019 | Pinot Noir | leaves | Kutna Hora (organic) |
| MT-M1 | September 2019 | Muller Thurgau | berries | Prague (conventional) |
| MT-M4 | September 2019 | Muller Thurgau | berries | Prague (conventional) |
| RR-M1 | September 2019 | Riesling Rheinhessen | berries | Prague (conventional) |
| RR-M2 | September 2019 | Riesling Rheinhessen | berries | Prague (conventional) |
| RS-M2 | September 2019 | Pinot Gris | berries | Prague (conventional) |
| Sample Code 1 | Sample Matrix | Endophyte Species Taxonomy | Ability to Produce Siderophores 2 | Antioxidant Activity (mgAA L−1) | Antifungal Activity 3 to: | ||
|---|---|---|---|---|---|---|---|
| Botrytis cinerea | Fusarium solani | Mucor plumbeus | |||||
| Z-MT-G-S6 | canes | Cladosporium cladosporioides | 1 | 12.4 | 2 | 2 | 2 |
| Z-RR-G-S | canes | Alternaria arborescens | 0 | 13.8 | 3 | 2 | 3 |
| Z-MT-KH-S1 | canes | Diatrype stigma | 3 | 17.5 | 2 | 2 | 2 |
| Z-MT-KH-S2 | canes | Didymella negriana | 0 | 4.8 | 3 | 2 | 2 |
| Z-RM-KH-S1 | canes | Aspergillus pseudodeflectus | 2 | 21.8 | 3 | 3 | 2 |
| Z-RM-KH-S2 | canes | Aspergillus niger | 3 | 13.4 | 2 | 4 | 4 |
| J-MT-G-L2 | leaves | Epicoccum nigrum | 2 | 17.6 | 3 | 3 | 2 |
| J- RR-G-S2 | canes | Pleurophoma ossicola | 0 | 2.7 | 2 | 3 | 3 |
| J-RS-KH-L1 | leaves | Sporocadus rosigena | 1 | 6.6 | 3 | 3 | 2 |
| J-RS-KH-L2 | leaves | Dendrophoma juglandina | 2 | 0 | 2 | 2 | 2 |
| J-MT-KH-S4 | canes | Pseudogymnoascus pannorum | 0 | 0 | 2 | 3 | 2 |
| J-RM-KH-S3 | canes | Aureobasidium pullulans | 1 | 6.4 | 2 | 2 | 2 |
| J-RM-KH-S4 | canes | Didymella sancta | 1 | 8.3 | 2 | 2 | 2 |
| J-RM-KH-S5 | canes | Cladosporium herbarum | 0 | 7.5 | 3 | 3 | 2 |
| J-RR-KH-S2 | canes | Phaeosphaeriaceae sp. | 0 | 7.8 | 2 | 2 | 2 |
| J-RR-KH-S3 | canes | Neosetophoma shoemakeri | 2 | 6.7 | 2 | 3 | 3 |
| L-MT-KH-L5 | leaves | Aspergillus fumigatus | 1 | 9.3 | 2 | 4 | 3 |
| L-RS-KH-L4 | leaves | Lophiostoma corticola | 1 | 8.3 | 4 | 1 | 1 |
| L-RR-KH-L4 | leaves | Cladosporium herbarum | 0 | 8.1 | 2 | 3 | 2 |
| L-RM-KH-S6 | canes | Aureobasidium pullulans | 1 | 0 | 3 | 3 | 2 |
| P-RM-G-L1 | leaves | Alternaria astroemeriae | 0 | 7.5 | 3 | 3 | 2 |
| P-RS-G-S2 | canes | Aureobasidium pullulans | 1 | 7.1 | 3 | 3 | 2 |
| P-MT-KH-L7 | leaves | Cladosporium herbarum | 0 | 0 | 1 | 1 | 1 |
| P-RM-KH-L7 | leaves | Didymella sancta | 1 | 3.4 | 3 | 3 | 3 |
| MT-M1 | berries | Penicillium crustosum | 1 | 10.5 | 3 | 3 | 2 |
| MT-M4 | berries | Penicillium crustosum | 3 | 9.3 | 4 | 5 | 2 |
| RR-M1 | berries | Penicillium crustosum | 1 | 13.9 | 2 | 3 | 2 |
| RR-M2 | berries | Penicillium crustosum | 3 | 23.9 | 4 | 5 | 3 |
| RS-M2 | berries | Penicillium crustosum | 3 | 19.1 | 4 | 5 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulišová, M.; Vrublevskaya, M.; Lovecká, P.; Vrchotová, B.; Stránská, M.; Kolařík, M.; Kolouchová, I. Fungal Endophytes of Vitis vinifera—Plant Growth Promotion Factors. Agriculture 2021, 11, 1250. https://doi.org/10.3390/agriculture11121250
Kulišová M, Vrublevskaya M, Lovecká P, Vrchotová B, Stránská M, Kolařík M, Kolouchová I. Fungal Endophytes of Vitis vinifera—Plant Growth Promotion Factors. Agriculture. 2021; 11(12):1250. https://doi.org/10.3390/agriculture11121250
Chicago/Turabian StyleKulišová, Markéta, Maria Vrublevskaya, Petra Lovecká, Blanka Vrchotová, Milena Stránská, Miroslav Kolařík, and Irena Kolouchová. 2021. "Fungal Endophytes of Vitis vinifera—Plant Growth Promotion Factors" Agriculture 11, no. 12: 1250. https://doi.org/10.3390/agriculture11121250