
Influence of Asafoetida Extract on the Virulence of the Entomopathogenic Nematode Steinernema carpocapsae and Its Symbiotic Bacterium Xenorhabdus nematophila in the Host Pyrrhocoris apterus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Culture
2.2. Asafoetida (ASF) Treatment, Dose Assessment, and Mortality
2.3. Nematode Deterrent Effect and Virulence on ASF-Treated P. apterus
2.4. S. carpocapsae Growth, Infectivity, and Virulence Assay on Host P. apterus
2.5. Isolation of Symbioitc Bacterium X. nematophila from P. apterus, Turbidity Test, and Its Virulance on P. apterus
2.6. Disc Diffusion Assay: Asafoetida Antimicrobial Activity Assessment Using Xenorhabdus spp.
2.7. P. apterus Immunity Assessment: Pro-Phenol Oxidase (PPO) and Phenoloxidase (PO) Assays
2.8. Data Presentation and Statistical Analyses
3. Results
3.1. Asafoetida (ASF) Dose and Toxicity Assessment with Nematodes
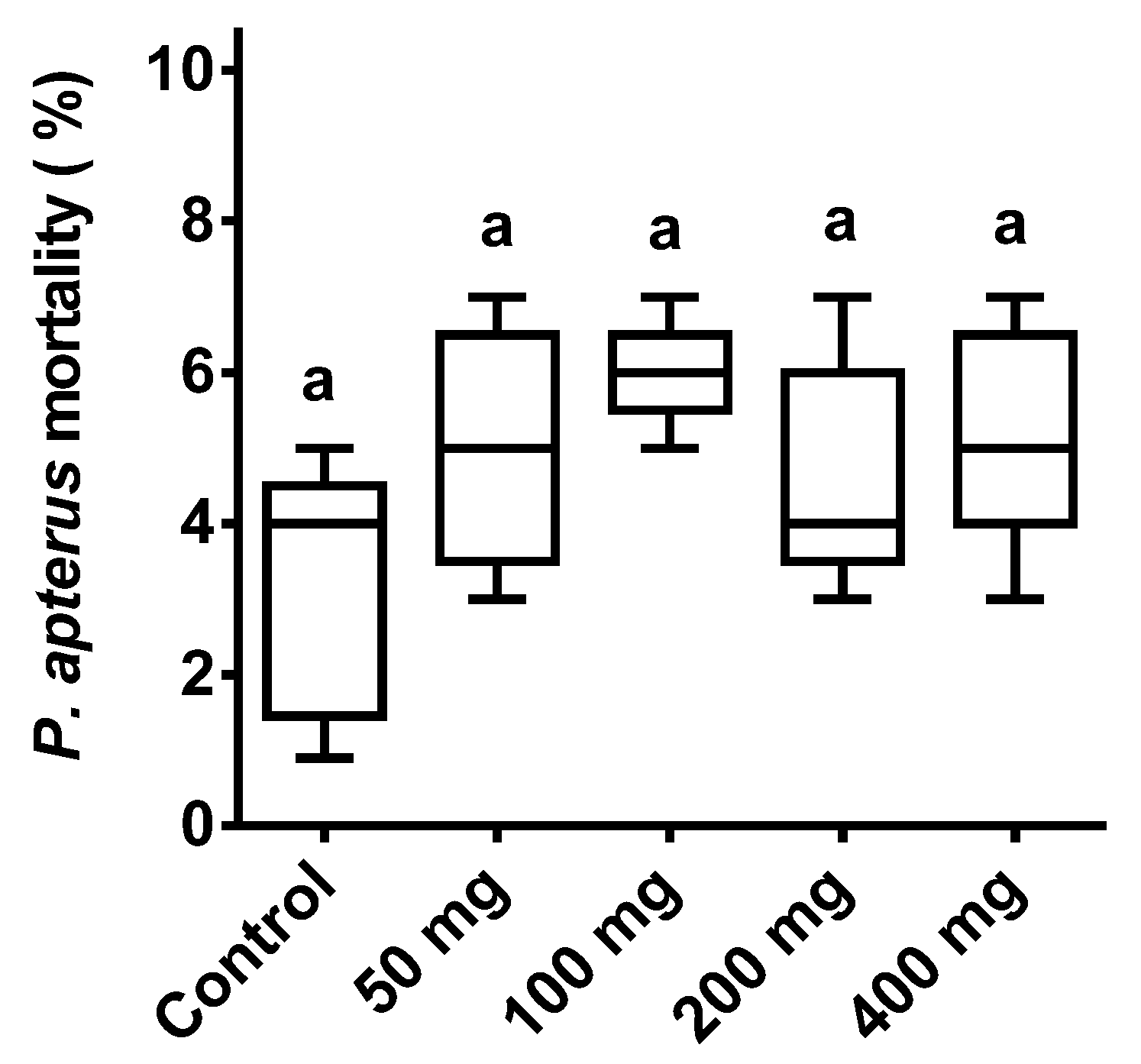
3.2. Asafoetida (ASF) Dose and Toxicity Assessment with Firebug P. apterus
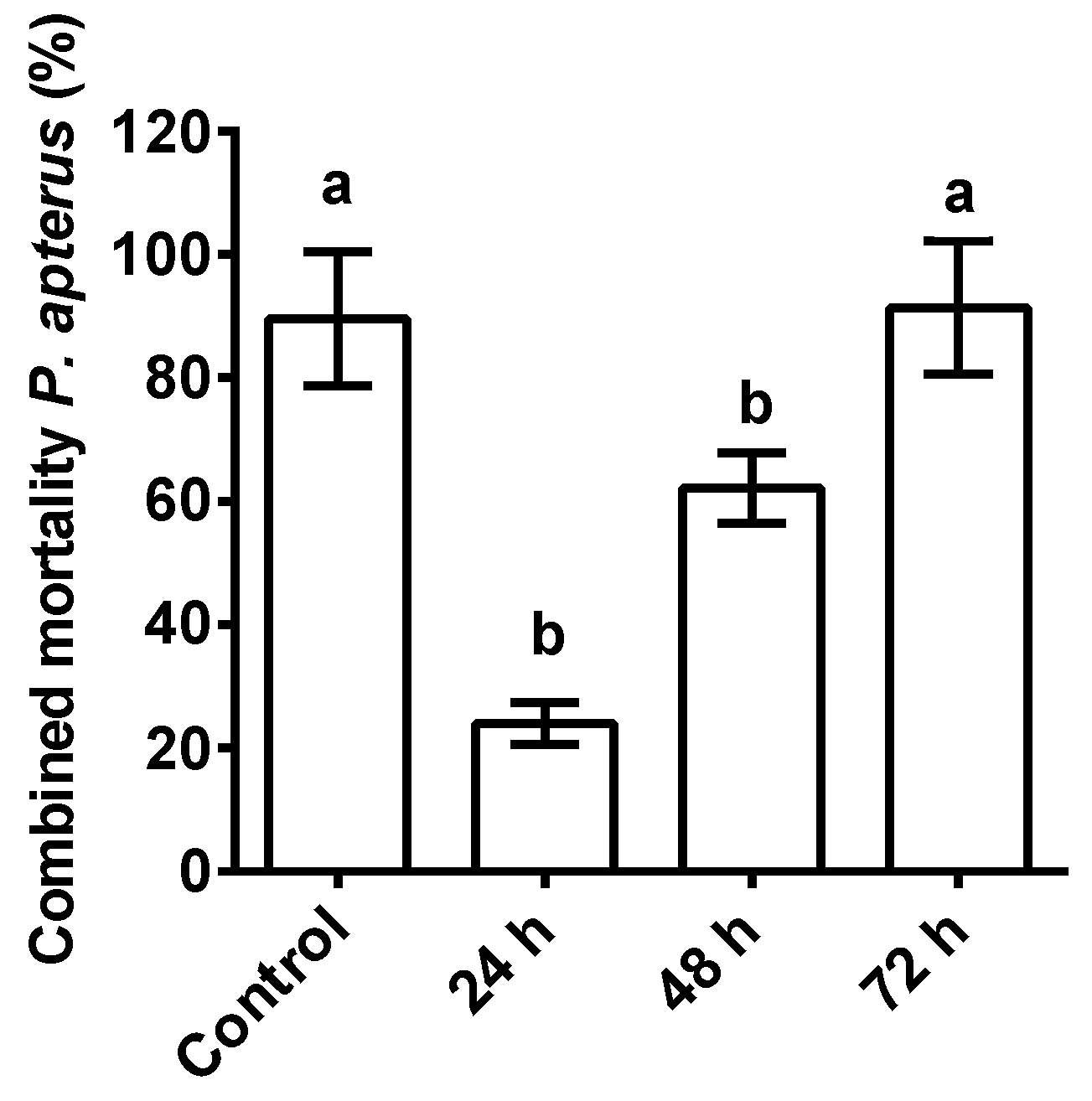
3.3. Combined Effect of ASF and Nematodes on Firebug P. apterus Mortality
3.4. Combined Effect of ASF Pre-Treated P. apterus on Nematode Infectivity and Deterrant Effect
3.5. Effect of ASF on Symbiotic Bacteria X. nematophila Growth, Turbidity Test, and Its Effect on P. apterus Mortality
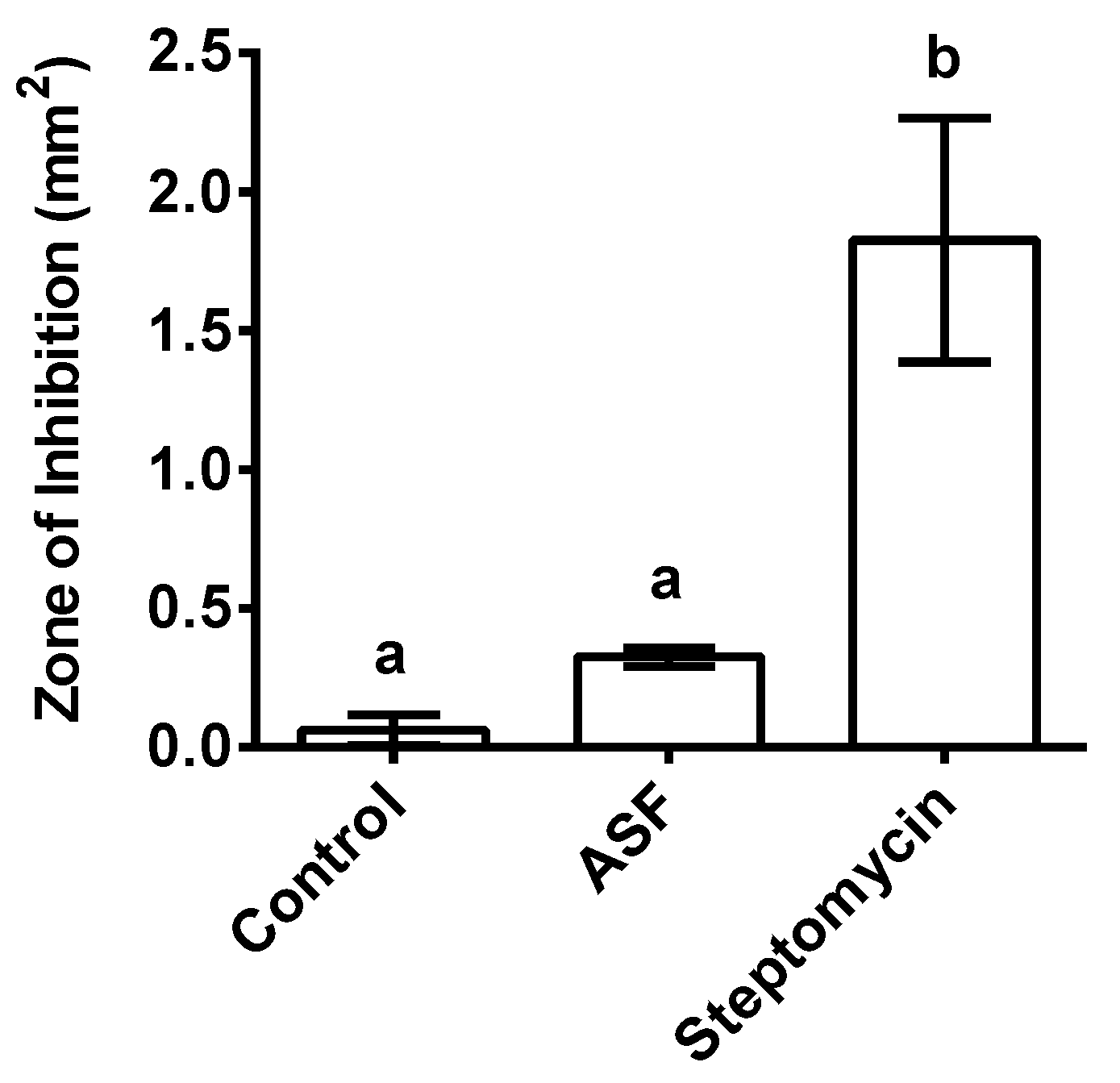
3.6. Growth of Symbiotic Bacteria X. nematophila on ASF: A Disc Diffusion Assay
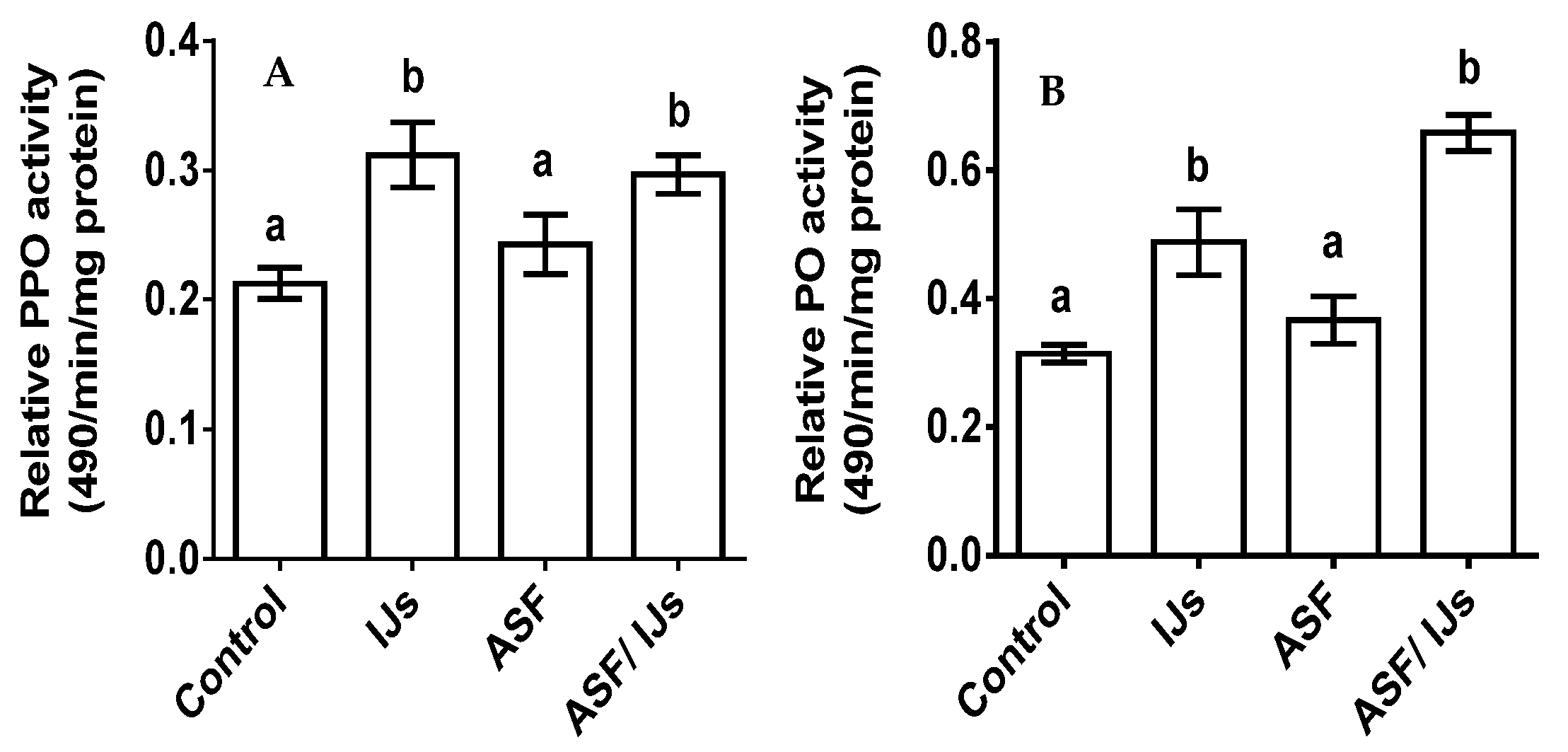
3.7. Effect of ASF on Pro-PO and PO on P. apterus: Assessments of Immunity Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EPNs | Entomopathogenic nematodes |
| IJs | Infective juveniles |
| IP | Intraperitoneal |
| ASF | Asafoetida |
| PPO | Pro-phenol oxidase |
| PO | Phenol oxidase |
References
- Brivio, M.F.; Mastore, M. Nematobacterial Complexes and Insect Hosts: Different Weapons for the Same War. Insects 2018, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Ji, D.; Cho, S.; Park, Y. Two Groups of Entomopathogenic Bacteria, Photorhabdus and Xenorhabdus, Share an Inhibitory Action against Phospholipase A2 to Induce Host Immunodepression. J. Invertebr. Pathol. 2005, 89, 258–264. [Google Scholar] [CrossRef]
- Elbrense, H.; Elmasry, A.M.A.; Seleiman, M.F.; Al-Harbi, M.S.; Abd El-Raheem, A.M. Can Symbiotic Bacteria (Xenorhabdus and Photorhabdus) Be More Efficient than Their Entomopathogenic Nematodes against Pieris rapae and Pentodon algerinus Larvae? Biology 2021, 10, 999. [Google Scholar] [CrossRef]
- Garriga, A.; Mastore, M.; Morton, A.; Garcia del Pino, F.; Brivio, M.F. Immune Response of Drosophila suzukii Larvae to Infection with the Nematobacterial Complex Steinernema carpocapsae–Xenorhabdus nematophila. Insects 2020, 11, 210. [Google Scholar] [CrossRef] [Green Version]
- Periago, M.V.; Bethony, J.M. Hookworm Virulence Factors: Making the Most of the Host. Microbes Infect. 2012, 14, 1451–1464. [Google Scholar] [CrossRef]
- Simões, N.; Caldas, C.; Rosa, J.S.; Bonifassi, E.; Laumond, C. Pathogenicity Caused by High Virulent and Low Virulent Strains of Steinernema Carpocapsae to Galleria Mellonella. J. Invertebr. Pathol. 2000, 75, 47–54. [Google Scholar] [CrossRef]
- Shaik, H.A.; Mishra, A.; Hussein, H.M.; Skoková Habuštová, O.; Sehnal, F. Competitive interactions between entomopatho genic nematodes and parasitoid venom. J. Appl. Entomol. 2020, 144, 481–490. [Google Scholar] [CrossRef]
- Zhang, X.; Machado, R.A.; Van Doan, C.; Arce, C.C.; Hu, L.; Robert, C.A. Entomopathogenic Nematodes Increase Predation Success by Inducing Cadaver Volatiles That Attract Healthy Herbivores. eLife 2019, 8, e46668. [Google Scholar] [CrossRef]
- Lu, D.; Macchietto, M.; Chang, D.; Barros, M.M.; Baldwin, J.; Mortazavi, A.; Dillman, A.R. Activated Entomopathogenic Nematode Infective Juveniles Release Lethal Venom Proteins. PLoS Pathog. 2017, 13, e1006302. [Google Scholar] [CrossRef] [Green Version]
- Sisay, B.; Tefera, T.; Wakgari, M.; Ayalew, G.; Mendesil, E. The Efficacy of Selected Synthetic Insecticides and Botanicals against Fall Armyworm, Spodoptera frugiperda, in Maize. Insects 2019, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Shaik, H.A.; Sinha, R.K.; Shah, B.R. Andrographolide: A Herbal-Chemosynthetic Approach for Enhancing Immunity, Combating Viral Infections, and Its Implication on Human Health. Molecules 2021, 26, 7036. [Google Scholar] [CrossRef]
- Filgueiras, C.C.; Willett, D.S. Non-Lethal Effects of Entomopathogenic Nematode Infection. Sci. Rep. 2021, 11, 17090. [Google Scholar] [CrossRef]
- Shakeri, M. A review on investigations on pomegranate neck worm in Iran. In Proceeding on Evaluation of Finding and Current Problems Associated with Spectrobates Ceratoniae Management in Pomegranate; Ministry of Jihad-e-Agriculture, Organization of Research and Education: Tehran, Iran, 2004; pp. 20–45. [Google Scholar]
- Noonari, A.M.; Abro, G.H.; Khuhro, R.D.; Buriro, A.S. Efficacy of biopesticides for management of sucking insect pests of cotton, Gossipium hirsutum (L.). J. Basic Appl. Sci. 2016, 12, 306–313. [Google Scholar]
- Shaik, H.A.; Mishra, A.; Sehnal, F. Silk recycling in larvae of the wax moth, galleria mellonella (lepidoptera: Pyralidae). Eur. J. Entomol. 2017, 114, 61–65. [Google Scholar] [CrossRef] [Green Version]
- El Deeb, H.K.; Al Khadrawy, F.M.; Abd El-Hameid, A.K. Inhibitory Effect of Ferula asafoetida L. (Umbelliferae) on Blastocystis Sp. Subtype 3 Growth In Vitro. Parasitol. Res. 2012, 111, 1213–1221. [Google Scholar] [CrossRef]
- Gautam, U.K.; Bohatá, A.; Shaik, H.A.; Zemek, R.; Kodrík, D. Adipokinetic Hormone Promotes Infection with Entomopathogenic Fungus Isaria Fumosorosea in the Cockroach Periplaneta Americana. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 229, 108677. [Google Scholar] [CrossRef]
- Seal, D.R.; Baniya, A.B.; Dyrdahl-Young, R.; Hochmuth, R.C.; Leppla, N.C.; Fenneman, D.K.; Broughton, R.D.T.; DiGennaro, P. Wireworm (Coleoptera: Elateridae) Species Composition and Management in Sweet Potato Grown in North Florida Using Chemical Insecticides and Entomopathogenic Nematodes. Environ. Entomol. 2020, 49, 1415–1426. [Google Scholar] [CrossRef]
- Farhadi, A.; Youssefi, M.R.; Abouhosseini Tabari, M. Evaluation of the anticestode and antinematode effects of the methanol extract of Ferula asafoetida on experimentally infected rats. J. Babol Univ. Med. Sci. 2016, 18, 47–51. [Google Scholar]
- Tavassoli, M.; Jalilzadeh-Amin, G.; Fard, V.R.B.; Esfandiarpour, R. The In Vitro Effect of Ferula asafoetida and Allium sativum Extracts on Strongylus spp. Ann. Parasitol. 2018, 64, 59–63. [Google Scholar]
- Baloch, G.N.; Tariq, S.; Ehteshamul-Haque, S.; Athar, M.; Sultana, V.; Ara, J. Management of Root Diseases of Eggplant and Watermelon with the Application of Asafoetida and Seaweeds. J. Appl. Bot. Food Qual. 2013, 86, 138–142. [Google Scholar] [CrossRef]
- Gautam, U.K.; Hlávková, D.; Shaik, H.A.; Karaca, I.; Karaca, G.; Sezen, K.; Kodrík, D. Adipokinetic Hormones Enhance the Efficacy of the Entomopathogenic Fungus Isaria fumosorosea in Model and Pest Insects. Pathogens 2020, 9, 801. [Google Scholar] [CrossRef]
- Kaya, H.K.; Stock, S.P. Chapter VI—Techniques in insect nematology. In Manual of Techniques in Insect Pathology; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Shaik, H.A.; Mishra, A.; Sehadová, H.; Kodrík, D. Responses of Sericotropin to Toxic and Pathogenic Challenges: Possible Role in Defense of the Wax Moth Galleria Mellonella. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 227, 108633. [Google Scholar] [CrossRef]
- Panwar, R.; Rana, S.; Dhawan, D.K.; Prasad, K.K. Chemopreventive Efficacy of Different Doses of Ferula asafoetida Oleo-Gum-Resin against 1,2-Dimethylhydrazine (DMH) Induced Rat Colon Carcinogenesis. J. Phytopharm. 2016, 4, 282–286. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Optimizing sampling and extraction methods for plant-parasitic and entomopathogenic nematodes. Plants 2021, 10, 629. [Google Scholar] [CrossRef]
- Wouts, W.M. Mass Production of the Entomogenous Nematode Heterorhabditis Heliothidis (Nematoda: Heterorhabditidae) on Artificial Media. J. Nematol. 1981, 13, 467–469. [Google Scholar]
- Mahar, A.N.; Munir, M.; Elawad, S.; Gowen, S.R.; Hague, N.G.M. Pathogenicity of Bacterium, Xenorhabdus Nematophila Isolated from Entomopathogenic Nematode (Steinernema Carpocapsae) and Its Secretion against Galleria Mellonella Larvae. J. Zhejiang Univ. Sci. B 2005, 6, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Cornet, S.; Franceschi, N.; Bollache, L.; Rigaud, T.; Sorci, G. Variation and Covariation in Infectivity, Virulence and Immunodepression in the Host-Parasite Association Gammarus Pulex-Pomphorhynchus Laevis. Proc. Biol. Sci. 2009, 276, 4229–4236. [Google Scholar] [CrossRef]
- Stoscheck, C.M. Quantitation of Protein. Methods Enzymol. 1990, 182, 50–68. [Google Scholar]
- Golden, J.W.; Riddle, D.L. A Pheromone Influences Larval Development in the Nematode Caenorhabditis Elegans. Science 1982, 218, 578–580. [Google Scholar] [CrossRef]
- Tsang, W.Y.; Sayles, L.C.; Grad, L.I.; Pilgrim, D.B.; Lemire, B.D. Mitochondrial Respiratory Chain Deficiency in Caenorhabditis Elegans Results in Developmental Arrest and Increased Life Span. J. Biol. Chem. 2001, 276, 32240–32246. [Google Scholar] [CrossRef] [Green Version]
- Rea, S.L.; Ventura, N.; Johnson, T.E. Relationship between Mitochondrial Electron Transport Chain Dysfunction, Development, and Life Extension in Caenorhabditis Elegans. PLoS Biol. 2007, 5, e259. [Google Scholar] [CrossRef] [Green Version]
- Iranshahi, M.; Amin, G.R.; Amini, M.; Shafiee, A. Sulfur Containing Derivatives from Ferula Persica Var. Latisecta. Phytochemistry 2003, 63, 965–966. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Iranshahi, M. Biological Activities of Essential Oils from the Genus Ferula (Apiaceae). Asian Biomed. 2010, 4, 835–847. [Google Scholar] [CrossRef] [Green Version]
- Zuckerman, B.M. Hypotheses and Possibilities of Intervention in Nematode Chemoresponses. J. Nematol. 1983, 15, 173–182. [Google Scholar]
- Zuckerman, B.M.; Jansson, H.-B. Nematode Chemotaxis and Possible Mechanisms of Host/prey Recognition. Annu. Rev. Phytopathol. 1984, 22, 95–113. [Google Scholar] [CrossRef]
- Butcher, R.A. Decoding chemical communication in nematodes. Nat. Prod. Rep. 2017, 34, 472–477. [Google Scholar] [CrossRef] [Green Version]
- Haseeb, M.A.; Fried, B. Chemical Communication in Helminths. Adv. Parasitol. 1988, 27, 169–207. [Google Scholar] [PubMed]
- Singh, C.; Parmar, R. Antimicrobial Activity of Resin of Asafoetida (Hing) against Certain Human Pathogenic Bacteria. Adv. Biores. 2018, 9, 161–164. [Google Scholar]
- Divya, K.; Ramalakshmi, K.; Murthy, P.S.; Jagan Mohan Rao, L. Volatile Oils from Ferula asafoetida Varieties and Their Antimicrobial Activity. Lebenson. Wiss. Technol. 2014, 59, 774–779. [Google Scholar] [CrossRef]
- Mishra, N.; Behal, K.K. Antimicrobial activity of some spices against selected microbes. Int. J. Pharm. Pharm. Sci. 2010, 2, 187–196. [Google Scholar]
- Moghadam, D.H.; Sani, A.M.; Sangatash, M.M. Effect of Oleo-Gum Resin of Ferula asafoetida on Growth of Some Food and Crop Contaminating Microbes. Int. J. Adv. Biol. Biomed. Res. 2014, 2, 2788–2794. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaik, H.A.; Mishra, A. Influence of Asafoetida Extract on the Virulence of the Entomopathogenic Nematode Steinernema carpocapsae and Its Symbiotic Bacterium Xenorhabdus nematophila in the Host Pyrrhocoris apterus. Microorganisms 2023, 11, 1678. https://doi.org/10.3390/microorganisms11071678
Shaik HA, Mishra A. Influence of Asafoetida Extract on the Virulence of the Entomopathogenic Nematode Steinernema carpocapsae and Its Symbiotic Bacterium Xenorhabdus nematophila in the Host Pyrrhocoris apterus. Microorganisms. 2023; 11(7):1678. https://doi.org/10.3390/microorganisms11071678
Chicago/Turabian StyleShaik, Haq Abdul, and Archana Mishra. 2023. "Influence of Asafoetida Extract on the Virulence of the Entomopathogenic Nematode Steinernema carpocapsae and Its Symbiotic Bacterium Xenorhabdus nematophila in the Host Pyrrhocoris apterus" Microorganisms 11, no. 7: 1678. https://doi.org/10.3390/microorganisms11071678