
Ablation of Gabra5 Influences Corticosterone Levels and Anxiety-like Behavior in Mice

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Husbandry
2.2. Model Generation
2.3. Western Blotting
2.4. Fecal Corticosterone Assessment and Housing
2.5. Testosterone Sampling and Elisa
2.6. Open Field
2.7. Elevated Plus Maze
2.8. Fear Conditioning
2.9. Slice Preparation
2.10. Electrophysiology
2.11. Indirect Calorimetry
2.12. In Vivo Body Composition Analysis
2.13. Statistical Analyses
3. Results
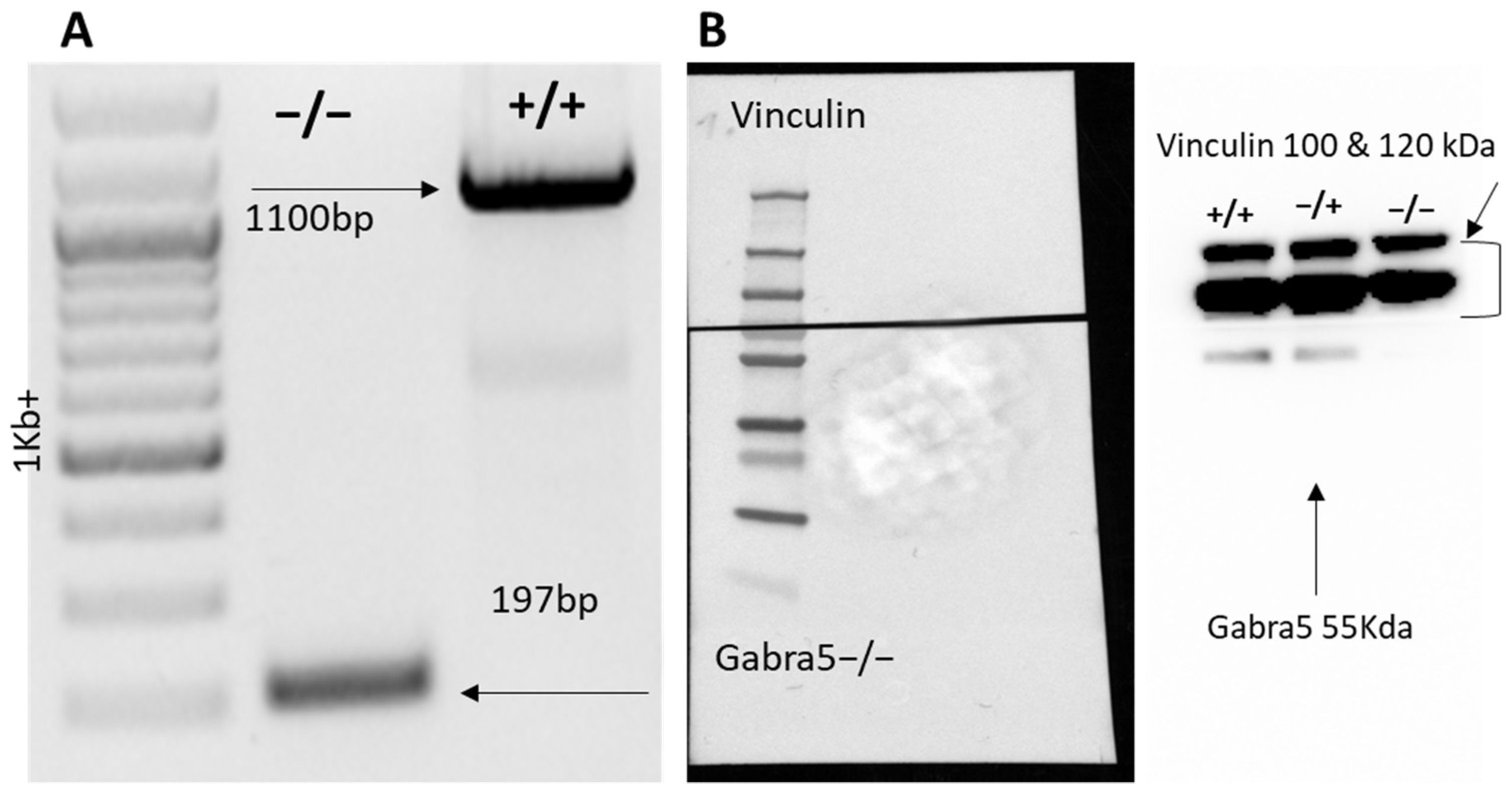
3.1. Generation of Gabra5−/− Mouse Model
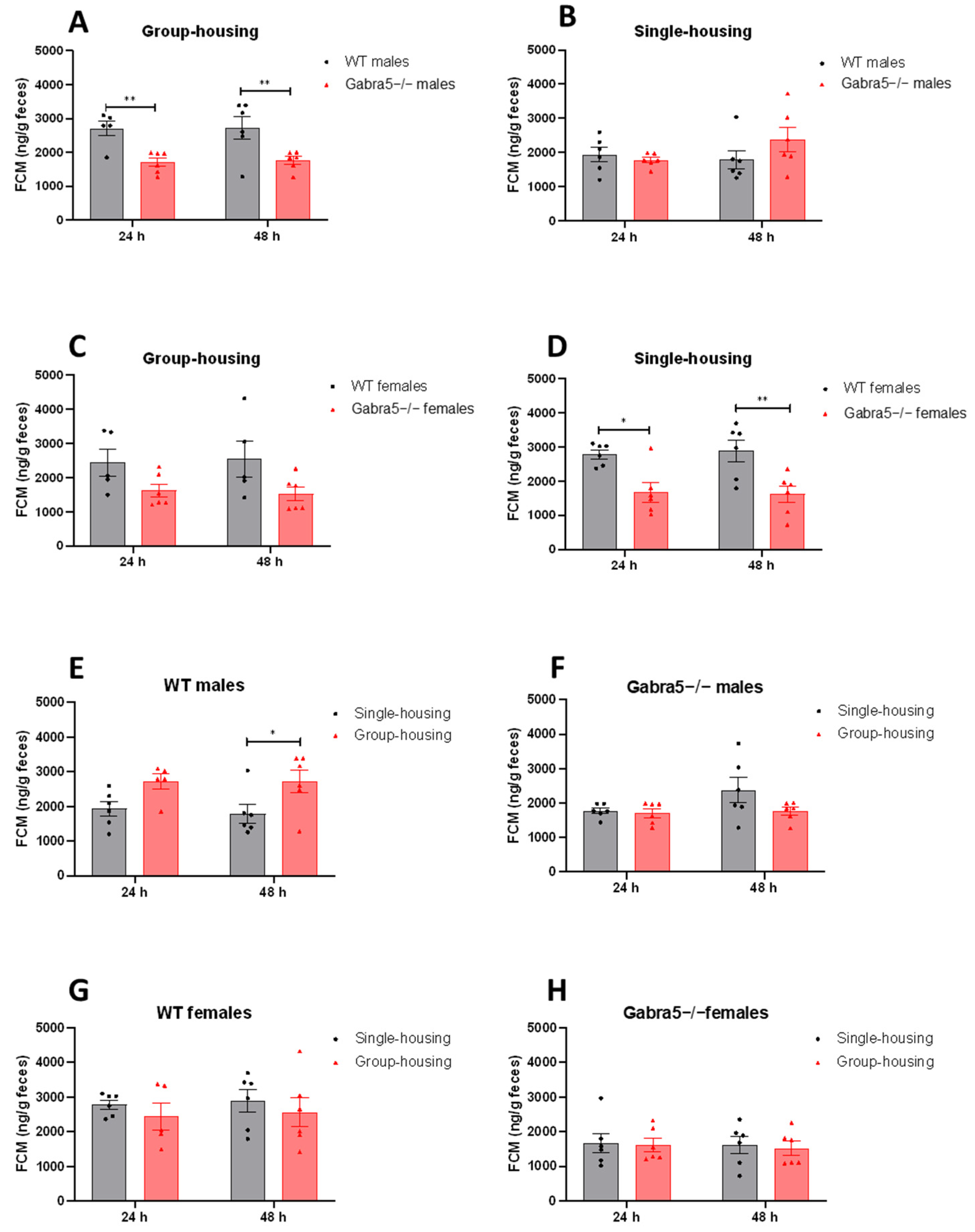
3.2. Gabra5−/− Mice Exhibit Decreased Levels of Fecal Corticosterone Metabolite
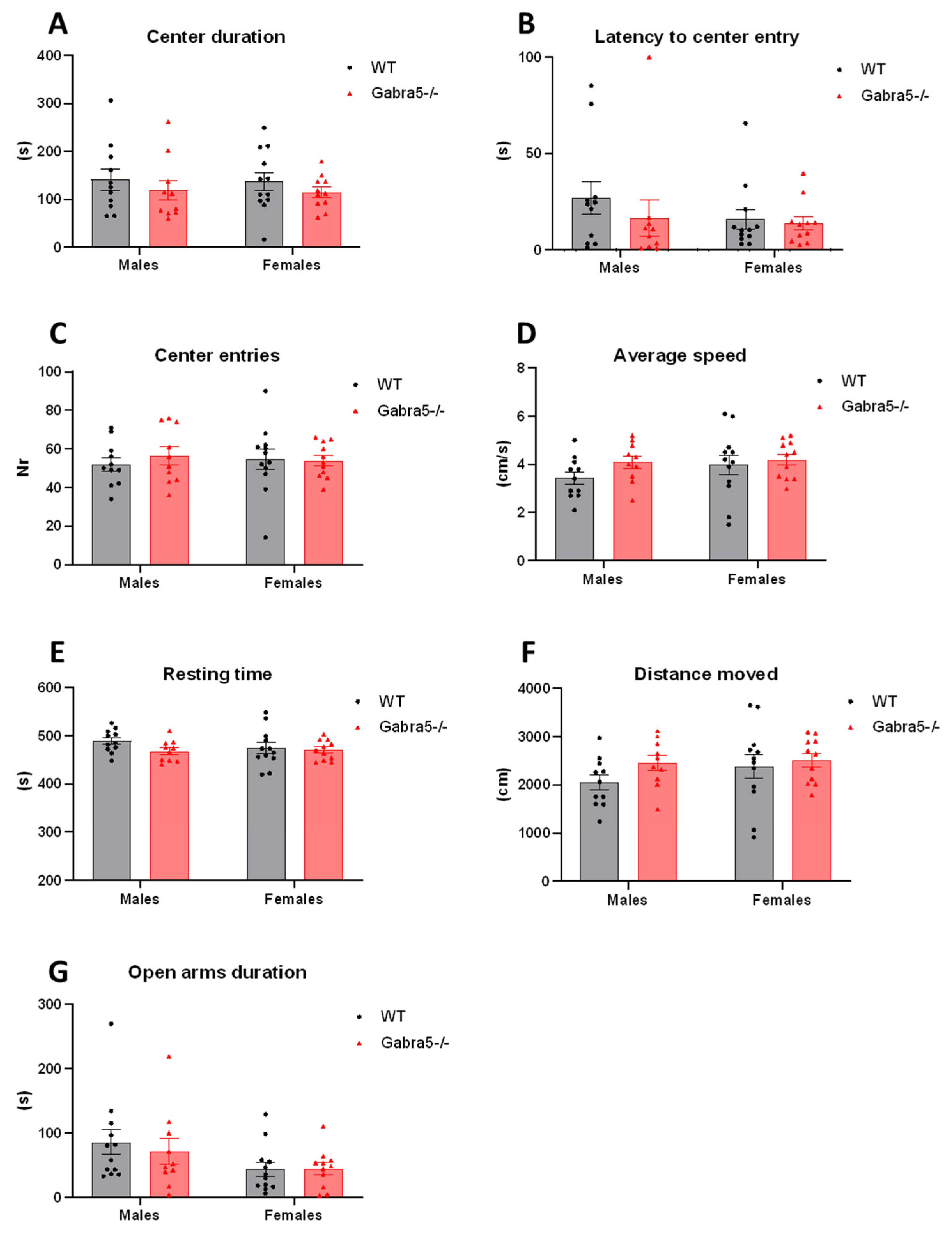
3.3. Open Field and Elevated Plus Maze Tests Do Not Suggest General Anxiety in Gabra5−/− Mice
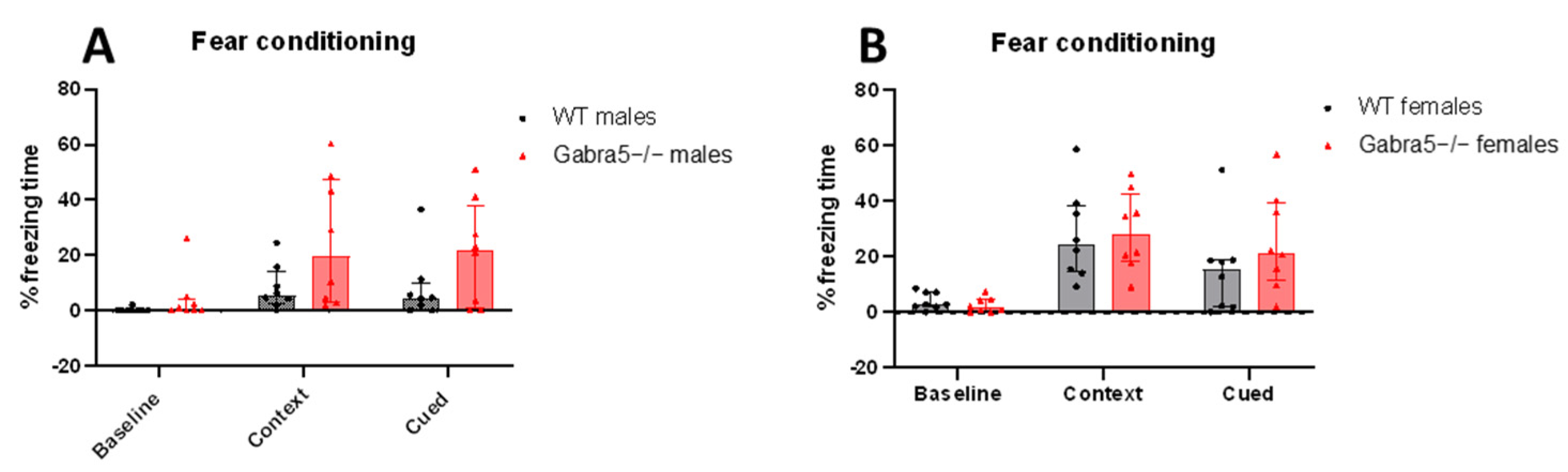
3.4. Gabra5−/− Animals Do Not Differ in Contextual and Delayed Cued Fear Conditioning
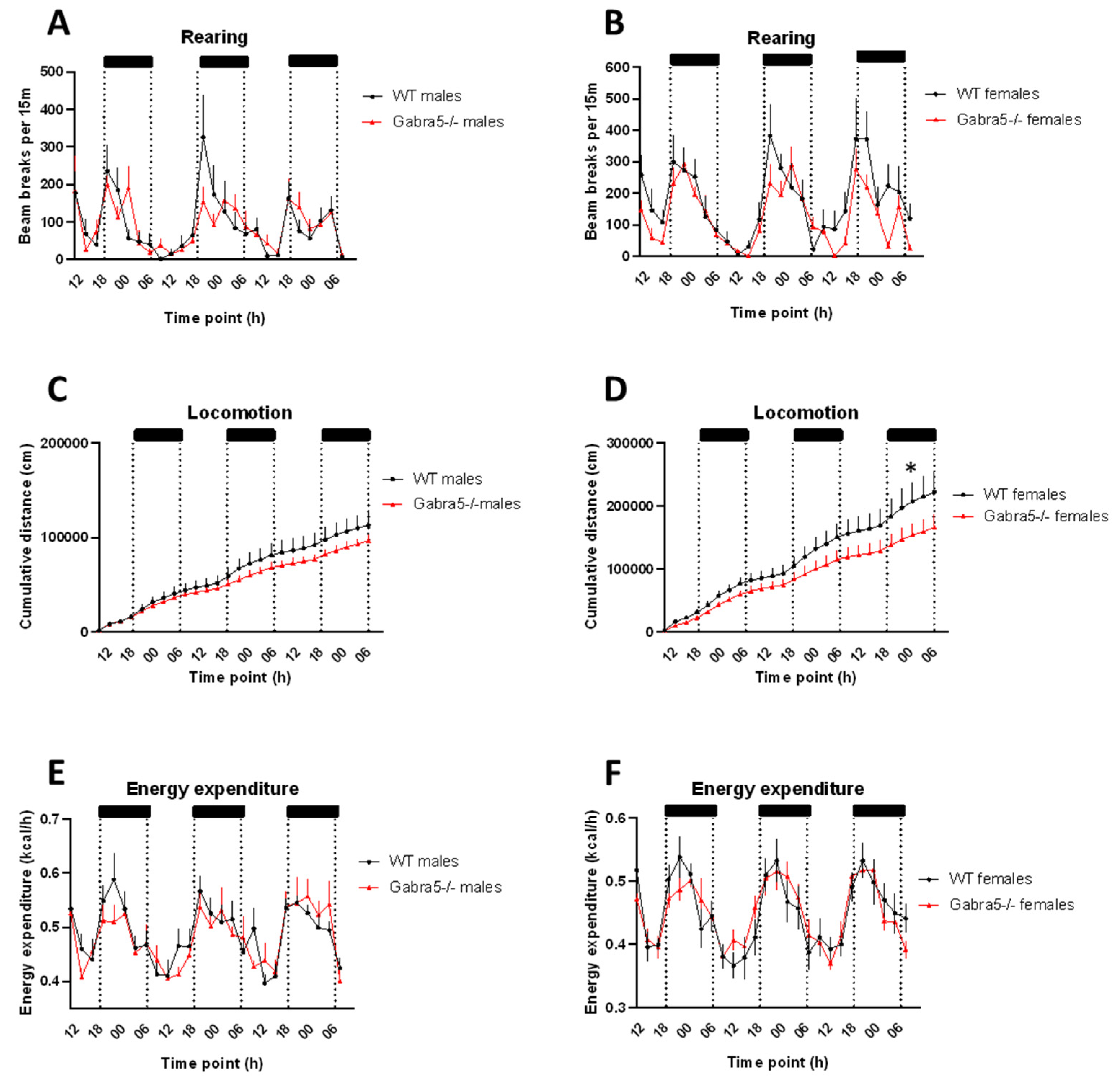
3.5. Gabra5−/− Animals Exhibit Decreased Rearing Behavior
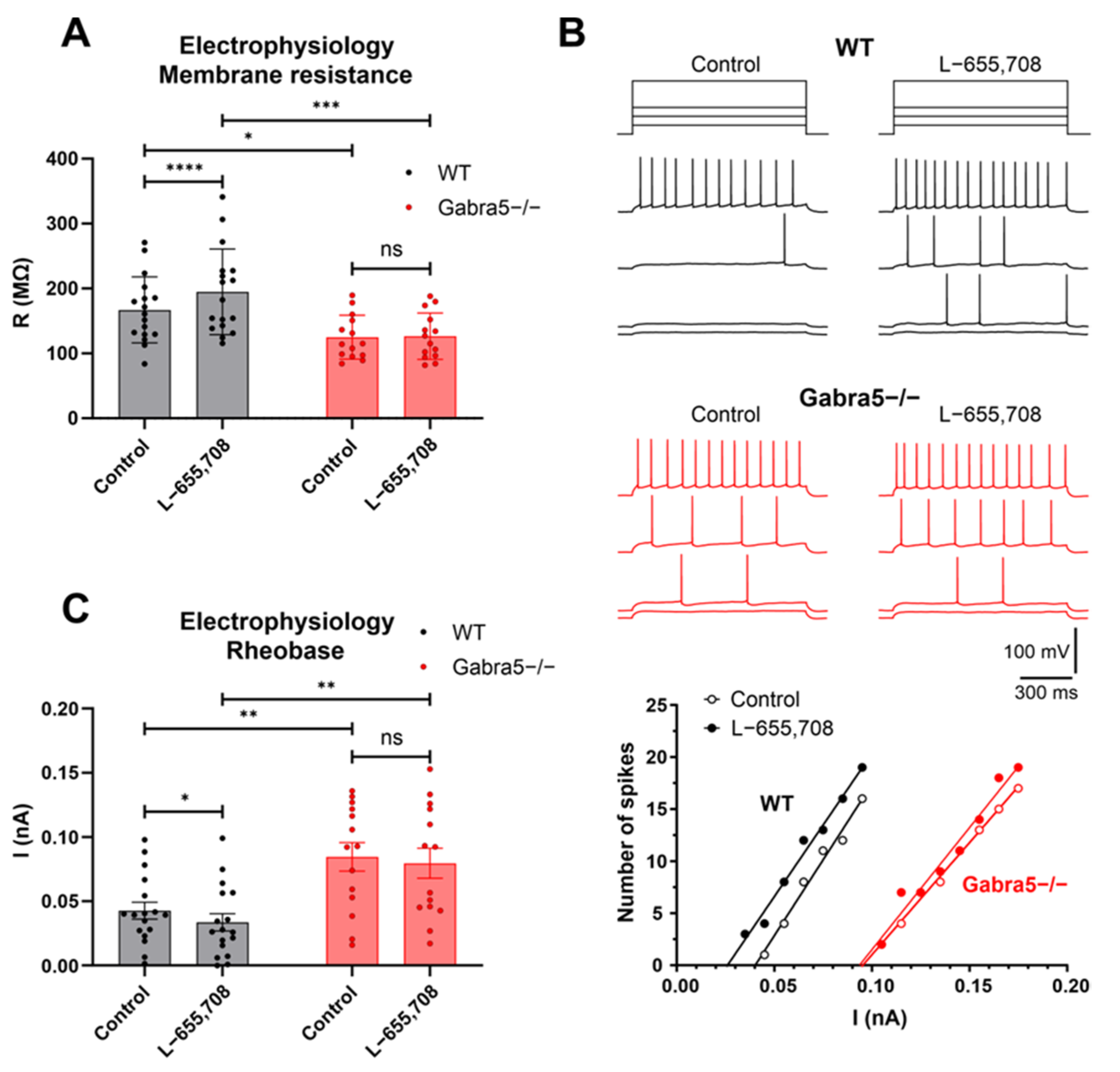
3.6. Hippocampal CA1 Pyramidal Neurons Exhibit Reduced Excitability in Gabra5−/− Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kagias, K.; Nehammer, C.; Pocock, R. Neuronal Responses to Physiological Stress. Front. Genet. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodnite, P.M. Stress: A concept analysis. Nurs. Forum 2014, 49, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.W. PVN pathways controlling energy homeostasis. Indian J. Endocrinol. Metab. 2012, 16 (Suppl. 3), S627–S636. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [CrossRef]
- Jubb, A.W.; Boyle, S.; Hume, D.A.; Bickmore, W.A. Glucocorticoid Receptor Binding Induces Rapid and Prolonged Large-Scale Chromatin Decompaction at Multiple Target Loci. Cell Rep. 2017, 21, 3022–3031. [Google Scholar] [CrossRef] [Green Version]
- Raglan, G.B.; Schmidt, L.A.; Schulkin, J. The role of glucocorticoids and corticotropin-releasing hormone regulation on anxiety symptoms and response to treatment. Endocr. Connect. 2017, 6, R1–R7. [Google Scholar] [CrossRef]
- Sapolsky, R.M. Why Zebras Don’t Get Ulcers: The Acclaimed Guide to Stress, Stress-Related Diseases, and Coping; Holt Paperbacks: New York, NY, USA, 2004. [Google Scholar]
- Cullinan, W.E.; Ziegler, D.R.; Herman, J.P. Functional role of local GABAergic influences on the HPA axis. Brain Struct. Funct. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Gunn, B.; Brown, A.R.; Lambert, J.J.; Belelli, D. Neurosteroids and GABAA Receptor Interactions: A Focus on Stress. Front. Neurosci. 2011, 5, 131. [Google Scholar] [CrossRef] [Green Version]
- Cullinan, W.E.; Helmreich, D.L.; Watson, S.J. Fos expression in forebrain afferents to the hypothalamic paraventricular nucleus following swim stress. J. Comp. Neurol. 1996, 368, 88–99. [Google Scholar] [CrossRef]
- Everington, E.A.; Gibbard, A.G.; Swinny, J.D.; Seifi, M. Molecular Characterization of GABA-A Receptor Subunit Diversity within Major Peripheral Organs and Their Plasticity in Response to Early Life Psychosocial Stress. Front. Mol. Neurosci. 2018, 11, 18. [Google Scholar] [CrossRef]
- Jacob, T.C. Neurobiology and Therapeutic Potential of α5-GABA Type A Receptors. Front. Mol. Neurosci. 2019, 12, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, J.; Wakefield, S.; MacKenzie, G.; Moss, S.J.; Maguire, J. Neurosteroidogenesis is required for the physiological response to stress: Role of neurosteroid-sensitive GABAA receptors. J. Neurosci. 2011, 31, 18198–18210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, L.M. Angelman syndrome: Review of clinical and molecular aspects. Appl. Clin. Genet. 2014, 7, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodges, L.M.; Fyer, A.J.; Weissman, M.M.; Logue, M.W.; Haghighi, F.; Evgrafov, O.; Rotondo, A.; Knowles, J.A.; Hamilton, S.P. Evidence for linkage and association of GABRB3 and GABRA5 to panic disorder. Neuropsychopharmacology 2014, 39, 2423–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnin, E.; Francavilla, R.; Amalyan, S.; Gervais, E.; David, L.S.; Luo, X.; Topolnik, L. Input-Specific Synaptic Location and Function of the α5 GABAA Receptor Subunit in the Mouse CA1 Hippocampal Neurons. J. Neurosci. 2019, 39, 788–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, J.F.; Burón, E.; Martın-López, M. Anxiogenic-like activity of L-655,708, a selective ligand for the benzodiazepine site of GABA(A) receptors which contain the α-5 subunit, in the elevated plus-maze test. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2002, 26, 1389–1392. [Google Scholar] [CrossRef] [PubMed]
- Behlke, L.M.; Foster, R.A.; Liu, J.; Benke, D.; Benham, R.S.; Nathanson, A.J.; Yee, B.K.; Zeilhofer, H.U.; Engin, E.; Rudolph, U. A Pharmacogenetic ‘Restriction-of-Function’ Approach Reveals Evidence for Anxiolytic-Like Actions Mediated by α5-Containing GABAA Receptors in Mice. Neuropsychopharmacology 2016, 41, 2492–2501. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.W.; Sieghart, W. GABA A receptors: Subtypes provide diversity of function and pharmacology. Neuropharmacology 2009, 56, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Myers, B.; McKlveen, J.M.; Herman, J.P. Glucocorticoid actions on synapses, circuits, and behavior: Implications for the energetics of stress. Front. Neuroendocrinol. 2014, 35, 180–196. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J.; Bonin, R.P.; Orser, B.A. The physiological properties and therapeutic potential of alpha5-GABAA receptors. Biochem. Soc. Trans. 2009, 37, 1334–1337. [Google Scholar] [CrossRef]
- Shin, L.M.; Liberzon, I. The neurocircuitry of fear, stress, and anxiety disorders. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2010, 35, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [PubMed] [Green Version]
- Cominski, T.P.; Jiao, X.; Catuzzi, J.E.; Stewart, A.L.; Pang, K.C.H. The role of the hippocampus in avoidance learning and anxiety vulnerability. Front. Behav. Neurosci. 2014, 8, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, J.C.; Su, K.; Goldberg, A.R.; Luna, V.M.; Biane, J.S.; Ordek, G.; Zhou, P.; Ong, S.K.; Wright, M.A.; Zweifel, L.; et al. Anxiety Cells in a Hippocampal-Hypothalamic Circuit. Neuron 2018, 97, 670–683.e6. [Google Scholar] [CrossRef] [Green Version]
- Caraiscos, V.B.; Elliott, E.M.; You-Ten, K.E.; Cheng, V.Y.; Belelli, D.; Newell, J.G.; Jackson, M.F.; Lambert, J.J.; Rosahl, T.W.; Wafford, K.A.; et al. Tonic inhibition in mouse hippocampal CA1 pyramidal neurons is mediated by alpha5 subunit-containing γ-aminobutyric acid type A receptors. Proc. Natl. Acad. Sci. USA 2004, 101, 3662–3667. [Google Scholar] [CrossRef] [Green Version]
- Collinson, N.; Kuenzi, F.M.; Jarolimek, W.; Maubach, K.A.; Cothliff, R.; Sur, C.; Smith, A.; Otu, F.M.; Howell, O.; Atack, J.R.; et al. Enhanced learning and memory and altered GABAergic synaptic transmission in mice lacking the α 5 subunit of the GABAA receptor. J. Neurosci. 2002, 22, 5572–5580. [Google Scholar] [CrossRef] [Green Version]
- Kulesskaya, N.; Voikar, V. Assessment of mouse anxiety-like behavior in the light-dark box and open-field arena: Role of equipment and procedure. Physiol. Behav. 2014, 133, 30–38. [Google Scholar] [CrossRef]
- Shoji, H.; Takao, K.; Hattori, S.; Miyakawa, T. Contextual and cued fear conditioning test using a video analyzing system in mice. J. Vis. Exp. 2014, 85, e50871. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J.; Zurek, A.A.; MacDonald, J.F.; Roder, J.C.; Jackson, M.F.; Orser, B.A. α5GABAAReceptor Activity Sets the Threshold for Long-Term Potentiation and Constrains Hippocampus-Dependent Memory. J. Neurosci. 2010, 30, 5269–5282. [Google Scholar] [CrossRef] [Green Version]
- Glykys, J.; Mody, I. Hippocampal network hyperactivity after selective reduction of tonic inhibition in GABA A receptor alpha5 subunit-deficient mice. J. Neurophysiol. 2006, 95, 2796–2807. [Google Scholar] [CrossRef]
- Morla, L.; Shore, O.; Lynch, I.J.; Merritt, M.E.; Wingo, C.S. A noninvasive method to study the evolution of extracellular fluid volume in mice using time-domain nuclear magnetic resonance. Am. J. Physiol. Ren. Physiol. 2020, 319, F115–F124. [Google Scholar] [CrossRef]
- Mitra, R.; Sapolsky, R.M. Acute corticosterone treatment is sufficient to induce anxiety and amygdaloid dendritic hypertrophy. Proc. Natl. Acad. Sci. USA 2008, 105, 5573–5578. [Google Scholar] [CrossRef] [Green Version]
- Sturman, O.; Germain, P.-L.; Bohacek, J. Exploratory rearing: A context- and stress-sensitive behavior recorded in the open-field test. Stress 2018, 21, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Izzi-Engbeaya, C.; Ma, Y.; Buckley, N.W.; Ratnasabapathy, R.; Richardson, E.; Counsell, J.R.; Fernandes-Freitas, I.; Norton, M.; Farooq, G.; Mirza, Z.; et al. Effects of corticosterone within the hypothalamic arcuate nucleus on food intake and body weight in male rats. Mol. Metab. 2020, 36, 100972. [Google Scholar] [CrossRef]
- Serwanski, D.R.; Miralles, C.P.; Christie, S.B.; Mehta, A.K.; Li, X.; De Blas, A.L. Synaptic and nonsynaptic localization of GABAA receptors containing the alpha5 subunit in the rat brain. J. Comp. Neurol. 2006, 499, 458–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonin, R.P.; Martin, L.J.; MacDonald, J.F.; Orser, B.A. Alpha5GABAA receptors regulate the intrinsic excitability of mouse hippocampal pyramidal neurons. J. Neurophysiol. 2007, 98, 2244–2254. [Google Scholar] [CrossRef] [PubMed]
- Syding, L.A.; Nickl, P.; Kasparek, P.; Sedlacek, R. CRISPR/Cas9 Epigenome Editing Potential for Rare Imprinting Diseases: A Review. Cells 2020, 9, 993. [Google Scholar] [CrossRef]
- Gasser, P.J.; Lowry, C.A.; Orchinik, M. 41—Rapid corticosteroid actions on behavior: Mechanisms and implications. In Hormones, Brain and Behavior, 2nd ed.; Pfaff, D.W., Joels, M., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 1365–1397. [Google Scholar]
- Lidster, K.; Owen, K.; Browne, W.J.; Prescott, M.J. Cage aggression in group-housed laboratory male mice: An international data crowdsourcing project. Sci. Rep. 2019, 9, 15211. [Google Scholar] [CrossRef] [Green Version]
- Giammanco, M.; Tabacchi, G.; Giammanco, S.; Di Majo, D.; La Guardia, M. Testosterone and aggressiveness. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2005, 11, RA136–RA145. [Google Scholar]
- Piantadosi, S.C.; French, B.J.; Poe, M.M.; Timić, T.; Marković, B.D.; Pabba, M.; Seney, M.L.; Oh, H.; Orser, B.A.; Savić, M.M.; et al. Sex-Dependent Anti-Stress Effect of an α5 Subunit Containing GABAA Receptor Positive Allosteric Modulator. Front. Pharmacol. 2016, 7, 446. [Google Scholar] [CrossRef] [Green Version]
- Lever, C.; Burton, S.; O’Keefe, J. Rearing on hind legs, environmental novelty, and the hippocampal formation. Rev. Neurosci. 2006, 17, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, D.C.; Blanchard, R.J.; Rodgers, R.J. Risk Assessment and animal models of anxiety. In Animal Models in Psychopharmacology; Olivier, B., Mos, J., Slangen, J.L., Eds.; Birkhäuser Basel: Basel, Switzerland, 1991; pp. 117–134. [Google Scholar]
- Cryan, J.F.; Sweeney, F.F. The age of anxiety: Role of animal models of anxiolytic action in drug discovery. Br. J. Pharmacol. 2011, 164, 1129–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balon, R.; Starcevic, V. Role of Benzodiazepines in Anxiety Disorders. Anxiety Disord. 2020, 1191, 367–388. [Google Scholar]
- El-Brolosy, M.A.; Stainier, D.Y.R. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genet. 2017, 13, e1006780. [Google Scholar] [CrossRef] [Green Version]
- Crestani, F.; Keist, R.; Fritschy, J.-M.; Benke, D.; Vogt, K.; Prut, L.; Bluthmann, H.; Mohler, H.; Rudolph, H. Trace fear conditioning involves hippocampal alpha5 GABA(A) receptors. Proc. Natl. Acad. Sci. USA 2002, 99, 8980–8985. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J.; Oh, G.H.T.; Orser, B.A. Etomidate targets alpha5 γ-aminobutyric acid subtype A receptors to regulate synaptic plasticity and memory blockade. Anesthesiology 2009, 111, 1025–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonin, R.P.; Zurek, A.A.; Yu, J.; Bayliss, D.A.; Orser, B.A. Hyperpolarization-Activated Current (Ih) Is Reduced in Hippocampal Neurons from Gabra5−/− Mice. PLoS ONE 2013, 8, e58679. [Google Scholar] [CrossRef]
- Glykys, J.; Mann, E.O.; Mody, I. Which GABA(A) receptor subunits are necessary for tonic inhibition in the hippocampus? J. Neurosci. 2008, 28, 1421–1426. [Google Scholar] [CrossRef] [Green Version]
- Brickley, S.G.; Revilla, V.; Cull-Candy, S.G.; Wisden, W.; Farrant, M. Adaptive regulation of neuronal excitability by a voltage-independent potassium conductance. Nature 2001, 409, 88–92. [Google Scholar] [CrossRef]
- Jin, C.; Kang, H.; Yoo, T.; Ryu, J.R.; Yoo, Y.-E.; Ma, R.; Zhang, Y.; Kang, H.R.; Kim, Y.; Seong, H.; et al. The Neomycin Resistance Cassette in the Targeted Allele of Shank3B Knock-Out Mice Has Potential Off-Target Effects to Produce an Unusual Shank3 Isoform. Front. Mol. Neurosci. 2021, 13, 614435. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Syding, L.A.; Kubik-Zahorodna, A.; Reguera, D.P.; Nickl, P.; Hruskova, B.; Kralikova, M.; Kopkanova, J.; Novosadova, V.; Kasparek, P.; Prochazka, J.; et al. Ablation of Gabra5 Influences Corticosterone Levels and Anxiety-like Behavior in Mice. Genes 2023, 14, 285. https://doi.org/10.3390/genes14020285
Syding LA, Kubik-Zahorodna A, Reguera DP, Nickl P, Hruskova B, Kralikova M, Kopkanova J, Novosadova V, Kasparek P, Prochazka J, et al. Ablation of Gabra5 Influences Corticosterone Levels and Anxiety-like Behavior in Mice. Genes. 2023; 14(2):285. https://doi.org/10.3390/genes14020285
Chicago/Turabian StyleSyding, Linn Amanda, Agnieszka Kubik-Zahorodna, David Pajuelo Reguera, Petr Nickl, Bohdana Hruskova, Michaela Kralikova, Jana Kopkanova, Vendula Novosadova, Petr Kasparek, Jan Prochazka, and et al. 2023. "Ablation of Gabra5 Influences Corticosterone Levels and Anxiety-like Behavior in Mice" Genes 14, no. 2: 285. https://doi.org/10.3390/genes14020285