
The First Insight into Polyhydroxyalkanoates Accumulation in Multi-Extremophilic Rubrobacter xylanophilus and Rubrobacter spartanus

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Cultivation and PHA Production
2.3. Analyses
2.4. Bioinformatics Analyses
3. Results
3.1. PHA Synthases in Genus Rubrobacter
3.2. PHA Accumulation by R. spartanus and R. xylanophilus
3.3. PHA Production on Various Substrates
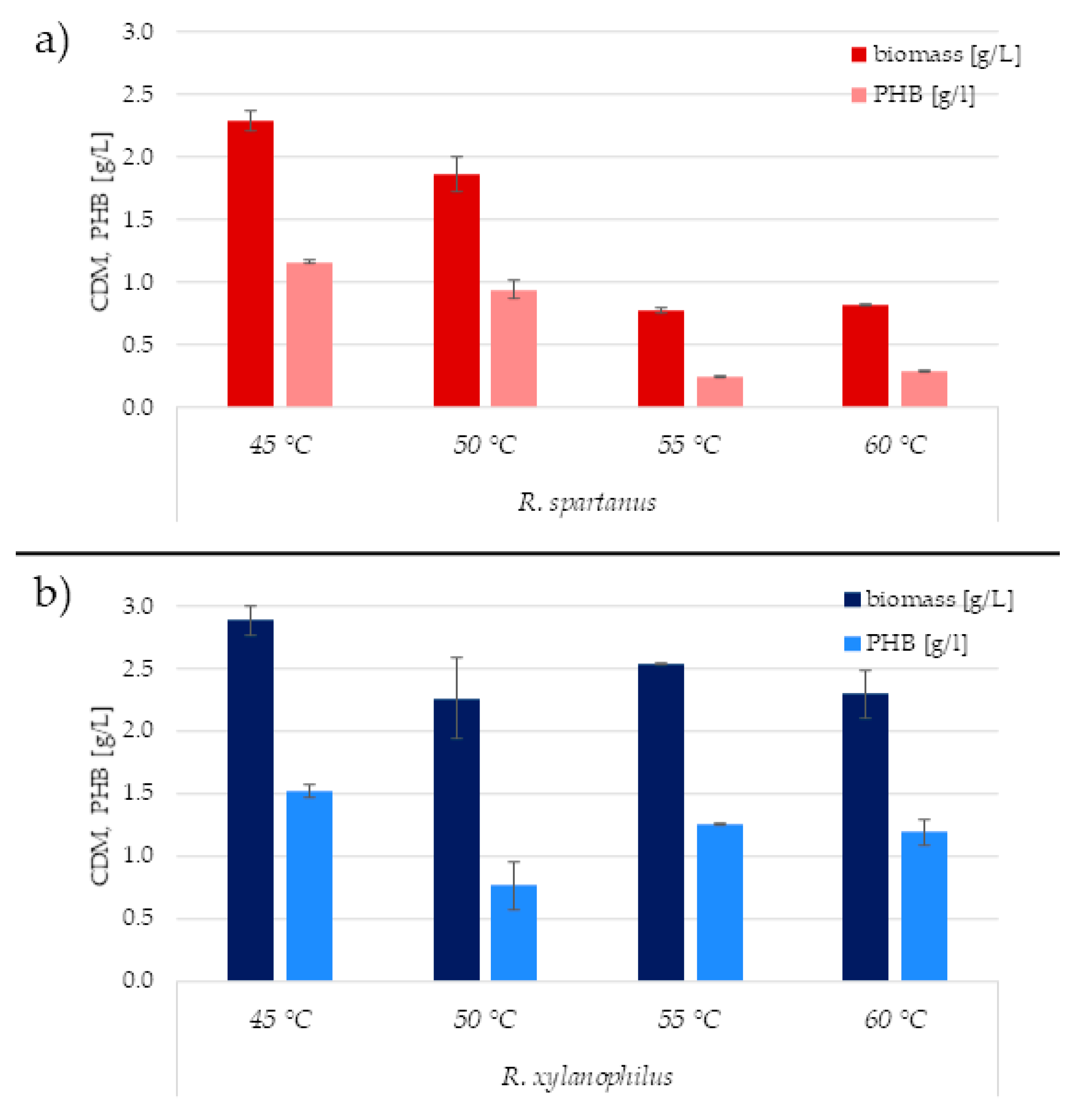
3.4. The Effect of Cultivation Temperature on PHA Accumulation
3.5. Production of PHA Copolymers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Castro, J.F.; Nouioui, I.; Asenjo, J.A.; Andrews, B.; Bull, A.T.; Goodfellow, M. New genus-specific primers for PCR identification of Rubrobacter strains. Antonie van Leeuwenhoek 2019, 112, 1863–1874. [Google Scholar] [CrossRef] [Green Version]
- Freed, S.; Ramaley, R.F.; Kyndt, J.A. Whole-Genome Sequence of the Novel Rubrobacter taiwanensis Strain Yellowstone, Isolated from Yellowstone National Park. Microbiol. Resour. Announc. 2019, 8, e00287-19. [Google Scholar] [CrossRef] [Green Version]
- Norman, J.S.; King, G.M.; Friesen, M.L. Rubrobacter spartanus sp. nov., a moderately thermophilic oligotrophic bacterium isolated from volcanic soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 3597–3602. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.-W.; Li, C.; He, Y.-Q.; Cui, L.-Q.; Long, L.-J.; Tian, X.-P. Rubrobacter tropicus sp. nov. and Rubrobacter marinus sp. nov., isolated from deep-sea sediment of the South China Sea. Int. J. Syst. Evol. Microbiol. 2020, 70, 5576–5585. [Google Scholar] [CrossRef] [PubMed]
- Terato, H.; Suzuki, K.; Nishioka, N.; Okamoto, A.; Shimazaki-Tokuyama, Y.; Inoue, Y.; Saito, T. Characterization and ra-dio-resistant function of manganese superoxide dismutase of Rubrobacter radiotolerans. J. Radiat. Res. 2011, 52, 735–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asgarani, E.; Terato, H.; Asagoshi, K.; Shahmohammadi, H.R.; Ohyama, Y.; Saito, T.; Yamamoto, O.; Ide, H. Purification and characterization of a novel DNA repair enzyme from the extremely radioresistant bacterium Rubrobacter radiotolerans. J. Radiat. Res. 2000, 41, 19–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Furukawara, M.; Omae, K.; Tadokoro, N.; Saito, Y.; Abe, K.; Kera, Y. A Highly Stable d-Amino Acid Oxidase of the Thermophilic Bacterium Rubrobacter xylanophilus. Appl. Environ. Microbiol. 2014, 80, 7219–7229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovács, K.; Bánóczi, G.; Varga, A.; Szabó, I.; Holczinger, A.; Hornyánszky, G.; Zagyva, I.; Paizs, C.; Vértessy, B.G.; Poppe, L. Expression and Properties of the Highly Alkalophilic Phenylalanine Ammonia-Lyase of Thermophilic Rubrobacter xylanophilus. PLoS ONE 2014, 9, e85943. [Google Scholar] [CrossRef] [Green Version]
- Gabani, P.; Singh, O.V. Radiation-resistant extremophiles and their potential in bio-technology and therapeutics. Appl. Microbiol. Biot. 2013, 97, 993–1004. [Google Scholar] [CrossRef]
- Webb, K.M.; DiRuggiero, J. Role of Mn2+and Compatible Solutes in the Radiation Resistance of Thermophilic Bacteria and Archaea. Archaea 2012, 2012, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.; Newton, G.L.; Fahey, R.C.; Rawat, M. Unusual production of glutathione in Actinobacteria. Arch. Microbiol. 2009, 191, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.-Q.; Jiang, X.-R. Next generation industrial biotechnology based on extremophilic bacteria. Curr. Opin. Biotechnol. 2018, 50, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Koller, M.; Mukherjee, A. Polyhydroxyalkanoates—Linking Properties, Applications and End-of-life Options. Chem. Biochem. Eng. Q. 2020, 34, 115–129. [Google Scholar] [CrossRef]
- Sedlacek, P.; Slaninova, E.; Koller, M.; Nebesarova, J.; Marova, I.; Krzyzanek, V.; Obruca, S. PHA granules help bacterial cells to preserve cell integrity when exposed to sudden osmotic imbalances. N. Biotechnol. 2019, 49, 129–136. [Google Scholar] [CrossRef]
- Alves, L.P.S.; Santana-Filho, A.P.; Sassaki, G.L.; Pedrosa, F.D.O.; de Souza, E.M.; Chubatsu, L.S.; Müller-Santos, M. 3-Hydroxybutyrate Derived from Poly-3-Hydroxybutyrate Mobilization Alleviates Protein Aggregation in Heat-Stressed Herbaspirillum seropedicae SmR. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Obruca, S.; Sedlacek, P.; Krzyzanek, V.; Mravec, F.; Hrubanova, K.; Samek, O.; Kucera, D.; Benesova, P.; Marova, I. Accu-mulation of poly(3-hydroxybutyrate) helps bacterial cells to survive freezing. PLoS ONE 2016, 11, e0157778. [Google Scholar] [CrossRef] [Green Version]
- Slaninova, E.; Sedlacek, P.; Mravec, F.; Mullerova, L.; Samek, O.; Koller, M.; Hesko, O.; Kucera, D.; Marova, I.; Obruca, S. Light scattering on PHA granules protects bacterial cells against the harmful effects of UV radiation. Appl. Microbiol. Biotechnol. 2018, 102, 1923–1931. [Google Scholar] [CrossRef]
- Batista, M.B.; Teixeira, C.S.; Sfeir, M.Z.T.; Alves, L.P.S.; Valdameri, G.; Pedrosa, F.D.O.; Sassaki, G.L.; Steffens, M.B.R.; De Souza, E.M.; Dixon, R.; et al. PHB Biosynthesis Counteracts Redox Stress in Herbaspirillum seropedicae. Front. Microbiol. 2018, 9, 472. [Google Scholar] [CrossRef]
- Müller-Santos, M.; Koskimäki, J.J.; Alves, L.P.S.; de Souza, E.M.; Jendrossek, D.; Pirttilä, A.M. The protective role of PHB and its degradation products against stress situations in bacteria. FEMS Microbiol. Rev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Obruca, S.; Sedlacek, P.; Koller, M. The underexplored role of diverse stress factors in microbial biopolymer synthesis. Bioresour. Technol. 2021, 326, 124767. [Google Scholar] [CrossRef]
- Carreto, L.; Moore, E.; Nobre, M.F.; Wait, R.; Riley, P.W.; Sharp, R.J.; Da Costa, M.S. Rubrobacter xylanophilus sp. nov., a New Thermophilic Species Isolated from a Thermally Polluted Effluent. Int. J. Syst. Bacteriol. 1996, 46, 460–465. [Google Scholar] [CrossRef]
- Obruca, S.; Benesova, P.; Oborna, J.; Marova, I. Application of protease-hydrolyzed whey as a complex nitrogen source to increase poly(3-hydroxybutyrate) production from oils by Cupriavidus necator. Biotechnol. Lett. 2014, 36, 775–781. [Google Scholar] [CrossRef]
- Obruca, S.; Sedlacek, P.; Mravec, F.; Krzyzanek, V.; Nebesarova, J.; Samek, O.; Kucera, D.; Benesova, P.; Hrubanova, K.; Milerova, M.; et al. The presence of PHB granules in cytoplasm protects non-halophilic bacterial cells against the harmful impact of hypertonic environments. New Biotechnol. 2017, 39, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2012, 41, D36–D42. [Google Scholar] [CrossRef] [Green Version]
- Bateman, A.; Martin, M.J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49. [Google Scholar] [CrossRef]
- Taboada, B.; Estrada, K.; Ciria, R.; Merino, E. Operon-mapper: A web server for precise operon identification in bacterial and archaeal genomes. Bioinform. 2018, 34, 4118–4120. [Google Scholar] [CrossRef] [PubMed]
- Knoll, M.; Hamm, T.M.; Wagner, F.; Martinez, V.; Pleiss, J. The PHA Depolymerase Engineering Database: A systematic analysis tool for the diverse family of polyhydroxyalkanoate (PHA) depolymerases. BMC Bioinform. 2009, 10, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stothard, P. The Sequence Manipulation Suite: JavaScript Programs for Analyzing and Formatting Protein and DNA Se-quences. BioTechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mravec, F.; Obruca, S.; Krzyzánek, V.; Sedlacek, P.; Hrubanova, K.; Samek, O.; Kucera, D.; Benesova, P.; Nebesářová, J. Accumulation of PHA granules in Cupriavidus necator as seen by confocal fluorescence microscopy. FEMS Microbiol. Lett. 2016, 363, 363. [Google Scholar] [CrossRef] [Green Version]
- Matias, F.; Bonatto, D.; Padilla, G.; Rodrigues, M.F.D.A.; Henriques, J.A.P. Polyhydroxyalkanoates production by actinobacteria isolated from soil. Can. J. Microbiol. 2009, 55, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Thakur, V.; Ambika; Kumar, S.; Singh, D. Bioplastic reservoir of diverse bacterial communities revealed along altitude gradient of Pangi-Chamba trans-Himalayan region. FEMS Microbiol. Lett. 2018, 365, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trakunjae, C.; Boondaeng, A.; Apiwatanapiwat, W.; Kosugi, A.; Arai, T.; Sudesh, K.; Vaithanomsat, P. Enhanced polyhy-droxybutyrate (PHB) production by newly isolated rare actinomycetes Rhodococcus sp. strain BSRT1-1 using response surface methodology. Sci. Rep. UK 2021, 11, 1–14. [Google Scholar]
- Altaee, N.; El-Hiti, G.A.; Fahdil, A.; Sudesh, K.; Yousif, E. Screening and Evaluation of Poly(3-hydroxybutyrate) with Rho-dococcus equi Using Different Carbon Sources. Arab. J. Sci. Eng. 2017, 42, 2371–2379. [Google Scholar] [CrossRef]
- Jia, K.; Cao, R.; Hua, D.H.; Li, P. Study of Class i and Class III Polyhydroxyalkanoate (PHA) Synthases with Substrates Con-taining a Modified Side Chain. Biomacromolecules 2016, 17, 1477–1485. [Google Scholar] [CrossRef] [Green Version]
- Tsuge, T.; Hyakutake, M.; Mizuno, K. Class IV polyhydroxyalkanoate (PHA) synthases and PHA-producing Bacillus. Appl. Microbiol. Biotechnol. 2015, 99, 6231–6240. [Google Scholar] [CrossRef]
- Obruca, S.; Sedlacek, P.; Slaninova, E.; Fritz, I.; Daffert, C.; Meixner, K.; Sedrlova, Z.; Koller, M. Novel unexpected functions of PHA granules. Appl. Microbiol. Biotechnol. 2020, 104, 4795–4810. [Google Scholar] [CrossRef]
- Obruca, S.; Sedlacek, P.; Koller, M.; Kucera, D.; Pernicova, I. Involvement of poly-hydroxyalkanoates in stress resistance of microbial cells: Biotechnological consequences and applications. Biotechnol. Adv. 2018, 36, 856–870. [Google Scholar] [CrossRef]
- Soto, G.; Setten, L.; Lisi, C.; Maurelis, C.; Mozzicafreddo, M.; Cuccioloni, M.; Angeletti, M.; Ayub, N.D. Hydroxybutyrate prevents protein aggregation in the halotolerant bacterium Pseudomonas sp. CT13 under abiotic stress. Extremophiles 2012, 16, 455–462. [Google Scholar] [CrossRef]
- Obruca, S.; Sedlacek, P.; Mravec, F.; Samek, O.; Marova, I. Evaluation of 3-hydroxybutyrate as an enzyme-protective agent against heating and oxidative damage and its potential role in stress response of poly(3-hydroxybutyrate) accumulating cells. Appl. Microbiol. Biotechnol. 2015, 100, 1365–1376. [Google Scholar] [CrossRef]
- De Almeida, A.; Catone, M.V.; Rhodius, V.A.; Gross, C.A.; Pettinari, M.J. Unexpected Stress-Reducing Effect of PhaP, a Poly(3-Hydroxybutyrate) Granule-Associated Protein, in Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 6622–6629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brojanigo, S.; Parro, E.; Cazzorla, T.; Favaro, L.; Basaglia, M.; Casella, S. Conversion of Starchy Waste Streams into Polyhy-droxyalkanoates Using Cupriavidus necator DSM. Polymers 2020, 12, 1496. [Google Scholar] [CrossRef] [PubMed]
- Raposo, R.S.; de Almeida, M.C.M.; Oliveira, M.D.C.M.D.; da Fonseca, M.M.; Cesário, M.T. A Burkholderia sacchari cell factory: Production of poly-3-hydroxybutyrate, xylitol and xylonic acid from xylose-rich sugar mixtures. New Biotechnol. 2017, 34, 12–22. [Google Scholar] [CrossRef]
- Pernicova, I.; Kucera, D.; Nebesarova, J.; Kalina, M.; Novackova, I.; Koller, M.; Obruca, S. Production of polyhydroxyalka-noates on waste frying oil employing selected Halomonas strains. Bioresource Technol. 2019, 292, 122028. [Google Scholar] [CrossRef]
- Singh, A.K.; Srivastava, J.K.; Chandel, A.K.; Sharma, L.; Mallick, N.; Singh, S.P. Biomedical applications of microbially engi-neered polyhydroxyalkanoates: An insight into recent advances, bottlenecks, and solutions. Appl. Microbiol. Biotechnol. 2019, 103, 2007–2032. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.T.; Song, H.; Raschbauer, M.; Emerstorfer, F.; Omann, M.; Stelzer, F.; Neureiter, M. Utilization of desugarized sugar beet molasses for the production of poly(3-hydroxybutyrate) by halophilic Bacillus megaterium uyuni S29. Process Biochem. 2019, 86, 9–15. [Google Scholar] [CrossRef]
- Sadykov, M.R.; Ahn, J.-S.; Widhelm, T.J.; Eckrich, V.M.; Endres, J.L.; Driks, A.; Rutkowski, G.E.; Wingerd, K.L.; Bayles, K.W. Poly(3-hydroxybutyrate) fuels the tricarboxylic acid cycle and de novo lipid biosynthesis during Bacillus anthracis sporulation. Mol. Microbiol. 2017, 104, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Kourilova, X.; Pernicova, I.; Sedlar, K.; Musilova, J.; Sedlacek, P.; Kalina, M.; Koller, M.; Obruca, S. Production of polyhy-droxyalkanoates (PHA) by a thermophilic strain of Schlegelella thermodepolymerans from xylose rich substrates. Bioresource Technol. 2020, 315, 123885. [Google Scholar] [CrossRef]
- Budde, C.F.; Riedel, S.L.; Willis, L.B.; Rha, C.; Sinskey, A.J. Production of Poly(3-Hydroxybutyrate-co-3-Hydroxyhexanoate) from Plant Oil by Engineered Ralstonia eutropha Strains. Appl. Environ. Microbiol. 2011, 77, 2847–2854. [Google Scholar] [CrossRef] [Green Version]



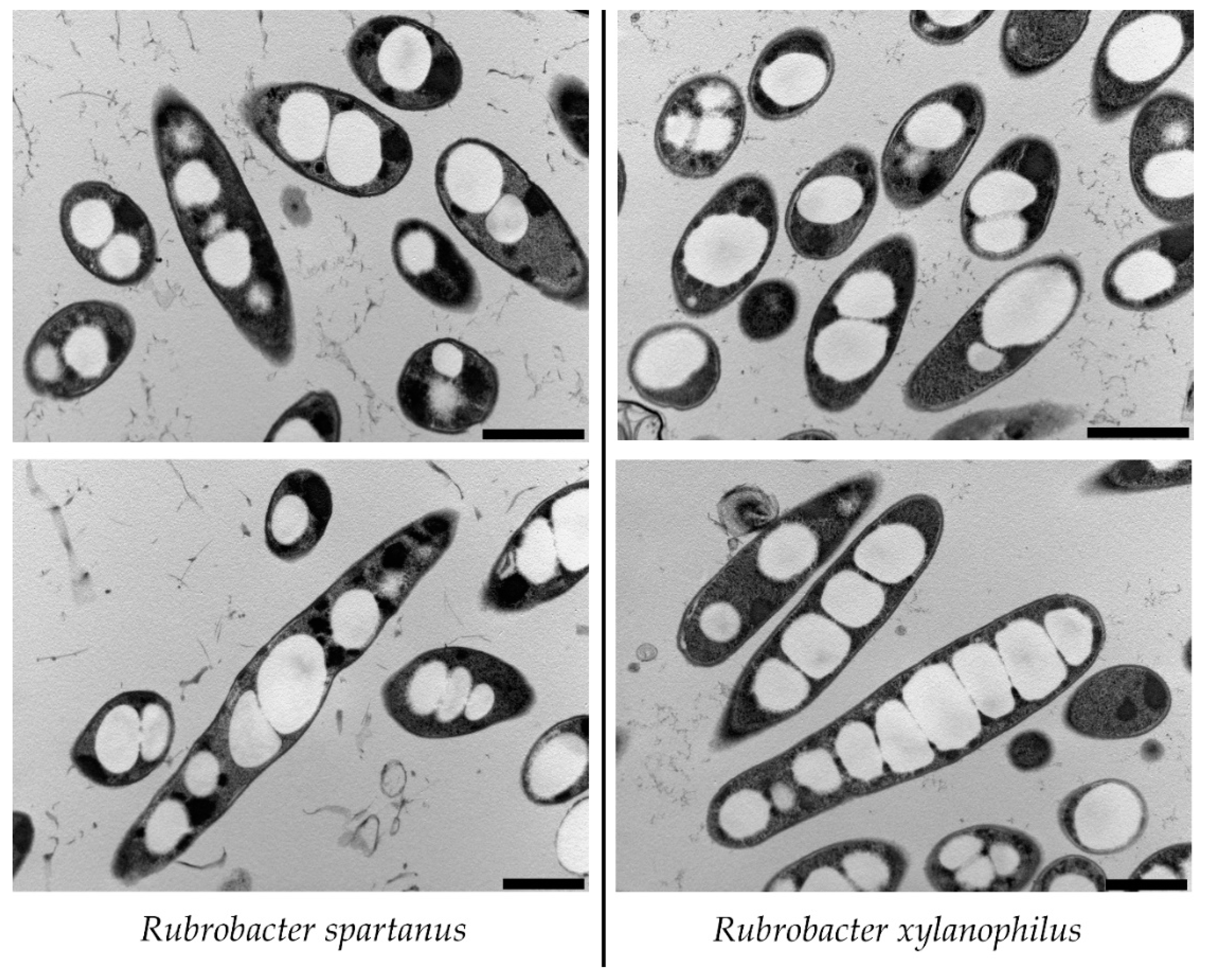{kind=link}
{kind=link}
| Organism | Accession No. | Putative PHA Class | Gene | Locus |
|---|---|---|---|---|
| Rubrobacter sp. SCSIO 52909 | CP045119.1 | Class I | phaC | GBA63_04560 |
| Class III | phaC | GBA63_04605 | ||
| phaE | GBA63_04610 | |||
| Rubrobacter sp. SCSIO 52915 | CP045121.1 | Class I | phaC | GBA65_16640 |
| Class III | phaC | GBA65_16620 | ||
| phaE | GBA65_16615 | |||
| R. xylanophilus AA3-22 | AP019791.1 | Class I | phaC | RxyAA322_15910 |
| Class III | phaC | RxyAA322_15900 | ||
| phaE | RxyAA322_15890 | |||
| R. xylanophilus DSM 9941 | CP000386.1 | Class I | phaC | Rxyl_2386 |
| Class III | phaC | Rxyl_2385 | ||
| phaE | Rxyl_2384 | |||
| R. radiotolerans RSPS-4 | CP007514.1 | Class I | phaC | RradSPS_2088 |
| Class I | phaC | RradSPS_2601 | ||
| Class III | phaC | RradSPS_2085 | ||
| phaE | RradSPS_2084 | |||
| R. indioceani SCSIO 08198 | NZ_CP031115.1 | Class I | phaC | DU509_RS10855 |
| Class I | phaC | DU509_RS11050 | ||
| Class III | phaC | DU509_RS10845 | ||
| phaE | DU509_RS15705 |
| Organism | Accession |
|---|---|
| Rubrobacter sp. MGR_bin435 | MBA3953681.1 |
| Rubrobacter sp. Co-bin5 | MBD0252984.1 |
| Rubrobacter radiotolerans RSPS-4 | AHY46637.1 |
| Rubrobacter radiotolerans DSM 5868 | SMC05048.1 |
| Rubrobacter sp. SCSIO 52909 | QIN83803.1 |
| Rubrobacter sp. MGR_bin143 | MBA2690884.1 |
| Rubrobacter sp. MGR_bin332 | MBA3609848.1 |
| Rubrobacter indicoceani SCSIO 08198 | WP_119067871.1 |
| Rubrobacter sp. MGR_bin27 | MBA3471765.1 |
| Rubrobacter sp. SCSIO 52915 | QIN79014.1 |
| Rubrobacter sp. MGR_bin77 | MBA3388321.1 |
| Rubrobacter sp. MGR_bin253 | MBA2376553.1 |
| Rubrobacter sp. MGR_bin199 | MBA2534341.1 |
| Rubrobacter sp. MGR_bin231 | MBA2443425.1 |
| Rubrobacter sp. MGR_bin80 | MBA3425602.1 |
| Substrate | CDM 1 [g/L] | PHB [% of CDM] | P3HB [g/L] | |
|---|---|---|---|---|
| R1 | arabinose | 0.47 ± 0.16 | 3.00 ± 0.04 | 0.01 ± 0.01 |
| fructose | 0.82 ± 0.11 | 4.21 ± 0.16 | 0.03 ± 0.01 | |
| galactose | 0.73 ± 0.10 | 7.28 ± 0.20 | 0.05 ± 0.01 | |
| glucose | 3.35 ± 0.03 | 51.53 ± 0.30 | 1.73 ± 0.07 | |
| glycerol | 0.92 ± 0.06 | 38.02 ± 0.13 | 0.35 ± 0.01 | |
| lactose | 2.38 ± 0.06 | 52.28 ± 0.14 | 1.24 ± 0.01 | |
| mannose | 0.96 ± 0.04 | 4.56 ± 0.19 | 0.04 ± 0.01 | |
| WFO | 0.49 ± 0.17 | 2.34 ± 0.05 | 0.01 ± 0.01 | |
| succrose | 2.10 ± 0.04 | 45.28 ± 8.41 | 0.95 ± 0.18 | |
| xylose | 1.46 ± 0.27 | 4.39 ± 0.23 | 0.06 ± 0.01 | |
| Substrate | CDM [g/L] | PHB [% of CDM] | P3HB [g/L] | |
| R2 | arabinose | n. d. 2 | n. d. | n. d. |
| fructose | n. d. | n. d. | n. d. | |
| galactose | n. d. | n. d. | n. d. | |
| glucose | 2.70 ± 0.04 | 38.59 ± 2.84 | 1.04 ± 0.08 | |
| glycerol | 2.27 ± 0.07 | 49.65 ± 0.18 | 1.13 ± 0.01 | |
| lactose | n. d. | n. d. | n. d. | |
| mannose | n. d. | n. d. | n. d. | |
| WFO | 2.50 ± 0.01 | 1.34 ± 0.05 | 0.03 ± 0.00 | |
| succrose | n. d. | n. d. | n. d. | |
| xylose | n. d. | n. d. | n. d. |
| n-Propanol | Valeric Acid | Levulinic Acid | n-Amyl Alcohol | Propionate | ||
|---|---|---|---|---|---|---|
| R1 | CDM [g/L] | 2.37 ± 0.01 | 0.85 ± 0.01 | n.d.* | 2.15 ± 0.08 | 0.58 ± 0.01 |
| PHA [% of CDM] | 31.38 ± 0.06 | 3.57 ± 0.03 | n.d. | 30.73 ± 1.31 | 4.63 ± 0.04 | |
| PHA [g/L] | 0.74 ± 0.01 | 0.03 ± 0.01 | n.d. | 0.66 ± 0.03 | 0.03 ± 0.01 | |
| 3HV [mol. %] | 4.27 | 8.42 | n.d. | 2.78 | 0.00 | |
| 3HB [mol. %] | 95.73 | 91.58 | n.d. | 97.22 | 100.00 | |
| R2 | CDM [g/L] | 2.23 ± 0.01 | n.d. | 0.45 ± 0.01 | 1.29 ± 0.04 | 2.08 ± 0.02 |
| PHA [% of CDM] | 43.86 ± 1.42 | n.d. | 13.26 ± 0.03 | 32.73 ± 0.73 | 20.62 ± 0.43 | |
| PHA [g/L] | 0.98 ± 0.03 | n.d. | 0.06 ± 0.01 | 0.42 ± 0.02 | 0.43 ± 0.01 | |
| 3HV [mol. %] | 0.77 | n.d. | 0.00 | 99.48 | 87.04 | |
| 3HB [mol. %] | 99.23 | n.d. | 100.00 | 0.52 | 12.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouřilová, X.; Schwarzerová, J.; Pernicová, I.; Sedlář, K.; Mrázová, K.; Krzyžánek, V.; Nebesářová, J.; Obruča, S. The First Insight into Polyhydroxyalkanoates Accumulation in Multi-Extremophilic Rubrobacter xylanophilus and Rubrobacter spartanus. Microorganisms 2021, 9, 909. https://doi.org/10.3390/microorganisms9050909
Kouřilová X, Schwarzerová J, Pernicová I, Sedlář K, Mrázová K, Krzyžánek V, Nebesářová J, Obruča S. The First Insight into Polyhydroxyalkanoates Accumulation in Multi-Extremophilic Rubrobacter xylanophilus and Rubrobacter spartanus. Microorganisms. 2021; 9(5):909. https://doi.org/10.3390/microorganisms9050909
Chicago/Turabian StyleKouřilová, Xenie, Jana Schwarzerová, Iva Pernicová, Karel Sedlář, Kateřina Mrázová, Vladislav Krzyžánek, Jana Nebesářová, and Stanislav Obruča. 2021. "The First Insight into Polyhydroxyalkanoates Accumulation in Multi-Extremophilic Rubrobacter xylanophilus and Rubrobacter spartanus" Microorganisms 9, no. 5: 909. https://doi.org/10.3390/microorganisms9050909