
Evaluation of Spatio-Temporal Patterns of Predation Risk to Forest Grouse Nests in the Central European Mountain Regions

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Potential Predator Monitoring
2.4. Statistical Analyses
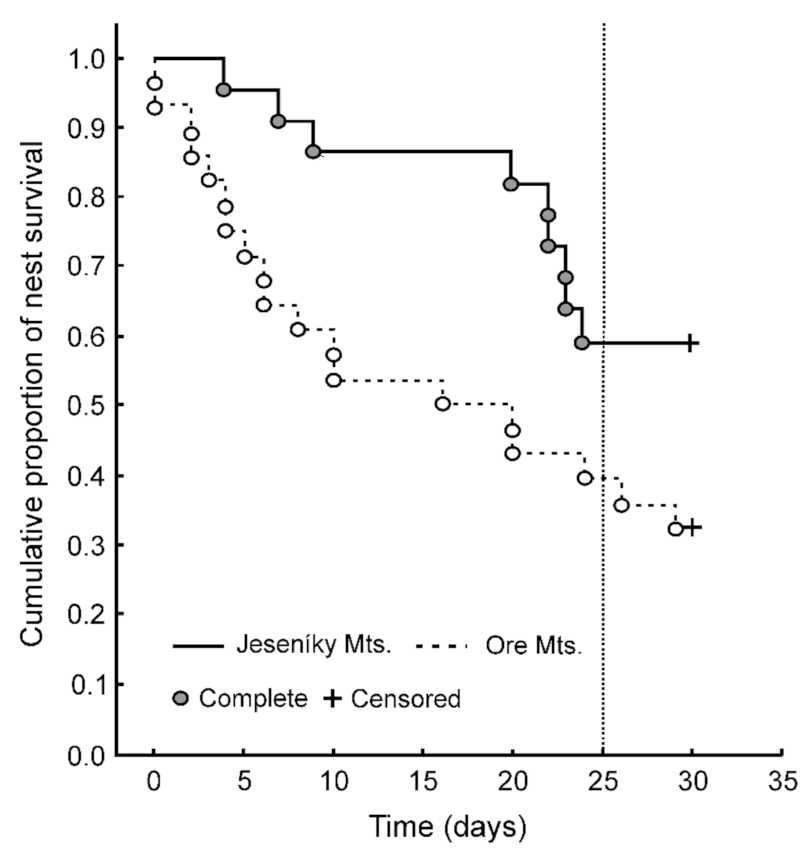
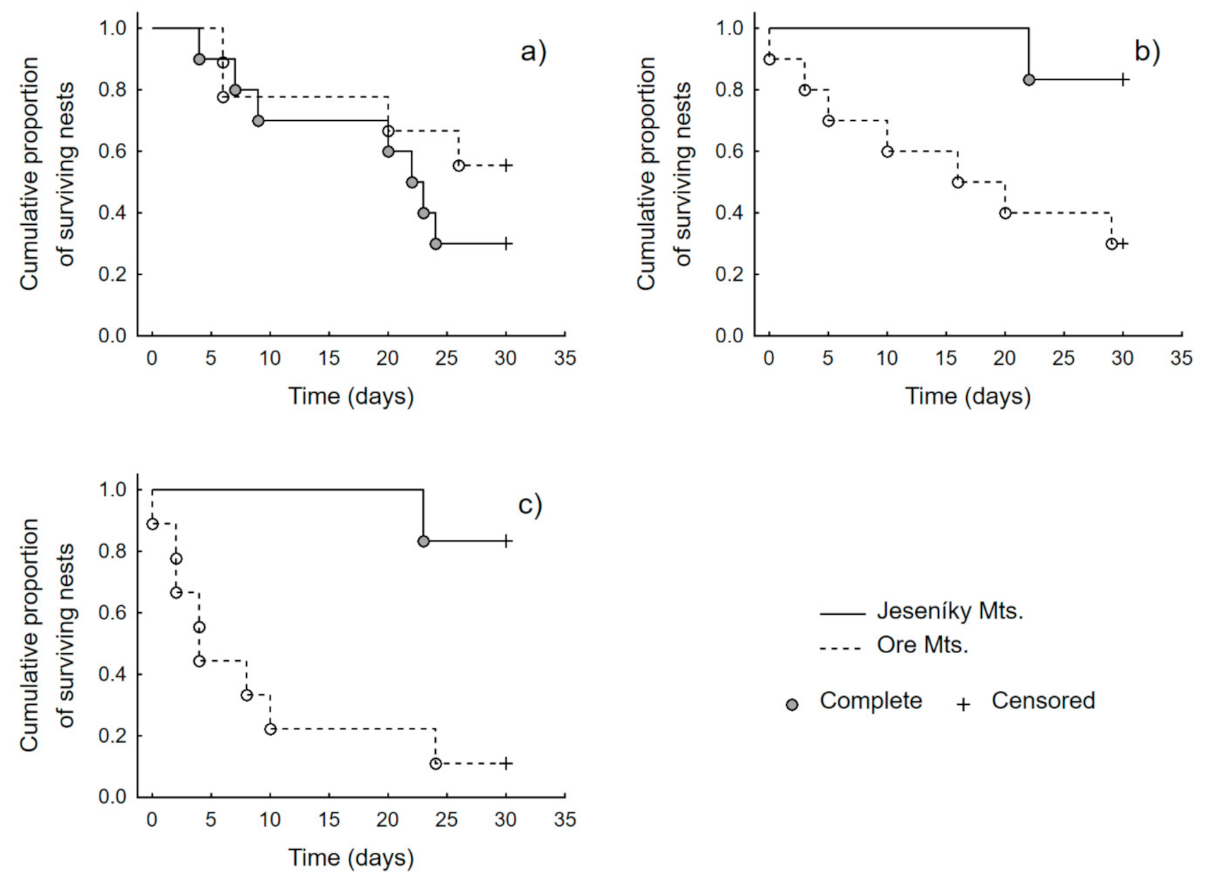
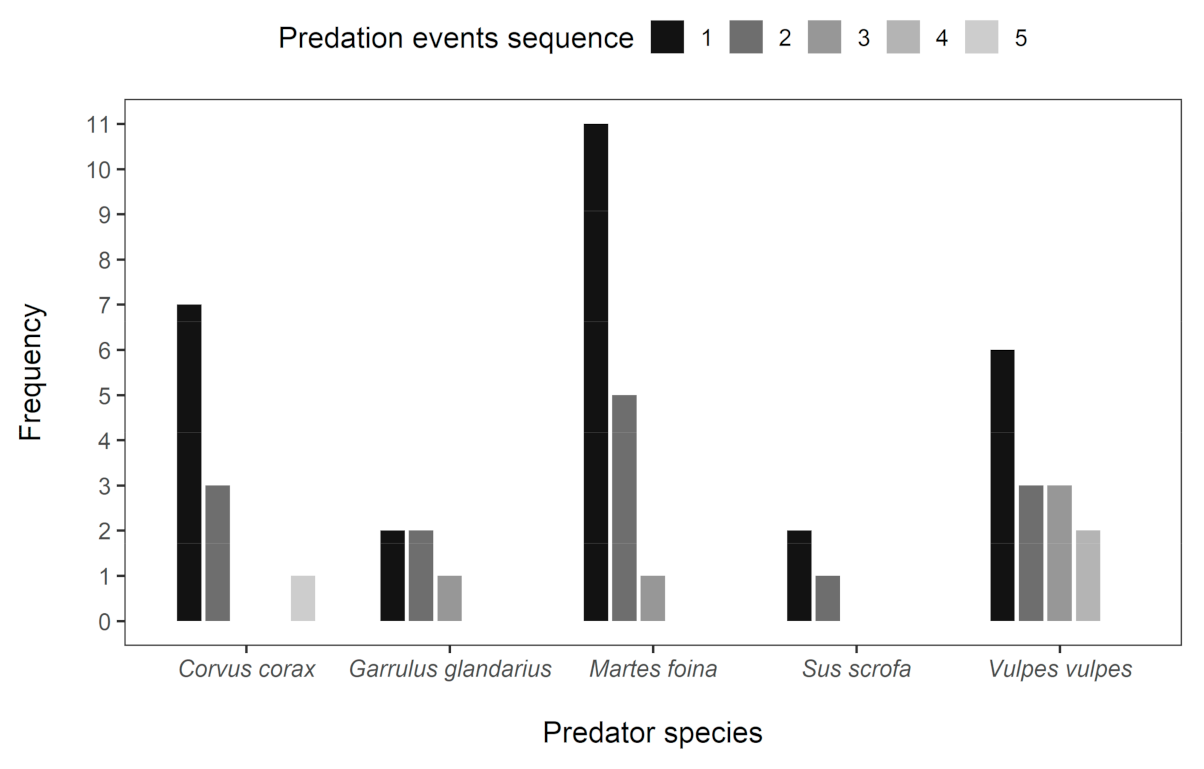
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chamberlain, D.E.; Fuller, R.J.; Bunce, R.G.H.; Duckworth, J.C.; Shrubb, M. Changes in the abundance of farmland birds in relation to the timing of agricultural intensification in England and Wales. J. Appl. Ecol. 2000, 37, 771–788. [Google Scholar] [CrossRef] [Green Version]
- Isaksson, D.; Wallander, J.; Larsson, M. Managing predation on ground-nesting birds: The effectiveness of nest exclosures. Biol. Conserv. 2007, 136, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Roos, S.; Smart, J.; Gibbons, D.W.; Wilson, J.D. A review of predation as a limiting factor for bird populations in mesopredator-rich landscapes: A case study of the UK. Biol. Rev. 2018, 93, 1915–1937. [Google Scholar] [CrossRef]
- Kaasiku, T.; Rannap, R.; Kaart, T. Managing coastal grasslands for an endangered wader species can give positive results only when expanding the area of open landscape. J. Nat. Conserv. 2019, 48, 12–19. [Google Scholar] [CrossRef]
- Macdonald, M.A.; Bolton, M. Predation on wader nests in Europe. IBIS 2008, 150, 54–73. [Google Scholar] [CrossRef]
- Tamis, W.L.M.; Heemskerk, P. A longitudinal study of the effects of trees, geese and avian predators on breeding wader meadow birds: The case of the Demmerik polder, the Netherlands. Eur. J. Wildl. Res. 2020, 66. [Google Scholar] [CrossRef]
- Warren, P.; Land, C.; Hesford, N.; Baines, D. Conserving Black Grouse Lyrurus tetrix in southern Scotland: Evidence for the need to retain large contiguous moorland habitat within a forest-moorland landscape. Bird Study 2019, 66, 494–502. [Google Scholar] [CrossRef]
- Jahren, T.; Storaas, T.; Willebrand, T.; Fossland Moa, P.; Hagen, B.R. Declining reproductive output in capercaillie and black grouse-16 countries and 80 years. Anim. Biol. 2016, 66, 363–400. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yang, L.; Wu, S.; Xia, W.; Yang, L.; Li, M.; Chen, M.; Luan, X. Use of historical data to improve conservation of the black grouse (Lyrurus tetrix) in Northeast China. Ecosphere 2020, 11. [Google Scholar] [CrossRef]
- Melin, M.; Mehtätalo, L.; Helle, P.; Ikonen, K.; Packalen, T. Decline of the boreal willow grouse (Lagopus lagopus) has been accelerated by more frequent snow-free springs. Sci. Rep. 2020, 10, 6987. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.A.; Gómez-Serrano, M.Á.; López-López, P. Population Decline of the Capercaillie Tetrao urogallus aquitanicus in the Central Pyrenees. Ardeola 2020, 67, 285. [Google Scholar] [CrossRef]
- Wilkinson, N.I.; Eaton, M.A.; Marshall, G.; Haysom, S. The population status of Capercaillie Tetrao urogallus in Scotland during winter 2015–2016. Bird Study 2018, 65, 20–35. [Google Scholar] [CrossRef]
- Baines, D. The implications of grazing and predator management on the habitats and breeding success of black grouse Tetrao Tetrix. J. Appl. Ecol. 1996, 33, 54–62. [Google Scholar] [CrossRef]
- Warren, P.K.; Baines, D. Dispersal, survival and causes of mortality in black grouse Tetrao tetrix in northern England. Wildl. Biol. 2002, 8, 91–97. [Google Scholar] [CrossRef]
- Warren, P.; Baines, D. Expanding the range of black grouse Lyrurus tetrix in northern England—Can wild females be successfully translocated? Wildl. Biol. 2018, 2018, wlb.00435. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, G.X.; Alatalo, R.V.; Helle, P.; Siitari, H. Individual and Environmental Determinants of Daily Black Grouse Nest Survival Rates at Variable Predator Densities. Ann. Zool. Fenn. 2010, 47, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Catt, D.C.; Dugan, D.; Green, R.E.; Moss, R.; Picozzi, N.; Summers, R.W.; Tyler, G.A. Collisions against Fences by Woodland Grouse in Scotland. Forestry 1994, 67, 105–118. [Google Scholar] [CrossRef]
- Kämmerle, J.-L.; Storch, I. Predation, predator control and grouse populations: A review. Wildl. Biol. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pekkola, M.; Alatalo, R.; Pöysä, H.; Siitari, H. Seasonal survival of young and adult black grouse females in boreal forests. Eur. J. Wildl. Res. 2014, 60, 477–488. [Google Scholar] [CrossRef]
- Ciach, M. Rapid decline of an isolated population of the black grouse Tetrao tetrix: The crisis at the southern limit of the range. Eur. J. Wildl. Res. 2015, 2015, 623–627. [Google Scholar] [CrossRef] [Green Version]
- Svobodová, J.; Segelbacher, G.; Höglund, J. Genetic variation in Black Grouse populations with different lekking systems in the Czech Republic. J. Ornithol. 2011, 152, 37–44. [Google Scholar] [CrossRef]
- Svobodová, J.; Bejček, V.; Málková, P.; Štastný, K. Low survival rate of Black Grouse (Tetrao tetrix) in maturating forest growths in the Krušné hory Mts. Sylvia 2011, 47, 77–89. [Google Scholar]
- Caizergues, A.; Ellison, L.N. Age-specific reproductive performance of Black Grouse Tetrao tetrix females. Bird Study 2000, 47, 344–351. [Google Scholar] [CrossRef]
- Svobodová, J.; Albrecht, T.; Šálek, M. The relationship between predation risk and occurrence of black grouse (Tetrao tetrix) in a highly fragmented landscape: An experiment based on artificial nests. Ecoscience 2004, 11, 421–427. [Google Scholar] [CrossRef]
- Svobodová, J. Undying star—The Black Grouse (Tetrao tetrix). Sylvia 2005, 41, 17–33. [Google Scholar]
- Merta, D.; Bobek, B.; Furtek, J.; Kolecki, M. Distribution and number of black grouse, Tetrao tetrix in southwestern Poland and the potential impact of predators upon nesting success of the species. Folia Zool. 2009, 58, 159–167. [Google Scholar]
- Storch, I. Grouse: Status Survey and Conservation Action Plan 2006–2010; World Pheasant Association: Fordingbridge, UK, 2007; ISBN 978-2-8317-1009-9. [Google Scholar]
- Šťastný, K.; Bejček, V.; Hudec, K. Atlas Hnízdního rozšíření Ptáků v České Republice (Nesting Distribution of Birds in the Czech Republic); Aventinum: Prague, Czech Republic, 2006. [Google Scholar]
- Wegge, P.; Ingul, H.; Pollen, V.O.; Halvorsrud, E.; Sivkov, A.V.; Hjeljord, O. Comparing predation on forest grouse nests by avian and mammalian predators in two contrasting boreal forest landscapes by the use of artificial nests. Ornis Fenn. 2012, 89, 145–156. [Google Scholar]
- Oja, R.; Pass, E.; Soe, E.; Ligi, K.; Anijalg, P.; Laurimaa, L.; Saarma, U.; Lõhmus, A.; Valdmann, H. Increased nest predation near protected capercaillie leks: A caveat against small reserves. Eur. J. Wildl. Res. 2018, 64. [Google Scholar] [CrossRef]
- Willebrand, T.; Marcström, V. On the Danger of Using Dummy Nests to Study Predation. Auk 1988, 105, 378–379. [Google Scholar] [CrossRef]
- Storaas, T. A Comparison of Losses in Artificial and Naturally Occurring Capercaillie Nests. J. Wildl. Manag. 1988, 52, 123–126. [Google Scholar] [CrossRef]
- Major, R.E.; Kendal, C.E. The contribution of artificial nest experiments to understanding avian reproductive success: A review of methods and conclusions. IBIS 2000, 138, 298–307. [Google Scholar] [CrossRef]
- Wilson, G.R.; Brittingham, M.C. How well do artificial nests estimate success of real nests? Condor 1998, 100, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Zanette, L. What do artificial nests tells us about nest predation? Biol. Conserv. 2002, 103, 323–329. [Google Scholar] [CrossRef]
- Krüger, H.; Väänänen, V.-M.; Holopainen, S.; Nummi, P. The new faces of nest predation in agricultural landscapes—A wildlife camera survey with artificial nests. Eur. J. Wildl. Res. 2018, 64, 76. [Google Scholar] [CrossRef] [Green Version]
- Einarsen, G.; Hausner, V.H.; Yoccoz, N.G.; Ims, R.A. Predation on artificial ground nests in birch forests fragmented by spruce plantations. Écoscience 2008, 15, 141–149. [Google Scholar] [CrossRef]
- Nour, N.; Matthysen, E.; Dhondt, A.A. Artificial nest predation and habitat fragmentation: Different trends in bird and mammal predators. Ecography 1993, 16, 111–116. [Google Scholar] [CrossRef]
- Volf, O.; Volfová, E.; Václavíková, E.; Čížková, Š.; Mariňáková, M.; Benediktová, V. Opatření na Podporu Tetřívka Obecného v Krušných horách—Measures Supporting Black Grouse in Krušné hory Mts; Ministry of the Environment of the Czech Republic: Prague, Czech Republic, 2019.
- Höglund, J.; Stöhr, S. A non-lekking population of Black Grouse Tetrao tetrix. J. Avian Biol. 1997, 28, 184–187. [Google Scholar] [CrossRef]
- Vacek, Z.; Cukor, J.; Linda, R.; Vacek, S.; Šimůnek, V.; Brichta, J.; Gallo, J.; Prokůpková, A. Bark stripping, the crucial factor affecting stem rot development and timber production of Norway spruce forests in Central Europe. For. Ecol. Manag. 2020, 474, 118360. [Google Scholar] [CrossRef]
- Soukupová, J.; Rock, B.N.; Albrechtová, J. Comparative study of two spruce species in a polluted mountainous region. New Phytol. 2001, 150, 133–145. [Google Scholar] [CrossRef]
- Špulák, O.; Dušek, D. Comparison of the impact of blue spruce and reed Calamagrostis villosa on forest soil chemical properties. J. For. Sci. 2009, 55, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Hering, S.; Irrgang, S. Conversion of substitute tree species stands and pure spruce stands in the Ore Mountains in saxony. J. For. Sci. 2005, 51, 519–525. [Google Scholar] [CrossRef] [Green Version]
- FMI. Forest Management Institute—Hunting Evidence Withouth National Parks and Military Areas; Forest Management Institute: Brandýs nad Labem, Czech Republic, 2019. [Google Scholar]
- Šťastný, K.; Bejček, V.; Hudec, K. Atlas Hnízdního Rozšíření práků v České Republice 1985–1989; H & H: Jinočany, Czech Republic, 1997. [Google Scholar]
- Koubek, P.; Banaš, M. Tetřevovití v Jeseníkách: Možnosti přežití. Tetraonid birds in the Jeseníky Mts: Chances for survival. In Tetřevovití—Tetraonidae na Přelomu Tisíciletí. Tetraonids—Tetraonidae at the Break of the Millennium; Málková, I., Ed.; KORŠACH: České Budějovice, Czech Republic, 2000; pp. 19–25. [Google Scholar]
- Selva, N.; Berezowska-Cnota, T.; Elguero-Claramunt, I. Unforeseen effects of supplementary feeding: Ungulate baiting sites as hotspots for ground-nest predation. PLoS ONE 2014, 9, e90740. [Google Scholar] [CrossRef] [PubMed]
- Oja, R.; Zilmer, K.; Valdmann, H. Spatiotemporal effects of supplementary feeding of wild boar (Sus scrofa) on artificial ground nest depredation. PLoS ONE 2015, 10, e135254. [Google Scholar] [CrossRef] [PubMed]
- Dahl, F.; Åhlén, P.A. Nest predation by raccoon dog Nyctereutes procyonoides in the archipelago of northern Sweden. Biol. Invasions 2019, 21, 743–755. [Google Scholar] [CrossRef] [Green Version]
- Storaas, T.; Wegge, P. Nesting Habitats and Nest Predation in Sympatric Populations of Capercaillie and Black Grouse. J. Wildl. Manag. 1987, 51, 167–172. [Google Scholar] [CrossRef]
- Willebrand, T. Breeding and age in female Black Grouse Tetrao tetrix. Ornis Scand. 1992, 23, 29–32. [Google Scholar] [CrossRef]
- Padyšáková, E.; Šálek, M.; Poledník, L.; Sedláček, F.; Albrecht, T. Removal of American mink increases the success of simulated nests in linear habitat. Wildl. Res. 2009, 36, 225. [Google Scholar] [CrossRef]
- Padyšáková, E.; Šálek, M.; Poledník, L.; Sedláček, F.; Albrecht, T. Predation on simulated duck nests in relation to nest density and landscape structure. Wildl. Res. 2010, 37, 597. [Google Scholar] [CrossRef]
- Bu, H.; Shen, X.; Li, S. Predation patterns on artificial nests of ground nesting pheasants in the montane forest, Southwest China. Acta Ornithol. 2019, 54, 35–43. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Core Team: Wien, Austria, 2020.
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Statistics for Biology and Health; Springer: New York, NY, USA, 2000; ISBN 978-1-4419-3161-0. [Google Scholar]
- Cox, D.R. Regression models and life tables (with Discussion). J. R. Stat. Soc. 1972, 34, 18–220. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete samples. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- TIBCO Software Inc. Statistica (Data Analysis Software System), Version 13. Available online: www.tibco.com (accessed on 5 January 2021).
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.10; Microcomputer Power: Ithaca, NY, USA, 2018. [Google Scholar]
- Batschelet, E. Circular Statistics in Biology; Academic Press: London, UK, 1981. [Google Scholar]
- Klaus, S.; Bergmann, H.-H.; Marti, C.; Müller, F.; Wiesner, J. Die Birkhühner; Die Neue Brehm-Bücherei: Lutherstadt Wittenberg, Germany, 1990. [Google Scholar]
- Valentine, E.C.; Apol, C.A.; Proppe, D.S. Predation on artificial avian nests is higher in forestsbordering small anthropogenic openings. IBIS 2019, 161, 662–673. [Google Scholar] [CrossRef]
- Šálek, M.; Kreisinger, J.; Sedláček, F.; Albrecht, T. Do prey densities determine preferences of mammalian predators for habitat edges in an agricultural landscape? Landsc. Urban Plan. 2010, 98, 86–91. [Google Scholar] [CrossRef]
- Červinka, J.; Šálek, M.; Pavluvčík, P.; Kreisinger, J. The fine-scale utilization of forest edges by mammalian mesopredators related to patch size and conservation issues in Central European farmland. Biodivers. Conserv. 2011, 20, 3459–3475. [Google Scholar] [CrossRef]
- Svobodová, J.; Kreisinger, J.; Šálek, M.; Koubová, M.; Albrecht, T. Testing mechanistic explanations for mammalian predator responses to habitat edges. Eur. J. Wildl. Res. 2011, 57, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Šálek, M.; Kreisinger, J.; Sedláček, F.; Albrecht, T. Corridor vs. hayfield matrix use by mammalian predators in an agricultural landscape. Agric. Ecosyst. Environ. 2009, 134, 8–13. [Google Scholar] [CrossRef]
- Donovan, T.M.; Jones, P.W.; Annand, E.M.; Thompson, F.R. Variation in local-scale edge effects: Mechanisms and landscape context. Ecology 1997, 78, 2064–2075. [Google Scholar] [CrossRef]
- Schneider, N.A.; Low, M.; Arlt, D.; Pärt, T. Contrast in edge vegetation structure modifies the predation risk of natural ground nests in an agricultural landscape. PLoS ONE 2012, 7, e31517. [Google Scholar] [CrossRef] [Green Version]
- Anglestam, P. Predation on Ground-Nesting Birds’ Nests in Relation to Predator Densities and Habitat Edge. Oikos 1986, 47, 365–373. [Google Scholar] [CrossRef]
- Ratti, J.T.; Reese, K.P. Preliminary Test of the Ecological Trap Hypothesis. J. Wildl. Manag. 1988, 52, 484–491. [Google Scholar] [CrossRef]
- Santos, T.; Tellería, J.L. Edge effects on nest predation in Mediterranean fragmented forests. Biol. Conserv. 1992, 60, 1–5. [Google Scholar] [CrossRef]
- Andren, H. Corvid Density and Nest Predation in Relation to Forest Fragmentation: A Landscape Perspective. Ecology 1992, 73, 794–804. [Google Scholar] [CrossRef]
- Ashby, K.R. Patterns of daily activity in mammals. Mamm. Rev. 1972, 1, 171–185. [Google Scholar] [CrossRef]
- Mason, L.R.; Smart, J.; Drewitt, A.L. Tracking day and night provides insights into the relative importance of different wader chick predators. IBIS 2018, 160, 71–88. [Google Scholar] [CrossRef]
- Jahren, T.; Storaas, T.; Fossland, M.P.; Hagen, B.R. Multi-fate nest predation in black grouse and capercaillie and effects of habitat—Preliminary results. In Proceedings of the International Grouse Symposium, Reykjavik, Iceland, 4–7 September 2015. [Google Scholar]
- Helldin, J.O. Population trends and harvest management of pine marten Martes martes in Scandinavia. Wildl. Biol. 2000, 6, 111–120. [Google Scholar] [CrossRef]
- Selås, V.; Vik, J.O. Possible impact of snow depth and ungulate carcasses on red fox (Vulpes vulpes) populations in Norway, 1897–1976. J. Zool. 2006, 269, 299–308. [Google Scholar] [CrossRef]
- Baines, D.; Aebischer, N.J.; Macleod, A. Increased mammalian predators and climate ch1ange predict declines in breeding success and density of Capercaillie Tetrao urogallus, an old stand specialist, in fragmented Scottish forests. Biodivers. Conserv. 2016, 25, 2171–2186. [Google Scholar] [CrossRef]
- Jahren, T.; Odden, M.; Linnell, J.D.C.; Panzacchi, M. The impact of human land use and landscape productivity on population dynamics of red fox in southeastern Norway. Mammal Res. 2020, 65, 503–516. [Google Scholar] [CrossRef]
- Giménez-Anaya, A.; Herrero, J.; Rosell, C.; Couto, S.; García-Serrano, A. Food habits of wild boar (Sus scrofa) in a Mediterranean coastal wetland. Wetlands 2008, 28, 197–203. [Google Scholar] [CrossRef]
- Ballari, S.A.; Barrios-García, M.N. A review of wild boar Sus scrofa diet and factors affecting food selection in native and introduced ranges. Mamm. Rev. 2014, 44, 124–134. [Google Scholar] [CrossRef]
- Bobek, B.; Furtek, J.; Bobek, J.; Merta, D.; Wojciuch-Ploskonka, M. Spatio-temporal characteristics of crop damage caused by wild boar in north-eastern Poland. Crop Prot. 2017, 93, 106–112. [Google Scholar] [CrossRef]
- Bragina, E.V.; Ives, A.R.; Pidgeon, A.M.; Balčiauskas, L.; Csányi, S.; Khoyetskyy, P.; Kysucká, K.; Lieskovsky, J.; Ozolins, J.; Randveer, T.; et al. Wildlife population changes across Eastern Europe after the collapse of socialism. Front. Ecol. Environ. 2018, 16, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Massei, G.; Coats, J.; Lambert, M.S.; Pietravalle, S.; Gill, R.; Cowan, D. Camera traps and activity signs to estimate wild boar density and derive abundance indices. Pest Manag. Sci. 2018, 74, 853–860. [Google Scholar] [CrossRef]
- Pejsak, Z.; Truszczyński, M.; Tarasiuk, K. Afrykański pomór świń u dzików. Med. Weter 2018, 74, 743–746. [Google Scholar]
- Rosvold, J.; Andersen, R. Zoologisk Rapport 2008-1 Jørgen Rosvold & Reidar Andersen Wild Boar in Norway—Is Climate a Limiting Factor? NTNU Vit-Enskapsmuseet Rapp. Zool. Ser. 2008, 1, 1–23. [Google Scholar]
- Oja, R.; Kaasik, A.; Valdmann, H. Winter severity or supplementary feeding—which matters more for wild boar? Acta Theriol. 2014, 59, 553–559. [Google Scholar] [CrossRef]
- Moynahan, B.; Lindberg, M.S.; Rotella, J.J.; Thomas, J.W. Factors Affecting Nest Survival of Greater Sage-Grouse in Northcentral Montana. J. Wildl. Manag. 2007, 71, 1773–1783. [Google Scholar] [CrossRef]
- Coates, P.S.; Connelly, J.W.; Delehanty, D.J. Predators of Greater Sage-Grouse nests identified by video. J. Field Ornithol. 2008, 79, 421–428. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Habitat | Predated Nests | Nonpredated Nests | Total |
|---|---|---|---|---|
| Jeseníky Mts. | Forest edge | 7 | 3 | 10 |
| Clearing | 1 | 5 | 6 | |
| Forest interior | 1 | 5 | 6 | |
| Ore Mts. | Forest edge | 4 | 5 | 9 |
| Clearing | 8 | 1 | 9 | |
| Forest interior | 7 | 3 | 10 | |
| Total | 28 | 22 | 50 |
| Independent Variable | Wald | df | p-Value |
|---|---|---|---|
| Study area | 5.99 | 1 | 0.014 |
| Habitat type | 0.69 | 2 | 0.709 |
| Study area*habitat type | 9.00 | 2 | 0.011 |
| Independent Variable | % of Explained Variability | Pseudo-F | p-Value |
|---|---|---|---|
| Habitat type | 21.7 | 3.2 | 0.004 |
| Category of predators’ behavior | 18.4 | 2.8 | 0.006 |
| Study area | 11.3 | 1.7 | 0.091 |
| Species | Predation | Predation Attempts | Pass-By Events | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Jeseníky | Ore Mts. | SUM | Jeseníky | Ore Mts. | SUM | Jeseníky | Ore Mts. | SUM | |
| Martes foina | 8 | 3 | 11 (39%) | — | 2 | 2 (10%) | — | 5 | 5 (16%) |
| Vulpes vulpes | 1 | 5 | 6 (22%) | — | 6 | 6 (27%) | 7 | 8 | 15 (47%) |
| Corvus corax | — | 7 | 7 (25%) | — | 4 | 4 (18%) | — | — | 0 |
| Garrulus glandarius | — | 2 | 2 (7%) | 5 | 1 | 6 (27%) | — | 2 | 2 (6%) |
| Sus scrofa | — | 2 | 2 (7%) | 1 | 3 | 4 (18%) | 2 | 5 | 7 (22%) |
| Martes martes | — | — | 0 | — | — | 0 | 2 | — | 2 (6%) |
| Meles meles | — | — | 0 | — | — | 0 | 1 | — | 1 (3%) |
| Independent Variable | df | AIC | % of Explained Variability | Chi | p-Value |
|---|---|---|---|---|---|
| Null model | 3 | 125.47 | |||
| Day/night | 4 | 87.98 | 33.1 | 39.49 | <0.001 |
| X hour * y hour | 6 | 71.06 | 50.5 | 60.41 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cukor, J.; Linda, R.; Andersen, O.; Eriksen, L.F.; Vacek, Z.; Riegert, J.; Šálek, M. Evaluation of Spatio-Temporal Patterns of Predation Risk to Forest Grouse Nests in the Central European Mountain Regions. Animals 2021, 11, 316. https://doi.org/10.3390/ani11020316
Cukor J, Linda R, Andersen O, Eriksen LF, Vacek Z, Riegert J, Šálek M. Evaluation of Spatio-Temporal Patterns of Predation Risk to Forest Grouse Nests in the Central European Mountain Regions. Animals. 2021; 11(2):316. https://doi.org/10.3390/ani11020316
Chicago/Turabian StyleCukor, Jan, Rostislav Linda, Oddgeir Andersen, Lasse Frost Eriksen, Zdeněk Vacek, Jan Riegert, and Martin Šálek. 2021. "Evaluation of Spatio-Temporal Patterns of Predation Risk to Forest Grouse Nests in the Central European Mountain Regions" Animals 11, no. 2: 316. https://doi.org/10.3390/ani11020316