
The Influence of Ozone on Net Ecosystem Production of a Ryegrass–Clover Mixture under Field Conditions

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
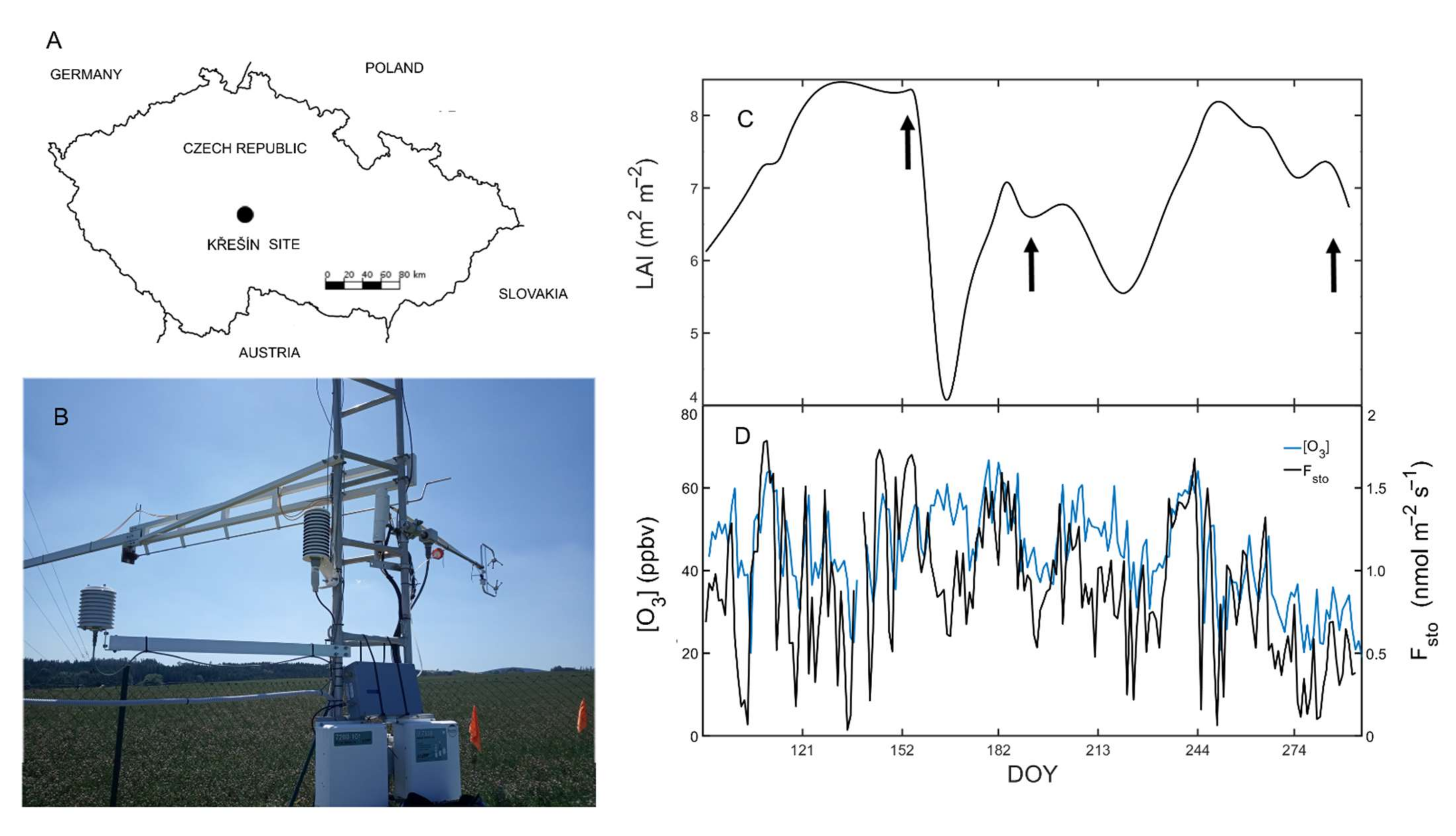
2.1. Experimental Site Description
2.2. Measurement of Microclimatic Parameters
2.3. Eddy Covariance Flux Measurements
2.4. Modelling of Stomatal O3 Flux
2.5. Modelling and Statistical Analyses
3. Results
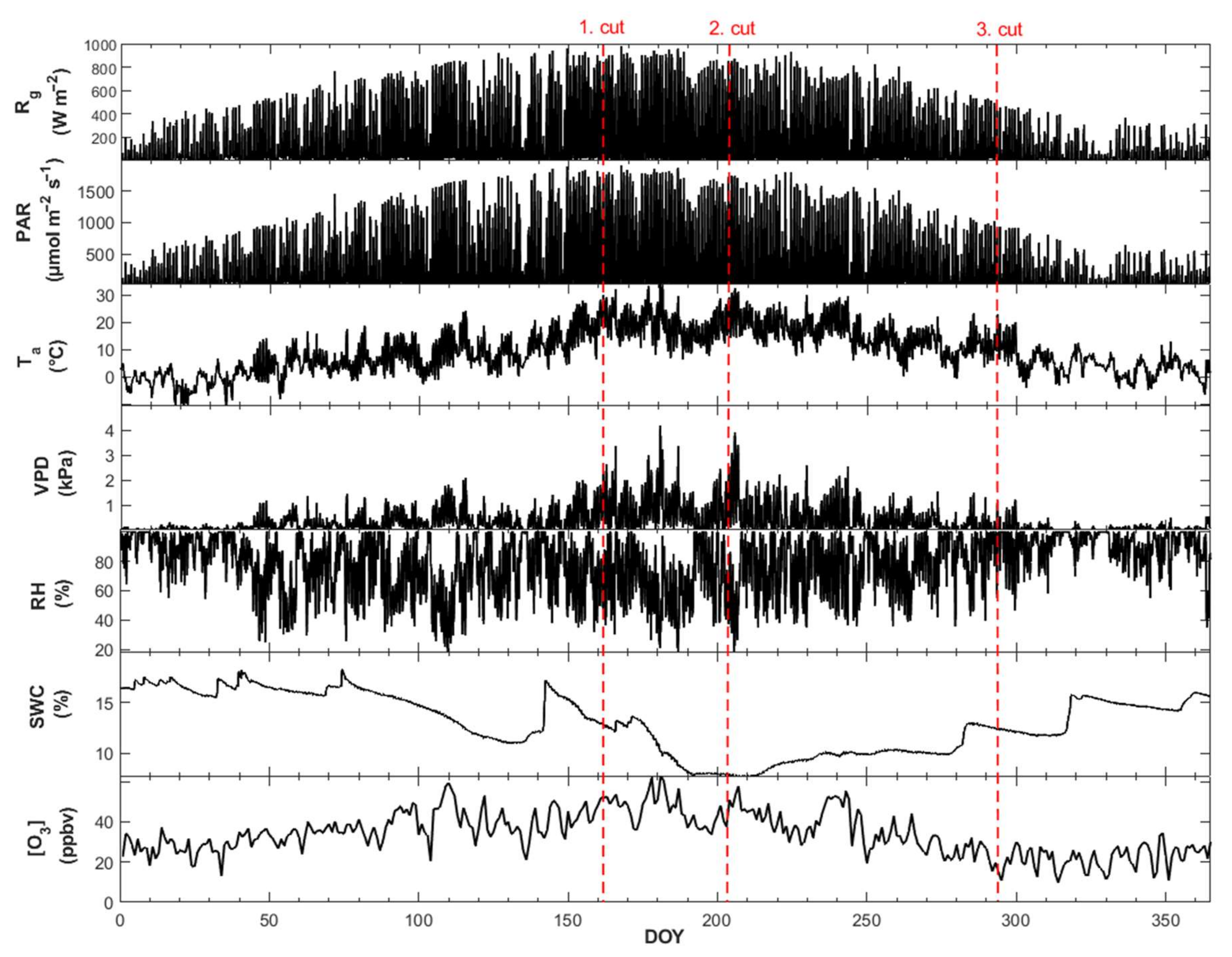
3.1. Microclimatic Growth Conditions
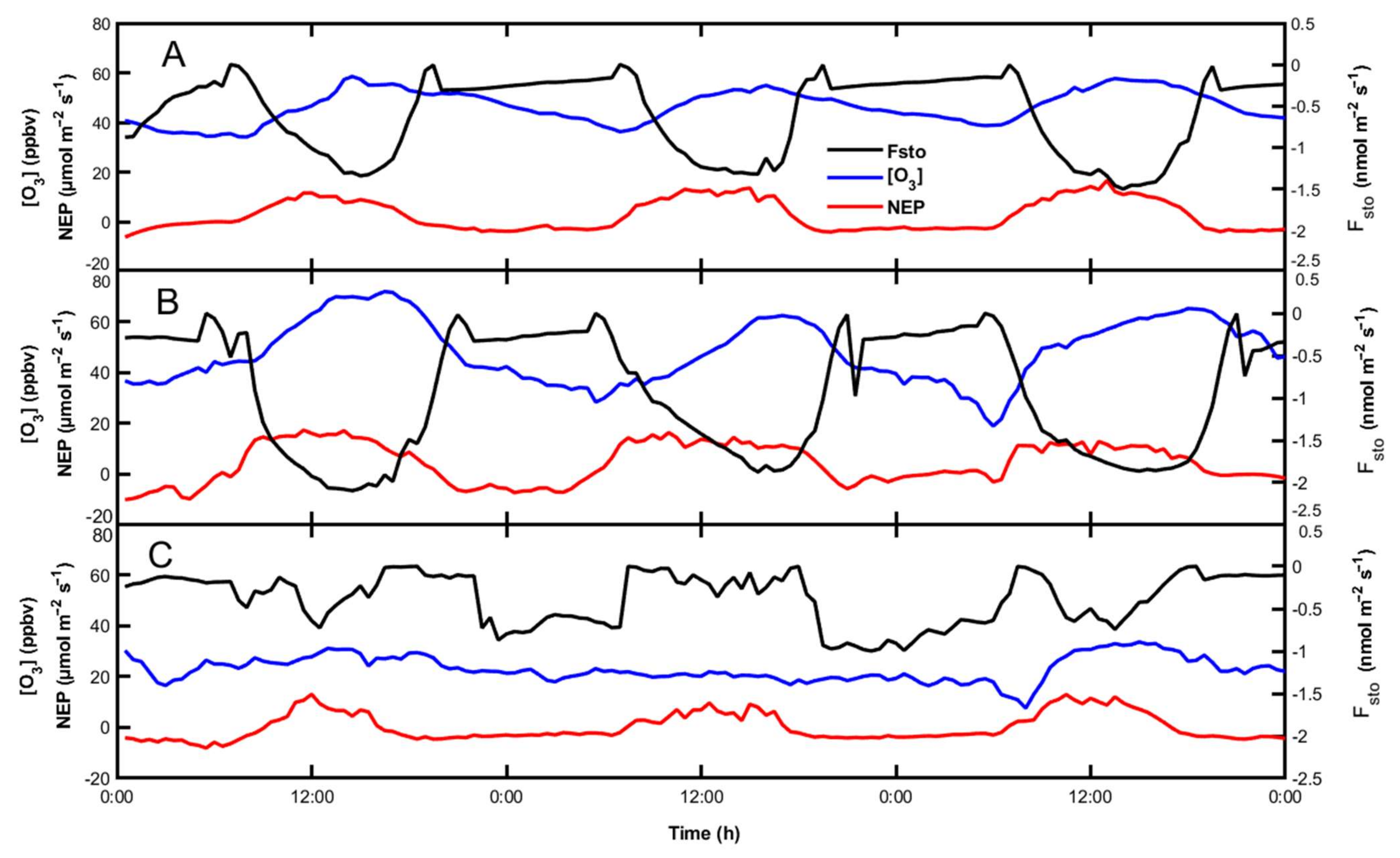
3.2. Daily Courses of [O3], Fsto and NEP
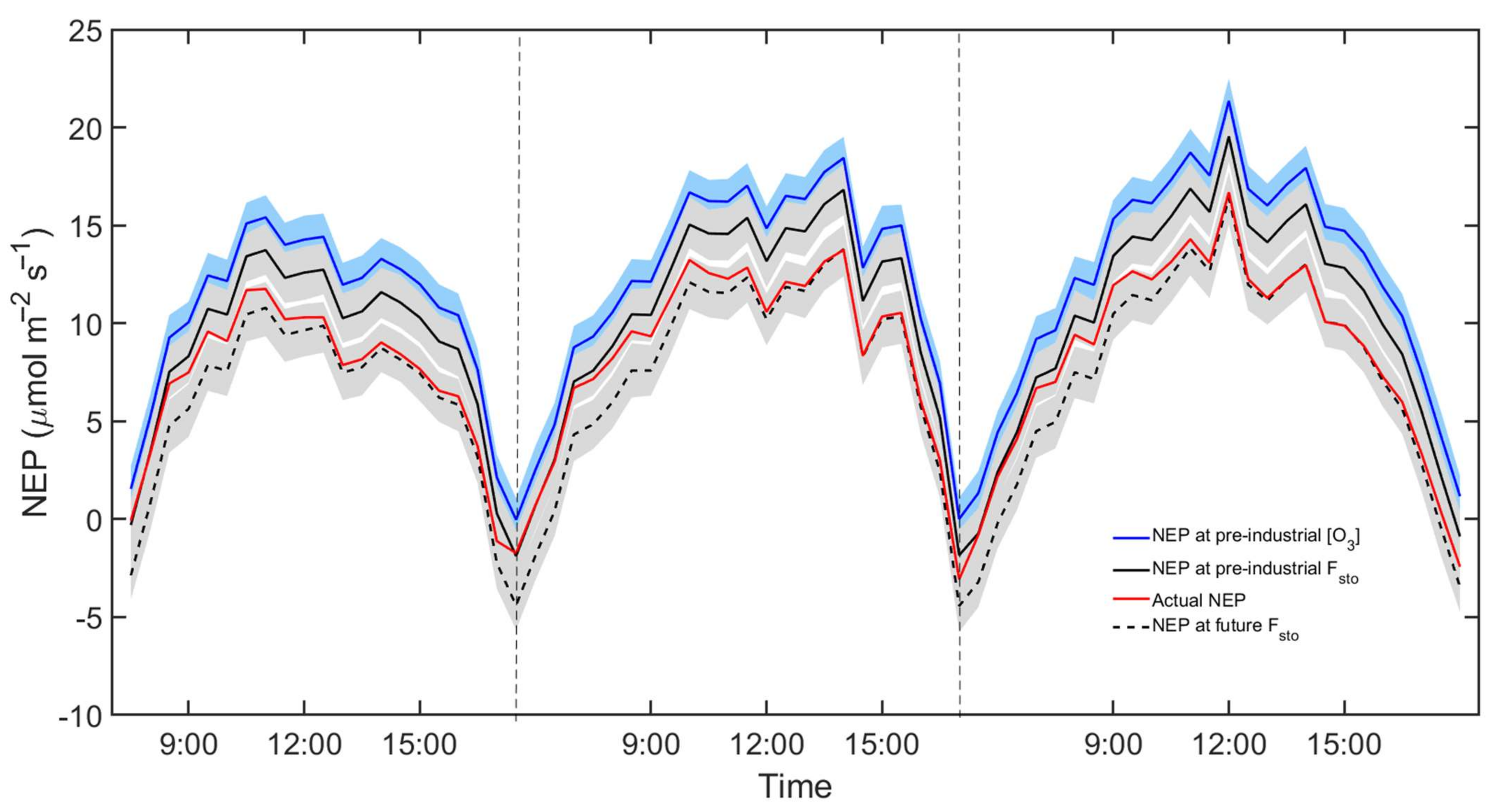
3.3. NEP Modelling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monks, P.S.; Granier, C.; Fuzzi, S.; Stohl, A.; Williams, M.L.; Akimoto, H.; Amann, M.; Baklanov, A.; Baltensperger, U.; Bey, I. Atmospheric composition change–global and regional air quality. Atmos. Environ. 2009, 43, 5268–5350. [Google Scholar] [CrossRef] [Green Version]
- Juráň, S.; Grace, J.; Urban, O. Temporal changes in ozone concentrations and their impact on vegetation. Atmosphere 2021, 12, 82. [Google Scholar] [CrossRef]
- Cooper, O.R.; Parrish, D.; Ziemke, J.; Balashov, N.; Cupeiro, M.; Galbally, I.; Gilge, S.; Horowitz, L.; Jensen, N.; Lamarque, J.-F. Global distribution and trends of tropospheric ozone: An observation-based reviewGlobal distribution and trends of tropospheric ozone. Elementa 2014, 2, 000029. [Google Scholar] [CrossRef]
- Dentener, F.; Stevenson, D.; Ellingsen, K.; van Noije, T.; Schultz, M.; Amann, M.; Atherton, C.; Bell, N.; Bergmann, D.; Bey, I. The global atmospheric environment for the next generation. Environ. Sci. Technol. 2006, 40, 3586–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granier, C.; Bessagnet, B.; Bond, T.; D’Angiola, A.; van Der Gon, H.D.; Frost, G.J.; Heil, A.; Kaiser, J.W.; Kinne, S.; Klimont, Z. Evolution of anthropogenic and biomass burning emissions of air pollutants at global and regional scales during the 1980–2010 period. Clim. Change 2011, 109, 163–190. [Google Scholar] [CrossRef]
- Agyei, T.; Juráň, S.; Ofori-Amanfo, K.K.; Šigut, L.; Urban, O.; Marek, M.V. The impact of drought on total ozone flux in a mountain Norway spruce forest. J. For. Sci. 2020, 66, 278–280. [Google Scholar] [CrossRef]
- Hůnová, I.; Livorová, H.; Ostatnická, J. Potential ambient ozone impact on ecosystems in the Czech Republic as indicated by exposure index AOT40. Eco. Indic. 2003, 3, 35–47. [Google Scholar] [CrossRef]
- Tai, A.P.; Martin, M.V.; Heald, C.L. Threat to future global food security from climate change and ozone air pollution. Nat. Clim. Change 2014, 4, 817–821. [Google Scholar] [CrossRef] [Green Version]
- Mills, G.; Hayes, F.; Simpson, D.; Emberson, L.; Norris, D.; Harmens, H.; Bueker, P. Evidence of widespread effects of ozone on crops and (semi-) natural vegetation in Europe (1990–2006) in relation to AOT40-and flux-based risk maps. Glob. Change Biol. 2011, 17, 592–613. [Google Scholar] [CrossRef] [Green Version]
- Mills, G.; Hayes, F.; Jones, M.; Cinderby, S. Identifying ozone-sensitive communities of (semi-) natural vegetation suitable for mapping exceedance of critical levels. Environ. Pollut. 2007, 146, 736–743. [Google Scholar] [CrossRef]
- Gandin, A.; Davrinche, A.; Jolivet, Y. Deciphering the main determinants of O3 tolerance in Euramerican poplar genotypes. Sci. Total Environ. 2019, 656, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Zapletal, M.; Juran, S.; Krpes, V.; Michna, K.; Cudlin, P.; Edwards, M. Effect of ozone flux on selected structural and antioxidant characteristics of a mountain norway spruce forest. Balt. For. 2018, 24, 261–267. [Google Scholar]
- Wittig, V.E.; Ainsworth, E.A.; Naidu, S.L.; Karnosky, D.F.; Long, S.P. Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: A quantitative meta-analysis. Glob. Change Biol. 2009, 15, 396–424. [Google Scholar] [CrossRef]
- Goumenaki, E.; Taybi, T.; Borland, A.; Barnes, J. Mechanisms underlying the impacts of ozone on photosynthetic performance. Environ. Exp. Bot. 2010, 69, 259–266. [Google Scholar] [CrossRef]
- Hoshika, Y.; Watanabe, M.; Inada, N.; Koike, T. Model-based analysis of avoidance of ozone stress by stomatal closure in Siebold’s beech (Fagus crenata). Ann. Bot. 2013, 112, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Di Baccio, D.; Castagna, A.; Paoletti, E.; Sebastiani, L.; Ranieri, A. Could the differences in O3 sensitivity between two poplar clones be related to a difference in antioxidant defense and secondary metabolic response to O3 influx? Tree Physiol. 2008, 28, 1761–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grantz, D.; Gunn, S.; Vu, H.B. O3 impacts on plant development: A meta-analysis of root/shoot allocation and growth. Plant Cell Environ. 2006, 29, 1193–1209. [Google Scholar] [CrossRef]
- Rodwell, J. British Plant Communities: Grasslands and Montane Communities; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Veen, P.; Jefferson, R.; De Smidt, J.; van der Straaten, J. Grasslands in Europe: Of High Nature Value; KNNV Publishing: Zeist, The Netherlands, 2009. [Google Scholar]
- Wilbourn, S.; Davison, A.; Ollerenshaw, J. The use of an unenclosed field fumigation system to determine the effects of elevated ozone on a grass–clover mixture. New Phytol. 1995, 129, 23–32. [Google Scholar] [CrossRef]
- Nussbaum, S.; Bungener, P.; Geissmann, M.; Fuhrer, J. Plant–plant interactions and soil moisture might be important in determining ozone impacts on grasslands. New Phytol. 2000, 147, 327–335. [Google Scholar] [CrossRef]
- Hayes, F.; Mills, G.; Ashmore, M. Effects of ozone on inter-and intra-species competition and photosynthesis in mesocosms of Lolium perenne and Trifolium repens. Environ. Pollut. 2009, 157, 208–214. [Google Scholar] [CrossRef]
- Evans, P.A.; Ashmore, M. The effects of ambient air on a semi-natural grassland community. Agric. Ecosyst. Environ. 1992, 38, 91–97. [Google Scholar] [CrossRef]
- Ren, W.; Tian, H.; Tao, B.; Chappelka, A.; Sun, G.; Lu, C.; Liu, M.; Chen, G.; Xu, X. Impacts of tropospheric ozone and climate change on net primary productivity and net carbon exchange of China's forest ecosystems. Glob. Ecol. Biogeogr. 2011, 20, 391–406. [Google Scholar] [CrossRef]
- Oliver, R.J.; Mercado, L.M.; Sitch, S.; Simpson, D.; Medlyn, B.E.; Lin, Y.-S.; Folberth, G.A. Large but decreasing effect of ozone on the European carbon sink. Biogeosciences 2018, 15, 4245–4269. [Google Scholar] [CrossRef] [Green Version]
- Felzer, B.; Kicklighter, D.; Melillo, J.; Wang, C.; Zhuang, Q.; Prinn, R. Effects of ozone on net primary production and carbon sequestration in the conterminous United States using a biogeochemistry model. Tellus B Chem. Phys. Meteorol. 2004, 56, 230–248. [Google Scholar] [CrossRef]
- Ren, W.; Tian, H.; Liu, M.; Zhang, C.; Chen, G.; Pan, S.; Felzer, B.; Xu, X. Effects of tropospheric ozone pollution on net primary productivity and carbon storage in terrestrial ecosystems of China. J. Geophys. Res. Atmos. 2007, 112, D22S09. [Google Scholar] [CrossRef]
- Felzer, B.S.; Cronin, T.; Reilly, J.M.; Melillo, J.M.; Wang, X. Impacts of ozone on trees and crops. Comptes Rendus Geosci. 2007, 339, 784–798. [Google Scholar] [CrossRef] [Green Version]
- Cailleret, M.; Ferretti, M.; Gessler, A.; Rigling, A.; Schaub, M. Ozone effects on European forest growth—Towards an integrative approach. J. Ecol. 2018, 106, 1377–1389. [Google Scholar] [CrossRef]
- Nussbaum, S.; Remund, J.; Rihm, B.; Mieglitz, K.; Gurtz, J.; Fuhrer, J. High-resolution spatial analysis of stomatal ozone uptake in arable crops and pastures. Environ. Int. 2003, 29, 385–392. [Google Scholar] [CrossRef]
- Hayes, F.; Wagg, S.; Mills, G.; Wilkinson, S.; Davies, W. Ozone effects in a drier climate: Implications for stomatal fluxes of reduced stomatal sensitivity to soil drying in a typical grassland species. Glob. Change Biol. 2012, 18, 948–959. [Google Scholar] [CrossRef] [Green Version]
- Juráň, S.; Šigut, L.; Holub, P.; Fares, S.; Klem, K.; Grace, J.; Urban, O. Ozone flux and ozone deposition in a mountain spruce forest are modulated by sky conditions. Sci. Total Environ. 2019, 672, 296–304. [Google Scholar] [CrossRef]
- Xu, J.; Zheng, Y.; He, Y.; Zhu, F.; Mai, B.; Wang, S.; Zhang, M.; Zhao, X.; Wang, L.; Xu, L.; et al. Estimating stomatal conductance and partitioning total ozone uptake over a winter wheat field. Atmos. Pollut. Res. 2019, 10, 904–912. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Yendrek, C.R.; Sitch, S.; Collins, W.J.; Emberson, L.D. The effects of tropospheric ozone on net primary productivity and implications for climate change. Annu. Rev. Plant Biol. 2012, 63, 637–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.; Mills, G.; Illidge, R.; Davies, W.J. How is ozone pollution reducing our food supply? J. Exp. Bot. 2012, 63, 527–536. [Google Scholar] [CrossRef]
- Long, S.P. Virtual Special Issue on food security–greater than anticipated impacts of near-term global atmospheric change on rice and wheat. Glob. Change Biol. 2012, 18, 1489–1490. [Google Scholar] [CrossRef]
- Emberson, L.; Ashmore, M.; Cambridge, H.; Simpson, D.; Tuovinen, J.-P. Modelling stomatal ozone flux across Europe. Environ. Pollut. 2000, 109, 403–413. [Google Scholar] [CrossRef]
- Dvorská, A.; Sedlák, P.; Schwarz, J.; Fusek, M.; Hanuš, V.; Vodička, P.; Trusina, J. Atmospheric station Křešín u Pacova, Czech Republic–a Central European research infrastructure for studying greenhouse gases, aerosols and air quality. Adv. Sci. Res. 2015, 12, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Integrated Carbon Observation System Home Page. Available online: https://www.icos-cp.eu (accessed on 24 September 2021).
- Global Mercury Observation System. Available online: http://www.gmos.eu (accessed on 27 September 2021).
- Aerosol, Clouds, and Trace Gases Research Infrastructure Network. Available online: https://www.actris.eu (accessed on 25 September 2021).
- Tan, C.-W.; Zhang, P.-P.; Zhou, X.-X.; Wang, Z.-X.; Xu, Z.-Q.; Mao, W.; Li, W.-X.; Huo, Z.-Y.; Guo, W.-S.; Yun, F. Quantitative monitoring of leaf area index in wheat of different plant types by integrating NDVI and Beer-Lambert law. Sci. Rep. 2020, 10, 929. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Buck, A.L. New equations for computing vapor pressure and enhancement factor. J. Appl. Meteorol. Climatol. 1981, 20, 1527–1532. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Zhao, F.; Voss, L.; Xu, L.; Sun, X.; Yu, G.; Meixner, F.X. The effects of different calibration and frequency response correction methods on eddy covariance ozone flux measured with a dry chemiluminescence analyzer. Agric. For. Meteorol. 2015, 213, 114–125. [Google Scholar] [CrossRef]
- OpenEddy Software. Available online: https://github.com/lsigut/openeddy (accessed on 26 September 2021).
- Mauder, M.; Cuntz, M.; Drüe, C.; Graf, A.; Rebmann, C.; Schmid, H.P.; Schmidt, M.; Steinbrecher, R. A strategy for quality and uncertainty assessment of long-term eddy-covariance measurements. Agric. For. Meteorol. 2013, 169, 122–135. [Google Scholar] [CrossRef]
- Foken, T.; Wichura, B. Tools for quality assessment of surface-based flux measurements. Agric. For. Meteorol. 1996, 78, 83–105. [Google Scholar] [CrossRef]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Change Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Liu, S.; Lu, L.; Mao, D.; Jia, L. Evaluating parameterizations of aerodynamic resistance to heat transfer using field measurements. Hydrol. Earth Syst. Sci. 2007, 11, 769–783. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2013. Available online: https://www.R-project.org/ (accessed on 21 October 2021).
- Knowles, J.; Frederick, C.; Whitworth, A. Tools for Analyzing Mixed Effect Regression Models; The Comprehensive R Archive Network (CRAN): Vienna, Austria, 2020. [Google Scholar]
- Juráň, S.; Edwards-Jonášová, M.; Cudlín, P.; Zapletal, M.; Šigut, L.; Grace, J.; Urban, O. Prediction of ozone effects on net ecosystem production of Norway spruce forest. iForest Biogeosci. For. 2018, 11, 743. [Google Scholar] [CrossRef] [Green Version]
- Hayes, F.; Mills, G.; Alonso, R.; González-Fernández, I.; Coyle, M.; Grünhage, L.; Gerosa, G.; Karlsson, P.E.; Marzuoli, R. A site-specific analysis of the implications of a changing ozone profile and climate for stomatal ozone fluxes in Europe. Water Air Soil Pollut. 2019, 230, 4. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhao, P.; Niu, J.; Sun, Z.; Zhu, L.; Ni, G. Canopy stomatal uptake of NOX, SO2 and O3 by mature urban plantations based on sap flow measurement. Atmos. Environ. 2016, 125, 165–177. [Google Scholar] [CrossRef]
- Hoshika, Y.; Fares, S.; Savi, F.; Gruening, C.; Goded, I.; De Marco, A.; Sicard, P.; Paoletti, E. Stomatal conductance models for ozone risk assessment at canopy level in two Mediterranean evergreen forests. Agric. For. Meteorol. 2017, 234, 212–221. [Google Scholar] [CrossRef]
- Emberson, L.D.; Büker, P.; Ashmore, M.R. Assessing the risk caused by ground level ozone to European forest trees: A case study in pine, beech and oak across different climate regions. Environ. Pollut. 2007, 147, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Schneuwly, J.; Ammann, C. Large regional differences of soil water limitation effect on ozone induced yield loss for wheat and potato in Switzerland. Sci. Total Environ. 2020, 718, 135257. [Google Scholar] [CrossRef] [PubMed]
- Otu-Larbi, F.; Conte, A.; Fares, S.; Wild, O.; Ashworth, K. Current and future impacts of drought and ozone stress on Northern Hemisphere forests. Glob. Change Biol. 2020, 26, 6218–6234. [Google Scholar] [CrossRef]
- Franks, P.J.; Farquhar, G.D. The mechanical diversity of stomata and its significance in gas-exchange control. Plant Physiol. 2007, 143, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Hoshika, Y.; Omasa, K.; Paoletti, E. Both ozone exposure and soil water stress are able to induce stomatal sluggishness. Environ. Exp. Bot. 2013, 88, 19–23. [Google Scholar] [CrossRef]
- Yang, X.; Wu, J.; Chen, X.; Ciais, P.; Maignan, F.; Yuan, W.; Piao, S.; Yang, S.; Gong, F.; Su, Y.; et al. A comprehensive framework for seasonal controls of leaf abscission and productivity in evergreen broadleaved tropical and subtropical forests. Innovation 2021, 2, 100154. [Google Scholar] [CrossRef]
- Monks, P.S. A review of the observations and origins of the spring ozone maximum. Atmos. Environ. 2000, 34, 3545–3561. [Google Scholar] [CrossRef]
- Emberson, L.D.; Pleijel, H.; Ainsworth, E.A.; van den Berg, M.; Ren, W.; Osborne, S.; Mills, G.; Pandey, D.; Dentener, F.; Büker, P. Ozone effects on crops and consideration in crop models. Eur. J. Agron. 2018, 100, 19–34. [Google Scholar] [CrossRef]
- Feng, Z.; Kobayashi, K. Assessing the impacts of current and future concentrations of surface ozone on crop yield with meta-analysis. Atmos. Environ. 2009, 43, 1510–1519. [Google Scholar] [CrossRef]
- Anav, A.; Menut, L.; Khvorostyanov, D.; Viovy, N. Impact of tropospheric ozone on the Euro-Mediterranean vegetation. Glob. Change Biol. 2011, 17, 2342–2359. [Google Scholar] [CrossRef]
- Luwe, M.; Heber, U. Ozone detoxification in the apoplasm and symplasm of spinach, broad bean and beech leaves at ambient and elevated concentrations of ozone in air. Planta 1995, 197, 448–455. [Google Scholar] [CrossRef]
- Kohut, R.J.; Laurence, J.A.; Amundson, R.G. Effects of Ozone and Sulfur Dioxide on Yield of Red Clover and Timothy. J. Environ. Qual. 1988, 17, 580–585. [Google Scholar] [CrossRef]
- Hewitt, D.; Mills, G.; Hayes, F.; Wilkinson, S.; Davies, W. Highlighting the threat from current and near-future ozone pollution to clover in pasture. Environ. Pollut. 2014, 189, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, D.; Mills, G.; Hayes, F.; Davies, W. The climate benefits of high-sugar grassland may be compromised by ozone pollution. Sci. Total Environ. 2016, 565, 95–104. [Google Scholar] [CrossRef]
- Otu-Larbi, F.; Conte, A.; Fares, S.; Wild, O.; Ashworth, K. FORCAsT-gs: Importance of stomatal conductance parameterisation to estimated ozone deposition velocity. J. Adv. Model. Earth Syst. 2021, 33, e2021MS002581. [Google Scholar] [CrossRef]
- Sicard, P.; De Marco, A.; Carrari, E.; Dalstein-Richier, L.; Hoshika, Y.; Badea, O.; Pitar, D.; Fares, S.; Conte, A.; Popa, I. Epidemiological derivation of flux-based critical levels for visible ozone injury in European forests. J. For. Res. 2020, 31, 1509–1519. [Google Scholar] [CrossRef]
- Hůnová, I. Ambient air quality in the Czech Republic: Past and present. Atmosphere 2020, 11, 214. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
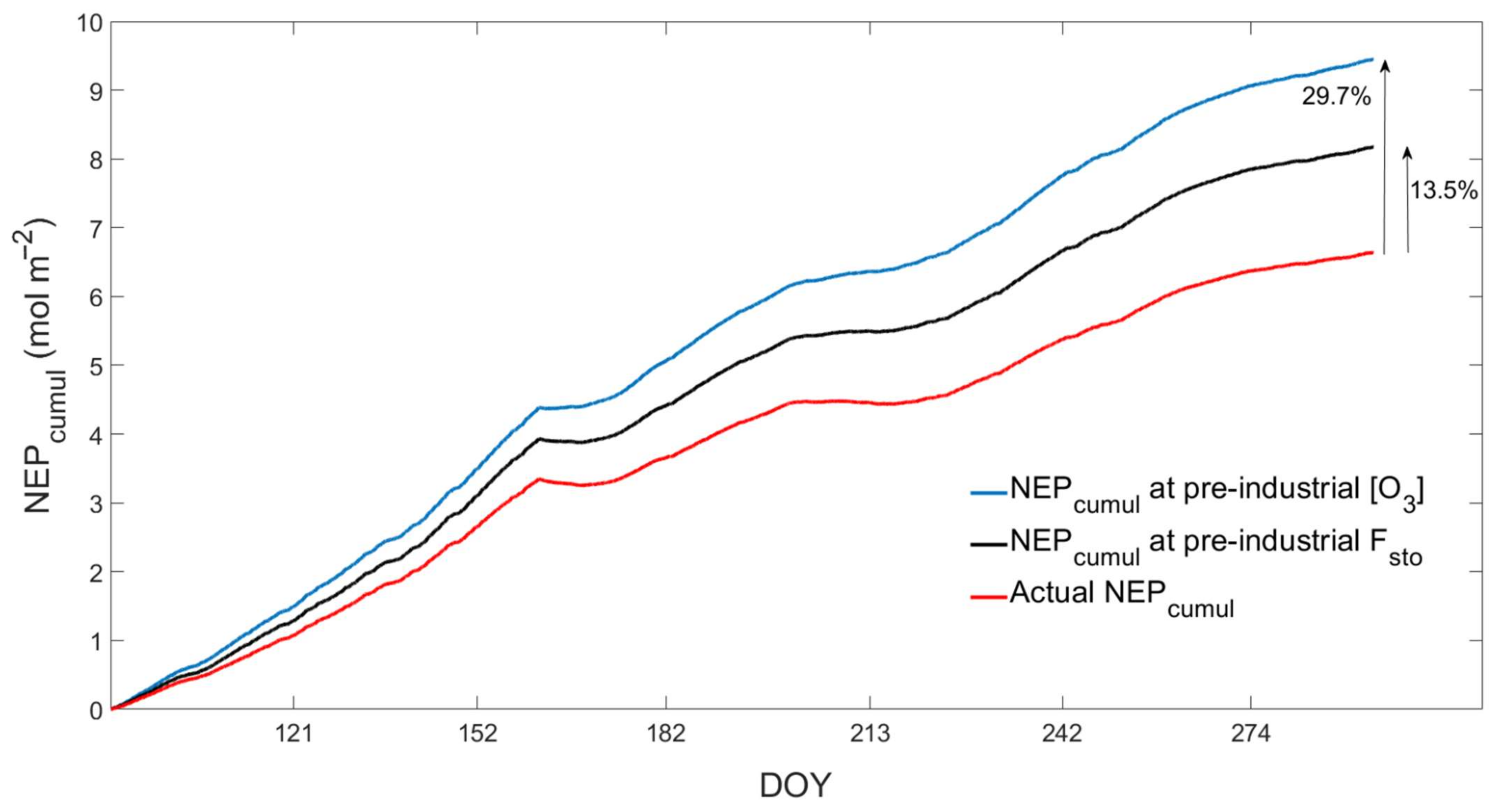{kind=link}
| Model | Predictor/ Interaction | Parametric Estimate | Goodness of Fit Std. Error | t-Value | Statistics p-Value |
|---|---|---|---|---|---|
| Model Fsto | Fsto | −1.767 | 0.069 | −25.45 | 2.00·10−16 ** |
| GPP | 1.055 | 0.005 | 223.95 | 2.00·10−16 ** | |
| GPP × Fsto | −0.022 | 0.004 | −6.18 | 7.18·10−16 ** | |
| Model [O3] | [O3] | −0.076 | 0.002 | −31.90 | 2.00·10−16 ** |
| GPP | 1.067 | 0.007 | 161.06 | 2.00·10−16 ** | |
| [O3] × GPP | −0.002 | 0.000 | −12.54 | 2.00·10−16 ** |
| Month | NEP (mol m−2) | ΔNEP (%) | ΔNEP (%) | |||
|---|---|---|---|---|---|---|
| EC | LM | Low [O3] | High [O3] | Low Fsto | High Fsto | |
| April | 10.729 | 10.708 | ↑ 28.1 | ↓ 13.9 | ↑ 16.6 | ↓ 12.1 |
| May | 15.769 | 15.764 | ↑ 21.1 | ↓ 13.3 | ↑ 13.1 | ↓ 8.1 |
| June | 10.017 | 10.050 | ↑ 36.2 | ↓ 7.9 | ↑ 23.5 | ↓ 6.9 |
| July | 7.967 | 7.982 | ↑ 38.8 | ↓ 18.2 | ↑ 26.2 | ↓ 13.8 |
| August | 9.256 | 9.276 | ↑ 33.5 | ↓ 16.2 | ↑ 21.0 | ↓ 13.0 |
| September | 9.962 | 9.968 | ↑ 23.7 | ↓ 28.7 | ↑ 15.9 | ↓ 7.0 |
| October | 2.671 | 2.670 | ↑ 30.2 | ↓ 86.6 | ↑ 17.1 | ↓ 61.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agyei, T.; Juráň, S.; Edwards-Jonášová, M.; Fischer, M.; Švik, M.; Komínková, K.; Ofori-Amanfo, K.K.; Marek, M.V.; Grace, J.; Urban, O. The Influence of Ozone on Net Ecosystem Production of a Ryegrass–Clover Mixture under Field Conditions. Atmosphere 2021, 12, 1629. https://doi.org/10.3390/atmos12121629
Agyei T, Juráň S, Edwards-Jonášová M, Fischer M, Švik M, Komínková K, Ofori-Amanfo KK, Marek MV, Grace J, Urban O. The Influence of Ozone on Net Ecosystem Production of a Ryegrass–Clover Mixture under Field Conditions. Atmosphere. 2021; 12(12):1629. https://doi.org/10.3390/atmos12121629
Chicago/Turabian StyleAgyei, Thomas, Stanislav Juráň, Magda Edwards-Jonášová, Milan Fischer, Marian Švik, Kateřina Komínková, Kojo Kwakye Ofori-Amanfo, Michal V. Marek, John Grace, and Otmar Urban. 2021. "The Influence of Ozone on Net Ecosystem Production of a Ryegrass–Clover Mixture under Field Conditions" Atmosphere 12, no. 12: 1629. https://doi.org/10.3390/atmos12121629