
Factors Influencing the Production of Extracellular Polysaccharides by the Green Algae Dictyosphaerium chlorelloides and Their Isolation, Purification, and Composition

Abstract
:1. Introduction
2. Materials and Methods
2.1. The Strain
2.2. Chemicals
2.3. Instruments
2.4. Vertical Column Photobioreactor (300 mL)
2.5. Airlift Column Photobioreactor (60 L)
2.6. Growth Medium
2.7. Biomass Cell Concentration Measurement
2.8. Quantification of Extracellular Polysaccharides
2.9. Viscosity Measurement
2.10. Staining of Extracellular Polysaccharides
2.11. Salinity Stress
2.12. Separation of Cells from the Culture Medium
2.13. Purification of Extracellular Polysaccharides
2.14. Determination of Extracellular Polysaccharides Composition
2.15. Statistical Analysis
3. Results
3.1. Laboratory Scale 300 mL Column Photobioreactors—Optimization of Cultivation Medium for Algal EPS Production
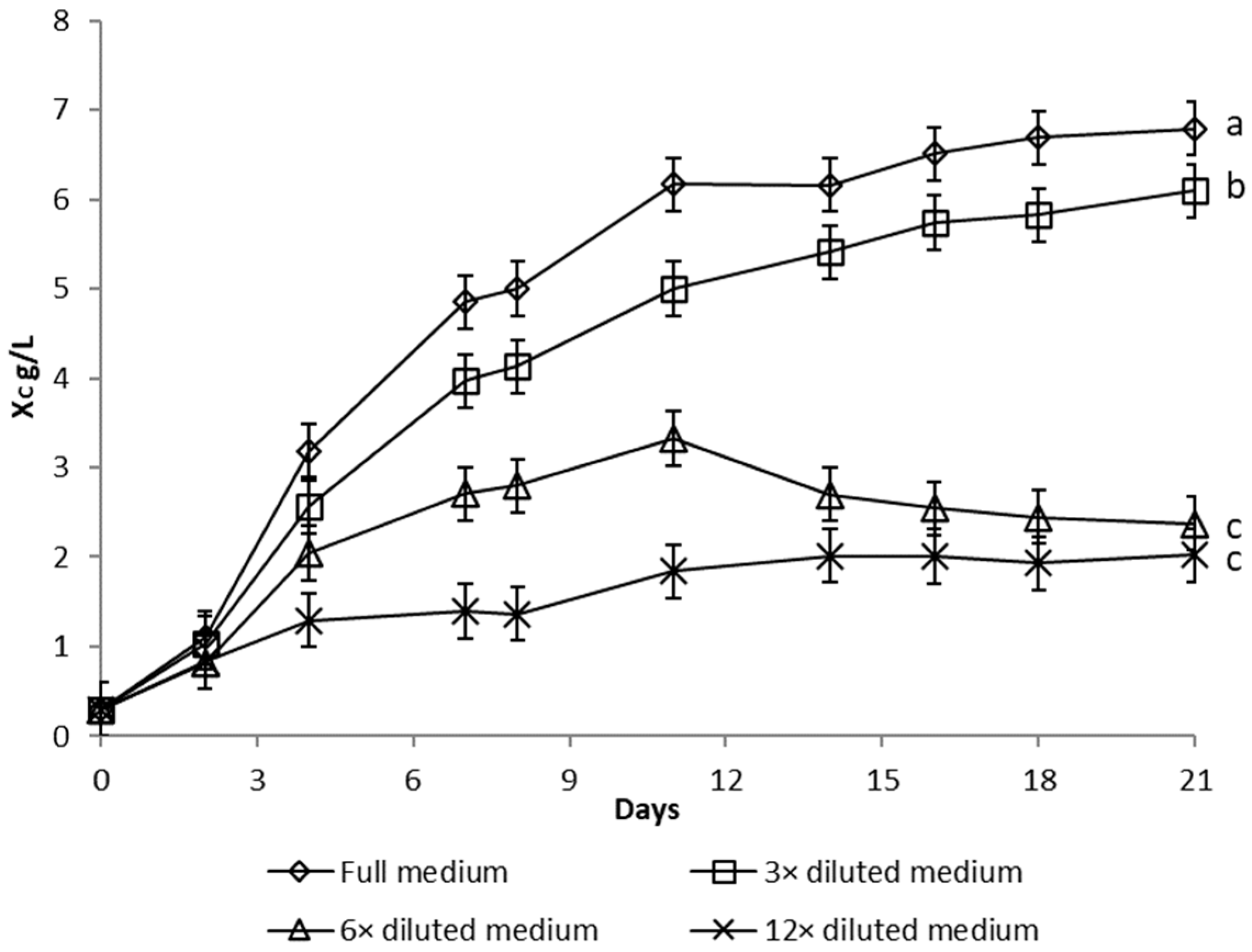
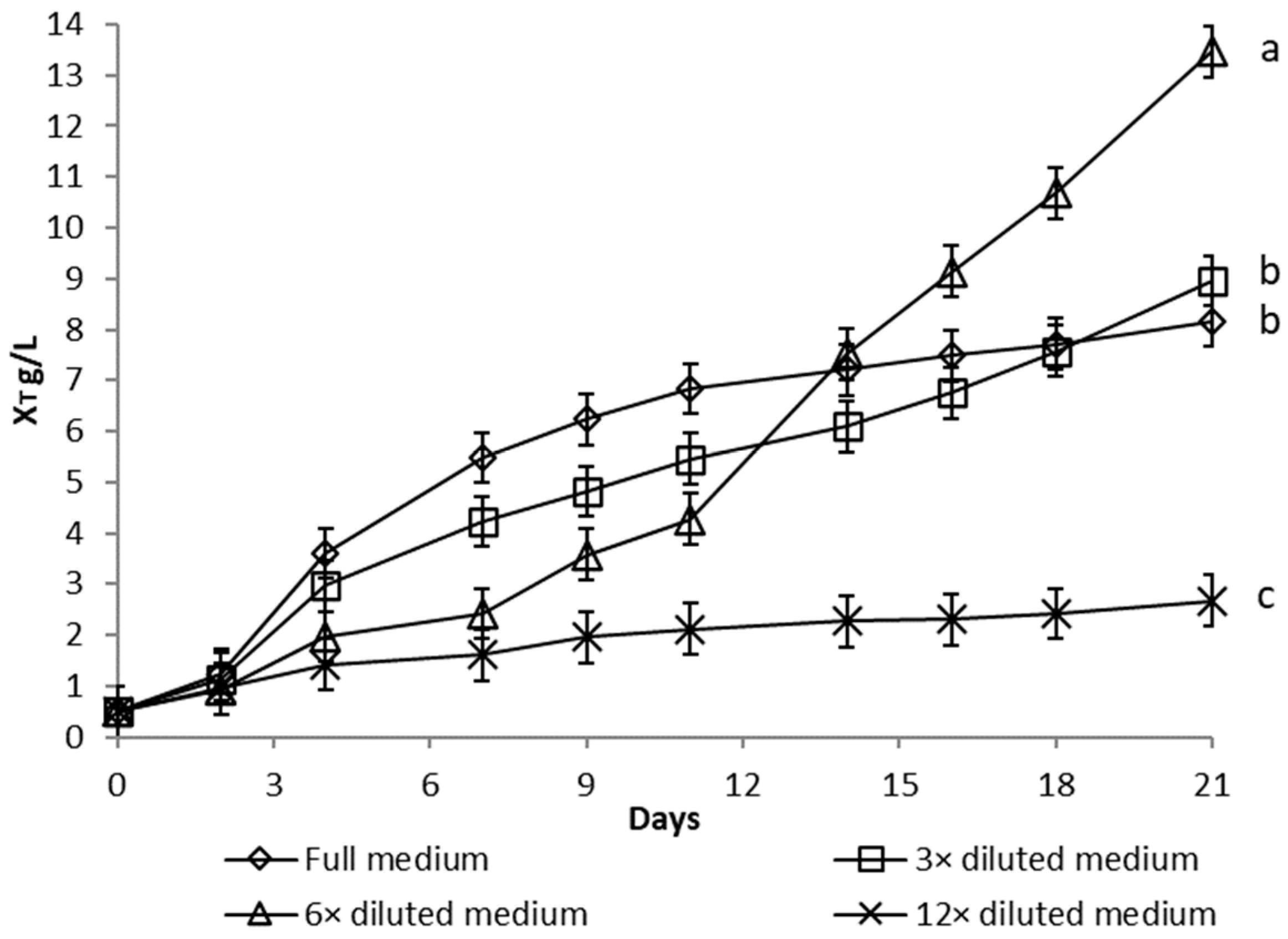
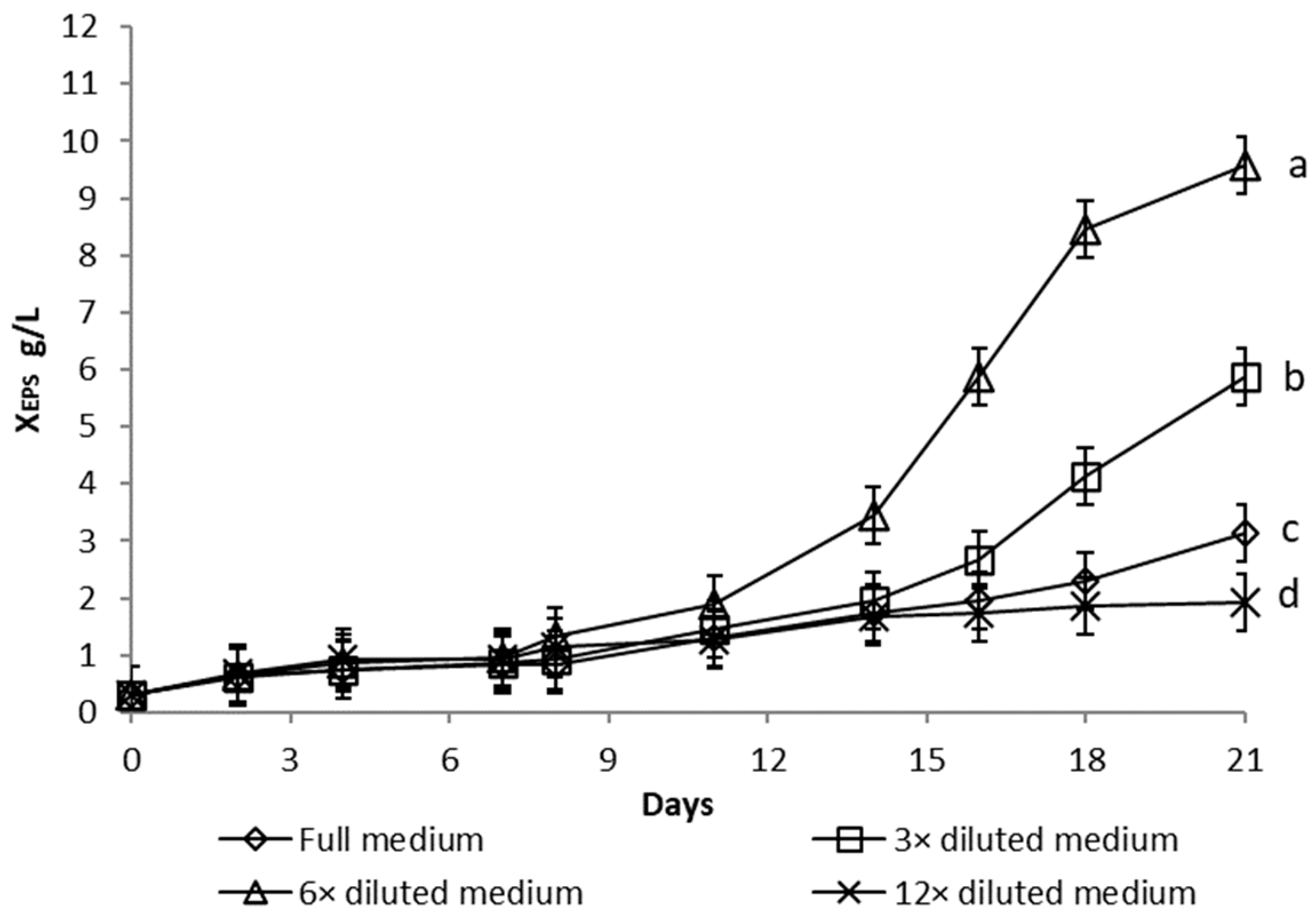
3.1.1. Nutrient Limitation
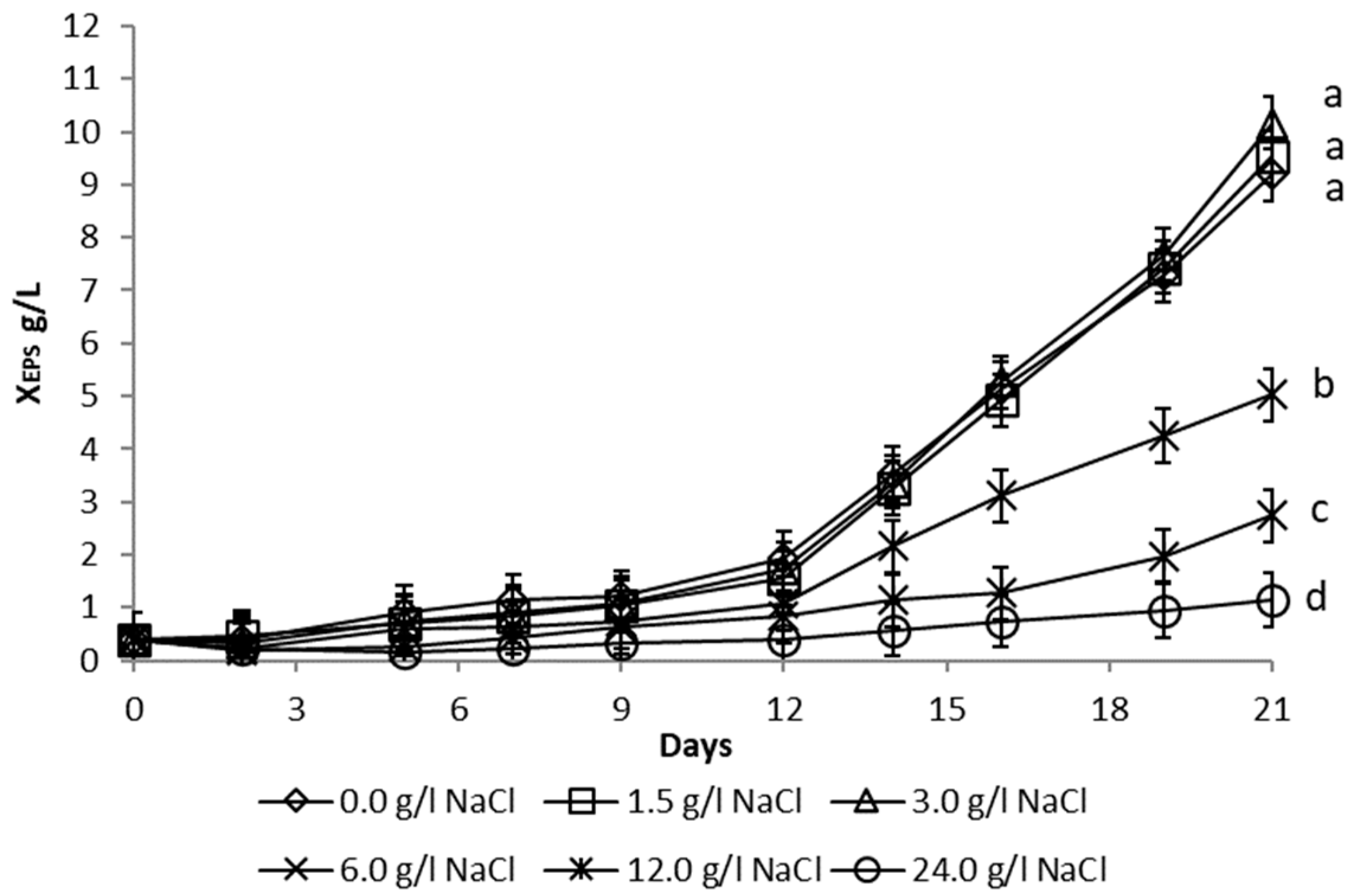
3.1.2. Salinity Stress
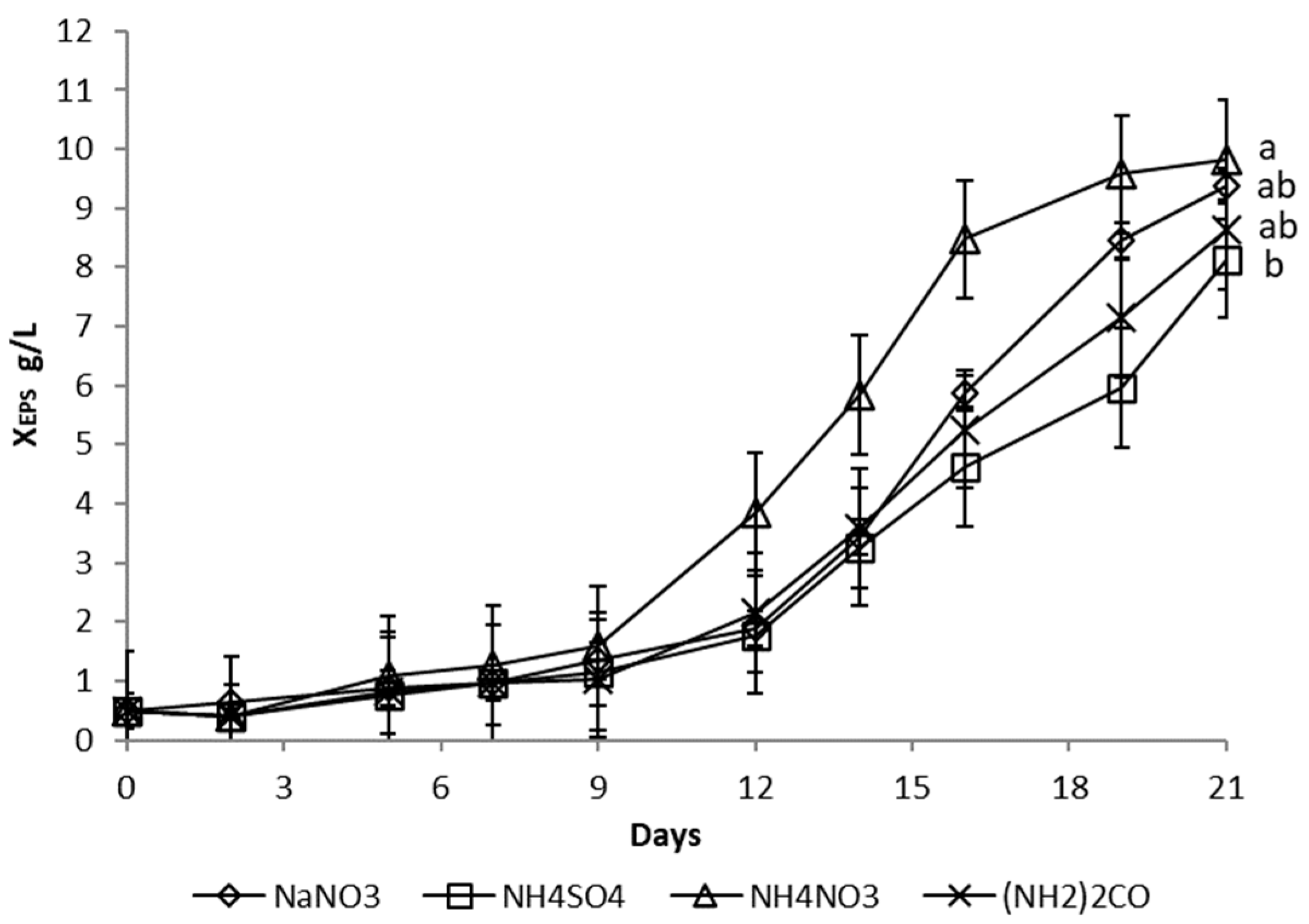
3.1.3. Nitrogen Source
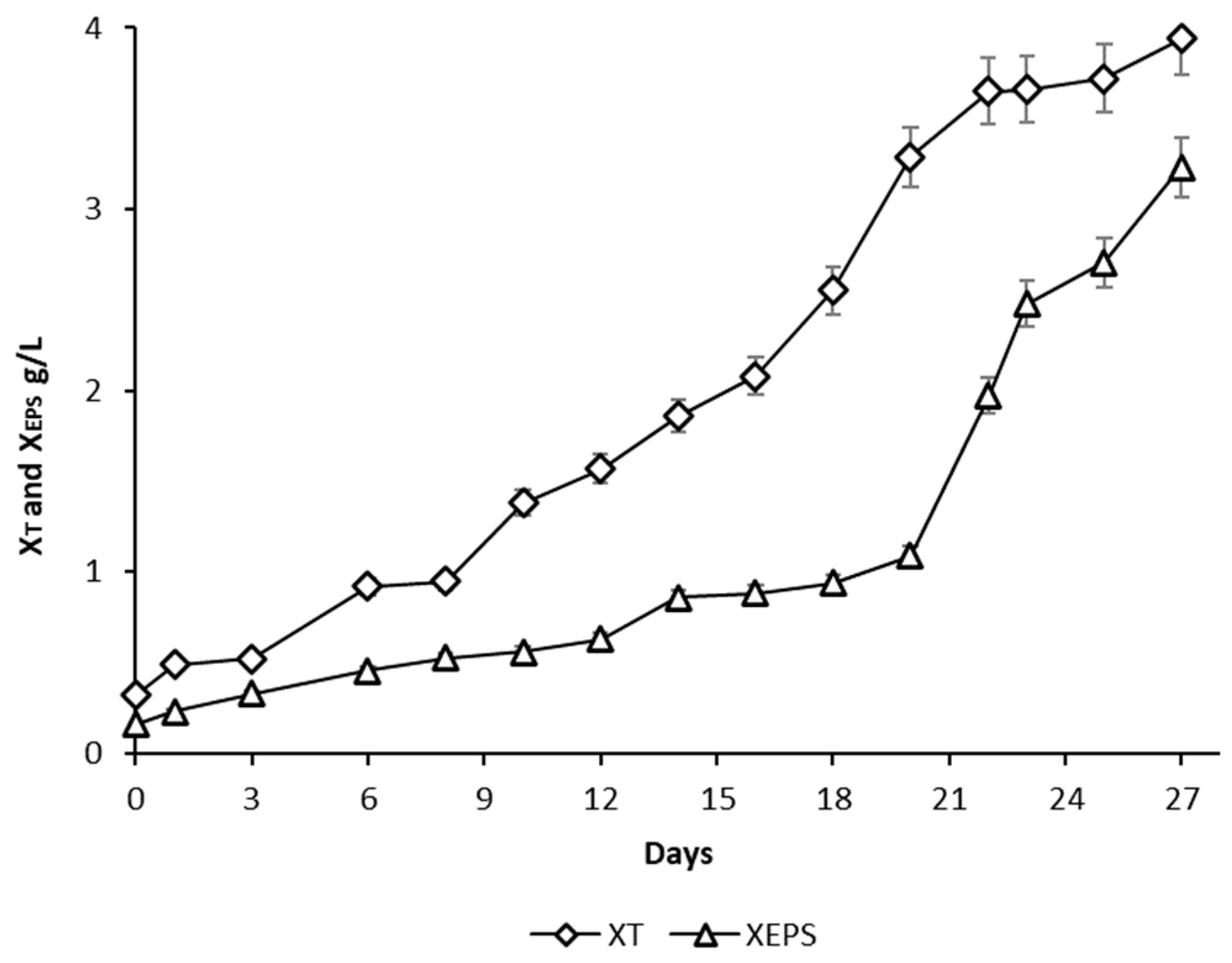
3.2. Algal Cultivation in 60 L Photobioreactor
3.3. Separation of EPS from Algae Cells
3.4. Purification of EPS
3.5. Determination of EPS Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ginzberg, A.; Cohen, M.; Sod-Moriah, A.; Shany, S.; Rosenshtrauch, A.; Arad, M. Chickens fed with biomass of the red microalga Porphyridium sp. have reduced blood cholesterol levels and modified fatty acids composition in egg yolk. J. Appl. Phycol. 2000, 12, 325–330. [Google Scholar] [CrossRef]
- Guzman, S.; Gato, A.; Lamela, M.; Freire-Garabal, M.; Calleja, M. Anti-Inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Hayashi, T.; Kojima, I. A natural sulphated polysaccharide, calcium spirulan, isolated from Spirulina platensis: In vitro and ex vivo evaluation of anti-herpes simplex virus and anti-human immunodeficiency virus. AIDS Res. Hum. Retrovir. 1996, 12, 1463–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majee, S.B.; Avlani, D.; Biswas, G.R. Pharmacological, pharmaceutical, cosmetic and diagnostic applications of sulphated polysaccharides from marine algae and bacteria. Afr. J. Pharm. Pharmacol. 2017, 11, 68–77. [Google Scholar]
- Pejin, B.; Tešanović, K.; Jakovljević, D.; Kaišarević, S.; Šibul, F.; Rašeta, M.; Karaman, M. The polysaccharide extracts from the fungi Coprinus comatus and Coprinellus truncorum do exhibit AChE inhibitory activity. Nat. Prod. Res. 2019, 33, 750–754. [Google Scholar] [CrossRef]
- Huang, J.; Liu, L.; Yu, Y.; Lin, W.; Chen, B.; Li, M. Reduction in the blood glucose level of exopolysaccharide of Porphyridium cruentum in alloxan-induced diabetic mice. J. Fujian Norm. Univ. 2006, 22, 77–80. [Google Scholar]
- Kanekiyo, K.; Lee, B.; Hayashi, K.; Takenaka, H.; Hayakawa, Y.; Endo, S.; Hayashi, T. Isolation of an antiviral polysaccharide, nostoflan, from a terrestrial cyanobacterium, Nostoc flagelliforme. J. Nat. Prod. 2005, 68, 1037–1041. [Google Scholar] [CrossRef]
- Karaman, M.; Janjušević, L.; Jakovljević, D.; Šibul, F.; Boris Pejin, B. Anti-hydroxyl radical activity, redox potential and anti-AChE activity of Amanita strobiliformis polysaccharide extract. Nat. Prod. Res. 2019, 33, 1522–1526. [Google Scholar] [CrossRef]
- Kim, M.; Yim, J.; Kim, S.; Kim, H.; Lee, W.; Kim, S.; Kang, P.; Lee, C. In vitro inhibition of influenza A virus infection by marine microalga-derived sulphated polysaccharide p-KG03. Antivir. Res. 2012, 93, 253–259. [Google Scholar] [CrossRef]
- Angelis, S.; Novak, A.; Sydney, E.; Soccol, V.; Carvalho, J.; Pandey, A.; Noseda, M.; Tholozan, J.; Lorquin, J.; Soccol, C. Co-Culture of Microalgae, Cyanobacteria, and Macromycetes for Exopolysaccharides Production: Process Preliminary Optimization and Partial Characterization. Appl. Biochem. Biotechnol. 2012, 167, 1092–1106. [Google Scholar] [CrossRef] [Green Version]
- Arad, M.; Friedman, O.; Rotem, A. Effect of nitrogen on polysaccharide production in a Porphyridium sp. Appl. Environ. Microbiol. 1988, 54, 2411–2414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arad, M.; Lerental, Y.; Dubinsky, O. Effect of nitrate and sulphate starvation on polysaccharide formation Rhodella reticulata. Bioresour. Technol. 1992, 42, 141–148. [Google Scholar] [CrossRef]
- Bafana, A. Characterization and optimization of production of exopolysaccharide from Chlamydomonas reinhardtii. Carbohydr. Polym. 2013, 95, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.C.; Mann, R. Physiological Aspects in Algal Mass Cultures. In Algal Biomass; Shelef, G., Soeder, C.J., Eds.; Elsevier/North-Holland Biomedical Press: Amsterdam, The Netherlands, 1989; pp. 343–353. [Google Scholar]
- Penna, A.; Berluti, S.; Penna, N.; Magnani, M. Influence of nutrient ratios on the in vitro extracellular polysaccharide production by marine diatoms from Adriatic Sea. J. Plankton Res. 1999, 21, 1681–1690. [Google Scholar] [CrossRef] [Green Version]
- Buetow, D.E. Growth, survival and biochemical alteration of Euglena gracilis in medium limited in sulfur. J. Cell Comp. Physiol. 1965, 66, 235–242. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Sili, C.; Tassinato, G.; Vincenzini, M.; Materassi, R. Effects of growth conditions on exopolysaccharide production by Cyanospira capsulata. Bioresour. Technol. 1991, 38, 101–104. [Google Scholar] [CrossRef]
- Richmond, A. Environmental Stress Physiology. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Richmond, A., Ed.; Blackwell Publishing Company: Hoboken, NJ, USA, 2004; pp. 57–82. [Google Scholar]
- Singh, A.; Nigam, P.S.; Murphy, J.D. Mechanism and challenges in the commercialization of algal biofuels. Bioresour. Technol. 2011, 102, 26–34. [Google Scholar] [CrossRef]
- Giroldo, D.; Vieira, H. An extracellular sulphated fucfucose-richlysaccharides produced by a tropical strain of C. obovata (Cryptophyceae). J. Appl. Phycol. 2002, 14, 185–191. [Google Scholar] [CrossRef]
- Dubinsky, O.; Barak, Z.; Geresh, S.; Arad, M. Composition of the cell-wall polysaccharide of the unicellular red alga Rhodella reticulata at two phases of growth. Recent Adv. Algal Biotechnol. 1990, 5, 17. [Google Scholar]
- Garozzo, D.; Impallomeni, G.; Spina, E.; Sturiale, L. The structure of the exocellular polysaccharide from the cyanobacterium Cyanospira capsulata. Carbohydr. Res. 1998, 307, 113–124. [Google Scholar] [CrossRef]
- Geresh, S.; Lupescu, N.; Arad, M. Fractionation and partial characterization of the sulphated polysaccharide of Porphyridium. Phytochemistry 1992, 31, 4181–4186. [Google Scholar] [CrossRef]
- Heaney, J.; Chapman, J. Structural studies on the extracellular polysaccharide of the red alga, Porphyridium cruentum. Carbohydr. Res. 1976, 52, 169–177. [Google Scholar] [CrossRef]
- Arad, M.; Levy, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, J.; Valdez, M.; Garcia, S.; Vazquez, S. Effect of climatic variation on the relative abundance of red alga Gelidium robustus in Baja California Sur, Mexico. J. Appl. Phycol. 2000, 177, 177–188. [Google Scholar] [CrossRef]
- Tsarenko, P.M.; Wasser, S.P. Trebouxiophyceae. In Algae of Ukraine: Diversity, Nomenclature, Taxonomy, Ecology and Geography; Tsarenko, P.M., Wasser, S.P., Nevo, E., Eds.; A.R. Gantner Verlag KG: Ruggell, Liechtenstein, 2011; pp. 61–108. [Google Scholar]
- Rios, S.D.; Torres, C.M.; Torras, C.; Salvado, J.; Mateo-Sanz, J.M.; Jimenez, L. Microalgae-based biodiesel: Economic analysis of downstream process realistic scenarios. Bioresour. Technol. 2013, 136, 617–625. [Google Scholar] [CrossRef]
- Stack, K.; Lee, R.; Richardson, D.; Lewis, T.; Garnier, G. Complex formation and stability of colloidal wood resin pitch suspensions with hemicellulose polymers. Colloids Surf. A 2014, 441, 101–108. [Google Scholar] [CrossRef]
- Liu, J.; Tao, Y.; Wua, J.; Zhu, Y.; Gao, B.; Tang, Y.; Li, A.; Zhang, C.; Zhang, Y. Effective flocculation of target microalgae with self-flocculating microalgae induced by pH decrease. Bioresour. Technol. 2014, 167, 367–375. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, F.; Wang, X.; Liu, X.; Hou, Y.; Zhang, Q. Extraction of the polysaccharides from five algae and their potential antioxidant activity in vitro. Carbohydr. Polym. 2010, 82, 118–121. [Google Scholar] [CrossRef]
- Chakraborty, M.; Miao, C.; McDonald, A.; Chen, S. Concomitant extraction of bio-oil and value added polysaccharides from Chlorella sorokiniana using a unique sequential hydrothermal extraction technology. Fuel 2011, 95, 63–70. [Google Scholar] [CrossRef]
- Mishra, A.; Jha, B. Isolation and characterization of extracellular polymeric substances from microalgae Dunaliella salina under salt stress. Bioresour. Technol. 2009, 100, 3382–3386. [Google Scholar] [CrossRef]
- Ye, H.; Zhou, C.H.; Bian, F. Study on degreasing of Sargassum sp. by super- critical carbon dioxide fluid. Sci. Technol. Food Ind. 2006, 27, 136–137. [Google Scholar]
- Ye, H.; Wang, K.; Zhou, C.; Liu, J.; Zeng, X. Purification, antitumor and anti-oxidant activities in vitro of polysaccharides from the brown seaweed Sargassum pallidum. Food Chem. 2008, 111, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Yoon, W.J.; Kim, K.N.; Ahn, G.N.; Kang, S.M.; Kang, D.H.; Affan, A.; Oh, C.; Jung, W.K.; Jeon, Y.J. Evaluation of the anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Gloaugen, V.; Ruiz, G.; Morvan, H.; Mouradi-Givernaud, A.; Maes, E.; Krausz, P.; Strecker, G. The extracellular polysaccharide of Porphyridium sp.: An NMR study of lithium-resistant oligosaccharidic fragments. Carbohydr. Res. 2004, 339, 97–103. [Google Scholar] [CrossRef]
- Wang, S.C.; Bligh, S.W.; Shi, S.S.; Wang, Z.T.; Hu, Z.B.; Crowder, J.; Branford-White, C.; Vella, C. Structural features and anti-HIV-1 activity of novel polysaccharides from red algae Grateloupia longifolia and Grateloupia filicina. Int. J. Biol. Macromol. 2007, 41, 369–375. [Google Scholar] [CrossRef]
- Patel, A.K.; Laroche, C.; Marcati, A.; Ursu, A.V.; Jubeau, S.; Marchal, L.; Petit, E.; Djelveh, G.; Michaud, P. Separation and fractionation of exopolysaccharides from Porphyridium cruentum. Bioresour. Technol. 2013, 145, 345–350. [Google Scholar] [CrossRef]
- Elain, A.; Nkounkou, C.; Le Fellic, M.; Donnart, K. Green extraction of polysaccharides from Arthrospira platensis using high pressure homogenization. J. Appl. Phycol. 2020, 32, 1719–1727. [Google Scholar] [CrossRef]
- Zhang, R.; Grimi, N.; Marchal, L.; Lebovka, N.; Vorobiev, E. Effect of ultrasonication, high pressure homogenization and their combination on the efficiency of extraction of bio-molecules from microalgae Parachlorella kessleri. Algal Res. 2019, 40, 101524. [Google Scholar] [CrossRef]
- Pugh, N.; Ross, S.A.; El Sohly, H.N.; El Sohly, M.A.; Pasco, D.S. Isolation of Three High Molecular Weight Polysaccharide Preparations with Potent Immunostimulatory Activity from Spirulina platensis, Aphanizomenon flos-aquae and Chlorella pyrenoidosa. Planta Med. 2001, 67, 737–742. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Bišová, K.; Kawano, S.; Cepák, V.; Zachleder, V.; Čížková, M.; Brányiková, I.; Vítová, M. The microalga Parachlorella kessleri–A novel highly efficient lipid producer. Biotechnol. Bioeng. 2012, 110, 97–107. [Google Scholar] [CrossRef]
- Moheimani, N.R.; Borowitzka, M.A.; Isdepsky, A.; Sing, S.F. Standard Methods for Measuring Growth of Algae and Their Composition. Algae Biofuels Energy 2013, 5, 265–284. [Google Scholar]
- Chen, F. High cell density culture of microalgae in heterotrophic growth. Trends Biotechnol. 1996, 14, 421–426. [Google Scholar] [CrossRef]
- Lívanský, K.; Doucha, J. Production of high-density Chlorella culture grown in fermenters. J. Appl. Phycol. 2011, 24, 35–43. [Google Scholar]
- Branyikova, I.; Bisova, K.; Branyik, T.; Doucha, J.; Marsalkova, B.; Vitova, M.; Zachleder, V. Microalgae—Novel Highly Efficient Starch Producers. Biotechnol. Bioeng. 2011, 108, 766–776. [Google Scholar] [CrossRef]
- IHC World Alcian Blue Protocol for Staining. 2013. Available online: http://www.ihcworld.com/_protocols/special_stains/alcian_blue_ellis.htm (accessed on 1 July 2022).
- Baeva, E.; Bleha, R.; Lavrova, E.; Sushytskyi, L.; Čopíková, J.; Jablonsky, I.; Klouček, P.; Synytsya, A. Polysaccharides from Basidiocarps of Cultivating Mushroom Pleurotus ostreatus: Isolation and Structural Characterization. Molecules 2019, 24, 2740. [Google Scholar] [CrossRef] [Green Version]
- Kraan, S. Algal Polysaccharides, Novel Applications, and Outlook. In Carbohydrates Comprehensive Studies on Glycobiology and Glycotechnology; Chang, C.-F., Ed.; Intech: London, UK, 2012; pp. 505–515. [Google Scholar]
- Soanen, N.; Da Silva, E.; Gardarin, C.; Michaud, P.; Laroche, C. Improvement of exopolysaccharide production by Porphyridium marinum. Bioresour. Technol. 2016, 213, 231–238. [Google Scholar] [CrossRef]
- Yang, Z.; Ma, Y.; Zheng, J.; Yang, W.; Liu, J.; Li, H. Proteomics to reveal metabolic network shifts towards lipid accumulation following nitrogen deprivation in the diatom Phaeodactylum tricornutum. J. Appl. Phycol. 2014, 26, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Gilmour, D.; Hipkins, M.; Boney, A. The effect of osmotic and ionic stress on the primary processes of photosynthesis in Dunaliella tertiolecta. Planta 1984, 163, 250–256. [Google Scholar] [CrossRef]
- Gonzalez, A.; Tabemero, A.; Sanchez, J.M.; Martin del Valle, E.M.; Galan, M.A. Effect of nitrogen source on growth and lipid accumulation in Scenedesmus abundance and Chlorella ellipsoidea. Bioresour. Technol. 2014, 173, 334–341. [Google Scholar] [CrossRef]
- Lupi, F.M.; Fernandes, H.M.L.; Tomé, M.M.; Sá-Correia, I.; Novais, J.M. Influence of nitrogen source and photoperiod on exopolysaccharide synthesis by the microalga Botryococcus braunii UC 58. Enzyme Microb. Technol. 1994, 16, 546–550. [Google Scholar] [CrossRef]
- Rajasri, Y.; Ramgopal Rao, S.; Rao, C.S. Lipid accumulation studies in Chlorella pyrenoidosa using customized photobioreactor-effect of nitrogen source, light intensity and mode of operation. Int. J. Eng. Res. Appl. 2012, 2, 2446–2453. [Google Scholar]
- Singh, S.; Arad, S.; Richmond, A. Extracellular polysaccharide production in outdoor mass cultures of Porphyridium sp. in flat plate glass reactors. J. Appl. Phycol. 2000, 12, 269–275. [Google Scholar] [CrossRef]
- Contipro Inc. (Czech Republic). Official Website as of 8 May 2022. Available online: https://www.contipro.com/portfolio/manufacturer-of-pharmaceutical-sodium-hyaluronate (accessed on 1 July 2022).
- Lubrizol Corporation Inc. (USA). Official Website as of 8 May 2022. Available online: https://www.lubrizol.com/Personal-Care/Products/Product-Finder/Products-Data/Kelco-Care-diutan-gum (accessed on 1 July 2022).
- Dhamodharan, P.; Ponnusamy, N.; Odumpatta, R.; Lulu, S.; Arumugam, M. Computational investigation of marine bioactive compounds against E6 oncoprotein of Human Papilloma Virus-HPV16. J. Appl. Pharm. Sci. 2018, 8, 23–32. [Google Scholar]
- Bhatt, A.; Arora, P.; Prajapati, S.K. Can Algal Derived Bioactive Metabolites Serve as Potential Therapeutics for the Treatment of SARS-CoV-2 Like Viral Infection? Front. Microbiol. 2020, 11, 596374. [Google Scholar] [CrossRef] [PubMed]
- Hans, N.; Malik, A.; Naik, S. Antiviral activity of sulfated polysaccharides from marine algae and its application in combating COVID-19: Mini review. Bioresour. Technol. Rep. 2021, 13, 100623. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pressure (Bars) | XC (g/L) | XEPS (g/L) |
|---|---|---|
| 0 | 0.96 | 1.11 |
| 250 | 0.58 | 1.25 |
| 500 | 0.24 | 1.31 |
| 1000 | 0.07 | 1.50 |
| 1500 | 0.17 | 1.51 |
| 2000 | 0.20 | 1.51 |
| Fraction | XEPS (g/L) | V (L) |
|---|---|---|
| Feed | 0.68 | 8.4 |
| Retentate | 3.39 | 1.4 |
| Permeate | <0.01 | 7.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kronusová, O.; Kaštánek, P.; Koyun, G.; Kaštánek, F.; Brányik, T. Factors Influencing the Production of Extracellular Polysaccharides by the Green Algae Dictyosphaerium chlorelloides and Their Isolation, Purification, and Composition. Microorganisms 2022, 10, 1473. https://doi.org/10.3390/microorganisms10071473
Kronusová O, Kaštánek P, Koyun G, Kaštánek F, Brányik T. Factors Influencing the Production of Extracellular Polysaccharides by the Green Algae Dictyosphaerium chlorelloides and Their Isolation, Purification, and Composition. Microorganisms. 2022; 10(7):1473. https://doi.org/10.3390/microorganisms10071473
Chicago/Turabian StyleKronusová, Olga, Petr Kaštánek, Görkem Koyun, František Kaštánek, and Tomáš Brányik. 2022. "Factors Influencing the Production of Extracellular Polysaccharides by the Green Algae Dictyosphaerium chlorelloides and Their Isolation, Purification, and Composition" Microorganisms 10, no. 7: 1473. https://doi.org/10.3390/microorganisms10071473