

Commensal Bacteria Impact on Intestinal Toll-like Receptor Signaling in Salmonella-Challenged Gnotobiotic Piglets
 , , , ,
, , , ,
Abstract
:1. Introduction
1.1. Pathogen- and Damage-Associated Molecular Patterns
1.2. Colonization of the Newborn Intestinal Tract and Salmonella Typhimurium
1.3. Pig Translational Model
2. Materials and Methods
2.1. Gnotobiotic Minipigs
2.2. Bacterial Cultures
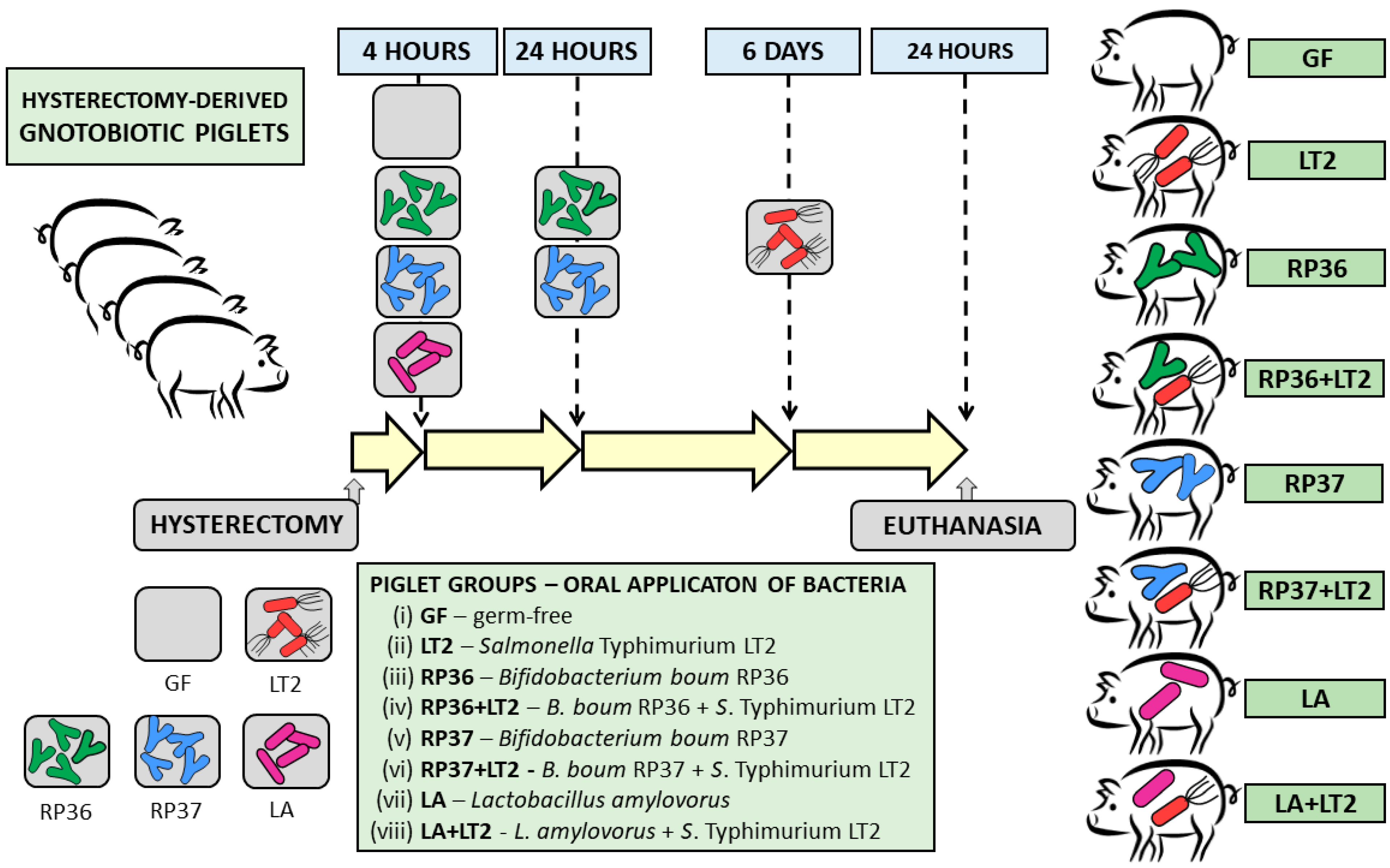
2.3. Experimental Design
2.4. Clinical Signs
2.5. Histological Evaluation
2.6. Blood Plasma
2.7. Total RNA Extraction and cDNA Synthesis
2.8. Real-Time PCR
2.9. IL-6 and IL-12/23p40 in Blood Plasma
2.10. Statistical Analysis
3. Results
3.1. Clinical Signs of Infection
3.2. Histological Assessment of the Terminal Ileum
3.3. Toll-like Receptors, Their Related Molecules, and RAGE mRNA Expression in the Ileum
3.4. Toll-like Receptors, Their Related Molecules, and RAGE mRNA Expression in the Colon
3.5. IL-6 and IL-12/23p40 Levels in Plasma
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medzhitov, R. The Spectrum of Inflammatory Responses. Science 2021, 374, 1070–1075. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr. Approaching the Asymptote? Evolution and Revolution in Immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 54 Pt 1, 1–13. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Chen, G.Y.; Nuñez, G. Sterile Inflammation: Sensing and Reacting to Damage. Nat. Rev. Immunol. 2010, 10, 826–837. [Google Scholar] [CrossRef]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-Sensing Receptors in Sterile Inflammation and Inflammatory Diseases. Nat. Rev. Immunol. 2020, 20, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Kono, H.; Rock, K.L. How Dying Cells Alert the Immune System to Danger. Nat. Rev. Immunol. 2008, 8, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The Role of Pattern-Recognition Receptors in Innate Immunity: Update on Toll-like Receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Kono, H.; Onda, A.; Yanagida, T. Molecular Determinants of Sterile Inflammation. Curr. Opin. Immunol. 2014, 26, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Paudel, Y.N.; Angelopoulou, E.; Piperi, C.; Balasubramaniam, V.R.M.T.; Othman, I.; Shaikh, M.F. Enlightening the Role of High Mobility Group Box 1 (HMGB1) in Inflammation: Updates on Receptor Signalling. Eur. J. Pharmacol. 2019, 858, 172487. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M. Signaling in Innate Immunity and Inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Cavaillon, J.-M.; Giamarellos-Bourboulis, E.J. Immunosuppression Is Inappropriately Qualifying the Immune Status of Septic and SIRS Patients. Shock 2019, 52, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Rock, K.L.; Latz, E.; Ontiveros, F.; Kono, H. The Sterile Inflammatory Response. Annu. Rev. Immunol. 2010, 28, 321–342. [Google Scholar] [CrossRef]
- Caroff, M.; Karibian, D.; Cavaillon, J.M.; Haeffner-Cavaillon, N. Structural and Functional Analyses of Bacterial Lipopolysaccharides. Microbes Infect. 2002, 4, 915–926. [Google Scholar] [CrossRef]
- Kagan, J.C. Lipopolysaccharide Detection across the Kingdoms of Life. Trends Immunol. 2017, 38, 696–704. [Google Scholar] [CrossRef]
- Dong, Y.; Glaser, K.; Speer, C.P. Late-Onset Sepsis Caused by Gram-Negative Bacteria in Very Low Birth Weight Infants: A Systematic Review. Expert Rev. Anti. Infect. Ther. 2019, 17, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Karki, R.; Kanneganti, T.-D. The “Cytokine Storm”: Molecular Mechanisms and Therapeutic Prospects. Trends Immunol. 2021, 42, 681–705. [Google Scholar] [CrossRef] [PubMed]
- Meizlish, M.L.; Franklin, R.A.; Zhou, X.; Medzhitov, R. Tissue Homeostasis and Inflammation. Annu. Rev. Immunol. 2021, 39, 557–581. [Google Scholar] [CrossRef]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, J.C. The Microbiome in Early Life: Implications for Health Outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef]
- Healy, D.B.; Ryan, C.A.; Ross, R.P.; Stanton, C.; Dempsey, E.M. Clinical Implications of Preterm Infant Gut Microbiome Development. Nat. Microbiol. 2022, 7, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhai, Q.; Zhang, H.; Chen, W.; Hill, C. Gut Colonization Mechanisms of Lactobacillus and Bifidobacterium: An Argument for Personalized Designs. Annu. Rev. Food Sci. Technol. 2021, 12, 213–233. [Google Scholar] [CrossRef] [PubMed]
- Kwoji, I.D.; Aiyegoro, O.A.; Okpeku, M.; Adeleke, M.A. Multi-Strain Probiotics: Synergy among Isolates Enhances Biological Activities. Biology 2021, 10, 322. [Google Scholar] [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Vasileva, E.V.; Odorskaya, M.V.; Marsova, M.V.; Kovalev, G.I.; Danilenko, V.N. A Multi-Strain Potential Probiotic Formulation of GABA-Producing Lactobacillus plantarum 90sk and Bifidobacterium adolescentis 150 with Antidepressant Effects. Probiotics Antimicrob. Proteins 2020, 12, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Ducarmon, Q.R.; Zwittink, R.D.; Hornung, B.V.H.; van Schaik, W.; Young, V.B.; Kuijper, E.J. Gut Microbiota and Colonization Resistance against Bacterial Enteric Infection. Microbiol. Mol. Biol. Rev. 2019, 83, e00007-19. [Google Scholar] [CrossRef] [PubMed]
- Herzog, M.K.-M.; Cazzaniga, M.; Peters, A.; Shayya, N.; Beldi, L.; Hapfelmeier, S.; Heimesaat, M.M.; Bereswill, S.; Frankel, G.; Gahan, C.G.M.; et al. Mouse Models for Bacterial Enteropathogen Infections: Insights into the Role of Colonization Resistance. Gut Microbes 2023, 15, 2172667. [Google Scholar] [CrossRef]
- Besser, J.M. Salmonella Epidemiology: A Whirlwind of Change. Food Microbiol. 2018, 71, 55–59. [Google Scholar] [CrossRef]
- Hurley, D.; McCusker, M.P.; Fanning, S.; Martins, M. Salmonella-Host Interactions—Modulation of the Host Innate Immune System. Front. Immunol. 2014, 5, 481. [Google Scholar] [CrossRef]
- Heredia, N.; García, S. Animals as Sources of Food-Borne Pathogens: A Review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef]
- Coburn, B.; Grassl, G.A.; Finlay, B.B. Salmonella, the Host and Disease: A Brief Review. Immunol. Cell Biol. 2007, 85, 112–118. [Google Scholar] [CrossRef]
- Ménard, S.; Lacroix-Lamandé, S.; Ehrhardt, K.; Yan, J.; Grassl, G.A.; Wiedemann, A. Cross-Talk Between the Intestinal Epithelium and Salmonella Typhimurium. Front. Microbiol. 2022, 13, 906238. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.A. Salmonella Infections in Immunocompromised Adults. J. Infect. 2008, 56, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.C.; Best, E.; Nourse, C. Non-Typhoidal Salmonella Infections in Children: Review of Literature and Recommendations for Management. J. Paediatr. Child Health 2017, 53, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Estellé, J.; Kiilerich, P.; Ramayo-Caldas, Y.; Xia, Z.; Feng, Q.; Liang, S.; Pedersen, A.Ø.; Kjeldsen, N.J.; Liu, C.; et al. A Reference Gene Catalogue of the Pig Gut Microbiome. Nat. Microbiol. 2016, 1, 16161. [Google Scholar] [CrossRef]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the Pig as a Human Biomedical Model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.; Sangild, P.T.; Stoll, B.; Thymann, T.; Buddington, R.; Marini, J.; Olutoye, O.; Shulman, R.J. Translational Advances in Pediatric Nutrition and Gastroenterology: New Insights from Pig Models. Annu. Rev. Anim. Biosci. 2020, 8, 321–354. [Google Scholar] [CrossRef]
- Zhang, Q.; Widmer, G.; Tzipori, S. A Pig Model of the Human Gastrointestinal Tract. Gut Microbes 2013, 4, 193–200. [Google Scholar] [CrossRef]
- Pabst, R. The Pig as a Model for Immunology Research. Cell Tissue Res. 2020, 380, 287–304. [Google Scholar] [CrossRef]
- Rothkötter, H.J.; Sowa, E.; Pabst, R. The Pig as a Model of Developmental Immunology. Hum. Exp. Toxicol. 2002, 21, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The Pig: A Model for Human Infectious Diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef]
- Roberts, R.M.; Green, J.A.; Schulz, L.C. The Evolution of the Placenta. Reproduction 2016, 152, R179–R189. [Google Scholar] [CrossRef]
- Bigler, N.A.; Bruckmaier, R.M.; Gross, J.J. Implications of Placentation Type on Species-Specific Colostrum Properties in Mammals. J. Anim. Sci. 2022, 100, skac287. [Google Scholar] [CrossRef] [PubMed]
- Salmon, H.; Berri, M.; Gerdts, V.; Meurens, F. Humoral and Cellular Factors of Maternal Immunity in Swine. Dev. Comp. Immunol. 2009, 33, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Fricke, W.F.; Ravel, J. Microbiome or No Microbiome: Are We Looking at the Prenatal Environment through the Right Lens? Microbiome 2021, 9, 9. [Google Scholar] [CrossRef]
- Kennedy, K.M.; de Goffau, M.C.; Perez-Muñoz, M.E.; Arrieta, M.-C.; Bäckhed, F.; Bork, P.; Braun, T.; Bushman, F.D.; Dore, J.; de Vos, W.M.; et al. Questioning the Fetal Microbiome Illustrates Pitfalls of Low-Biomass Microbial Studies. Nature 2023, 613, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Perez-Muñoz, M.E.; Arrieta, M.-C.; Ramer-Tait, A.E.; Walter, J. A Critical Assessment of the “Sterile Womb” and “in Utero Colonization” Hypotheses: Implications for Research on the Pioneer Infant Microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef]
- Miniats, O.P.; Jol, D. Gnotobiotic Pigs-Derivation and Rearing. Can. J. Comp. Med. 1978, 42, 428–437. [Google Scholar] [PubMed]
- Taylor, D.M. The Use of Laminar Flow for Obtaining Germ-Free Mice. Lab. Anim. 1975, 9, 337–343. [Google Scholar] [CrossRef]
- Ruff, W.E.; Greiling, T.M.; Kriegel, M.A. Host-Microbiota Interactions in Immune-Mediated Diseases. Nat. Rev. Microbiol. 2020, 18, 521–538. [Google Scholar] [CrossRef]
- Splichal, I.; Rychlik, I.; Splichalova, I.; Karasova, D.; Splichalova, A. Toll-like Receptor 4 Signaling in the Ileum and Colon of Gnotobiotic Piglets Infected with Salmonella Typhimurium or Its Isogenic ∆rfa Mutants. Toxins 2020, 12, 545. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.C.; Gyles, C.L. Virulence of Wild and Mutant Strains of Salmonella Typhimurium in Ligated Intestinal Segments of Calves, Pigs, and Rabbits. Am. J. Vet. Res. 1987, 48, 504–510. [Google Scholar] [PubMed]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The Microbiome and Innate Immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Splichalova, A.; Slavikova, V.; Splichalova, Z.; Splichal, I. Preterm Life in Sterile Conditions: A Study on Preterm, Germ-Free Piglets. Front. Immunol. 2018, 9, 220. [Google Scholar] [CrossRef] [PubMed]
- Pechar, R.; Killer, J.; Mekadim, C.; Geigerová, M.; Rada, V. Classification of Culturable Bifidobacterial Population from Colonic Samples of Wild Pigs (Sus Scrofa) Based on Three Molecular Genetic Methods. Curr. Microbiol. 2017, 74, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Splichalova, A.; Pechar, R.; Killer, J.; Splichalova, Z.; Bunesova, V.N.; Vlkova, E.; Salmonova, H.S.; Splichal, I. Colonization of Germ-Free Piglets with Mucinolytic and Non-Mucinolytic Bifidobacterium boum Strains Isolated from the Intestine of Wild Boar and Their Interference with Salmonella Typhimurium. Microorganisms 2020, 8, 2002. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Poznanski, E.; Fuentes, S.; Akkermans, A.D.L.; Smidt, H.; de Vos, W.M. Lactobacillus sobrius Sp. Nov., Abundant in the Intestine of Weaning Piglets. Int. J. Syst. Evol. Microbiol. 2006, 56, 29–32. [Google Scholar] [CrossRef]
- Trebichavsky, I.; Dlabac, V.; Rehakova, Z.; Zahradnickova, M.; Splichal, I. Cellular Changes and Cytokine Expression in the Ilea of Gnotobiotic Piglets Resulting from Peroral Salmonella Typhimurium Challenge. Infect. Immun. 1997, 65, 5244–5249. [Google Scholar] [CrossRef]
- McClelland, M.; Sanderson, K.E.; Spieth, J.; Clifton, S.W.; Latreille, P.; Courtney, L.; Porwollik, S.; Ali, J.; Dante, M.; Du, F.; et al. Complete Genome Sequence of Salmonella Enterica Serovar Typhimurium LT2. Nature 2001, 413, 852–856. [Google Scholar] [CrossRef]
- Splichal, I.; Donovan, S.M.; Splichalova, Z.; Neuzil Bunesova, V.; Vlkova, E.; Jenistova, V.; Killer, J.; Svejstil, R.; Skrivanova, E.; Splichalova, A. Colonization of Germ-Free Piglets with Commensal Lactobacillus amylovorus, Lactobacillus mucosae, and Probiotic E. Coli Nissle 1917 and Their Interference with Salmonella Typhimurium. Microorganisms 2019, 7, 273. [Google Scholar] [CrossRef] [PubMed]
- Splichalova, A.; Jenistova, V.; Splichalova, Z.; Splichal, I. Colonization of Preterm Gnotobiotic Piglets with Probiotic Lactobacillus rhamnosus GG and Its Interference with Salmonella Typhimurium. Clin. Exp. Immunol. 2019, 195, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Gal-Mor, O. Persistent Infection and Long-Term Carriage of Typhoidal and Nontyphoidal Salmonellae. Clin. Microbiol. Rev. 2019, 32, e00088-18. [Google Scholar] [CrossRef] [PubMed]
- Skrzypek, T.; Valverde Piedra, J.L.; Skrzypek, H.; Kazimierczak, W.; Biernat, M.; Zabielski, R. Gradual Disappearance of Vacuolated Enterocytes in the Small Intestine of Neonatal Piglets. J. Physiol. Pharmacol. 2007, 58 (Suppl. S3), 87–95. [Google Scholar]
- Zabielski, R.; Godlewski, M.M.; Guilloteau, P. Control of Development of Gastrointestinal System in Neonates. J. Physiol. Pharmacol. 2008, 59 (Suppl. S1), 35–54. [Google Scholar]
- Shirkey, T.W.; Siggers, R.H.; Goldade, B.G.; Marshall, J.K.; Drew, M.D.; Laarveld, B.; Van Kessel, A.G. Effects of Commensal Bacteria on Intestinal Morphology and Expression of Proinflammatory Cytokines in the Gnotobiotic Pig. Exp. Biol. Med. 2006, 231, 1333–1345. [Google Scholar] [CrossRef]
- Zhang, K.; Griffiths, G.; Repnik, U.; Hornef, M. Seeing Is Understanding: Salmonella’s Way to Penetrate the Intestinal Epithelium. Int. J. Med. Microbiol. 2018, 308, 97–106. [Google Scholar] [CrossRef]
- Keestra-Gounder, A.M.; Tsolis, R.M.; Bäumler, A.J. Now You See Me, Now You Don’t: The Interaction of Salmonella with Innate Immune Receptors. Nat. Rev. Microbiol. 2015, 13, 206–216. [Google Scholar] [CrossRef]
- Trebichavsky, I.; Splichal, I.; Rada, V.; Splichalova, A. Modulation of Natural Immunity in the Gut by Escherichia Coli Strain Nissle 1917. Nutr. Rev. 2010, 68, 459–464. [Google Scholar] [CrossRef]
- Jakava-Viljanen, M.; Murros, A.; Palva, A.; Björkroth, K.J. Lactobacillus sobrius Konstantinov et al. 2006 Is a Later Synonym of Lactobacillus amylovorus Nakamura 1981. Int. J. Syst. Evol. Microbiol. 2008, 58, 910–913. [Google Scholar] [CrossRef]
- Tükel, C.; Nishimori, J.H.; Wilson, R.P.; Winter, M.G.; Keestra, A.M.; van Putten, J.P.M.; Bäumler, A.J. Toll-like Receptors 1 and 2 Cooperatively Mediate Immune Responses to Curli, a Common Amyloid from Enterobacterial Biofilms. Cell. Microbiol. 2010, 12, 1495–1505. [Google Scholar] [CrossRef] [PubMed]
- Rapsinski, G.J.; Wynosky-Dolfi, M.A.; Oppong, G.O.; Tursi, S.A.; Wilson, R.P.; Brodsky, I.E.; Tükel, Ç. Toll-like Receptor 2 and NLRP3 Cooperate to Recognize a Functional Bacterial Amyloid, Curli. Infect. Immun. 2015, 83, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Capurso, L. Thirty Years of Lactobacillus rhamnosus GG: A Review. J. Clin. Gastroenterol. 2019, 53 (Suppl. S1), S1–S41. [Google Scholar] [CrossRef] [PubMed]
- Splichal, I.; Donovan, S.M.; Kindlova, Z.; Stranak, Z.; Neuzil Bunesova, V.; Sinkora, M.; Polakova, K.; Valaskova, B.; Splichalova, A. Release of HMGB1 and Toll-like Receptors 2, 4, and 9 Signaling Are Modulated by Bifidobacterium animalis Subsp. Lactis BB-12 and Salmonella Typhimurium in a Gnotobiotic Piglet Model of Preterm Infants. Int. J. Mol. Sci. 2023, 24, 2329. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.; Al-Nakeeb, K.; Koza, A.; Zeidan, A.A. Updated Genome Sequence for the Probiotic Bacterium Bifidobacterium animalis Subsp. Lactis BB-12. Microbiol. Resour. Announc. 2021, 10, e0007821. [Google Scholar] [CrossRef]
- Splichal, I.; Donovan, S.M.; Jenistova, V.; Splichalova, I.; Salmonova, H.; Vlkova, E.; Neuzil Bunesova, V.; Sinkora, M.; Killer, J.; Skrivanova, E.; et al. High Mobility Group Box 1 and TLR4 Signaling Pathway in Gnotobiotic Piglets Colonized/Infected with L. Amylovorus, L. Mucosae, E. Coli Nissle 1917 and S. Typhimurium. Int. J. Mol. Sci. 2019, 20, 6294. [Google Scholar] [CrossRef]
- Henker, J.; Laass, M.; Blokhin, B.M.; Bolbot, Y.K.; Maydannik, V.G.; Elze, M.; Wolff, C.; Schulze, J. The Probiotic Escherichia Coli Strain Nissle 1917 (EcN) Stops Acute Diarrhoea in Infants and Toddlers. Eur. J. Pediatr. 2007, 166, 311–318. [Google Scholar] [CrossRef]
- Henker, J.; Laass, M.W.; Blokhin, B.M.; Maydannik, V.G.; Bolbot, Y.K.; Elze, M.; Wolff, C.; Schreiner, A.; Schulze, J. Probiotic Escherichia Coli Nissle 1917 versus Placebo for Treating Diarrhea of Greater than 4 Days Duration in Infants and Toddlers. Pediatr. Infect. Dis. J. 2008, 27, 494–499. [Google Scholar] [CrossRef]
- Splichalova, A.; Trebichavsky, I.; Rada, V.; Vlkova, E.; Sonnenborn, U.; Splichal, I. Interference of Bifidobacterium choerinum or Escherichia Coli Nissle 1917 with Salmonella Typhimurium in Gnotobiotic Piglets Correlates with Cytokine Patterns in Blood and Intestine. Clin. Exp. Immunol. 2011, 163, 242–249. [Google Scholar] [CrossRef]
- Grozdanov, L.; Zähringer, U.; Blum-Oehler, G.; Brade, L.; Henne, A.; Knirel, Y.A.; Schombel, U.; Schulze, J.; Sonnenborn, U.; Gottschalk, G.; et al. A Single Nucleotide Exchange in the Wzy Gene Is Responsible for the Semirough O6 Lipopolysaccharide Phenotype and Serum Sensitivity of Escherichia Coli Strain Nissle 1917. J. Bacteriol. 2002, 184, 5912–5925. [Google Scholar] [CrossRef]
- Foster, N.; Lovell, M.A.; Marston, K.L.; Hulme, S.D.; Frost, A.J.; Bland, P.; Barrow, P.A. Rapid Protection of Gnotobiotic Pigs against Experimental Salmonellosis Following Induction of Polymorphonuclear Leukocytes by Avirulent Salmonella Enterica. Infect. Immun. 2003, 71, 2182–2191. [Google Scholar] [CrossRef]
- Foster, N.; Hulme, S.; Lovell, M.; Reed, K.; Barrow, P. Stimulation of Gp91 Phagocytic Oxidase and Reactive Oxygen Species in Neutrophils by an Avirulent Salmonella Enterica Serovar Infantis Strain Protects Gnotobiotic Piglets from Lethal Challenge with Serovar Typhimurium Strain F98 without Inducing Intestinal Pathology. Infect. Immun. 2005, 73, 4539–4547. [Google Scholar] [CrossRef] [PubMed]
- Splichal, I.; Trebichavsky, I.; Splichalova, A.; Barrow, P.A. Protection of Gnotobiotic Pigs against Salmonella Enterica Serotype Typhimurium by Rough Mutant of the Same Serotype Is Accompanied by the Change of Local and Systemic Cytokine Response. Vet. Immunol. Immunopathol. 2005, 103, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Splichalova, A.; Splichalova, Z.; Karasova, D.; Rychlik, I.; Trevisi, P.; Sinkora, M.; Splichal, I. Impact of the Lipopolysaccharide Chemotype of Salmonella Enterica Serovar Typhimurium on Virulence in Gnotobiotic Piglets. Toxins 2019, 11, 534. [Google Scholar] [CrossRef] [PubMed]
- Splichalova, A.; Splichal, I.; Chmelarova, P.; Trebichavsky, I. Alarmin HMGB1 Is Released in the Small Intestine of Gnotobiotic Piglets Infected with Enteric Pathogens and Its Level in Plasma Reflects Severity of Sepsis. J. Clin. Immunol. 2011, 31, 488–497. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Andersson, U. Targeting Inflammation Driven by HMGB1. Front. Immunol. 2020, 11, 484. [Google Scholar] [CrossRef]
- Andersson, U.; Yang, H. HMGB1 Is a Critical Molecule in the Pathogenesis of Gram-Negative Sepsis. J. Intensive Med. 2022, 2, 156–166. [Google Scholar] [CrossRef]
- Burkey, T.E.; Skjolaas, K.A.; Dritz, S.S.; Minton, J.E. Expression of Toll-like Receptors, Interleukin 8, Macrophage Migration Inhibitory Factor, and Osteopontin in Tissues from Pigs Challenged with Salmonella Enterica Serovar Typhimurium or Serovar Choleraesuis. Vet. Immunol. Immunopathol. 2007, 115, 309–319. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, Y.; Huang, Y.; Deng, H. Pathophysiology of RAGE in Inflammatory Diseases. Front. Immunol. 2022, 13, 931473. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, A.M.; Shanahan, F. The Gut Flora as a Forgotten Organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Bescucci, D.M.; Moote, P.E.; Ortega Polo, R.; Uwiera, R.R.E.; Inglis, G.D. Salmonella Enterica Serovar Typhimurium Temporally Modulates the Enteric Microbiota and Host Responses To Overcome Colonization Resistance in Swine. Appl. Environ. Microbiol. 2020, 86, e01569-20. [Google Scholar] [CrossRef]
- Finamore, A.; Roselli, M.; Imbinto, A.; Seeboth, J.; Oswald, I.P.; Mengheri, E. Lactobacillus amylovorus Inhibits the TLR4 Inflammatory Signaling Triggered by Enterotoxigenic Escherichia Coli via Modulation of the Negative Regulators and Involvement of TLR2 in Intestinal Caco-2 Cells and Pig Explants. PLoS ONE 2014, 9, e94891. [Google Scholar] [CrossRef] [PubMed]
- Pierrakos, C.; Velissaris, D.; Bisdorff, M.; Marshall, J.C.; Vincent, J.-L. Biomarkers of Sepsis: Time for a Reappraisal. Crit. Care 2020, 24, 287. [Google Scholar] [CrossRef]
- Cavaillon, J.-M.; Singer, M.; Skirecki, T. Sepsis Therapies: Learning from 30 Years of Failure of Translational Research to Propose New Leads. EMBO Mol. Med. 2020, 12, e10128. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Control of Adaptive Immunity by the Innate Immune System. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Park, D.W.; Moon, S.; Cho, H.-J.; Park, J.H.; Seok, H.; Choi, W.S. Diagnostic and Prognostic Value of Interleukin-6, Pentraxin 3, and Procalcitonin Levels among Sepsis and Septic Shock Patients: A Prospective Controlled Study According to the Sepsis-3 Definitions. BMC Infect. Dis. 2019, 19, 968. [Google Scholar] [CrossRef]
- Huang, F.-C. The Interleukins Orchestrate Mucosal Immune Responses to Salmonella Infection in the Intestine. Cells 2021, 10, 3492. [Google Scholar] [CrossRef]
- Splichalova, A.; Splichal, I. Local and Systemic Occurrences of HMGB1 in Gnotobiotic Piglets Infected with E. Coli O55 Are Related to Bacterial Translocation and Inflammatory Cytokines. Cytokine 2012, 60, 597–600. [Google Scholar] [CrossRef]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the Mucosal Barrier to Infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, C.; Fieschi, C.; Lammas, D.A.; Picard, C.; Dorman, S.E.; Sanal, O.; MacLennan, J.M.; Holland, S.M.; Ottenhoff, T.H.M.; Casanova, J.-L.; et al. Interleukin (IL)-12 and IL-23 Are Key Cytokines for Immunity against Salmonella in Humans. J. Infect. Dis. 2004, 190, 1755–1757. [Google Scholar] [CrossRef] [PubMed]
- Awoniyi, M.; Miller, S.I.; Wilson, C.B.; Hajjar, A.M.; Smith, K.D. Homeostatic Regulation of Salmonella-Induced Mucosal Inflammation and Injury by IL-23. PLoS ONE 2012, 7, e37311. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.A.; Kuchroo, V.K. IL-12 Family Cytokines: Immunological Playmakers. Nat. Immunol. 2012, 13, 722–728. [Google Scholar] [CrossRef]
- Stecher, B. Establishing Causality in Salmonella-Microbiota-Host Interaction: The Use of Gnotobiotic Mouse Models and Synthetic Microbial Communities. Int. J. Med. Microbiol. 2021, 311, 151484. [Google Scholar] [CrossRef] [PubMed]
- Brugiroux, S.; Berry, D.; Ring, D.; Barnich, N.; Daims, H.; Stecher, B. Specific Localization and Quantification of the Oligo-Mouse-Microbiota (OMM12) by Fluorescence In Situ Hybridization (FISH). Curr. Protoc. 2022, 2, e548. [Google Scholar] [CrossRef]
- Horvathova, K.; Modrackova, N.; Splichal, I.; Splichalova, A.; Amin, A.; Ingribelli, E.; Killer, J.; Doskocil, I.; Pechar, R.; Kodesova, T.; et al. Defined Pig Microbiota with a Potential Protective Effect against Infection with Salmonella Typhimurium. Microorganisms 2023, 11, 1007. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GF | RP36 | RL37 | LA | |
|---|---|---|---|---|
| Villus height (μm) | 704.9 ± 76.2 | 614.2 ± 71.4 | 614.8 ± 40.1 | 635.9 ± 56.5 |
| Crypt depth (μm) | 74.7 ± 3.8 | 77.2 ± 7.8 | 78.7±4.3 | 77.2 ± 6.1 |
| Height/Depth (ratio) | 9.1 ± 2.1 | 8.2 ± 1.6 | 6.9 ± 0.8 | 8.0 ± 1.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Splichalova, A.; Kindlova, Z.; Killer, J.; Neuzil Bunesova, V.; Vlkova, E.; Valaskova, B.; Pechar, R.; Polakova, K.; Splichal, I. Commensal Bacteria Impact on Intestinal Toll-like Receptor Signaling in Salmonella-Challenged Gnotobiotic Piglets. Pathogens 2023, 12, 1293. https://doi.org/10.3390/pathogens12111293
Splichalova A, Kindlova Z, Killer J, Neuzil Bunesova V, Vlkova E, Valaskova B, Pechar R, Polakova K, Splichal I. Commensal Bacteria Impact on Intestinal Toll-like Receptor Signaling in Salmonella-Challenged Gnotobiotic Piglets. Pathogens. 2023; 12(11):1293. https://doi.org/10.3390/pathogens12111293
Chicago/Turabian StyleSplichalova, Alla, Zdislava Kindlova, Jiri Killer, Vera Neuzil Bunesova, Eva Vlkova, Barbora Valaskova, Radko Pechar, Katerina Polakova, and Igor Splichal. 2023. "Commensal Bacteria Impact on Intestinal Toll-like Receptor Signaling in Salmonella-Challenged Gnotobiotic Piglets" Pathogens 12, no. 11: 1293. https://doi.org/10.3390/pathogens12111293