Replacement of Albumin by Preovulatory Oviductal Fluid in Swim-Up Sperm Preparation Method Modifies Boar Sperm Parameters and Improves In Vitro Penetration of Oocytes

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
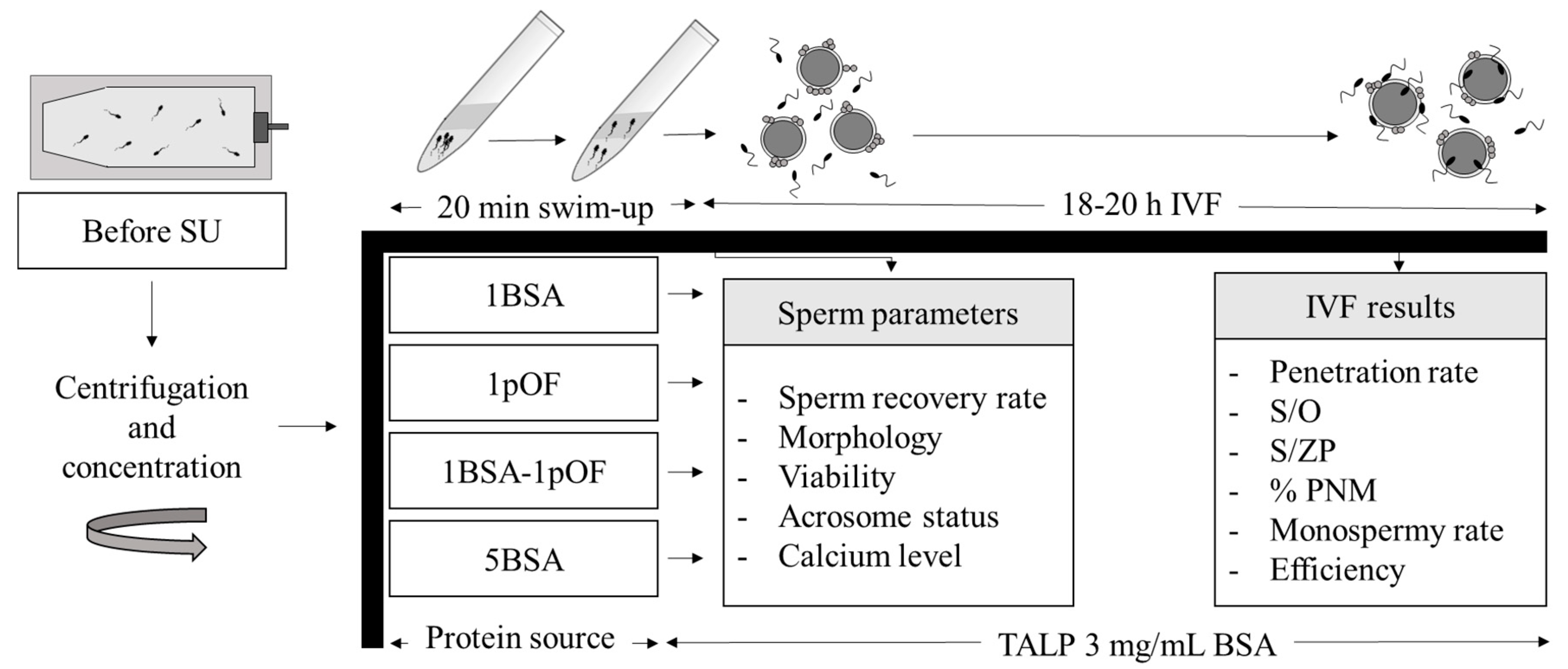
2. Materials and Methods
2.1. Ethics
2.2. Sperm Preparation
2.3. Evaluation of Sperm Functionality Parameters
2.3.1. Sperm Concentration and Morphology
2.3.2. Analysis of Motion Parameters by CASA
2.3.3. Sperm Viability and Acrosome Reaction
2.3.4. Intracellular Sperm Calcium Concentration
2.4. In Vitro Zygote Production
2.4.1. In Vitro Maturation of Cumulus-Oocyte Complexes (COCs)
2.4.2. In Vitro Fertilization (IVF)
2.5. Statistical Analysis
2.6. Experimental Design
3. Results
3.1. Sperm Recovery and Morphology
3.2. Sperm Motility, Motion Parameters (CASA), Sperm Viability and Acrosome Reaction
3.3. Intracellular Sperm Calcium
3.4. In Vitro Fertilization Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sakkas, D.; Ramalingam, M.; Garrido, N.; Barratt, C.L.R. Sperm selection in natural conception: What can we learn from Mother Nature to improve assisted reproduction outcomes? Hum. Reprod. Update 2015, 21, 711–726. [Google Scholar] [CrossRef] [Green Version]
- Luongo, C.; González-Brusi, L.; Cots-Rodríguez, P.; Izquierdo-Rico, M.J.; Avilés, M.; García-Vázquez, F.A. Sperm proteome after interaction with reproductive fluids in porcine: From the ejaculation to the fertilization site. Int. J. Mol. Sci. 2020, 21, 6060. [Google Scholar] [CrossRef] [PubMed]
- Matás, C.; Vieira, L.; García-Vázquez, F.A.; Avilés-López, K.; López-Úbeda, R.; Carvajal, J.A.; Gadea, J. Effects of centrifugation through three different discontinuous Percoll gradients on boar sperm function. Anim. Reprod. Sci. 2011, 127, 62–72. [Google Scholar] [CrossRef]
- Matás, C.; Sansegundo, M.; Ruiz, S.; García-Vázquez, F.A.; Gadea, J.; Romar, R.; Coy, P. Sperm treatment affects capacitation parameters and penetration ability of ejaculated and epididymal boar spermatozoa. Theriogenology 2010, 74, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, M.A.M.M.; Henning, H.H.W.; Stout, T.A.E.; Vos, P.L.A.M.; Gadella, B.M. Designing 3-Dimensional In Vitro Oviduct Culture Systems to Study Mammalian Fertilization and Embryo Production. Ann. Biomed. Eng. 2017, 45, 1731–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romar, R.; Cánovas, S.; Matás, C.; Gadea, J.; Coy, P. Pig in vitro fertilization: Where are we and where do we go? Theriogenology 2019, 137, 113–121. [Google Scholar] [CrossRef]
- Alvarez, J.G.; Lasso, J.L.; Blasco, L.; Nuñez, R.C.; Heyner, S.; Caballero, P.P.; Storey, B.T. Centrifugation of human spermatozoa induces sublethal damage; separation of human spermatozoa from seminal plasma by a dextran swim-up procedure without centrifugation extends their motile lifetime. Hum. Reprod. 1993, 8, 1087–1092. [Google Scholar] [CrossRef]
- Amiri, I.; Ghorbani, M.; Heshmati, S. Comparison of the DNA fragmentation and the sperm parameters after processing by the density gradient and the swim up methods. J. Clin. Diagn. Res. 2012, 6, 1451–1453. [Google Scholar] [CrossRef] [PubMed]
- Parrish, J.J.; Krogenaes, A.; Susko-Parrish, J.L. Effect of bovine sperm separation by either swim-up or Percoll method on success of in vitro fertilization and early embryonic development. Theriogenology 1995, 44, 859–869. [Google Scholar] [CrossRef]
- Arias, M.E.; Andara, K.; Briones, E.; Felmer, R. Bovine sperm separation by Swim-up and density gradients (Percoll and BoviPure): Effect on sperm quality, function and gene expression. Reprod. Biol. 2017, 17, 126–132. [Google Scholar] [CrossRef]
- Clarke, R.N.; Johnson, L.A. Effect of liquid storage and cryopreservation of boar spermatozoa on acrosomal integrity and the penetration of zona-free hamster ova in vitro. Gamete Res. 1987, 16, 193–204. [Google Scholar] [CrossRef]
- Holt, W.V.; Hernandez, M.; Warrell, L.; Satake, N. The long and the short of sperm selection in vitro and in vivo: Swim-up techniques select for the longer and faster swimming mammalian sperm. J. Evol. Biol. 2010, 23, 598–608. [Google Scholar] [CrossRef]
- Cánovas, S.; Ivanova, E.; Romar, R.; García-Martínez, S.; Soriano-Úbeda, C.; García-Vázquez, F.A.; Saadeh, H.; Andrews, S.; Kelsey, G.; Coy, P. DNA methylation and gene expression changes derived from assisted reproductive technologies can be decreased by reproductive fluids. Elife 2017, 6, e23670. [Google Scholar] [CrossRef]
- García-Martínez, S.; Sánchez Hurtado, M.A.; Gutiérrez, H.; Sánchez Margallo, F.M.; Romar, R.; Latorre, R.; Coy, P.; López Albors, O. Mimicking physiological O2 tension in the female reproductive tract improves assisted reproduction outcomes in pig. MHR Basic Sci. Reprod. Med. 2018, 24, 260–270. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, S.; Latorre, R.; Sánchez-Hurtado, M.A.; Sánchez-Margallo, F.M.; Bernabò, N.; Romar, R.; López-Albors, O.; Coy, P. Mimicking the temperature gradient between the sow’s oviduct and uterus improves in vitro embryo culture output. Mol. Hum. Reprod. 2020, 26, 748–759. [Google Scholar] [CrossRef]
- Hamze, J.G.; Jiménez-Movilla, M.; Romar, R. Sperm-Binding Assay Using an In Vitro 3D Model of the Mammalian Cumulus-Oocyte Complex. Curr. Protoc. Toxicol. 2020, 86. [Google Scholar] [CrossRef] [PubMed]
- Coy, P.; Gadea, J.; Romar, R.; Matás, C.; García, E. Effect of in vitro fertilization medium on the acrosome reaction, cortical reaction, zona pellucida hardening and in vitro development in pigs. Reproduction 2002, 124, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Romar, R.; Funahashi, H.; Coy, P. In vitro fertilization in pigs: New molecules and protocols to consider in the forthcoming years. Theriogenology 2016, 85, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Acuña, O.S.; Avilés, M.; López-Úbeda, R.; Guillén-Martínez, A.; Soriano-Úbeda, C.; Torrecillas, A.; Coy, P.; Izquierdo-Rico, M.J. Differential gene expression in porcine oviduct during the oestrous cycle. Reprod. Fertil. Dev. 2017, 29, 2387–2399. [Google Scholar] [CrossRef]
- Küçük, N.; Lopes, J.S.; Soriano-Úbeda, C.; Hidalgo, C.O.; Romar, R.; Gadea, J. Effect of oviductal fluid on bull sperm functionality and fertility under non-capacitating and capacitating incubation conditions. Theriogenology 2020, 158, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Day, B.N.; Lim, J.G.; Lee, H.T.; Chung, K.S. Effects of oviductal fluid and heparin on fertility and characteristics of porcine spermatozoa. Zygote 1997, 5, 61–65. [Google Scholar] [CrossRef]
- McCauley, T.C.; Buhi, W.C.; Wu, G.M.; Mao, J.; Caamaño, J.; Didion, B.A.; Day, B.N. Oviduct-Specific Glycoprotein Modulates Sperm-Zona Binding and Improves Efficiency of Porcine Fertilization In Vitro1. Biol. Reprod. 2003, 69, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Coy, P.; Lloyd, R.; Romar, R.; Satake, N.; Matas, C.; Gadea, J.; Holt, W.V. Effects of porcine pre-ovulatory oviductal fluid on boar sperm function. Theriogenology 2010, 74, 632–642. [Google Scholar] [CrossRef]
- Kumaresan, A.; Johannisson, A.; Humblot, P.; Bergqvist, A.-S. Oviductal fluid modulates the dynamics of tyrosine phosphorylation in cryopreserved boar spermatozoa during capacitation. Mol. Reprod. Dev. 2012, 79, 525–540. [Google Scholar] [CrossRef]
- Kumaresan, A.; González, R.; Johannisson, A.; Berqvist, A.S. Dynamic quantification of intracellular calcium and protein tyrosine phosphorylation in cryopreserved boar spermatozoa during short-time incubation with oviductal fluid. Theriogenology 2014, 82, 1145–1153. [Google Scholar] [CrossRef]
- Zapata-Carmona, H.; Soriano-Úbeda, C.; París-Oller, E.; Matás, C. Periovulatory oviductal fluid decreases sperm protein kinase A activity, tyrosine phosphorylation, and in vitro fertilization in pig. Andrology 2020, 8, 756–768. [Google Scholar] [CrossRef] [PubMed]
- Gadea, J.; Sellés, E.; Marco, M.A.; Coy, P.; Matás, C.; Romar, R.; Ruiz, S. Decrease in glutathione content in boar sperm after cryopreservation. Effect of the addition of reduced glutathione to the freezing and thawing extenders. Theriogenology 2004, 62, 690–701. [Google Scholar] [CrossRef] [PubMed]
- González-Abreu, D.; García-Martínez, S.; Fernández-Espín, V.; Romar, R.; Gadea, J. Incubation of boar spermatozoa in viscous media by addition of methylcellulose improves sperm quality and penetration rates during in vitro fertilization. Theriogenology 2017, 92, 14–23. [Google Scholar] [CrossRef]
- Yeste, M.; Fernández-Novell, J.M.; Ramió-Lluch, L.; Estrada, E.; Rocha, L.G.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Concha, I.I.; Ramírez, A.; Rodríguez-Gil, J.E. Intracellular calcium movements of boar spermatozoa during “in vitro” capacitation and subsequent acrosome exocytosis follow a multiple-storage place, extracellular calcium-dependent model. Andrology 2015, 3, 729–747. [Google Scholar] [CrossRef] [Green Version]
- Petters, R.M.; Wells, K.D. Culture of pig embryos. Biosci. Proc. 1993, 48, 61–73. [Google Scholar] [CrossRef]
- Lopes, J.S.; Canha-Gouveia, A.; París-Oller, E.; Coy, P. Supplementation of bovine follicular fluid during in vitro maturation increases oocyte cumulus expansion, blastocyst developmental kinetics, and blastocyst cell number. Theriogenology 2018, 126. [Google Scholar] [CrossRef]
- Abaigar, T.; Holt, W.V.; Harrison, R.A.P.; Del Barrio, G. Sperm subpopulations in Boar (Sus scrofa) and Gazelle (Gazella dama mhorr) semen as revealed by pattern analysis of computer-assisted motility assessments. Biol. Reprod. 1999, 60, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballester, L.; Romero-Aguirregomezcorta, J.; Soriano-Úbeda, C.; Matás, C.; Romar, R.; Coy, P. Timing of oviductal fluid collection, steroid concentrations, and sperm preservation method affect porcine in vitro fertilization efficiency. Fertil. Steril. 2014, 102, 1762–1768. [Google Scholar] [CrossRef] [PubMed]
- Coy, P.; Canovas, S.; Mondejar, I.; Saavedra, M.D.; Romar, R.; Grullon, L.; Matas, C.; Aviles, M. Oviduct-specific glycoprotein and heparin modulate sperm-zona pellucida interaction during fertilization and contribute to the control of polyspermy. Proc. Natl. Acad. Sci. USA 2008, 105, 15809–15814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.K.; Jee, B.C.; Kim, S.H. Histone methylation and acetylation in ejaculated human sperm: Effects of swim-up and smoking. Fertil. Steril. 2015, 103, 1425–1431. [Google Scholar] [CrossRef]
- Magdanz, V.; Boryshpolets, S.; Ridzewski, C.; Eckel, B.; Reinhardt, K. The motility-based swim-up technique separates bull sperm based on differences in metabolic rates and tail length. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Park, C.H.; Lee, S.G.; Choi, D.H.; Lee, C.K. A modified swim-up method reduces polyspermy during in vitro fertilization of porcine oocytes. Anim. Reprod. Sci. 2009, 115, 169–181. [Google Scholar] [CrossRef]
- Boerke, A.; Tsai, P.S.; Garcia-Gil, N.; Brewis, I.A.; Gadella, B.M. Capacitation-dependent reorganization of microdomains in the apical sperm head plasma membrane: Functional relationship with zona binding and the zona-induced acrosome reaction. Theriogenology 2008, 70, 1188–1196. [Google Scholar] [CrossRef]
- Espinosa, F.; López-González, I.; Muñoz-Garay, C.; Felix, R.; De la Vega-Beltrán, J.L.; Kopf, G.S.; Visconti, P.E.; Darszon, A. Dual regulation of the T-type Ca(2+) current by serum albumin and beta-estradiol in mammalian spermatogenic cells. FEBS Lett. 2000, 475, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Alcântara-Neto, A.S.; Fernandez-Rufete, M.; Corbin, E.; Tsikis, G.; Uzbekov, R.; Garanina, A.S.; Coy, P.; Almiñana, C.; Mermillod, P. Oviduct fluid extracellular vesicles regulate polyspermy during porcine invitro fertilisation. Reprod. Fertil. Dev. 2020, 32. [Google Scholar] [CrossRef]
- Navarro-Serna, S.; Hachem, A.; Canha-Gouveia, A.; Hanbashi, A.; Garrappa, G.; Lopes, J.S.; Paris-Oller, E.; Bermejo-Álvarez, P.; Matas, C.; Romar, R.; et al. Generation of Nonmosaic, Two-Pore Channel 2 Biallelic Knockout Pigs in One Generation by CRISPR-Cas9 Microinjection Before Oocyte Insemination. Cris. J. 2021, 4, 1–16. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Boar Ejaculated Semen | pOF Concentration | Sperm Treatment | Main Results | Reference |
|---|---|---|---|---|
| Fresh | Pure, 1–5% (v/v) | Centrifugation followed by 1.5 h incubation | ↑ sperm capacitation and prevents acrosome reaction | [21] |
| F-T | 10 µg/mL glycoproteins | 1 h incubation followed by centrifugation | = acrosome reaction (induced by calcium ionophore) | [22] |
| Fresh | 50 µg/mL protein fraction (< or >100 kDa) | Percoll followed by 20 min incubation | ↑ sperm viability, acrosome integrity and polyspermy ↓ sperm membrane fluidity | [23] |
| F-T | Pure, 20% (v/v) | Centrifugation followed by 6 h incubation | ↑ pTyr | [24] |
| F-T | Pure, 20% (v/v) | Centrifugation followed by 1 h incubation | ↑ pTyr, sperm intracellular calcium and motility | [25] |
| Fresh | Pure, 1% (v/v) | Swim-up for 20 min * | ↑ IVF efficiency and improve epigenetic patterns ** | [13] |
| Fresh | Pure, 1% (v/v) | 3 h incubation | ↓ pTyr, PKA and penetration rate | [26] |
| Group | n | Sperm Recovery Rate (%) |
|---|---|---|
| 1BSA | 16 | 1.11 ± 0.10 ab |
| 1pOF | 16 | 1.02 ± 0.10 a |
| 1BSA-1pOF | 16 | 1.20 ± 0.12 ab |
| 5BSA | 16 | 1.52 ± 0.11 b |
| p-value | 0.01 |
| Group | n | Normal Morphology (%) | Bent Tails (%) | Distal Droplets (%) | Proximal Droplets (%) | Total Droplets (%) |
|---|---|---|---|---|---|---|
| Before SU | 10 | 82.95 ± 1.47 a | 2.05 ± 0.65 a | 9.60 ± 0.99 a | 4.65 ± 0.44 a | 14.25 ± 1.12 a |
| 1BSA | 10 | 98.50 ± 0.40 b | 0.40 ± 0.22 bc | 0.50 ± 0.22 b | 0.50 ± 0.22 b | 1.00 ± 0.21 b |
| 1pOF | 10 | 97.10 ± 0.48 b | 0.50 ± 0.22 bc | 1.50 ± 0.40 b | 0.90 ± 0.31 b | 2.40 ± 0.56 b |
| 1BSA-1pOF | 10 | 97.50 ± 0.45 b | 0.30 ± 0.21 c | 1.60 ± 0.37 b | 0.60 ± 0.22 b | 2.20 ± 0.30 b |
| 5BSA | 10 | 93.70 ± 0.67 c | 1.30 ± 0.30 ab | 3.50 ± 0.58 c | 1.30 ± 0.34 b | 4.80 ± 0.65 c |
| p-value | < 0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Group | n | Slow Velocity—Non-Progressive (%) | Medium Velocity—Non-Progressive (%) | Medium Velocity—Progressive (%) | Fast Velocity—Progressive (%) |
|---|---|---|---|---|---|
| 1BSA | 1188 | 36.53 ± 1.40 a | 30.05 ± 1.33 ab | 23.99 ± 1.24 a | 9.43 ± 0.85 a |
| 1pOF | 2104 | 43.42 ± 1.48 b | 33.00 ± 1.40 a | 19.13 ± 1.17 b | 4.45 ± 0.62 b |
| 1BSA-1pOF | 1124 | 39.83 ± 1.07 ab | 29.42 ± 0.99 b | 20.87 ± 0.89 ab | 9.89 ± 0.65 a |
| 5BSA | 2329 | 36.11 ± 1.00 a | 28.30 ± 0.93 b | 21.47 ± 0.85 ab | 14.13 ± 0.72 c |
| p-value | <0.01 | 0.01 | 0.04 | <0.01 |
| Group | n | Viable Sperm with Intact Acrosome (%) | Viable Sperm with Damaged Acrosome (%) | Total Sperm with Acrosome Damaged (%) |
|---|---|---|---|---|
| Before SU | 14 | 79.71 ± 1.32 a | 9.62 ± 1.13 a | 17.54 ± 1.62 a |
| 1BSA | 14 | 89.34 ± 1.34 b | 0.74 ± 0.09 b | 3.61 ± 0.46 b |
| 1pOF | 14 | 90.67 ± 0.92 b | 0.35 ± 0.04 c | 2.54 ± 0.31 b |
| 1BSA-1pOF | 14 | 87.85 ± 1.18 b | 0.45 ± 0.11 c | 3.32 ± 0.33 b |
| 5BSA | 14 | 87.06 ± 1.74 b | 0.89 ± 0.11 b | 4.16 ± 0.54 b |
| p-value | <0.01 | <0.01 | <0.01 |
| Group | n | Viable Sperm with High Calcium Concentration (%) | Total Sperm with High Calcium Concentration (%) | Total Viable Sperm (%) |
|---|---|---|---|---|
| Before SU | 10 | 28.88 ± 9.03 a | 55.72 ± 10.99 | 64.48 ± 2.45 |
| 1BSA | 10 | 24.75 ± 2.36 a | 58.35 ± 6.32 | 59.57 ± 4.38 |
| 1pOF | 10 | 30.24 ± 1.87 a | 54.99 ± 6.90 | 69.54 ± 5.72 |
| 1BSA-1pOF | 10 | 27.76 ± 1.57 a | 52.62 ± 7.01 | 69.03 ± 7.18 |
| 5BSA | 10 | 17.19 ± 1.15 b | 46.65 ± 7.92 | 60.91 ± 6.20 |
| p-value | 0.03 | 0.85 | 0.83 |
| Group | n | Penetration (%) | S/O | S/ZP | PNM (%) | Monospermy (%) | Efficiency (%) |
|---|---|---|---|---|---|---|---|
| 1BSA | 178 | 74.16 ± 3.29 a | 2.65 ± 0.17 a | 11.28 ± 0.80 a | 99.42 ± 0.58 | 34.09 ± 4.14 ab | 25.28 ± 3.27 |
| 1pOF | 189 | 86.77 ± 2.47 b | 2.49 ± 0.12 a | 9.60 ± 0.49 ab | 100 | 28.66 ± 3.54 a | 24.87 ± 3.15 |
| 1BSA-1pOF | 197 | 76.65 ± 3.02 ab | 2.25 ± 0.10 ab | 7.69 ± 0.45 bc | 98.94 ± 0.75 | 31.79 ± 3.80 a | 24.37 ± 3.07 |
| 5BSA | 199 | 59.30 ± 3.49 c | 1.92 ± 0.11 b | 6.19 ± 0.51 c | 100 | 48.31 ± 4.62 b | 28.64 ± 3.21 |
| p-value | <0.01 | <0.01 | <0.01 | 0.312 | <0.01 | 0.754 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Serna, S.; París-Oller, E.; Simonik, O.; Romar, R.; Gadea, J. Replacement of Albumin by Preovulatory Oviductal Fluid in Swim-Up Sperm Preparation Method Modifies Boar Sperm Parameters and Improves In Vitro Penetration of Oocytes. Animals 2021, 11, 1202. https://doi.org/10.3390/ani11051202
Navarro-Serna S, París-Oller E, Simonik O, Romar R, Gadea J. Replacement of Albumin by Preovulatory Oviductal Fluid in Swim-Up Sperm Preparation Method Modifies Boar Sperm Parameters and Improves In Vitro Penetration of Oocytes. Animals. 2021; 11(5):1202. https://doi.org/10.3390/ani11051202
Chicago/Turabian StyleNavarro-Serna, Sergio, Evelyne París-Oller, Ondrej Simonik, Raquel Romar, and Joaquín Gadea. 2021. "Replacement of Albumin by Preovulatory Oviductal Fluid in Swim-Up Sperm Preparation Method Modifies Boar Sperm Parameters and Improves In Vitro Penetration of Oocytes" Animals 11, no. 5: 1202. https://doi.org/10.3390/ani11051202