
Reduced Temperature Sensitivity of Maximum Latewood Density Formation in High-Elevation Corsican Pines under Recent Warming

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
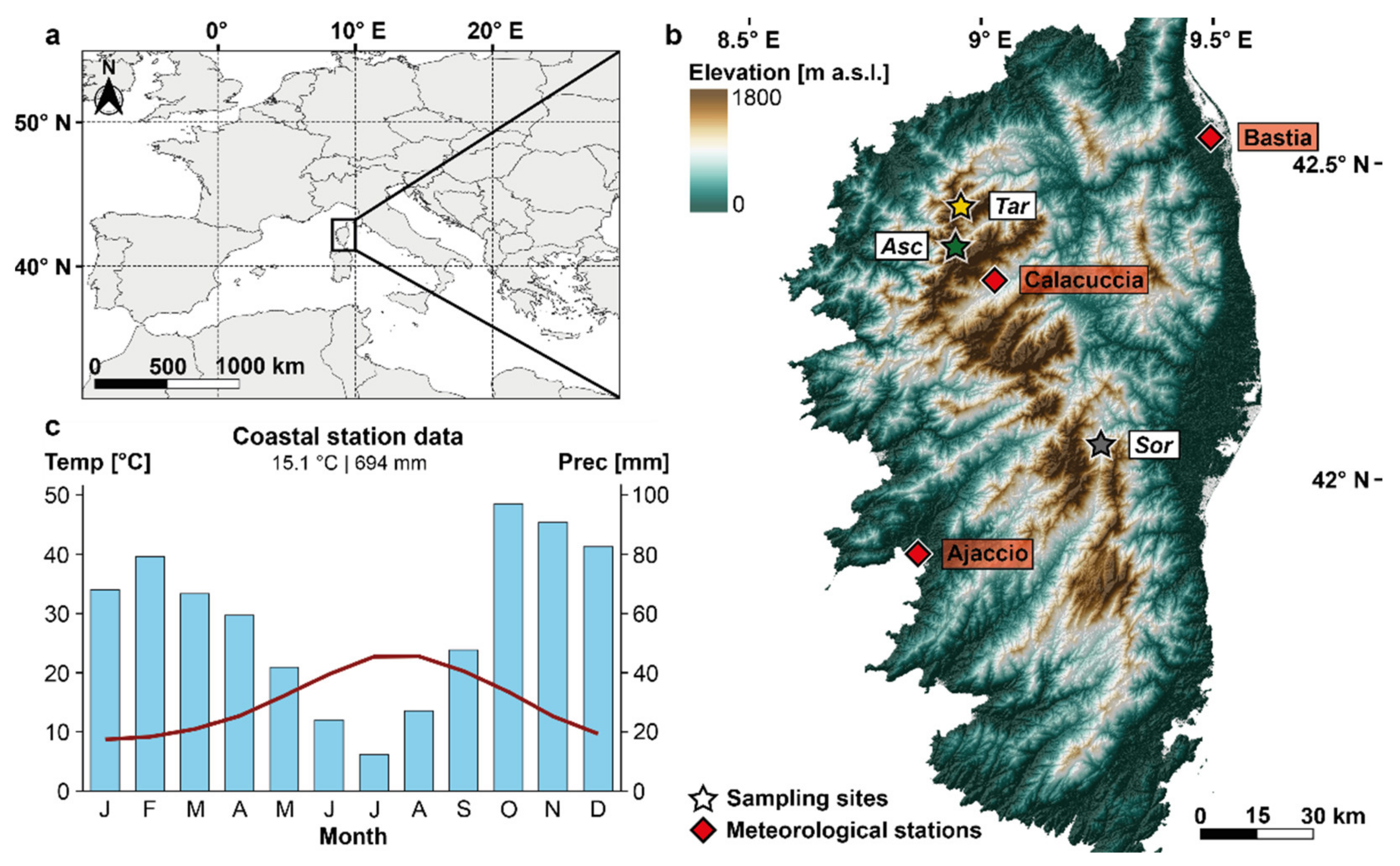
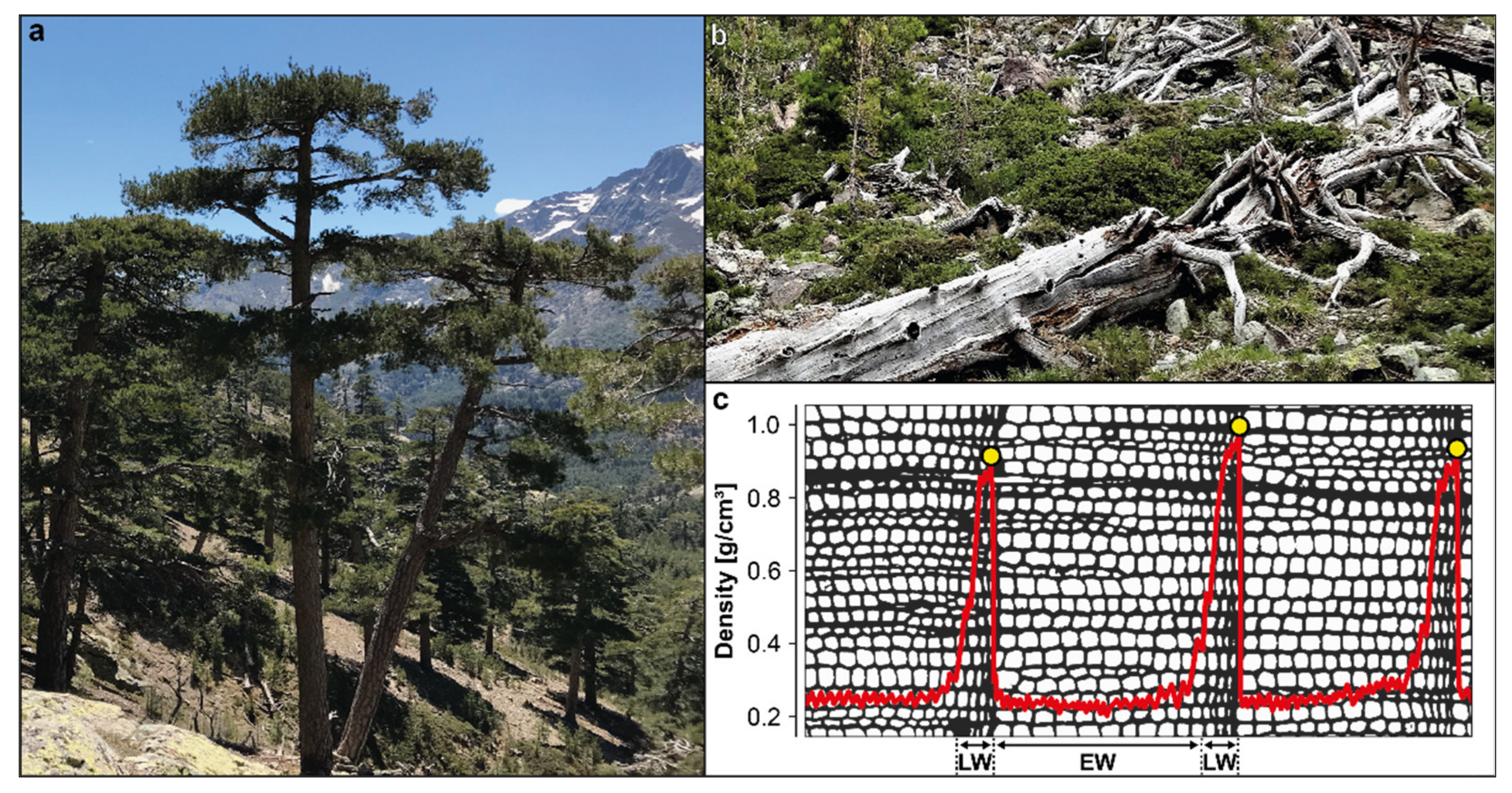
2.1. Wood Density Data and Chronology Development
2.2. Climate Data and Signal Estimation
3. Results
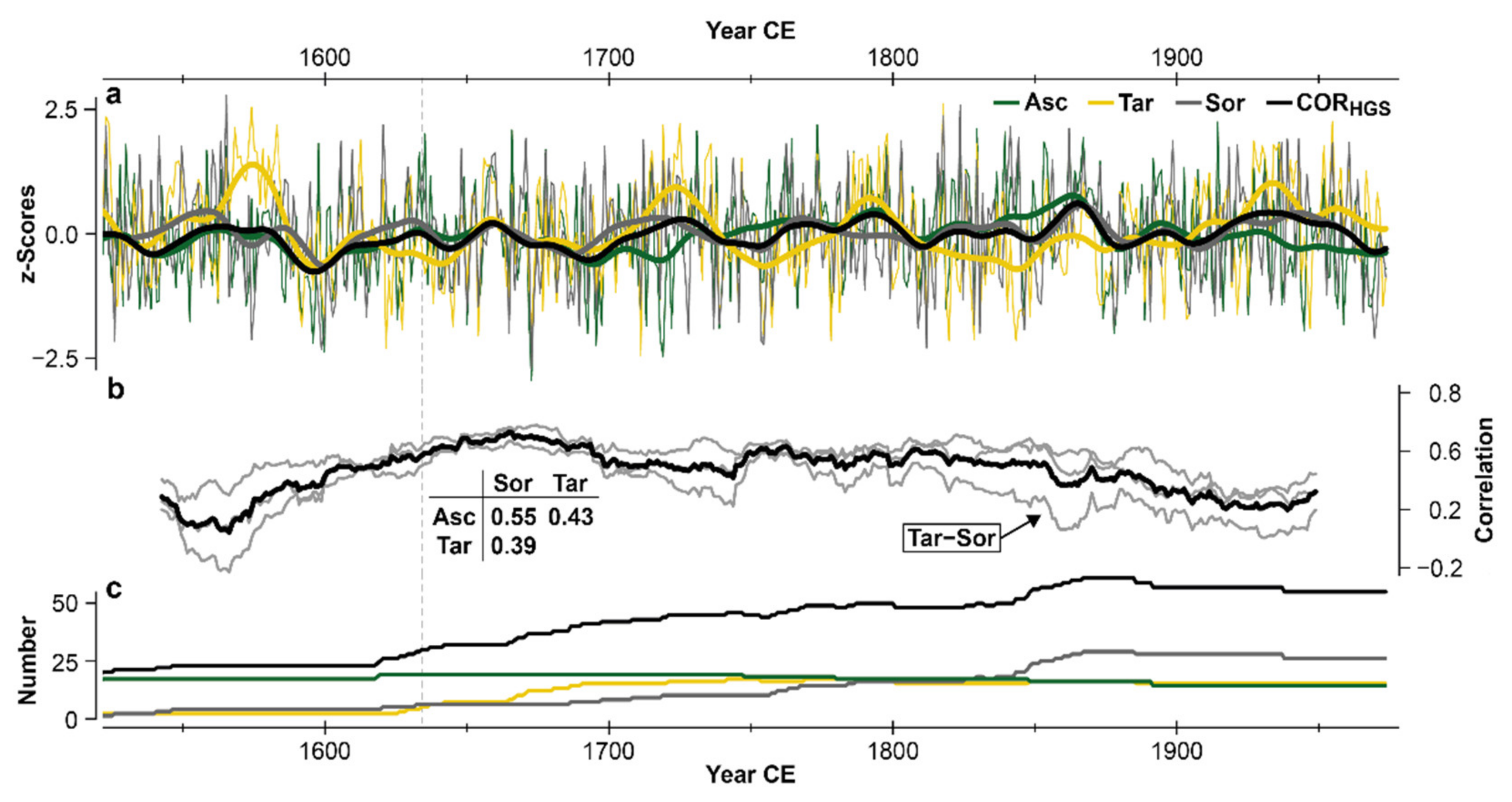
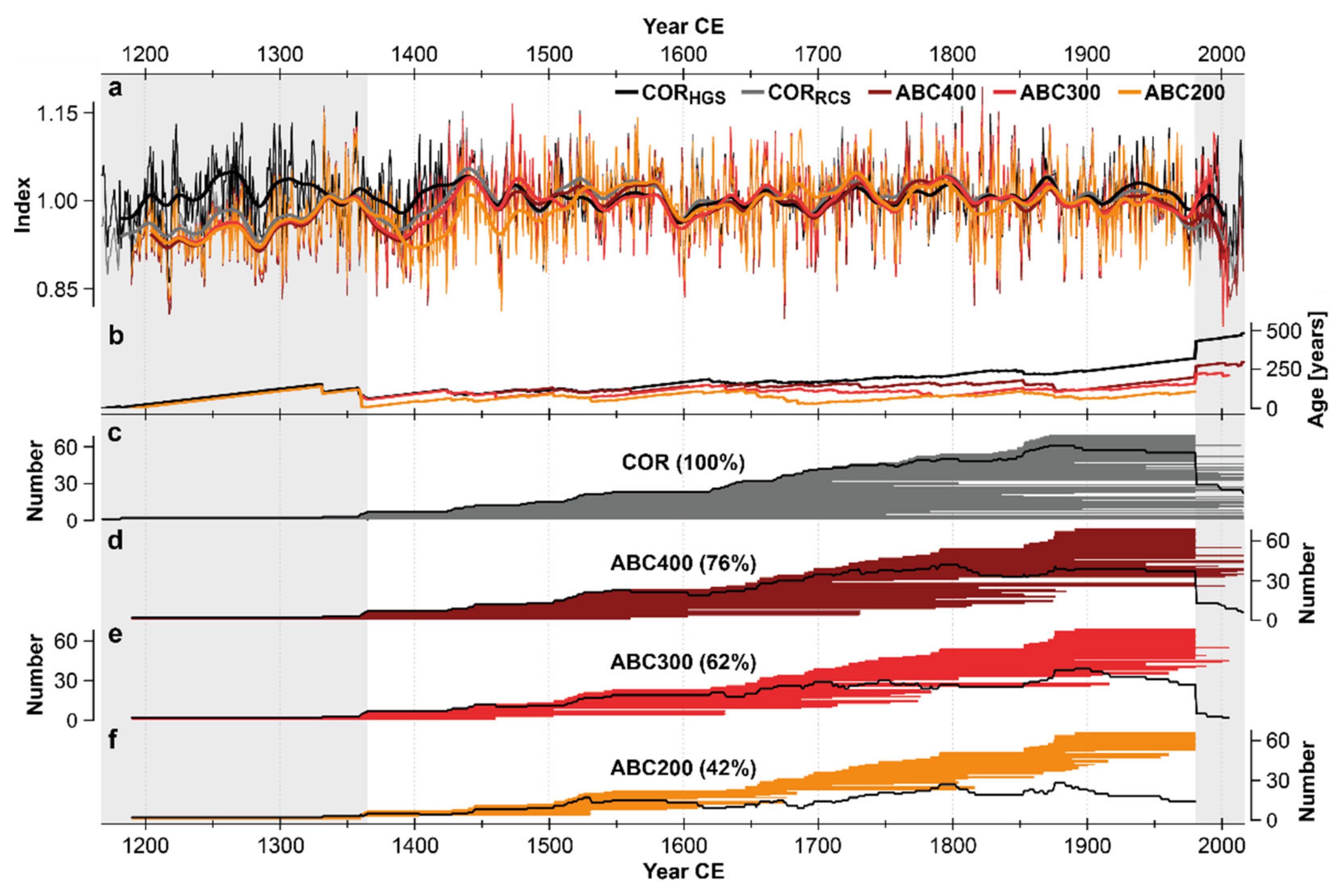
3.1. MXD Chronology Characteristics
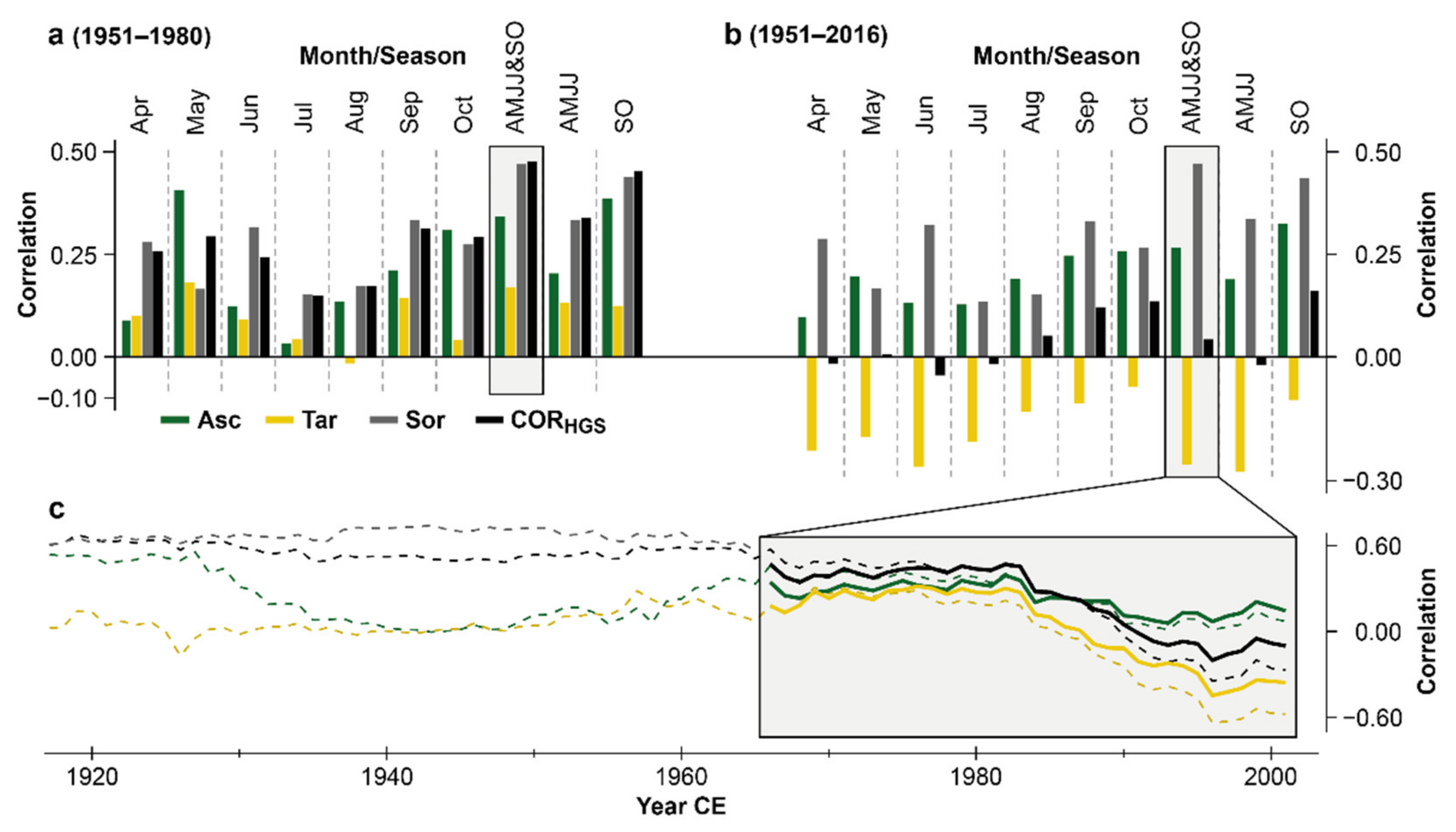
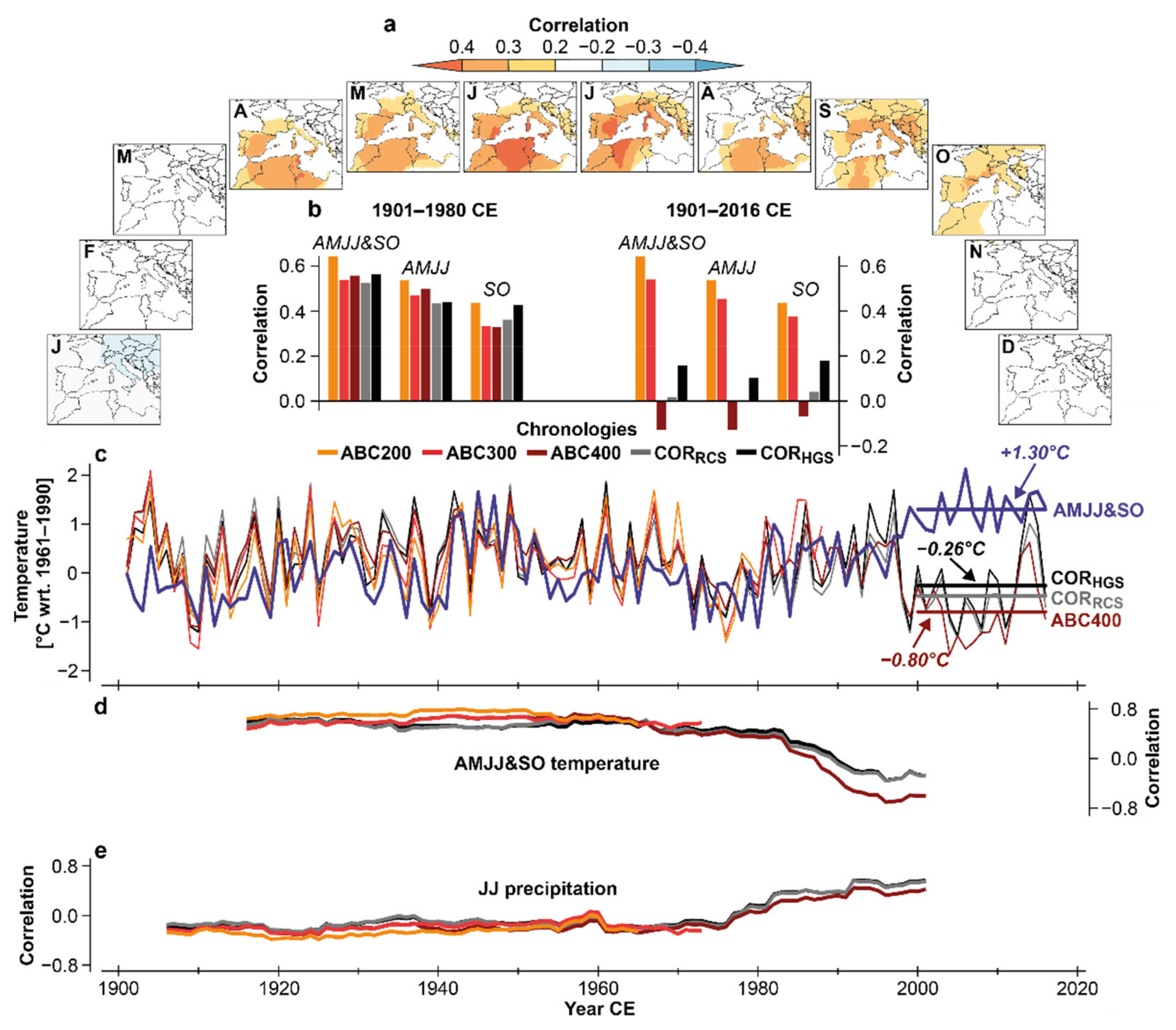
3.2. Climate Sensitivity of Corsican MXD
4. Discussion
4.1. Chronology Variants and Characteristics
4.2. Bimodal Temperature Signal
4.3. Fading Temperature Sensitivity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diffenbaugh, N.S.; Giorgi, F. Climate change hotspots in the CMIP5 global climate model ensemble. Clim. Chang. 2012, 114, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Giorgi, F. Increased aridity in the Mediterranean region under greenhouse gas forcing estimated from high resolution simulations with a regional climate model. Glob. Planet. Chang. 2008, 62, 195–209. [Google Scholar] [CrossRef]
- Hertig, E.; Jacobeit, J. Downscaling future climate change: Temperature scenarios for the Mediterranean area. Glob. Planet. Chang. 2008, 63, 127–131. [Google Scholar] [CrossRef]
- Beniston, M.; Stephenson, D.; Christensen, O.B.; Ferro, C.; Frei, C.; Goyette, S.; Halsnaes, K.; Holt, T.; Jylhä, K.; Koffi, B.; et al. Future extreme events in European climate: An exploration of regional climate model projections. Clim. Chang. 2007, 81, 71–95. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, E.; Gallardo, C.; Gaertner, M.A.; Arribas, A.; Castro, M. Future climate extreme events in the Mediterranean simulated by a regional climate model: A first approach. Glob. Planet. Chang. 2004, 44, 163–180. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate, 1st ed.; Blackburn Press: Caldwell, NJ, USA, 1976; ISBN 978-1-930665-39-2. [Google Scholar]
- Hetzer, T. Xylem Variability as a Proxy for Environmental and Climate Change in Corsica During the Past Millennium. Ph.D. Thesis, Friedrich-Alexander-University Erlangen-Nürnberg, Erlangen, Germany, 2013. [Google Scholar]
- Szymczak, S.; Joachimski, M.M.; Bräuning, A.; Hetzer, T.; Kuhlemann, J. A 560 yr summer temperature reconstruction for the western Mediterranean basin based on stable carbon isotopes from Pinus nigra ssp. laricio (Corsica/France). Clim. Past 2012, 8, 1737–1749. [Google Scholar] [CrossRef] [Green Version]
- Szymczak, S.; Hetzer, T.; Bräuning, A.; Joachimski, M.M.; Leuschner, H.-H.; Kuhlemann, J. Combining wood anatomy and stable isotope variations in a 600-year multi-parameter climate reconstruction from Corsican black pine. Quat. Sci. Rev. 2014, 101, 146–158. [Google Scholar] [CrossRef]
- Knerr, I.; Trachte, K.; Garel, E.; Huneau, F.; Santoni, S.; Bendix, J. Partitioning of large-scale and local-scale precipitation events by means of spatio-temporal precipitation regimes on Corsica. Atmosphere 2020, 11, 417. [Google Scholar] [CrossRef] [Green Version]
- Rome, S.; Giorgetti, J.-P. La montagne corse et ses caractéristiques climatiques. Météorologie 2007, 59, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Koninklijk Nederlands Meteorologisch Instituut (KNMI) Climate Explorer. Climate Data from Ajaccio (WMO Station Code 7761) and Bastia (WMO Station Code 7790). Available online: http://climexp.knmi.nl (accessed on 5 October 2020).
- Rome, S.; Giorgetti, J.-P. Du climat de la montagne corse et ses aléas. In Les Risques Liés au Temps et au Climat, XIXe Colloque de l’Association Internationale de Climatologie, Épernay, France, 6–9 Septembre 2006; Beltrando, G., Madelin, M.M., Quénel, H., Eds.; Association Internationale de Climatologie: Épernay, France, 2006; pp. 486–492. ISBN 2-901560-70-9. [Google Scholar]
- Médail, F.; Verlaque, R. Ecological characteristics and rarity of endemic plants from southeast France and Corsica: Implications for biodiversity conversation. Biol. Conserv. 1997, 80, 269–281. [Google Scholar] [CrossRef]
- Eilmann, B.; Rigling, A. Tree-growth analyses to estimate tree species’ drought tolerance. Tree Physiol. 2012, 32, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Pérez, L.; Villar-Salvador, P.; Martínez-Vilalta, J.; Toca, A.; Zavala, M.A. Distribution of pines in the Iberian peninsula agrees with species differences in foliage frost tolerance, not with vulnerability to freezing-induced xylem embolism. Tree Physiol. 2018, 38, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Luterbacher, J.; García-Herrera, R.; Akcer-On, S.; Allan, R.; Alvarez-Castro, M.C.; Benito, G.; Booth, J.; Büntgen, U.; Cagatay, N.; Colombaroli, D.; et al. A review of 2000 years of paleoclimatic evidence in the Mediterranean. In The Climate of the Mediterranean Region: From the Past to the Future, 1st ed.; Lionello, P., Ed.; Elsevier Insights: Amsterdam, The Netherlands, 2012; Volume 1, pp. 87–185. ISBN 978-0-12-416042-2. [Google Scholar]
- Tintner, J.; Smidt, E. Resistance of wood from Black pine (Pinus nigra var. austriaca) against weathering. J. Wood Sci. 2018, 64, 816–822. [Google Scholar] [CrossRef] [Green Version]
- Seim, A.; Treydte, K.; Trouet, V.; Frank, D.; Fonti, P.; Tegel, W.; Panayotov, M.; Fernández-Donado, L.; Krusic, P.; Büntgen, U. Climate sensitivity of Mediterranean pine growth reveals distinct east-west dipole. Int. J. Climatol. 2015, 35, 2503–2513. [Google Scholar] [CrossRef]
- Touchan, R.; Anchukaitis, K.J.; Meko, D.M.; Kerchouche, D.; Slimani, S.; Ilmen, R.; Hasnaoui, F.; Guibal, F.; Camarero, J.J.; Sánchez-Salguero, R.; et al. Climate controls on tree growth in the Western Mediterranean. Holocene 2017, 27, 1429–1442. [Google Scholar] [CrossRef]
- Esper, J.; Konter, O.; Klippel, L.; Krusic, P.J.; Büntgen, U. Pre-instrumental summer precipitation variability in northwestern Greece from a high-elevation Pinus heldreichii network. Int. J. Climatol. 2021, 41, 2828–2839. [Google Scholar] [CrossRef]
- Griggs, C.; DeGaetano, A.; Kuniholm, P.; Newton, M. A regional high-frequency reconstruction of May–June precipitation in the north Aegean from oak tree rings, A.D. 1089–1989. Int. J. Climatol. 2007, 27, 1075–1089. [Google Scholar] [CrossRef]
- Ruiz-Labourdette, D.; Génova, M.; Schmitz, M.F.; Urrutia, R.; Pineda, F.D. Summer rainfall variability in European Mediterranean mountains from the sixteenth to the twentieth century reconstructed from tree rings. Int. J. Biometeorol. 2014, 58, 1627–1639. [Google Scholar] [CrossRef] [Green Version]
- Touchan, R.; Meko, D.M.; Aloui, A. Precipitation reconstruction for northwestern Tunisia from tree rings. J. Arid Environ. 2008, 72, 1887–1896. [Google Scholar] [CrossRef]
- Cook, E.R.; Seager, R.; Kushnir, Y.; Briffa, K.R.; Büntgen, U.; Frank, D.; Krusic, P.J.; Tegel, W.; van der Schrier, G.; Andreu-Hayles, L.; et al. Old World megadroughts and pluvials during the Common Era. Sci. Adv. 2015, 1, e1500561. [Google Scholar] [CrossRef] [Green Version]
- Esper, J.; Frank, D.; Büntgen, U.; Verstege, A.; Luterbacher, J.; Xoplaki, E. Long-term drought severity variations in Morocco. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef] [Green Version]
- Klippel, L.; Krusic, P.J.; Brandes, R.; Hartl, C.; Belmecheri, S.; Dienst, M.; Esper, J. A 1286-year hydro-climate reconstruction for the Balkan peninsula. Boreas 2018, 47, 1218–1229. [Google Scholar] [CrossRef]
- Nicault, A.; Alleaume, S.; Brewer, S.; Carrer, M.; Nola, P.; Guiot, J. Mediterranean drought fluctuation during the last 500 years based on tree-ring data. Clim. Dyn. 2008, 31, 227–245. [Google Scholar] [CrossRef]
- Tejedor, E.; de Luis, M.; Cuadrat, J.M.; Esper, J.; Saz, M.A. Tree-ring-based drought reconstruction in the Iberian range (east of Spain) since 1694. Int. J. Biometeorol. 2016, 60, 361–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büntgen, U.; Frank, D.; Trouet, V.; Esper, J. Diverse climate sensitivity of Mediterranean tree-ring width and density. Trees 2010, 24, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Esper, J.; Schneider, L.; Smerdon, J.E.; Schöne, B.R.; Büntgen, U. Signals and memory in tree-ring width and density data. Dendrochronologia 2015, 35, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Esper, J.; Frank, D.; Timonen, M.; Zorita, E.; Wilson, R.; Luterbacher, J.; Holzkämper, S.; Fischer, N.; Wagner, S.; Nievergelt, D.; et al. Orbital forcing of tree-ring data. Nat. Clim. Chang. 2012, 2, 862–866. [Google Scholar] [CrossRef]
- Esper, J.; Düthorn, E.; Krusic, P.; Timonen, M.; Büntgen, U. Northern European summer temperature variations over the Common Era from integrated tree-ring density records. J. Quat. Sci. 2014, 29, 487–494. [Google Scholar] [CrossRef]
- Büntgen, U.; Frank, D.C.; Nievergelt, D.; Esper, J. Summer temperature variations in the European Alps, A.D. 755–2004. J. Clim. 2006, 19, 5606–5623. [Google Scholar] [CrossRef]
- Büntgen, U.; Krusic, P.J.; Verstege, A.; Sangüesa-Barreda, G.; Wagner, S.; Camarero, J.J.; Ljungqvist, F.C.; Zorita, E.; Oppenheimer, C.; Konter, O.; et al. New tree-ring evidence from the Pyrenees reveals western Mediterranean climate variability since medieval times. J. Clim. 2017, 30, 5295–5318. [Google Scholar] [CrossRef]
- Dorado-Liñán, I.; Büntgen, U.; González-Rouco, F.; Zorita, E.; Montavez, J.P.; Gómez-Navarro, J.J.; Brunet, M.; Heinrich, I.; Helle, G.; Gutiérrez, E. Estimating 750 years of temperature variations and uncertainties in the Pyrenees by tree-ring reconstructions and climate simulations. Clim. Past 2012, 8, 919–933. [Google Scholar] [CrossRef] [Green Version]
- Esper, J.; Hartl, C.; Tejedor, E.; de Luis, M.; Günther, B.; Büntgen, U. High-resolution temperature variability reconstructed from Black pine tree ring densities in southern Spain. Atmosphere 2020, 11, 748. [Google Scholar] [CrossRef]
- Esper, J.; Klippel, L.; Krusic, P.J.; Konter, O.; Raible, C.C.; Xoplaki, E.; Luterbacher, J.; Büntgen, U. Eastern Mediterranean summer temperatures since 730 CE from Mt. Smolikas tree-ring densities. Clim. Dyn. 2020, 54, 1367–1382. [Google Scholar] [CrossRef]
- Klippel, L.; Krusic, P.J.; Konter, O.; St. George, S.; Trouet, V.; Esper, J. A 1200+ year reconstruction of temperature extremes for the northeastern Mediterranean region. Int. J. Climatol. 2019, 39, 2336–2350. [Google Scholar] [CrossRef] [Green Version]
- Leonelli, G.; Coppola, A.; Salvatore, M.C.; Baroni, C.; Battipaglia, G.; Gentilesca, T.; Ripullone, F.; Borghetti, M.; Conte, E.; Tognetti, R.; et al. Climate signals in a multispecies tree–ring network from central and southern Italy and reconstruction of the late summer temperatures since the early 1700s. Clim. Past 2017, 13, 1451–1471. [Google Scholar] [CrossRef] [Green Version]
- Schweingruber, F.H. NOAA/WDS Paleoclimatology—Schweingruber—Col de Sorba Mount Renoso—PINI—ITRDB FRAN027. NOAA National Centers for Environmental Information. Available online: https://www.ncdc.noaa.gov/paleo-search/study/4389 (accessed on 5 October 2020).
- Björklund, J.; Von Arx, G.; Nievergelt, D.; Wilson, R.; Van den Bulcke, J.; Günther, B.; Loader, N.J.; Rydval, M.; Fonti, P.; Scharnweber, T.; et al. Scientific merits and analytical challenges of tree-ring densitometry. Rev. Geophys. 2019, 57, 1224–1264. [Google Scholar] [CrossRef] [Green Version]
- Eschbach, W.; Nogler, P.; Schär, E.; Schweingruber, F.H. Technical advances in the radiodensitometrical determination of wood density. Dendrochronologia 1995, 13, 155–168. [Google Scholar]
- Schweingruber, F.H.; Fritts, H.C.; Bräker, O.U.; Drew, L.G.; Schär, E. The X-ray technique as applied to dendroclimatology. Tree Ring Bull. 1978, 38, 61–91. [Google Scholar]
- Cook, E.R.; Krusic, P.J. Program ARSTAN, A Tree-Ring Standardization Program Based on Detrending and Autoregressive Timeseries Modeling with Interactive Graphics; Lamont-Doherty Earth Observatory, Columbia University: Palisades, NY, USA, 2005. [Google Scholar]
- Cook, E.; Peters, K. Calculating unbiased tree-ring indices for the study of climatic and environmental change. Holocene 1997, 7, 361–370. [Google Scholar] [CrossRef]
- Frank, D.; Esper, J.; Cook, E.R. Adjustment for proxy number and coherence in a large-scale temperature reconstruction. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R. A Time Series Analysis Approach to Tree-Ring Standardization. Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 1985. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Appl. Meteorol. Clim. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Esper, J.; Cook, E.R.; Krusic, P.J.; Peters, K.; Schweingruber, F.H. Tests of the RCS method for preserving low-frequency variability in long tree-ring chronologies. Tree Ring Res. 2003, 59, 81–98. [Google Scholar]
- Fang, K.; Gou, X.; Peters, K.; Li, J.; Zhang, F. Removing biological trends from tree-ring series: Testing modified Hugershoff curves. Tree Ring Res. 2010, 66, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R.; Briffa, K.R.; Meko, D.M.; Graybill, D.A.; Funkhouser, G. The ’segment length curse’ in long tree-ring chronology development for palaeoclimatic studies. Holocene 1995, 5, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Briffa, K.R.; Jones, P.D.; Bartholin, T.S.; Eckstein, D.; Schweingruber, F.H.; Karlén, W.; Zetterberg, P.; Eronen, M. Fennoscandian summers from AD 500: Temperature changes on short and long timescales. Clim. Dyn. 1992, 7, 111–119. [Google Scholar] [CrossRef]
- Esper, J.; Krusic, P.J.; Ljungqvist, F.C.; Luterbacher, J.; Carrer, M.; Cook, E.; Davi, N.K.; Hartl, C.; Kirdyanov, A.; Konter, O.; et al. Ranking of tree-ring based temperature reconstructions of the past millennium. Quat. Sci. Rev. 2016, 145, 134–151. [Google Scholar] [CrossRef] [Green Version]
- Briffa, K.R.; Osborn, T.J.; Schweingruber, F.H.; Harris, I.C.; Jones, P.D.; Shiyatov, S.G.; Vaganov, E.A. Low-frequency temperature variations from a northern tree ring density network. J. Geophys. Res. Atmos. 2001, 106, 2929–2941. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Schneider, U.; Finger, P.; Rustemeier, E.; Ziese, M.; Becker, A. Global Precipitation Analysis Products of the GPCC; Deutscher Wetterdienst (DWD), Abt. Hydrometeorologie, Weltzentrum für Niederschlagsklimatologie: Offenbach am Main, Germany, 2021. [Google Scholar]
- Trouet, V.; Van Oldenborgh, G.J. KNMI climate explorer: A web-based research tool for high-resolution paleoclimatology. Tree Ring Res. 2013, 69, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Météo-France Climate Data from Calacuccia Weather Station (20224). Available online: https://meteofrance.com/previsions-meteo-france/calacuccia/20224 (accessed on 7 January 2021).
- Schweingruber, F.H. NOAA/WDS Paleoclimatology—Schweingruber—Sierra de Crispo—PILE—ITRDB ITAL015. NOAA National Centers for Environmental Information. Available online: https://www.ncdc.noaa.gov/paleo-search/study/4644 (accessed on 5 October 2020).
- Esper, J.; Cook, E.R.; Schweingruber, F.H. Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. Science 2002, 295, 2250–2253. [Google Scholar] [CrossRef] [Green Version]
- Briffa, K.R.; Jones, P.D.; Schweingruber, F.H.; Karlen, W.; Shiyatov, S.G. Tree-ring variables as proxy-climate indicators: Problems with low-frequency signals. In Climatic Variations and Forcing Mechanisms of the Last 2000 Years, 1st ed.; Jones, P.D., Bradley, R.S., Jouzel, J., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; Volume 41, pp. 9–41. ISBN 978-3-642-61113-1. [Google Scholar]
- Frank, D.; Esper, J. Temperature reconstructions and comparisons with instrumental data from a tree-ring network for the European Alps. Int. J. Climatol. 2005, 25, 1437–1454. [Google Scholar] [CrossRef]
- Esper, J.; Büntgen, U.; Frank, D.; Nievergelt, D.; Treydte, K.; Verstege, A. Multiple tree-ring parameters from Atlas cedar (Morocco) and their climatic signal. In Schriften des Forschungszentrums Jülich, Reihe Umwelt: TRACE, Tree Rings in Archaeology, Climatology and Ecology, 4th ed.; Heidrich, I., Gärtner, H., Monbaron, M., Schleser, G., Eds.; Forschungszentrum Jülich: Jülich, Germany, 2006; Volume 61, pp. 46–55. ISBN 978-3-89336-425-1. [Google Scholar]
- Camarero, J.J.; Olano, J.M.; Parra, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef]
- De Micco, V.; Campelo, F.; De Luis, M.; Bräuning, A.; Grabner, M.; Battipaglia, G.; Cherubini, P. Intra-annual density fluctuations in tree rings: How, when, where, and why? IAWA J. 2016, 37, 232–259. [Google Scholar] [CrossRef]
- Trouet, V. A tree-ring based late summer temperature reconstruction (AD 1675–1980) for the northeastern Mediterranean. Radiocarbon 2014, 56, 69–78. [Google Scholar] [CrossRef]
- Szymczak, S.; Häusser, M.; Garel, E.; Santoni, S.; Huneau, F.; Knerr, I.; Trachte, K.; Bendix, J.; Bräuning, A. How do Mediterranean pine trees respond to drought and precipitation events along an elevation gradient? Forests 2020, 11, 758. [Google Scholar] [CrossRef]
- Hetzer, T.; Bräuning, A.; Leuschner, H.H. High-resolution climatic analysis of wood anatomical features in Corsican pine from Corsica (France) using latewood tracheid profiles. Trees 2014, 28, 1279–1288. [Google Scholar] [CrossRef]
- Martín-Benito, D.; Beeckman, H.; Cañellas, I. Influence of drought on tree rings and tracheid features of Pinus nigra and Pinus sylvestris in a mesic Mediterranean forest. Eur. J. For. Res. 2013, 132, 33–45. [Google Scholar] [CrossRef]
- Petrucco, L.; Nardini, A.; von Arx, G.; Saurer, M.; Cherubini, P. Isotope signals and anatomical features in tree rings suggest a role for hydraulic strategies in diffuse drought-induced dieback of Pinus nigra. Tree Physiol. 2017, 37, 523–535. [Google Scholar] [CrossRef]
- Pittermann, J.; Sperry, J.S.; Wheeler, J.K.; Hacke, U.G.; Sikkema, E.H. Mechanical reinforcement of tracheids compromises the hydraulic efficiency of conifer xylem. Plant Cell Environ. 2006, 29, 1618–1628. [Google Scholar] [CrossRef] [Green Version]
- Fonti, P.; von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef]
- Piermattei, A.; Crivellaro, A.; Carrer, M.; Urbinati, C. The “blue ring”: Anatomy and formation hypothesis of a new tree-ring anomaly in conifers. Trees 2015, 29, 613–620. [Google Scholar] [CrossRef]
- Lebourgeois, F. Climatic signals in earlywood, latewood and total ring width of Corsican pine from western France. Ann. For. Sci. 2000, 57, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Gričar, J.; Čufar, K.; Oven, P.; Schmitt, U. Differentiation of terminal latewood tracheids in silver fir trees during autumn. Ann. Bot. 2005, 95, 959–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl-Meier, C.; Dittmar, C.; Zang, C.; Rothe, A. Mountain forest growth response to climate change in the northern limestone Alps. Trees 2014, 28, 819–829. [Google Scholar] [CrossRef]
- Häusser, M.; Szymczak, S.; Garel, E.; Santoni, S.; Huneau, F.; Bräuning, A. Growth variability of two native pine species on Corsica as a function of elevation. Dendrochronologia 2019, 54, 49–55. [Google Scholar] [CrossRef]
- Szymczak, S.; Bräuning, A.; Häusser, M.; Garel, E.; Huneau, F.; Santoni, S. The relationship between climate and the intra-annual oxygen isotope patterns from pine trees: A case study along an elevation gradient on Corsica, France. Ann. For. Sci. 2019, 76, 1–76. [Google Scholar] [CrossRef]
- Briffa, K.R.; Schweingruber, F.H.; Jones, P.D.; Osborn, T.J.; Shiyatov, S.G.; Vaganov, E.A. Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature 1998, 391, 678–682. [Google Scholar] [CrossRef]
- Briffa, K.R.; Osborn, T.J.; Schweingruber, F.H. Large-scale temperature inferences from tree-rings: A review. Glob. Planet. Chang. 2004, 40, 11–26. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Wilson, R.; Liepert, B.; Cherubini, P. On the ‘divergence problem’ in northern forests: A review of the tree-ring evidence and possible causes. Glob. Planet. Chang. 2008, 60, 289–305. [Google Scholar] [CrossRef]
- Schweingruber, F.H.; Briffa, K.R. Tree-ring density networks for climate reconstruction. In Climatic Variations and Forcing Mechanisms of the Last 2000 Years, 1st ed.; Jones, P.D., Bradley, R.S., Jouzel, J., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; Volume 41, pp. 43–66. ISBN 978-3-642-61113-1. [Google Scholar]
- Laffoley, D.; Baxter, J.; Pergent-Martini, C.; Pergent, G.; Otero, M.M.; Simard, F. Climate Change and the Marine Environment in Corsica, Report Card 2018; International Union for Conservation of Nature (IUCN): Gland, Switzerland, 2018. [Google Scholar]
- Cook, B.I.; Anchukaitis, K.J.; Touchan, R.; Meko, D.M.; Cook, E.R. Spatiotemporal drought variability in the Mediterranean over the last 900 years. J. Geophys. Res. Atmos. 2016, 121, 2060–2074. [Google Scholar] [CrossRef]
- Spinoni, J.; Naumann, G.; Vogt, J.V.; Barbosa, P. The biggest drought events in Europe from 1950 to 2012. J. Hydrol. Reg. Stud. 2015, 3, 509–524. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulías, J.; Flexas, J. Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Lévy, G.; Aussenac, G.; Clerc, B.; Willm, F. Influence of soil drying on leaf water potential, photosynthesis, stomatal conductance and growth in two black pine varieties. Ann. Sci. For. 1998, 55, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Hartl-Meier, C.; Zang, C.; Büntgen, U.; Esper, J.; Rothe, A.; Göttlein, A.; Dirnböck, T.; Treydte, K. Uniform climate sensitivity in tree-ring stable isotopes across species and sites in a mid-latitude temperate forest. Tree Physiol. 2015, 35, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Olano, J.M.; Linares, J.C.; García-Cervigón, A.I.; Arzac, A.; Delgado, A.; Rozas, V. Drought-induced increase in water-use efficiency reduces secondary tree growth and tracheid wall thickness in a Mediterranean conifer. Oecologia 2014, 176, 273–283. [Google Scholar] [CrossRef]
- Büntgen, U.; Frank, D.; Neuenschwander, T.; Esper, J. Fading temperature sensitivity of Alpine tree growth at its Mediterranean margin and associated effects on large-scale climate reconstructions. Clim. Chang. 2012, 114, 651–666. [Google Scholar] [CrossRef] [Green Version]
- Castagneri, D.; Nola, P.; Motta, R.; Carrer, M. Summer climate variability over the last 250 years differently affected tree species radial growth in a mesic Fagus–Abies–Picea old-growth forest. For. Ecol. Manag. 2014, 320, 21–29. [Google Scholar] [CrossRef]
- Galván, J.D.; Camarero, J.J.; Ginzler, C.; Büntgen, U. Spatial diversity of recent trends in Mediterranean tree growth. Environ. Res. Lett. 2014, 9, 084001. [Google Scholar] [CrossRef] [Green Version]
- Martín-Benito, D.; del Río, M.; Cañellas, I. Black pine (Pinus nigra Arn.) growth divergence along a latitudinal gradient in western Mediterranean mountains. Ann. For. Sci. 2010, 67, 401. [Google Scholar] [CrossRef]
- Thiel, D.; Nagy, L.; Beierkuhnlein, C.; Huber, G.; Jentsch, A.; Konnert, M.; Kreyling, J. Uniform drought and warming responses in Pinus nigra provenances despite specific overall performances. For. Ecol. Manag. 2012, 270, 200–208. [Google Scholar] [CrossRef]
- Mouillot, F.; Ratte, J.-P.; Joffre, R.; Moreno, J.M.; Rambal, S. Some determinants of the spatio-temporal fire cycle in a mediterranean landscape (Corsica, France). Landsc. Ecol. 2003, 18, 665–674. [Google Scholar] [CrossRef]
- Soulères, O. Les incendies de Haute-Corse. Rev. For. Fr. 2000, 52, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Fulé, P.Z.; Ribas, M.; Gutiérrez, E.; Vallejo, R.; Kaye, M.W. Forest structure and fire history in an old Pinus nigra forest, eastern Spain. For. Ecol. Manag. 2008, 255, 1234–1242. [Google Scholar] [CrossRef]
- Szymczak, S.; Bräuning, A.; Häusser, M.; Garel, E.; Huneau, F.; Santoni, S. A dendroecological fire history for central Corsica/France. Tree Ring Res. 2020, 76, 40–53. [Google Scholar] [CrossRef]
- Touchan, R.; Baisan, C.; Mitsopoulos, I.D.; Dimitrakopoulos, A.P. Fire history in European black pine (Pinus nigra Arn.) forests of the Valia Kalda, Pindus mountains, Greece. Tree Ring Res. 2012, 68, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Llovet, J.; Rodrigo, A.; Vallejo, R. Are wildfires a disaster in the Mediterranean basin?—A review. Int. J. Wildland Fire 2008, 17, 713–723. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
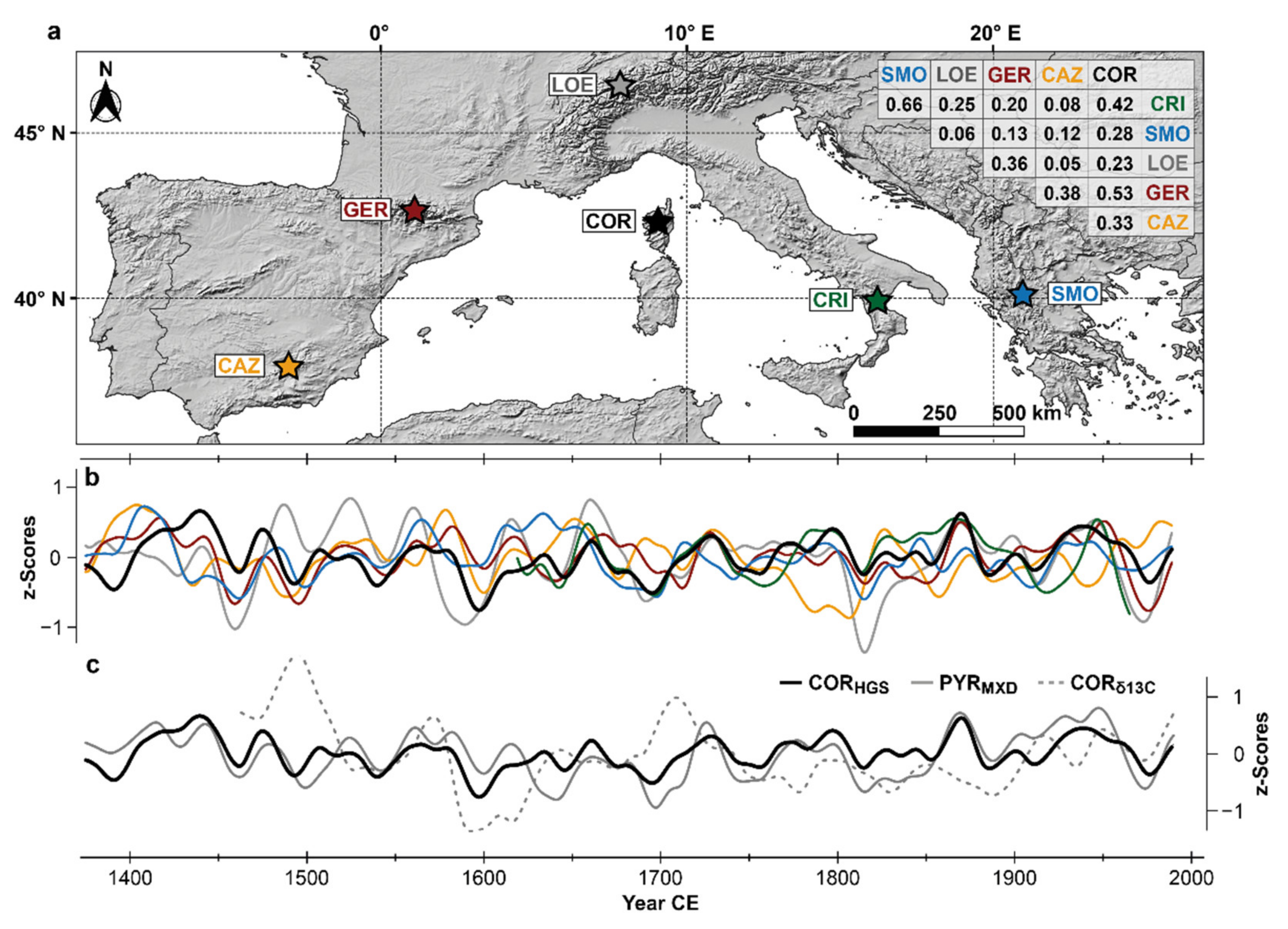
| Code | Site | Species | Radii | Period | Season | Correlation | Distance |
|---|---|---|---|---|---|---|---|
| CAZ | Cazorla | PINI | 88 | 1288–2014 | FMAM and SO | 0.53|0.45 | 1150 km |
| COR | Corsica | PINI | 69 | 1360–2016 | AMJJ and SO | 0.56|0.16 | 0 km |
| CRI | Serra di Crispo | PIHE | 21 | 1604–1980 | JAS | 0.60|0.60 | 650 km |
| GER | Gerber | PIUN | 414 | 1095–2014 | MJ and AS | 0.59|0.38 | 650 km |
| LOE | Loetschental | LADE | 180 | 748–2004 | JJAS | 0.77|0.78 | 450 km |
| SMO | Mt. Smolikas | PIHE | 192 | 672–2017 | AS | 0.52|0.56 | 1050 km |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Römer, P.; Hartl, C.; Schneider, L.; Bräuning, A.; Szymczak, S.; Huneau, F.; Lebre, S.; Reinig, F.; Büntgen, U.; Esper, J. Reduced Temperature Sensitivity of Maximum Latewood Density Formation in High-Elevation Corsican Pines under Recent Warming. Atmosphere 2021, 12, 804. https://doi.org/10.3390/atmos12070804
Römer P, Hartl C, Schneider L, Bräuning A, Szymczak S, Huneau F, Lebre S, Reinig F, Büntgen U, Esper J. Reduced Temperature Sensitivity of Maximum Latewood Density Formation in High-Elevation Corsican Pines under Recent Warming. Atmosphere. 2021; 12(7):804. https://doi.org/10.3390/atmos12070804
Chicago/Turabian StyleRömer, Philipp, Claudia Hartl, Lea Schneider, Achim Bräuning, Sonja Szymczak, Frédéric Huneau, Sébastien Lebre, Frederick Reinig, Ulf Büntgen, and Jan Esper. 2021. "Reduced Temperature Sensitivity of Maximum Latewood Density Formation in High-Elevation Corsican Pines under Recent Warming" Atmosphere 12, no. 7: 804. https://doi.org/10.3390/atmos12070804