3.1. Screening of Temperature Regime for Lipid Production in Metschnikowia Strains
Based on an experimental study, it was found that one of the factors that significantly affect not only biomass yield but also lipid production is temperature. Increased lipid accumulation has been shown to be positively affected by lower temperatures compared to room temperature. This fact also seems to be advantageous in terms of sterility, as lower temperatures are not optimal for the growth of most microorganisms, especially bacteria [
2].
Yeasts cultivated at different temperatures than the optimum one are exposed to temperature stress. Temperature stress causes various changes in yeast metabolism, which affects their growth and production properties [
9]. The influence of temperature stress due to low temperature (CT −15 °C) compared to the laboratory (LT −25 °C) on
Metschnikowia yeast was monitored on optimal glucose media with decreased C/N ratio (you can see at
Section 2.3).
Cultivation of yeasts that had been kept at laboratory temperature for the whole time resulted in higher biomass production, compared to cultivation in environments with low temperature (
Table 3). The low values of the produced biomass were generally lower in the
Metschnikowia yeast, but on the other hand, the change in temperature did not significantly affect them. Slowed metabolism and division of yeast cells by reduced temperature resulted in increased lipid production in all studied
Metschnikowia strains. Yeasts (
Table 3) show the same trend as in the previous work [
9].
3.1.1. Lipid Overproduction in Metschnikowia Strains at Low Temperature
Changing the temperature regime of the cultivation towards lower temperatures had a positive effect on lipid production, as higher yields of lipids were achieved in all yeasts cultivated at low temperatures. As shown in
Table 4, higher biomass production was most often achieved by cultivation at room temperature. Overall, most lipids were contained in the cells of the yeast
M. andauensis 1241 (7.9%) at reduced temperature, which also made it the highest producer of lipids compared to other yeasts.
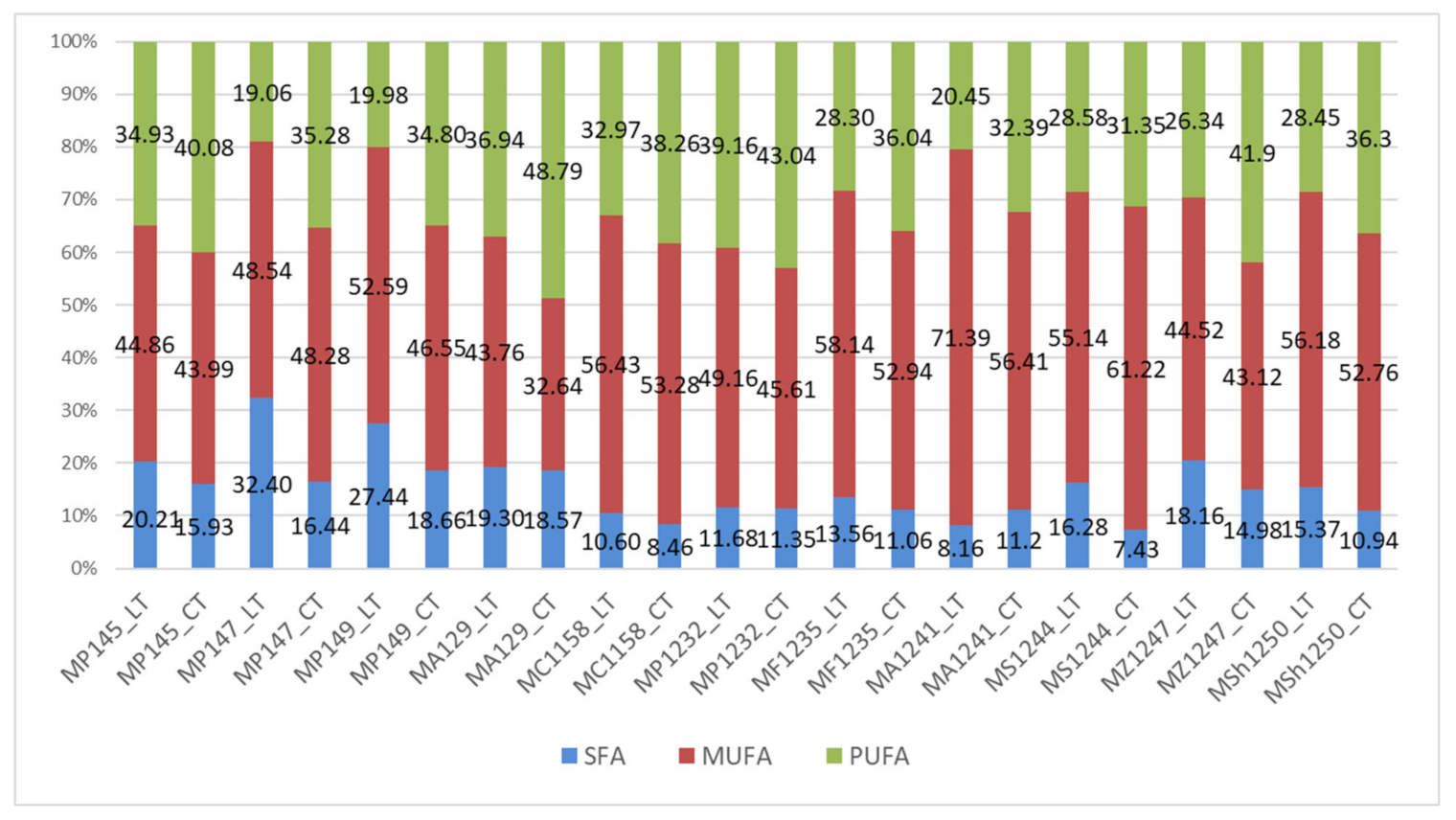
The fatty acid profile was determined on laboratory (LT) and reduced temperature (CT) control media where glucose was the main carbon source.
The influence of low temperature on the composition of fatty acids in microbial lipids was evident in all the yeasts studied. Results described in
Figure 1 show a stable trend of increased production of SFA in laboratory temperature between the majority of studied
Metschnikowia strains. This phenomenon is most markedly observed in the strains
Metschnikowia p. 147,
Metschnikowia s. 1244, and
Metschnikowia p. 149, where the highest production of 32.40% and 27.44% was achieved. In the case of other studied strains, the difference in SFA production depending on the cultivation temperature was lower and ranged from 0.5 to 4.0%. The application of reduced temperature had a positive effect on the increased accumulation of unsaturated fatty acids on the expense of SFA and MUFA. The results show that the increase in UFA in most strains is primarily in favor of the formation of polyunsaturated fatty acids. The highest PUFA content, more than 40%, was measured in
Metschnikowia p. 145,
Metschnikowia a. 129,
Metschnikowia p. 1232, and
Metschnikowia p. 1247.
Oleic acid had the highest concentration in the yeast cells. In some strains, it accounted for more than half of the fatty acid profile. However, a decrease in oleic acid content of almost 17% was observed in the cultivation at reduced temperature (CT). On the other hand, the amount of linoleic acid increased. Thus, it can be concluded that lower temperature conditions are likely to be more suitable to produce polyunsaturated fatty acids, mainly linoleic acid. A-linolenic acid was also formed in a small proportion. The low temperature has been shown to be a supporting factor for the increase in polyunsaturated fatty acids in almost all the cultivations studied.
Table 4 shows the fatty acid profile of
M. andauensis 1241, an dthe majority of the lipids produced in both temperature regimes had oleic acid. However, when using a low temperature, we can observe a significant decrease in oleic acid and increase in linoleic acid as compared to the culture at room temperature.
The low temperature even enhanced the production of other significant unsaturated FA that were not produced at room temperature at all, such as γ-linolenic acid, eicosadienic acid, arachidonic acid, or eiosapentaenoic acid in all the yeasts studied.
The effect of temperature on lipid production demonstrated in this work is consistent with the results of the study, where the stimulating effect of reduced temperature on increased lipid production was also demonstrated [
2,
9].
In addition, it is necessary to highlight the very significant effect of reduced temperature on the composition of FA of microbial lipids. These changes in the fatty acid composition are probably closely related to the effect of low temperature on the membranes, which reduces their fluidity.
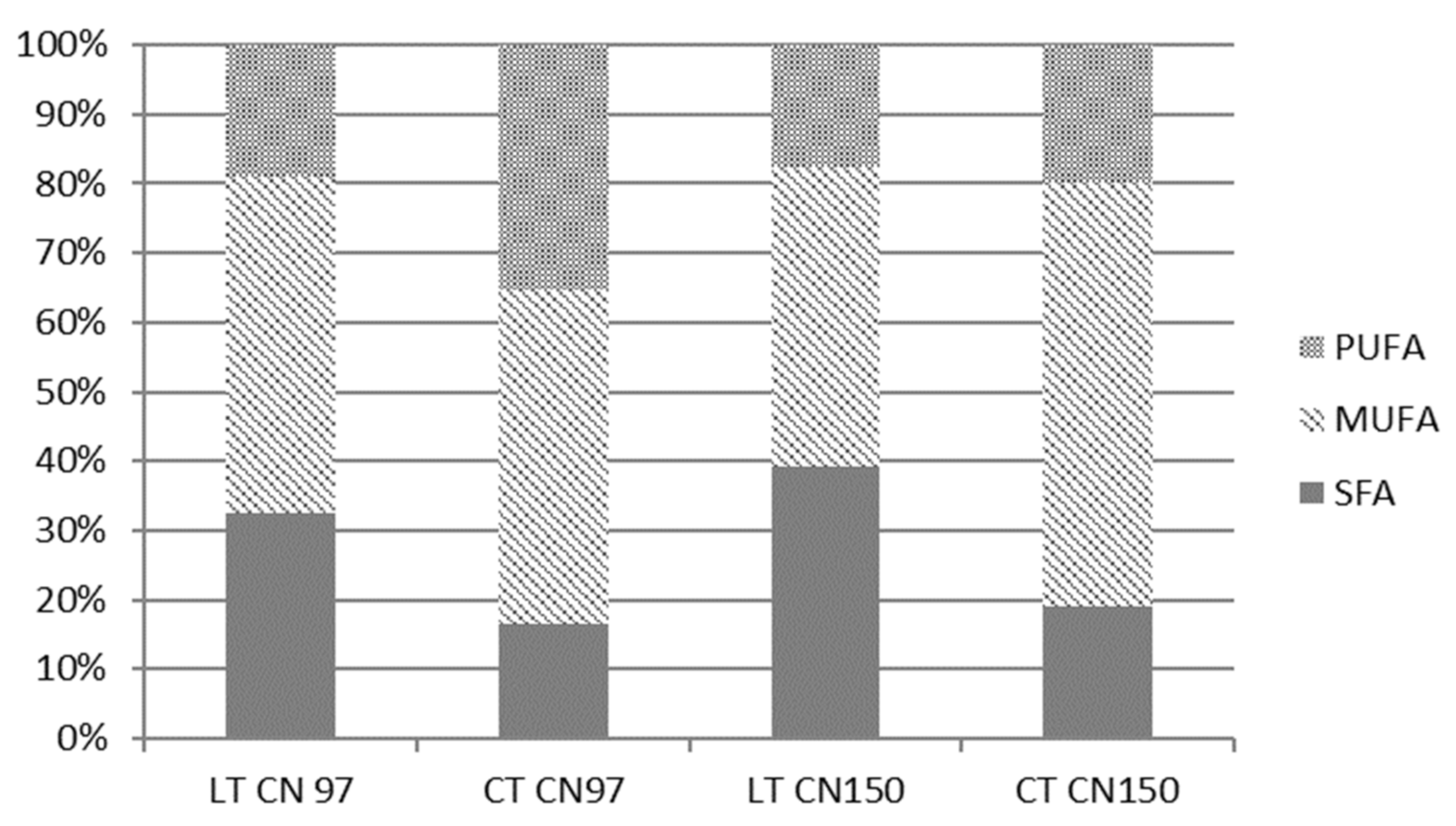
The effect of the temperature regime on lipid profile was therefore screened in addition to the optimal glucose medium with low C/N ratio also in media with increased C/N ratio, namely 97 and 150 in selected yeast strains.
For better clarity for the reader, the results are not shown except for in
Figure 2. In general, however, low temperature (CT) and an increased C/N ratio caused increased production of MUFA and PUFA in
Metschnikowia yeasts.
Figure 2 demonstrates the changes in saturation and unsaturation of fatty acids (FA) in the yeast
Metschnikowia pulcherrima 147. Both in the previous section (3.1.1) and this section, the cold temperature had an effect on the increase in PUFAs, in particular linoleic acid, compared to laboratory temperature, in cultures at all C/N ratios. Additionally, the cold temperature had the effect of reducing SFAs compared to laboratory temperature at all C/N ratios.
The effects of temperature and C/N ratio on the production properties of the studied Metschnikowia yeast proved to be significant and was demonstrated in all studied yeast cultures.
The results obtained show that with increasing C/N ratio, the amount of MUFA also increases at cold temperature. If we want to biotechnologically focus on higher PUFA production in yeasts of the genus Metschnikowia, we should choose a cold culture temperature and a C/N ratio of 97 or close to 100.
3.1.2. Combined Effect of Low Temperature and C/N ratio on Lipid Production
Lipid production in oleaginous yeast is influenced by several factors such as carbon and nitrogen source, C/N ratio, medium aeration, temperature, and pH. Different studies have therefore sought to optimize culture conditions to achieve the highest possible lipid production in yeast cells [
2]. The biomass production and increased lipid accumulation in oleaginous yeasts are opposing phenomena that are controlled by a number of external and internal cultivation factors such as aeration, sufficient nutrients, and temperature.
Based on the results of increased lipid accumulation at a reduced temperature [
9], the experiment continued by examining the effect of the C/N ratio on lipid production. Lipid accumulation is usually optimal at a C/N ratio higher than 65 and close to 100 [
2]. The results of combined low temperature and different C/N ratios on lipid accumulation in multiple yeast species are shown in
Table 5.
The production of biomass (g/L) and percent content of fatty acids per biomass weight (%) has been monitored in each cultivation for the chosen yeast strains according to the used type of carbon source (glucose) and chosen C/N ratio.
Metschnikowia yeast strains were cultivated in media where glucose was used as the main carbon source. Different contents of peptone and yeast extract were used in each medium as a source of nitrogen depending on the selected C/N ratios of 24, 97, and 150.
The results clearly show that as the C/N ratio increases, the amount of biomass in almost every yeast strain decreases. However, these differences are not so significant for most strains and range from 0.2–1.0 g/L. A significant decrease in biomass production with increasing C/N ratio was observed in four strains of M. chrysoperlae 1158, M. pulcherrima 1232, M. fructicola 1235, and M. zizyphicola 1247. In the case of these strains, the decrease in biomass production at the highest C/N ratio was more than 2 g/L compared to the lowest C/N ratio. Lipid production increases with increasing C/N ratio in all strains. By cultivating the yeast on glucose media with a C/N 97 ratio under reduced temperature conditions (15 °C), a relatively high lipid production was achieved. In almost half of the strains used, lipid production was more than 12% by volume of biomass.
By increasing the C/N 150 ratio in the glucose medium, relatively high microbial lipid productions were also observed. Some strains achieved almost 20% lipids from the dry weight of biomass; in particular, the yeast M. andauensis 1241 reached 19.6 ± 1.6% lipids in the production of 8.2 g/L biomass. This phenomenon was caused by the lower content of nitrogen source in the C/N 150 medium, which led to its rapid depletion, and the cells stopped their growth and began to accumulate lipids.
There were no significant variations in lipid or biomass production on both culture media. From this we can conclude that the yeast showed a similar trend on media with the same substrate. When the C/N ratio was changed, only the percentual lipid content and biomass production changed. In general, at higher C/N ratios, yeast showed a higher percentage of microbial lipid production [
9].
3.2. Screening of Lipid Production in Stressed Metschnikowia Strains Grown on Glycerol as a Carbon Source
More than 95% of the raw materials for biodiesel production come from edible oils, which is the cause of deforestation in some countries, due to the increase in required agricultural land. The main by-product of biodiesel is crude glycerol, but refining crude glycerol to pure glycerol is very expensive. According to research from 2016, crude glycerol can be used as the main carbon source for lipid-producing organisms [
17,
19,
20].
In the next part of the work, therefore, the carbon source of glucose was replaced by glycerol. The other conditions were maintained as in the previous part of the study, namely low temperature and C/N ratios of 24, 97, and 150.
Relatively high biomass production in g/L was recorded for all strains cultivated on glycerol medium with C/N 24. The most biomass with a volume of 27.7 g/L was produced by the yeast
M. sinensis 1244 (
Table 6), which was more than 2 times higher than glucose media. The results show that in general, glycerol is a better carbon substrate in terms of biomass production.
However, despite the same amounts of biomass, these strains did not show identical production properties in terms of lipid formation. In the strain M. zizyphicola 1247, up to 35.5% of lipids from the total biomass content were synthesized, which was also the highest measured value of lipid content from the total biomass.
In the monitoring of lipid production by the yeasts, M. pulcherrima 145, 147, and M.andauensis 129 on glycerol media, there were no significant deviations depending on the ratios of C/N used, namely 24, 97, and 150. Compared to the previous cultivation of yeast on glucose as a carbon source, there were no significant changes in the production of either biomass or lipids, as is the case with other monitored yeast strains.
In these other strains, in comparison with media where glucose was the main carbon source, higher biomass yields were achieved on glycerol and at the same time higher yields of microbial lipids. Thus, it can be argued that the glycerol content effectively promoted microbial growth in most of the strains studied.
The use of glycerol as a waste substrate therefore has the great advantage of its easy availability and low cost. At the same time, its use shows better production properties.
In the biotechnological process, glycerol serves as one of many potential sources to produce microbial lipids by oleogenic microorganisms. Therefore, the ability to utilize pure or crude glycerol has been the subject of several studies. The yeast
M. pulcherrima, whose lipid content was determined to be almost 40% after 14 days of cultivation under reduced conditions (15 °C), was also subjected to such studies. This confirmed that the yeast is able to grow and at the same time utilize the glycerol contained in the media in high concentrations [
2].
In the present paper, the best production properties were achieved by the yeast M. pulcherrima 1232, while the content of accumulated lipids in its cells was up to 36.31%. Very good yields of lipids from biomass also showed Metschnikowia sinensis 1244 for all measured media. However, maximum lipid production of nearly 30% was achieved for media with a C/N ratio of 150. For M. zizyphicola 1247, the maximum was at 35.5% for media with a C/N ratio of 24.
Compared to the cited study, no significant difference in the amount of lipids produced is observed. The mutual comparison of studies and the results of the work proves that the C/N ratio, temperature conditions, and the cultivation time itself significantly affect the production of lipids in oleogenic yeasts. High lipid yields on media where glycerol acted as the primary carbon source have been demonstrated in other yeasts, such as e.g.,
Yarrowia lipolytica,
Candida curvata, or
Rhodotorula graminis. By cultivation of glycerol, microbial lipid yields represented between 28 and 53% of the biomass content [
17]. Glycerol is an important carbon source for the yeast of the genus
Metchnikowia targeted for the biotechnological production of high-value lipids. Thus, we can consider that with targeted conversion of lipid-rich waste—such as animal fat, which is a rich source for TAG—we can contribute to the reduction of the burden on the environment by decreasing the amount of waste substrates.
Monitoring of Fatty Acid Profile in Stressed Metschnikowia Strains
The fatty acid profile of the individual strains cultivated on glucose and glycerol as the main carbon source was determined by gas chromatography. Depending on the type of medium used and the C/N ratio, the production strains of the examined strains were compared with each other.
The results of the analysis of the fatty acid produced by
Metschnikowia yeasts cultivated on different C/N ratios show a stable production of individual types of fatty acids in all studied yeast strains on both types of carbon sources. As the amount of carbon in the medium increases, we observe a slight increase in the content of unsaturated fatty acids. Thus, it can be stated that increasing the amount of carbon in the medium has a positive effect on the production of unsaturated fatty acids, but only to a certain extent. On media with a large excess of carbon (C/N 150), the results show the opposite effect, where the production of unsaturated fatty acids decreases to values comparable to media with a low C/N ratio or even less. The complete GC results describing the content of fatty acids are shown in
Tables S1 and S2 in the
Supplementary Material.
The majority of fatty acids produced by
Metschnikowia yeasts cultivated on both glucose (
Section 3.1.2) and glycerol (
Section 3.2) media with the same C/N ratio was oleic acid and linoleic acid. The use of glycerol as a carbon source led to higher yields of linoleic acid compared to glucose medium. Palmitic acid, palmitoleic acid, and stearic acid accounted for a significant proportion of fatty acids. In addition, it can be seen immediately that the yeast profile cultivated on glycerol is more diverse, enriched with significant unsaturated FA such as α-linolenic acid or arachidonic acid.
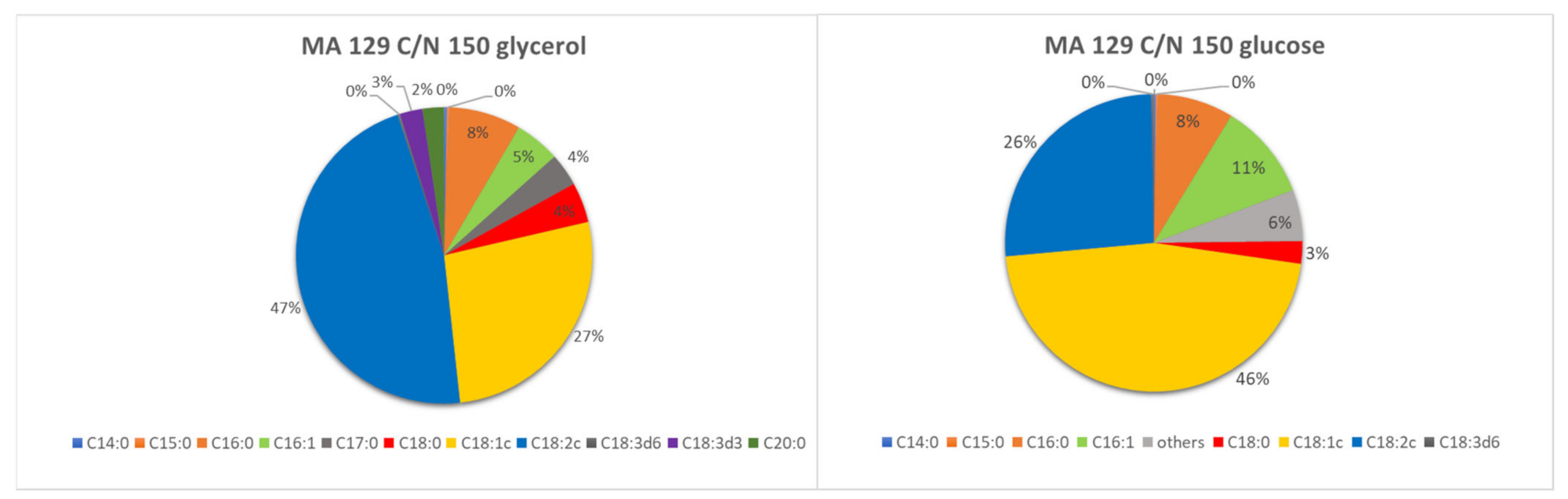
The yeast
Metschnikowia andauensis 129 (MA129) differed in the proportion of fatty acids at the same C/N ratio but different carbon sources (
Figure 3). While oleic acid, linoleic acid, and palmitoleic acid predominated in the glucose medium, linoleic acid predominated in the glycerol medium. Complete results of fatty acid profile analysis are listed in
Tables S1 and S2.
Cultivation on glycerol medium resulted in gamma and alpha-linolenic acid, which did not occur during cultivation on glucose. Although the fatty acid profile of the yeast M. andauensis 129 was strongly dependent on the carbon substrate used, the individual C/N ratios studied (24, 97, and 150) did not significantly affect it, and thus the proportions on the individual glycerol and glucose media are very similar.
The other studied strains of M. pulcherrima (145, 147, and 149) showed a higher dependence of the FA profile on the ratio of carbon and nitrogen in the medium compared to M. andauensis 129. The yeasts of genus Metschnikowia showed a C/N ratio of 97 on glycerol as the most preferred for linoleic acid formation, as well as to glucose media.
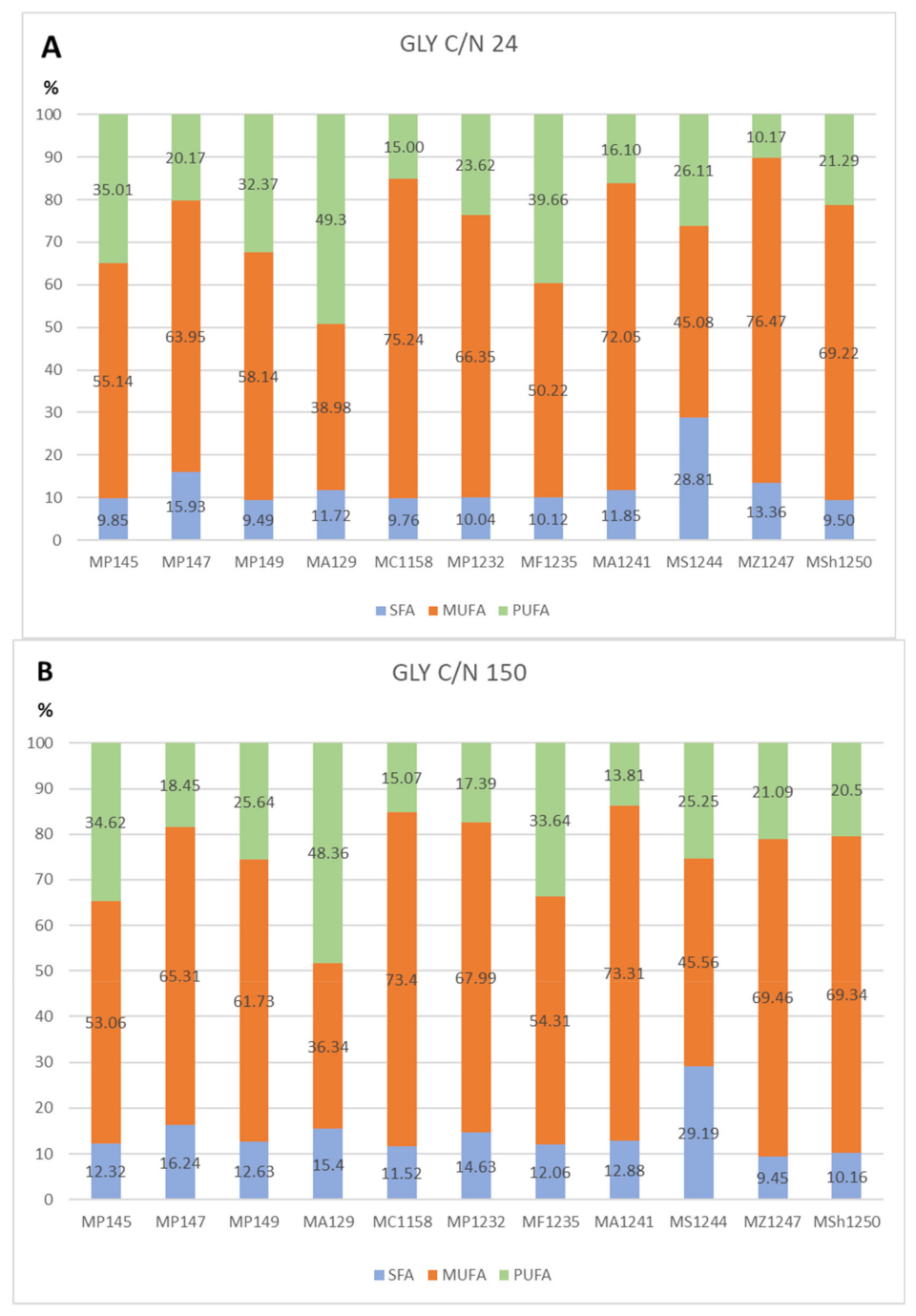
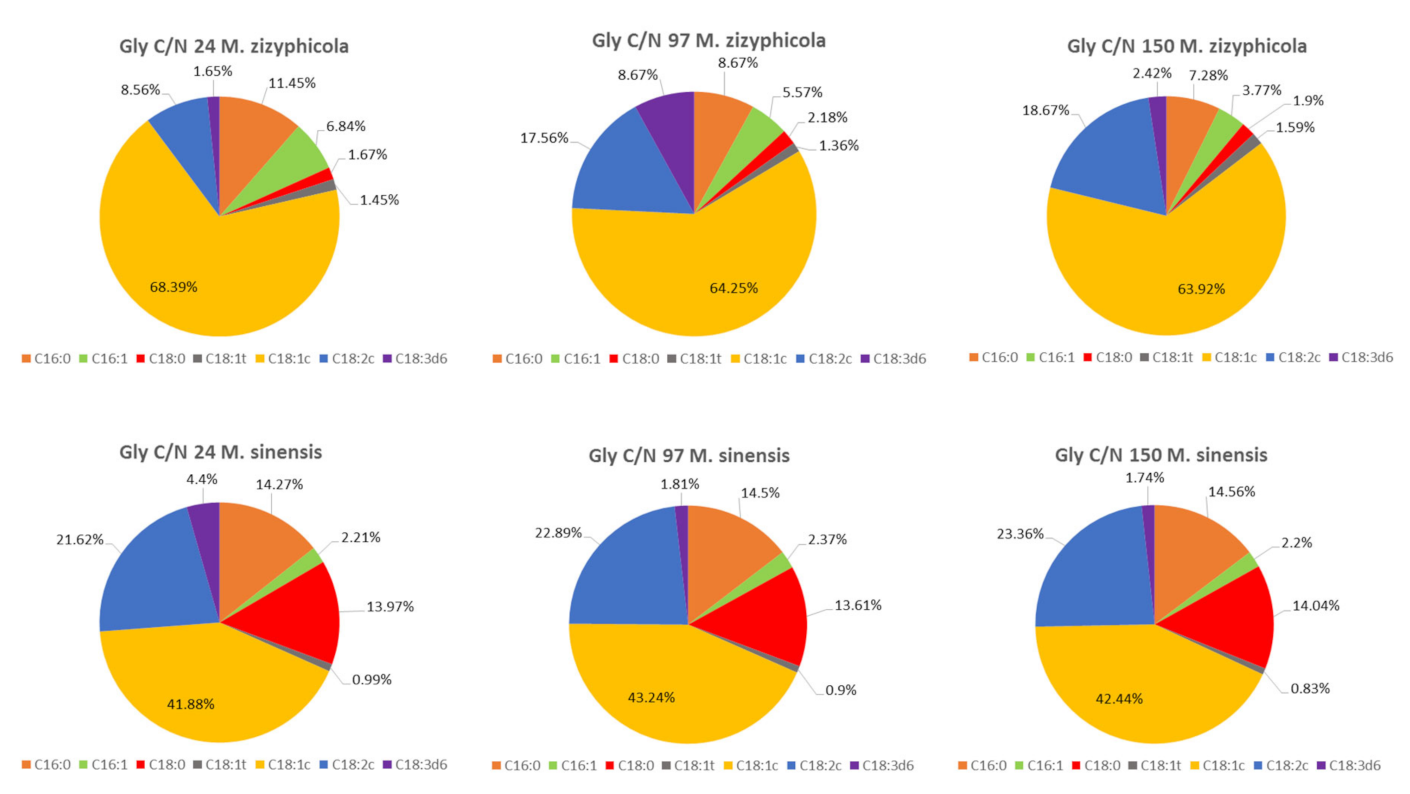
All strains showed a very similar fatty acid profile when growing in glycerol media with various C/N ratios (
Figure 4A,B see selected C/N ratio 24 and 150). The only exception was the yeast
M. sinensis 1244 (
Figure 4 and
Figure 5), for which the production of some types of fatty acids was different. As the only strain, it was able to produce the highest content of palmitic and stearic acid on all glycerol as well as glucose media. At the same time, it was also the one of lowest producers of oleic acid in these media. Thus, the difference in fatty acid production could probably be related to the genetic makeup of this yeast. The most common acids were oleic acid and linoleic acid. Palmitic, stearic, and palmitoleic acids also formed a certain proportion of the profile. Alpha-linolenic acid was also present in glycerol media, which was not in the fatty acid profile of glucose media.
The results of other cultured strains of
Metschnikowia yeast show a stable trend of fatty acid production. Even in media with different C/N ratios, the yeast maintains a relatively stable FA profile, which varies only in the range of 1–5%. In general, with an increased C/N ratio, we observe a slight increase in PUFA production at the expense of MUFA (
Figure 4 and
Figure 5).
As shown in
Figure 5 the production of oleic acid by increasing C/N in the yeast
M. zizyphicola 1232 tends to decrease. As C/N increased, so did the content of palmitic and palmitoleic acid. However, the positive effect of increasing C/N was demonstrated in the formation of linoleic acid, where the % proportion of acid also increased with increasing C/N. The direct effect of C/N on stearic acid production has not been demonstrated, as there has been no decrease or increase in C/N production with increasing C/N. More results are shown in
Table S2.
An increase or decrease in the C/N ratio did not have a direct effect on oleic acid production in the yeast M. sinensis 1244. However, the increasing C/N ratio had a favorable effect on the production of palmitic and linoleic acid, the content of which increased proportionally even with increasing C/N. Stearic acid did not differ in amount in different media but reached the highest values in all studied yeasts of the genus Metschnikowia when cultivated on glucose or glycerol at different C/N ratios. Compared to glucose media, with the biotechnological focus on increased PUFA production in Metschnikowia yeasts, there is no marked difference between C/N ratios of 97 and 150 on glycerol media. Thus, it can be concluded that the yeasts of the genus Metschnikowia are very stable in the area of fatty acid production and are not affected by the different C/N ratios. As a result, mainly the total lipid production and the associated fatty acid production can be increased by increasing the C/N ratio. By exchanging a simple carbon source, the fatty acid profile can then be modulated to some extent.
3.3. Production of Lipids in Metschnikowia Strains Cultivated on Crude Animal Fat as a Carbon Source
With the growing expansion of the meat industry, more and more animal fat is being produced as a by-product, which is currently considered to be one of the main sources in biodiesel production [
21,
22,
23]. From a chemical point of view, animal fats are triacylglycerols in which glycerol is esterified with three fatty acids. Animal fats are characterized by a higher content of saturated fatty acids or long chain fatty acids [
24].
Again, the media were prepared to maintain the selected C/N ratio, and in particular 24, 97, and 150 ratios at cold temperature were used (see
Section 2.3)
In the comparison to glucose and glycerol media, the relatively lowest biomass yields were achieved on fat media at the C/N ratio 24–150 (
Table 7), similar to the study [
11] where crude fat was used to cultivate carotenogenic yeast. Limitation of the nitrogen source in C/N 150 fat media slightly increased lipid accumulation in yeast cells. In the fat medium C/N 97, the yeast
M. andauensis 1241 showed the highest lipid production. The lipid content was 14.4% by weight of the biomass.
The results confirm that all studied strains of Metschnikowia yeast were able to utilize crude animal fat. All yeasts were able to accumulate more than 5% lipids in their cells. The yeast M. pulcherrima 1232, which achieved very good production on glycerol media, was rather average on fat media. Additionally, the yeast M. sinensis 1244, a good producer of glycerol media, achieved the lowest production of lipids on fat media with C/N ratio 150.
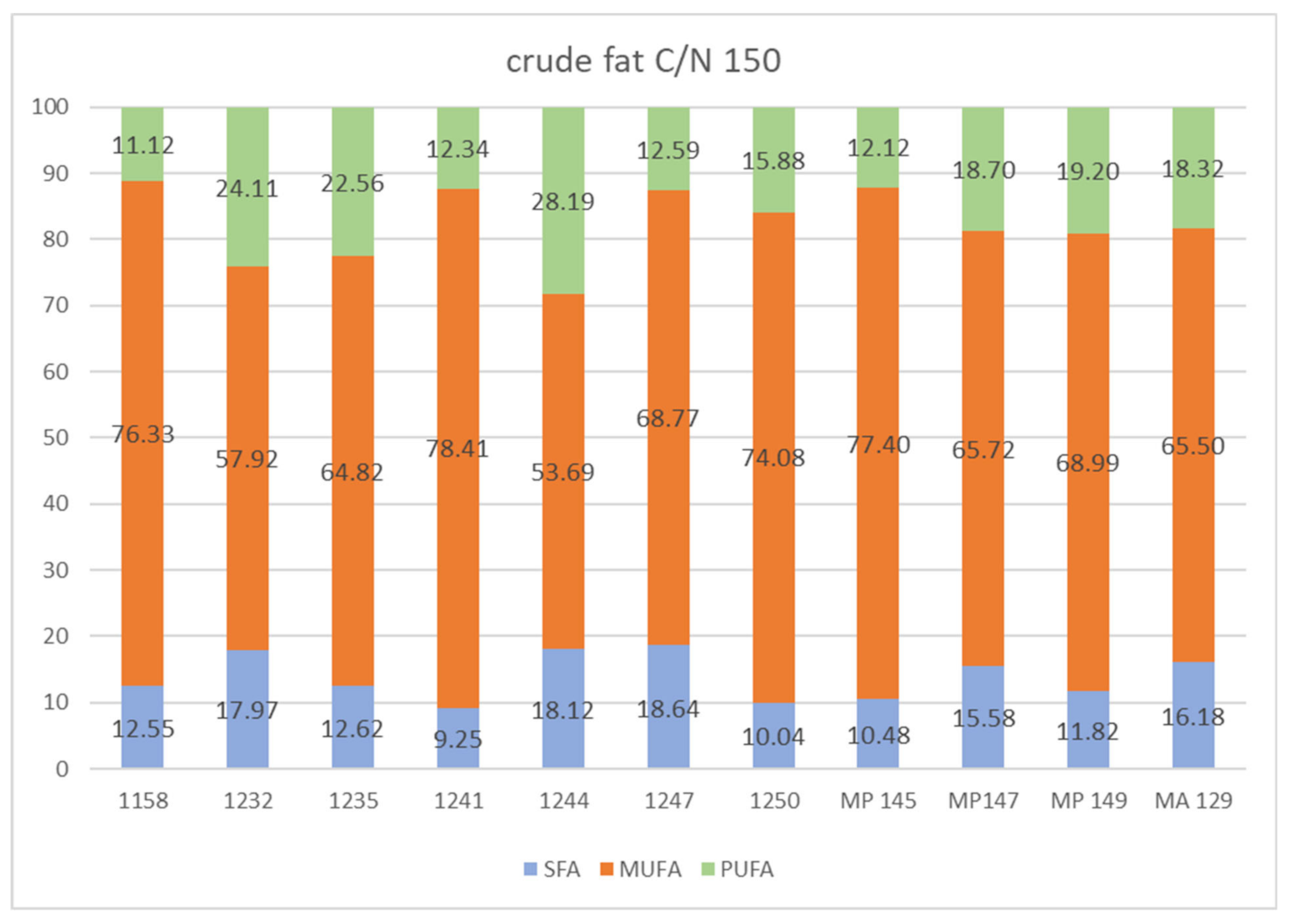
In the comparison of fat media, the relatively lowest biomass yields were achieved at a C/N ratio of 150. Limitation of the nitrogen source in C/N 150 fat media slightly increased the lipid accumulation in yeast cells. Overall, of all fat media at the C/N ratio of 150, the largest amount of lipids was recorded, around 20.35% in the yeast M. zizyphicola 1247.
The lowest biomass production was recorded by cultivation on waste fat substrate. This phenomenon may have been since the yeast was probably not able to utilize pure fat in sufficient quantities due to its solid state in cultivation conditions. The carbon contained in the substrate was difficult for them to access, which was reflected in the amount of biomass produced. In order to make the fat more accessible to yeast, it would be appropriate to use an emulsifier in the future or to modify the fat technologically, for example by hydrolysis. After hydrolysis of the fat, carbon is released into the medium mainly in the form of fatty acids. A minor proportion is glycerol, which can serve as a rapid source of energy for yeast sufficient for the initial growth of biomass. Subsequently, yeast metabolism focuses on fatty acid processing.
The fatty acid profile accumulated in the cells was monitored in all studied yeasts of the genus Metschnikowia using the gas chromatography technique. The fatty acid profile was compared between strains according to the C/N ratio of the medium used.
The presence of palmitic and palmitoleic acid was confirmed in all strains examined by cultivation on waste fat with a C/N ratio of 24. Stearic acid was also present in small amounts in all yeasts. Of all the fatty acids, oleic acid was the most produced. In the cells of some yeasts, its content was up to 74.82% (M. zizyphicola 1247), 74.77% (M. chrysoperlae 1158), and 72.92% (M. pulcherrima 145) of total fatty acids. When cultivated on C/N 24, the oleic acid content of all strains always accounted for more than half of the fatty acid content. As in glycerol cultures, yeasts of the genus Metschnikowia have shown the ability to produce linoleic and α-linolenic acid on the fat medium. The content of α-linolenic acid ranged from 0.83% (M. pulcherrima 147) to 4.68% (M. sinensis 1244).
As with the lower C/N medium, yeast when cultivated at a C/N ratio of 97 showed a similar trend in fatty acid production. Again, the presence of oleic, palmitic, stearic, and palmitoleic acids was detected in all yeasts examined. As with C/N 24 and C/N 97, an increased content of Ω-3 fatty α-linolenic acid was observed in M. sinensis strain 1244 among other yeasts. At C/N 24, the content of α-linolenic acid was 4.68%, and at C/N 97, the content slightly increased to 5.93%.
Figure 6 shows the content of fatty acids present in the biomass of yeast cultivated on a fat medium in C/N 150. The production of palmitic, palmitoleic, and stearic acids was recorded on this medium. Relatively high palmitic acid production was observed in
M. pulcherrima 1232
strain compared to the yields obtained on the medium with C/N 97. The highest yield of palmitic acid was obtained on the fat medium with C/N 150. Relatively increased yields were also observed for stearic acid production ranging from 0.88% (
M. andauensis 1241) to 14.06% (
M. zizyphicola 1247). Oleic acid had the highest proportion of all fatty acids. The oleic acid content of
M. pulcherrima 145 was up to 71.42%. It can also be seen from
Figure 6 that the presence of α-linoleic acid was detected in a smaller amount in the cells of all examined yeasts, but it was mostly contained again in the yeast
M. sinensis 1244 (5.67%).
From the results given above and in
Figure 6, which summarize the proportion of saturated and unsaturated fatty acids in the obtained microbial lipids, it can be clearly stated that in all strains of yeasts of the genus
Metschnikowia there was a significant predominance of monounsaturated fatty acids.
The obtained results demonstrate the ability of all monitored strains of yeasts of the genus Metschnikowia to use waste animal fat for their growth and lipid production.
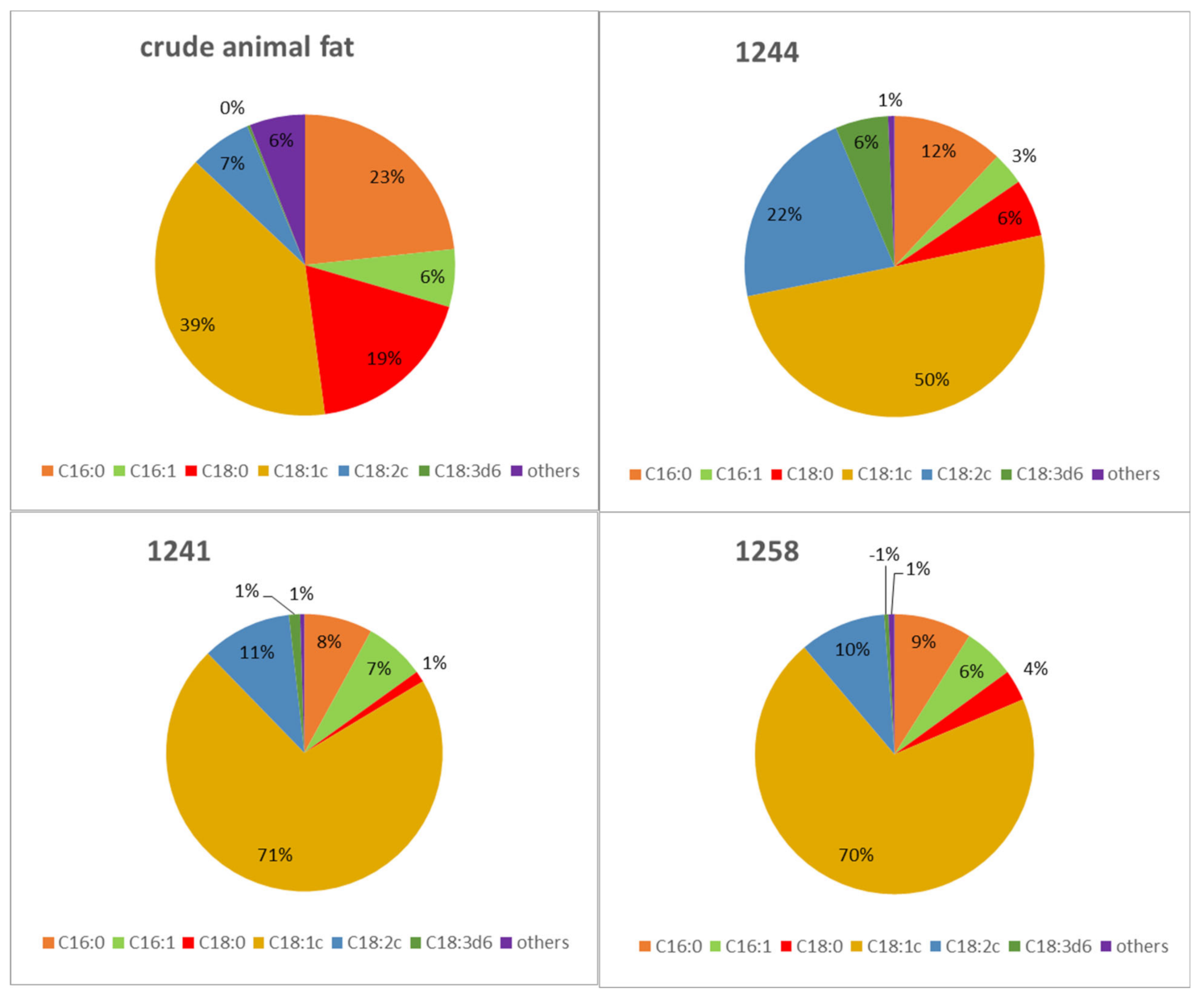
As mentioned above, the fatty acids from crude animal fat were composed of the sum of palmitic acid (23.3%), stearic acid (18.4%), and oleic acid (39.1%), while the summary content of linoleic and linolenic acids represented about 7% [
11].
Figure 7 shows conversion of animal fat into other lipid metabolites for selected yeasts of the genus
Metschnikowia. The content of selected fatty acids is different from the original animal fat and is dependent on the given strain.
Using different biotechnological applications, the potential to grow on raw animal fat for the selected yeasts of the genus
Metschnikowia was investigated. For this carbon source, several strains showed specific growth, biomass, and lipid production, which was higher compared to glucose or glycerol medium. Moreover, monitoring of the fatty acid profile suggests that animal fat could be used as a source of carbon in yeast cultivation for biomass production and conversion to other types of fatty acids. More data and dedicated results are mentioned in
Table S3 in the
Supplementary Material.
As can be seen from
Table S3, the conversion of animal fat by yeast increased the amount of oleic and linolenic acid; oleic acid in turn became the most abundant fatty acid. Compared to the original crude animal fat, which contained a relatively high amount of saturated fatty acids, namely stearic and palmitic, for selected yeasts, the content of these acids is lower. The only strain that differs from other yeast strains is again
Metschnikowia sinensis 1244, which produces higher amounts of stearic acid compared to the other strains. However, this strain has the highest amount of polyunsaturated fatty acids. We would like to note that while crude animal fat contains approximately 20% stearic acid, this fatty acid is significantly degraded in yeast metabolism and converted to other lipid metabolites. The highest potential in the transformation of waste fat with a high SFA content compared to MUFA was observed in strains
M. chrysoperlae 1158,
M. andauensis 1241, and
M. pulcherrima 145, where these strains accumulated more than 70% oleic acid. In contrast, in
M. pulcherrima 1232,
M. fructicola 1235, and
M. sinensis 1244 strains, the transformation of SFA to unsaturated fatty acids also focuses on the formation of PUFA, which reaches more than 20% (C/N 150). The results show that the ability of yeast to transform saturated fatty acids from waste fat to unsaturated fatty acids increases with increasing C/N ratio. Thus, utilization of animal fat by yeasts of the genus
Metschnikowia seems to be very promising for different biotechnological applications. Additionally, the production of various lipid metabolites depending on the strain and culture conditions is possible.
If we compare the results of
Metschnikowia strains grown on glycerol or waste fat with other yeast species, we see that lipid production is relatively lower. The genus
Yarrowia is able to produce 45–55% lipids on waste lipid-based media [
25,
26]. Representatives of carotenogenic yeasts, e.g., the genus
Sporidiobolus [
11], are also able to achieve identical productions of up to 55% on lipid wastes.
However, unlike the above-mentioned species
Yarrowia or
Sporidiobolus, the main biotechnological significance of the genus
Metschnikowia stems from its genetic makeup and ability to transform and synthesize higher fatty acids. These yeasts are thus able to transform wastes with a high SFA content into microbial oils with a high content of PUFA and MUFA. In the case of red yeast, a suitable waste substrate such as coffee oil must be used to achieve biomass with a wide range of unsaturated fatty acids [
11].
3.4. Production of Lipolytic Enzymes by Yeast of the Genus Metschnikowia Cultivated on Animal Fat Substrate
Yeast biomass production in general on waste fat was relatively low. The reason for the low production of fatty media could probably be the low concentration of waste fat, which did not represent a sufficient amount of carbon in the medium, or the carbon contained in the substrate was difficult to obtain for them. However, it should be noted that yeasts of the genus
Metschnikowia were in the past included in the genus
Candida, based on several similarities between these currently two genera [
27]. Some yeasts of the genus
Candida are known to overproduce lipases [
28]. In the present study, it was hypothesized that yeasts of the genus
Metschnikowia could also show increased lipase activity, which could be related to the ability to easily degrade the animal fat present in the medium, utilize it, and accumulate lipids at the same time. By using waste fat with yeast, we can obtain new biotechnologically important products from processed industrial waste and at the same time reduce the burden on the environment [
29,
30].
It was proved that the yeasts of the genus
Metschnikowia showed a potential ability to utilize waste substrates in the form of animal fat. Thus, in the next step, we monitored their ability to produce hydrolytic enzymes, namely lipases. Quantitative screening of lipase enzyme activity was performed on selected yeasts of the genus
Metschnikowia using a p-nitrophenyl palmitate spectrophotometric method [
31]. Consequently, the amount of p-nitrophenol released by the action of the enzyme on p-NPP was measured. The lipase activity of the individual strains was expressed in units of nmol/mL·min. Culture media containing waste fat as the main carbon source were used to determine the enzymatic activity of
Metschnikowia yeast lipases. The culture media were prepared as the optimal medium (see
Section 2.3), where instead of glucose a reference amount of fat (47 g/L) was added as a carbon source. In addition, an emulsifier in the form of polysorbate 80 (Tween 80) was added to one culture series containing fat. In order to increase the activity of extracellular lipases, surfactants are often added to the medium [
32,
33]. In the present work, Tween 80 (a non-ionic polyoxyethylene detergent) was used to investigate lipase activities, which stimulates the biosynthesis and secretion of lipases because it increases cell permeability and enables lipase export across the cell membrane.
Table 8 shows a comparison of the lipolytic activity which selected yeast samples showed on fat media and on media where Tween 80 was added. Lipase production was confirmed in all strains under investigation.
It can be seen from
Table 8 that overall, the lowest enzymatic activity of lipases on the unmodified waste fat was recorded for
M. zizyphicola strain 1247 (0.57 nmol/min mL). In spite of the fact that lipid substrates mostly support lipase production, in this case, the yeast produced a small amount of lipolytic enzymes. The fat substrate was most likely a complex carbon source for this yeast.
However, by adding Tween 80 to the medium, an increase in lipase activity to 0.73 nmol/min mL can be observed. Thus, for this strain, it can be assumed that in the presence of an emulsifier, the waste fat was used more efficiently. A similar trend of lipase activities can be observed in other studied strains. It can be concluded that Tween partially contributed to better emulsification of the fat medium, thus showing higher lipase activity in these strains.
The highest lipolytic activity observed on pure fat medium was recorded in M. chrysoperlae strain 1158 1.13 nmol/min mL and in the yeast M. sinensis 1244 with a value of 1.05 nmol/min mL. Moreover, the highest enzymatic activity of lipases was observed in this strain (M. sinensis 1244) on media containing Tween 80, where it was 1.30 nmol/min mL. From these experiments, a positive effect of the emulsifying properties of Tween 80 can be readily observed.
However, compared to other yeasts, the measured enzymatic activities of
Metschnikowia yeast lipases were relatively small. In the yeast
Y. lipolytica, after 72 h of cultivation, the production of extracellular lipases was determined to be 2.8 ∙ 10
4 nmol/min ∙ ml, in its mutant strains, the production increased even up to 1 × 10
6 nmol min mL [
34]. We could observe that majority of yeasts show higher values of lipase activity during the later exponential phase.
A possible explanation of lower lipase activities could be the fact that the activity was not observed during the later exponential phase of the growth curve but at the end of the stationary phase of growth. This finding is also supported by the amounts of residual fat that were determined after the supernatant cultivation. We found that in some strains, fat was used up to 80%. However, on average, fat was used at 60%.
Previous studies have shown that olive oil is a possible carbon source substrate that can increase lipase synthesis. It contains approximately 55–83% oleic acid, which acts as an inhibitor for the production of a gene encoding lipase synthesis [
33,
34].
The addition of an emulsifier or the addition of oleic acid to the medium can induce an increased production of lipases in yeasts of the genus Metschnikowia and thus a better use of animal fat for the production of lipid metabolites. Higher amounts of lipases can break down TAG from animal fat, and the released glycerol will be used as a carbon source. From the results mentioned above, it can be seen that yeasts can utilize glycerol and have excellent production properties of both biomass and lipid metabolites. Therefore, the yeast conversion of crude processed animal fat into value-added products is a valuable process for the food industry.
3.5. Rapid Analysis of Lipids during Cultivation by Raman Spectroscopy
Fatty acid profiles were measured by GC. Simultaneously, we monitored the metabolic states of yeasts in order to optimize the cultivation process for biotechnological applications using Raman spectroscopy.
Raman spectroscopy offers a powerful alternative analytical method for the detection and identification of different substances in biological samples, such as bacteria, algae, and yeast. Raman spectroscopy can be exploited in instances where fast and accurate monitoring/determination of samples is required [
35,
36].
Here, attention is given to the oil-producing yeast strain genus
Metschnikowia to exploit its potential applications in the biotechnology field. In order to utilize selected microorganisms for efficient biotechnological production, the influence of different cultivation parameters (such as the effects of temperature regime and medium composition) on cells can be monitored using different instrumentation [
9].
In this study, Raman spectroscopy was used for monitoring changes in saturation degree of fatty acids (iodine number) in yeast cells. Raman spectroscopy can be utilized for fast and accurate lipids estimation as the intensity ratios of specific, selected Raman bands (lipid CH2 scissoring 1.445 cm
−1, lipid C=C stretching 1.656 cm
−1). Raman spectroscopy proved to be a very efficient tool for the rapid quantitative/qualitative analyses of oil produced by yeast. It can take several minutes [
12,
13].
The figure (
Figure S1) shows the Raman spectra of the yeast
M. sinensis 1244 cultivated on glycerol medium at two C/N ratios, 97 and 150. The work compared the results obtained from gas chromatography and Raman spectroscopy using iodine number (
Table S4). It can be seen from the figure and the table that the yeast
M. sinensis had a similar representation of saturated and unsaturated fatty acids on both mentioned media. This is also proven by the values of iodine numbers, which differ only minimally. We know from the results of gas chromatography that the yeast
M. sinensis 1244 had a similar proportion of fatty acids and their amount in both culture media. In a previous study, Raman spectroscopy was successfully used to monitor unsaturation during cultivation on glucose media [
9].
On the contrary is the case of
M. pulcherrima 1232 and
M. sinensis 1244 cultured on the same media with glycerol of C/N ratio 150. In (
Figure S2) and (
Table S5), it is seen that greater quantities of unsaturated fatty acids were produced by yeast
M pulcherrima 1232 versus
M. sinensis 1244. This is also evidenced by the iodine number calculated from the Raman spectrum. The work shows only illustrative examples due to the large number of spectra obtained.
By detecting the presence of fatty acids by the iodine number method [
13], it was demonstrated that the results obtained from the Raman spectra approximately corresponded to the results obtained by gas chromatography with slight deviations. Thus, the results support the use of Raman spectroscopy in the analysis of microbial lipids already during the culture process, as this method provides a rapid and relatively accurate estimate of the unsaturation of the lipids produced. Thanks to Raman spectroscopy, we can quickly estimate the success or failure of targeted lipid production by manipulating culture conditions [
36].


 and
and 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}