
Plant Growth-Promoting Endophytic Bacteria Isolated from Miscanthus giganteus and Their Antifungal Activity
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Bacterial Endophytes
2.2. Identification of Bacterial Isolates
2.3. Detection of ACC-Deaminase Activity
2.4. Siderophore Detection
2.5. Phosphate Solubilization
2.6. Potential for Nitrogen Fixation
2.7. Phytohormones Detection
2.8. Antifungal Activity of Endophytic Bacteria
2.9. Immobilization of Endophytic Microorganisms in Agar and Testing of Their Antifungal Activities
2.10. Immobilization of Endophytic Microorganisms into Dextrin-Coated Cellulose Carriers and Testing of Their Antifungal Activities
3. Results
3.1. Isolation and Identification of Endophytic Bacteria
3.2. Plant Growth-Promoting Potential of Bacterial Isolates
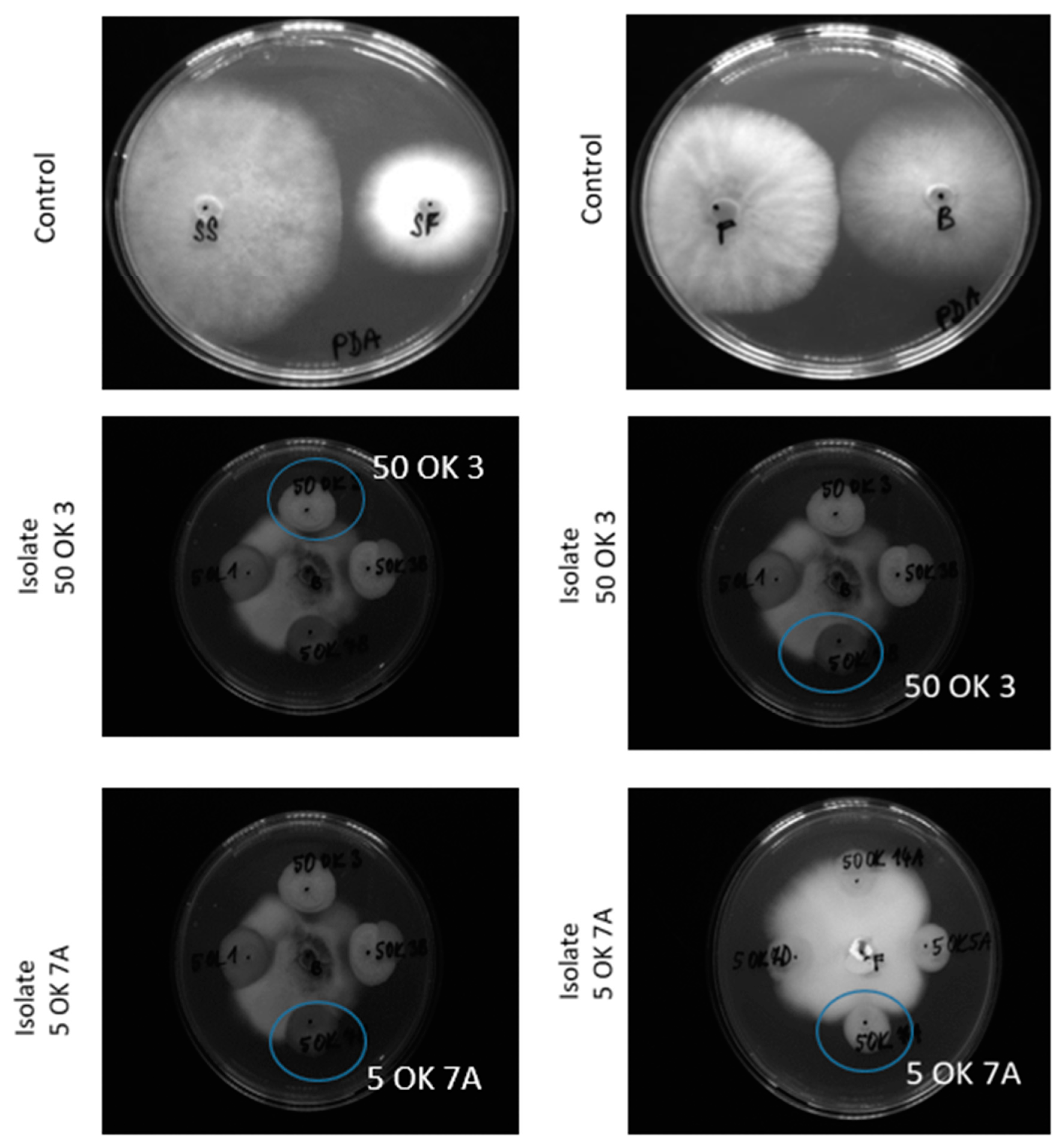
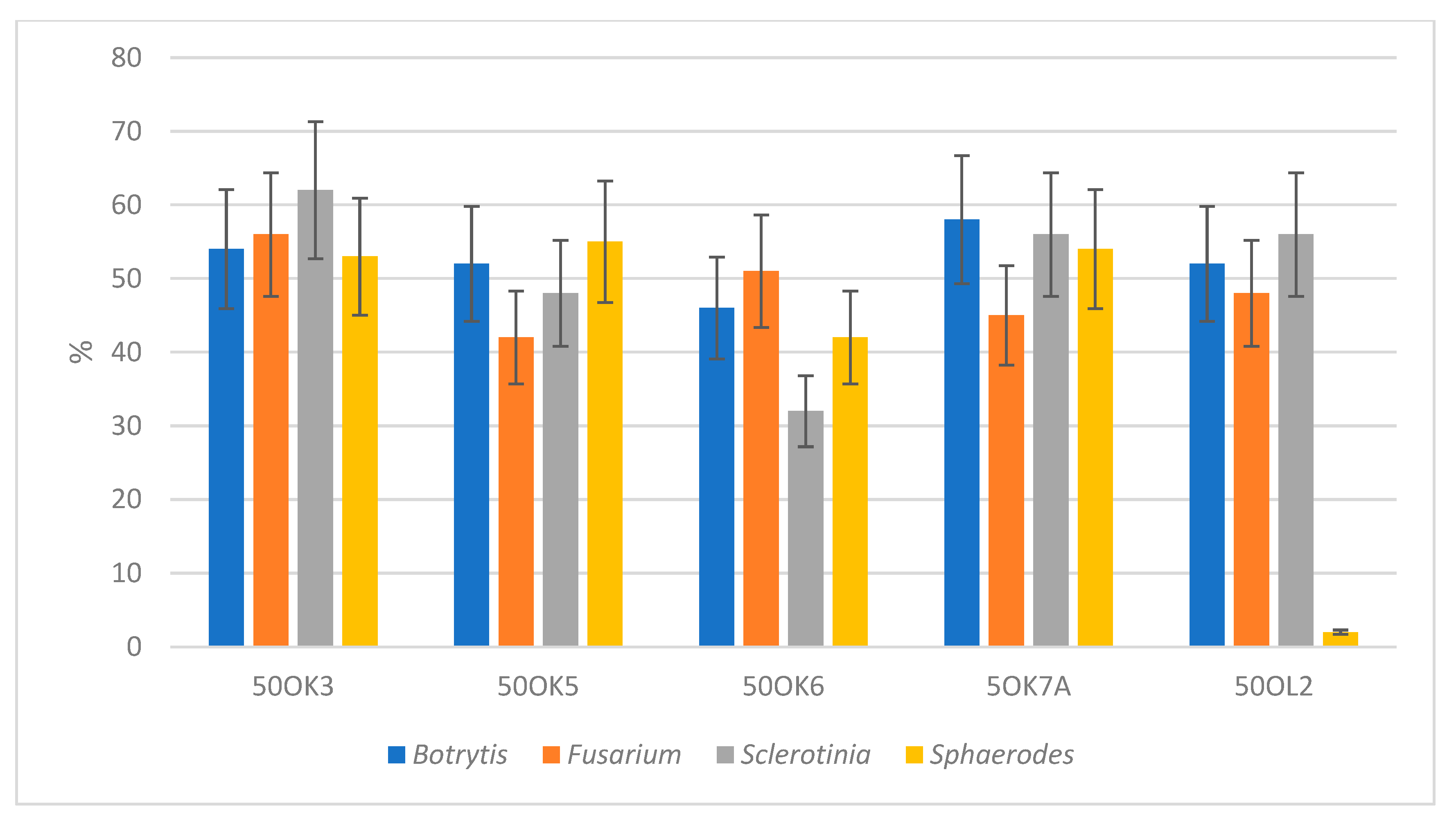
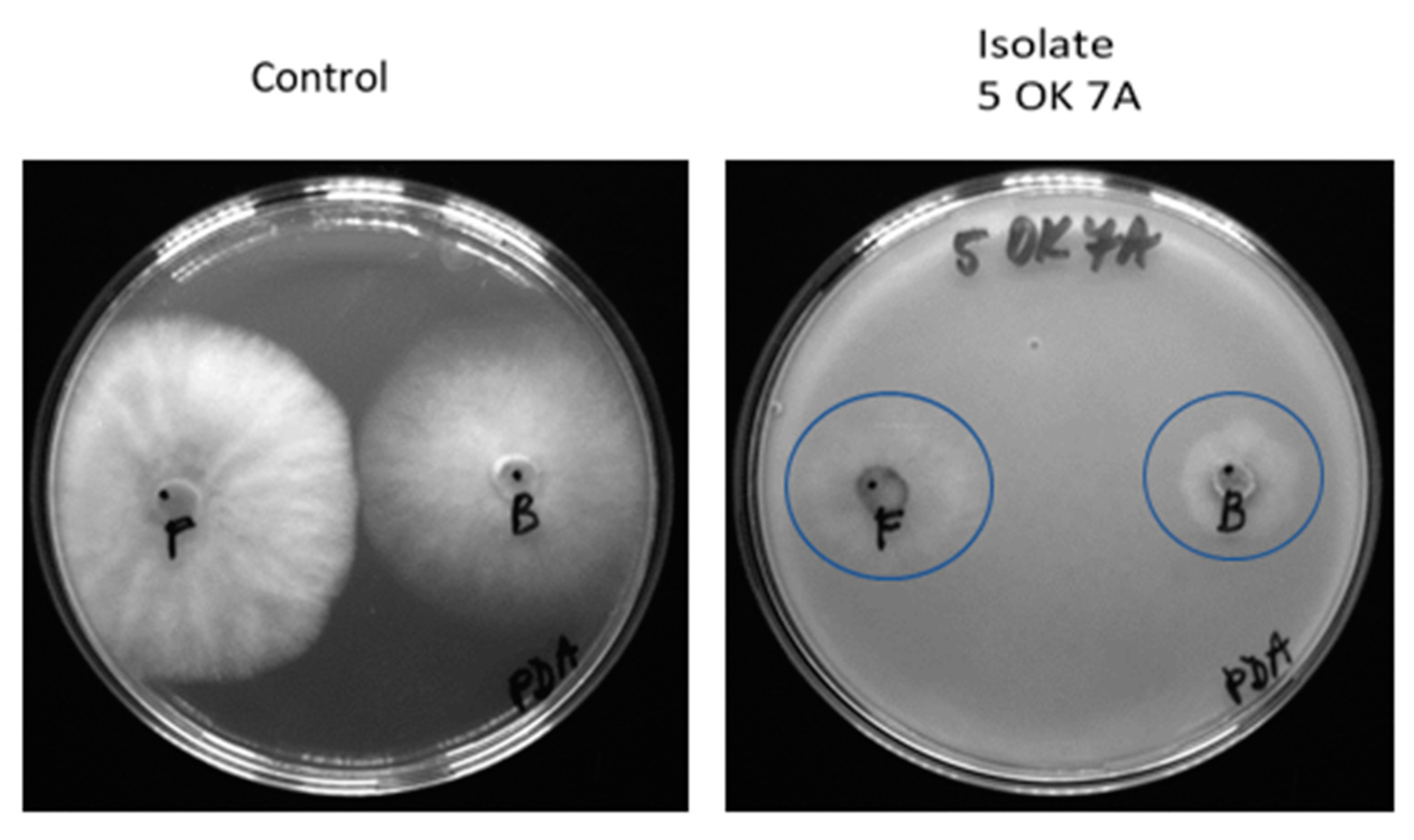
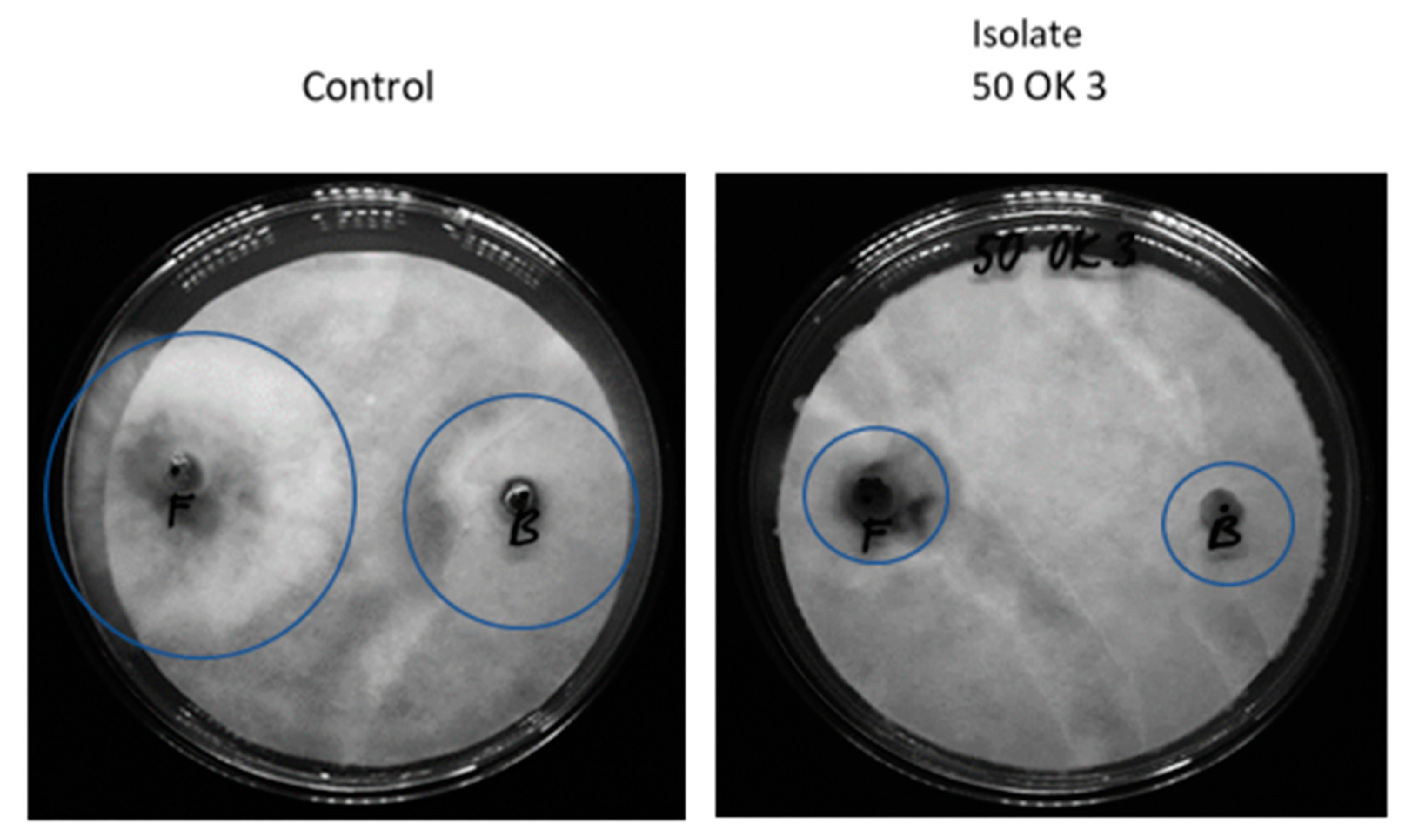
3.3. Antifungal Activity of Bacterial Isolates
3.4. Evaluation of Antifungal Activity of Immobilized Endophytic Isolates
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ouattara, M.S.; Laurent, A.; Berthou, M.; Borujerdi, E.; Butier, A.; Malvoisin, P.; Romelot, D.; Loyce, C. Identifying Factors Explaining Yield Variability of Miscanthus x giganteus and Miscanthus sinensis Across Contrasting Environments: Use of an Agronomic Diagnosis Approach. Bioenergy Res. 2022, 15, 672–685. [Google Scholar] [CrossRef]
- Hudson, A.O.; Ahmad, N.H.; Van Buren, R.; Savka, M.A. Sugarcane and Grapevine Endophytic Bacteria: Isolation, Detection of Quorum Sensing Signals and Identification by 16S v3 rDNA Sequence Analysis. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Formatex Research Center: Badajoz, Spain, 2011; pp. 801–806. [Google Scholar]
- Li, H.; Sharma, V.K.; Newcombe, G.; BarbosaTrivella, D.B.; Soni, R. Biotechnology application of bacterial endophytes in ariculture, environment and industry. Front. Microbiol. 2023, 17, 1269279. [Google Scholar]
- Malfanova, N.V. Endophytic Bacteria with Plant Growth Promoting and Biocontrol Abilities. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 2013. [Google Scholar]
- Reed, M.L.E.; Glick, B.R. Applications of plant growth-promoting bacteria for plant and soil systems. In Application of Microbial Engineering; Editor Gupta, V.K., Schmoll, M., Mazutti, M.A., Maki, M., Tuohy, M.G., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 181–228. [Google Scholar]
- Afzal, M.; Khan, Q.M.; Sessitsch, A. Endophytic bacteria: Prospects and applications for the phytoremediation of organic pollutants. Chemosphere 2014, 117, 232–242. [Google Scholar] [CrossRef]
- Kushwaha, P.; Kashyap, P.L.; Srivastava, A.K.; Tiwari, R.K. Plant growth promoting and antifungal activity in endophytic Bacillus strains from pearl millet (Pennisetum glaucum). Braz. J. Microbiol. 2020, 51, 229–241. [Google Scholar] [CrossRef]
- Gupta, G.; Panwar, J.; Akhtar, M.S.; Jha, P.N. Endophytic nitrogen-fixing bacteria as biofertilizer. Sustain. Agric. Rev. 2012, 11, 183–221. [Google Scholar]
- Rana, K.L.; Kour, D.; Kaur, T. Endophytic microbes: Biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Antonie Van Leeuwenhoek 2020, 113, 1075–1107. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Bhowmick, S.; Yadav, S.; Rashid, M.; Chouhan, G.; Vaishya, J.; Verma, J. Re-vitalizing of endophytic microbes for soil health management and plant protection. 3 Biotech. 2021, 11, 399. [Google Scholar] [CrossRef]
- Gunjal, A.; Waghmode, M.; Patil, N.; Kapadnis, B. Endophytes and Their Role in Phytoremediation and Biotransformation Process. In Microbial Biotechnology in Environmental Monitoring and Cleanup; IGI Global, Sharma, A., Eds.; Engineering Science Reference: Harshey, PA, USA, 2018; pp. 238–251. [Google Scholar]
- Khan, M.S.; Gao, J.; Chen, X.; Zhang, M.; Yang, F.; Du, Y.; Moe, T.S.; Munir, I.; Xue, J.; Zhang, X. The Endophytic Bacteria Bacillus velezensis Lle-9, Isolated from Lilium leucanthum, Harbors Antifungal Activity and Plant Growth-Promoting Effects. J. Microbiol. Biotechnol. 2020, 30, 668–680. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.S.; Dunfield, K.E. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Elucidating Mechanisms of Endophytes Used in Plant Protection and Other Bioactivities with Multifunctional Prospects. Front. Bioeng. Biotechnol. 2020, 8, 467–487. [Google Scholar] [CrossRef] [PubMed]
- Morales-Cedeno, L.R.; Orozco-Mosqueda, M.d.C.; Loeza-Lara, P.D.; Parra-Cota, F.I.; de los Santos-Villalobos, S.; Santoyo, G. Plant growth-promoting bacterial endophytes as biocontrol agents of pre- and post-harvest diseases: Fundamentals, methods of application and future perspectives. Microbiol. Res. 2021, 242, 126612. [Google Scholar] [CrossRef]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.A.; Hassan, S.E.-D. Plant Growth-Promoting Endophytic Bacterial Community Inhabiting the Leaves of Pulicaria incisa (Lam.) DC Inherent to Arid Regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.d.C.; Flores, A.; Rojas-Sánchez, B.; Urtis-Flores, C.A.; Morales-Cedeño, L.R.; Valencia-Marin, M.F.; Chávez-Avila, S.; Rojas-Solis, D.; Santoyo, G. Plant Growth-Promoting Bacteria as Bioinoculants: Attributes and Challenges for Sustainable Crop Improvement. Agronomy 2021, 11, 1167. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O.; Santoyo, G.; Perazzolli, M. The Potential Role of Microbial Biostimulants in the Amelioration of Climate Change-Associated Abiotic Stresses on Crops. Front. Microbiol. 2022, 12, 829099. [Google Scholar] [CrossRef]
- Schmidt, C.S.; Mrnka, L.; Frantík, T.; Lovecká, P.; Vosádka, M. Plant growth promotion of Miscanthus × giganteus by endophytic bacteria and fungi on non-polluted and polluted soils. World J. Microbiol. Biotechnol. 2018, 13, 48. [Google Scholar] [CrossRef]
- Koubek, J.; Uhlík, O.; Ječná, K.; Junková, P.; Vrkoslavová, J.; Lipov, J.; Kurzawová, V.; Macek, T.; Macková, M. Whole-cell MALDI-TOF: Rapid screening method in environmental microbiology. Int. Biodeterior. Biodegrad. 2012, 69, 82–86. [Google Scholar] [CrossRef]
- Li, Z.; Chang, S.; Lin, L.; Li, Y.; An, Q. A colorimetric assay of 1-aminocyclopropane-1-carboxylate (ACC) based on ninhydrin reaction for rapid screening of bacteria containing ACC deaminase. Lett. Appl. Microbiol. 2011, 53, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of Blue Agar CAS Assay for Siderophore Detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef]
- Jasim, B.; Jimtha, C.J.; Jyothis, M.; Radhakrishnan, E.K. Plant growth promoting potential of endophytic bacteria isolated from Piper nigrum. Plant Growth Regul. 2013, 71, 1–11. [Google Scholar] [CrossRef]
- Dadarwal, K.R.; Kundu, B.S.; Tauro, P. In vitro and in vivo nitrogenase activity of Rhizobium mutants and their symbiotic effectivity. J. Biosci. 1981, 3, 117–124. [Google Scholar] [CrossRef]
- Hajšlová, J.; Fenclová, M.; Zachariašová, M. Methodology for the Rapid Screening of Isolates of Endophytic Microorganisms and Identification of Strains with Phytohormonal Activity; UCT Prague: Prague, Czech Republic, 2013; ISBN 978-80-7080-869-6. (In Czech) [Google Scholar]
- Landa, B.B.; Hervás, A.; Bettiol, W.; Jiménez-Díaz, R.M. Antagonistic activity of Bacteria from the chickpea rhizosphere against Fusarium Oxysporum f. sp. Ciceris. Phytoparasitica 1997, 25, 305–318. [Google Scholar] [CrossRef]
- Manka, M.; Visconti, A.; Chełkowski, J.; Baottalico, A. Pathogenicity of Fusarium Isolates from Wheat, Rye and Triticale Towards Seedlings and their Ability to Produce Trichothecenes and Zearalenone. J. Phytopathol. 2008, 113, 24–29. [Google Scholar] [CrossRef]
- Loper, J.E.; Gross, H. Genomic analysis of antifungal metabolite production by Pseudomonas fluorescens Pf-5. Eur. J. Plant Pathol. 2007, 119, 265–278. [Google Scholar] [CrossRef]
- Coutinho, T.A.; Venter, S.N. Pantoea ananatis: An unconventional plant pathogen. Mol. Plant Pathol. 2009, 10, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Scales, B.S.; Dickson, R.P.; LiPuma, J.J.; Huffnagle, G.B. Microbiology, genomics, and clinical significance of the Pseudomonas fluorescens species complex, an unappreciated colonizer of humans. Clin. Microbiol. Rev. 2014, 27, 927–948. [Google Scholar] [CrossRef] [PubMed]
- Johnston-Monje, D.; Raizada, M.N. Conservation and diversity of seed associated endophytes in Zea across boundaries of evolution, ethnography and ecology. PLoS ONE 2011, 6, e20396. [Google Scholar] [CrossRef]
- Verma, P.; Yadav, A.N.; Kazy, S.K.; Saxena, A.K.; Suman, A. Evaluating the diversity and phylogeny of plant growth promoting bacteria associated with wheat (Triticum aestivum) growing in central zone of India. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 432–447. [Google Scholar]
- Zhang, Y.; He, L.; Chen, Z.; Wang, Q.; Qian, M.; Sheng, X. Characterization of ACC deaminase-producing endophytic bacteria isolated from copper-tolerant plants and their potential in promoting the growth and copper accumulation of Brassica napus. Chemosphere 2011, 83, 57–62. [Google Scholar] [CrossRef]
- Trivedi, P.; Spann, T.; Wang, N. Isolation and characterization of beneficial bacteria associated with citrus roots in Florida. Microb. Ecol. 2011, 62, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Borisov, A.Y.; Tikhonovich, I.A.; Kluge, C.; Preisfeld, A.; et al. Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2011, 47, 642–652. [Google Scholar] [CrossRef]
- Oliveira, C.A.; Alves, V.M.C.; Marriel, I.E.; Gomes, E.A.; Scotti, M.R.; Carneiro, N.P.; Guimaraes, C.T.; Schaffert, R.E.; Sá, N.M.H. Phosphate solubilizing microorganisms isolated from rhizosphere of maize cultivated in an oxisol of the Brazilian Cerrado Biome. Soil Biol. Biochem. 2009, 41, 1782–1787. [Google Scholar] [CrossRef]
- Cornelis, P. Iron uptake and metabolism in pseudomonads. Appl. Microbiol. Biotechnol. 2010, 86, 1637–1645. [Google Scholar] [CrossRef]
- Chen, L.; Dodd, I.C.; Theobald, J.C.; Belimov, A.A.; Davies, W.J. The rhizobacterium Variovorax paradoxus 5C-2, containing ACC deaminase, promotes growth and development of Arabidopsis thaliana via an ethylene-dependent pathway. J. Exp. Bot. 2013, 64, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Flores-Duarte, N.J.; Pérez-Pérez, J.; Navarro-Torre, S.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Improved Medicago sativa Nodulation under Stress Assisted by Variovorax sp. Endophytes. Plants 2022, 11, 1091. [Google Scholar] [CrossRef]
- Méndez, M.O.; Maier, R.M. Phytoestabilisation of mine tailings in arid and semiarid environments: An emerging remediation technology. Environmetal Health Perspecives 2008, 116, 278–283. [Google Scholar] [CrossRef]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Kido, K.; Hasegawa, M.; Matsumoto, H.; Kobayashi, M.; Takikawa, Y. Pantoea ananatis strains are differentiated into three groups based on reactions of tobacco and welsh onion and on genetic characteristics. J. Gen. Plant Pathol. 2010, 76, 208–218. [Google Scholar] [CrossRef]
- Pérez-Martínez, I.; Zhao, Y.; Murillo, J.; Sundin, G.W.; Ramos, C. Global genomic analysis of Pseudomonas savastanoi pv. savastanoi plasmids. J. Bacteriol. 2008, 190, 625–635. [Google Scholar] [PubMed]
- Kochar, M.; Upadhyay, A.; Srivastava, S. Indole-3-acetic acid biosynthesis in the biocontrol strain Pseudomonas fluorescens Psd and plant growth regulation by hormone overexpression. Res. Microbiol. 2011, 162, 426–435. [Google Scholar] [CrossRef]
- Jha, P.N.; Gupta, G.; Jha, P.; Mehrotra, R. Association of rhizospheric/endophytic bacteria with plants: A potential gateway to sustainable agriculture. Greener J. Agric. Sci. 2013, 3, 73–84. [Google Scholar]
- Stewart, A.; Cromey, M. Identifying disease threats and management practices for bio-energy crops. Curr. Opin. Environ. Sustain. 2011, 3, 75–80. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates from Plant Tissue | Identification | ACCd | P-SOL | SID | NITRO |
|---|---|---|---|---|---|
| 50 OK 3 | Pseudomonas libanensis | - | +++ | + | +++ |
| 50 OK 5 | Variovorax paradoxus | - | - | - | ++ |
| 50 OK 6 | Pseudomonas marginalis | - | + | +++ | ++ |
| 5 OK 7A | Pseudomonas libanensis | - | +++ | + | ++ |
| 50 OL 2 | Pantoea ananatis | + | ++ | +++ | + |
| Isolates from Plant Tissue | Identification | Phytohormones (ng·mL−1) | ||
|---|---|---|---|---|
| iP | iPR | IAA | ||
| 50 OK 3 | Pseudomonas libanensis | 5.5 | n.d. | n.d. |
| 50 OK 5 | Variovorax paradoxus | 3.8 | n.d. | n.d. |
| 50 OK 6 | Pseudomonas marginalis | 3.7 | n.d. | n.d. |
| 5 OK 7A | Pseudomonas libanensis | 11.7 | 8.9 | 12.6 |
| 50 OL 2 | Pantoea ananatis | 1.1 | n.d. | n.d. |
| Isolates from Plant Tissue | Identification | Botrytis cinerea | Fusarium sporotrichioides | Sclerotinia sclerotiniorum | Sphaerodes fimicola |
|---|---|---|---|---|---|
| 50 OK 3 | Pseudomonas libanensis | + | + | + | + |
| 50 OK 5 | Variovorax paradoxus | + | + | + | + |
| 50 OK 6 | Pseudomonas marginalis | + | + | + | + |
| 5 OK 7A | Pseudomonas libanensis | + | + | + | + |
| 50 OL 2 | Pantoea ananatis | + | + | + | + |
| Isolates from Plant Tissue | Identification | Botrytis cinerea | Fusarium sporotrichioides | Sclerotinia sclerotiniorum | Sphaerodes fimicola |
|---|---|---|---|---|---|
| 50 OK 3 | Pseudomonas libanensis | + | + | + | + |
| 50 OK 5 | Variovorax paradoxus | + | - | + | + |
| 50 OK 6 | Pseudomonas marginalis | + | + | - | + |
| 5 OK 7A | Pseudomonas libanensis | + | + | + | + |
| 50 OL 2 | Pantoea ananatis | + | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lovecká, P.; Kroneislová, G.; Novotná, Z.; Röderová, J.; Demnerová, K. Plant Growth-Promoting Endophytic Bacteria Isolated from Miscanthus giganteus and Their Antifungal Activity. Microorganisms 2023, 11, 2710. https://doi.org/10.3390/microorganisms11112710
Lovecká P, Kroneislová G, Novotná Z, Röderová J, Demnerová K. Plant Growth-Promoting Endophytic Bacteria Isolated from Miscanthus giganteus and Their Antifungal Activity. Microorganisms. 2023; 11(11):2710. https://doi.org/10.3390/microorganisms11112710
Chicago/Turabian StyleLovecká, Petra, Gabriela Kroneislová, Zuzana Novotná, Jana Röderová, and Kateřina Demnerová. 2023. "Plant Growth-Promoting Endophytic Bacteria Isolated from Miscanthus giganteus and Their Antifungal Activity" Microorganisms 11, no. 11: 2710. https://doi.org/10.3390/microorganisms11112710