Taxonomic Diversity Not Associated with Gross Karyotype Differentiation: The Case of Bighead Carps, Genus Hypophthalmichthys (Teleostei, Cypriniformes, Xenocyprididae)

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling
2.2. Chromosome Preparation and Conventional Cytogenetics
2.3. DNA Isolation and Preparation of FISH Probes
2.4. FISH Analysis
2.5. Microscopic Analyses and Image Processing
3. Results
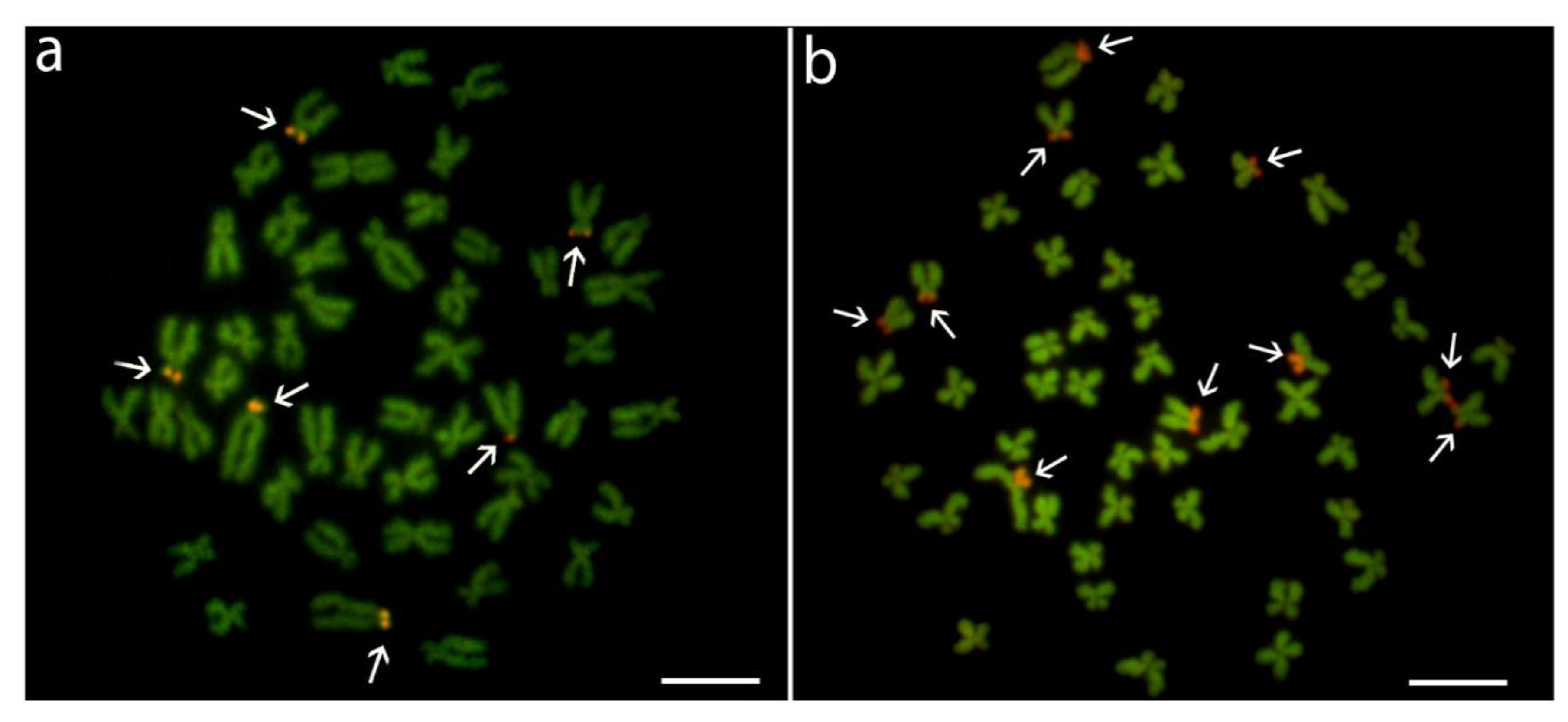
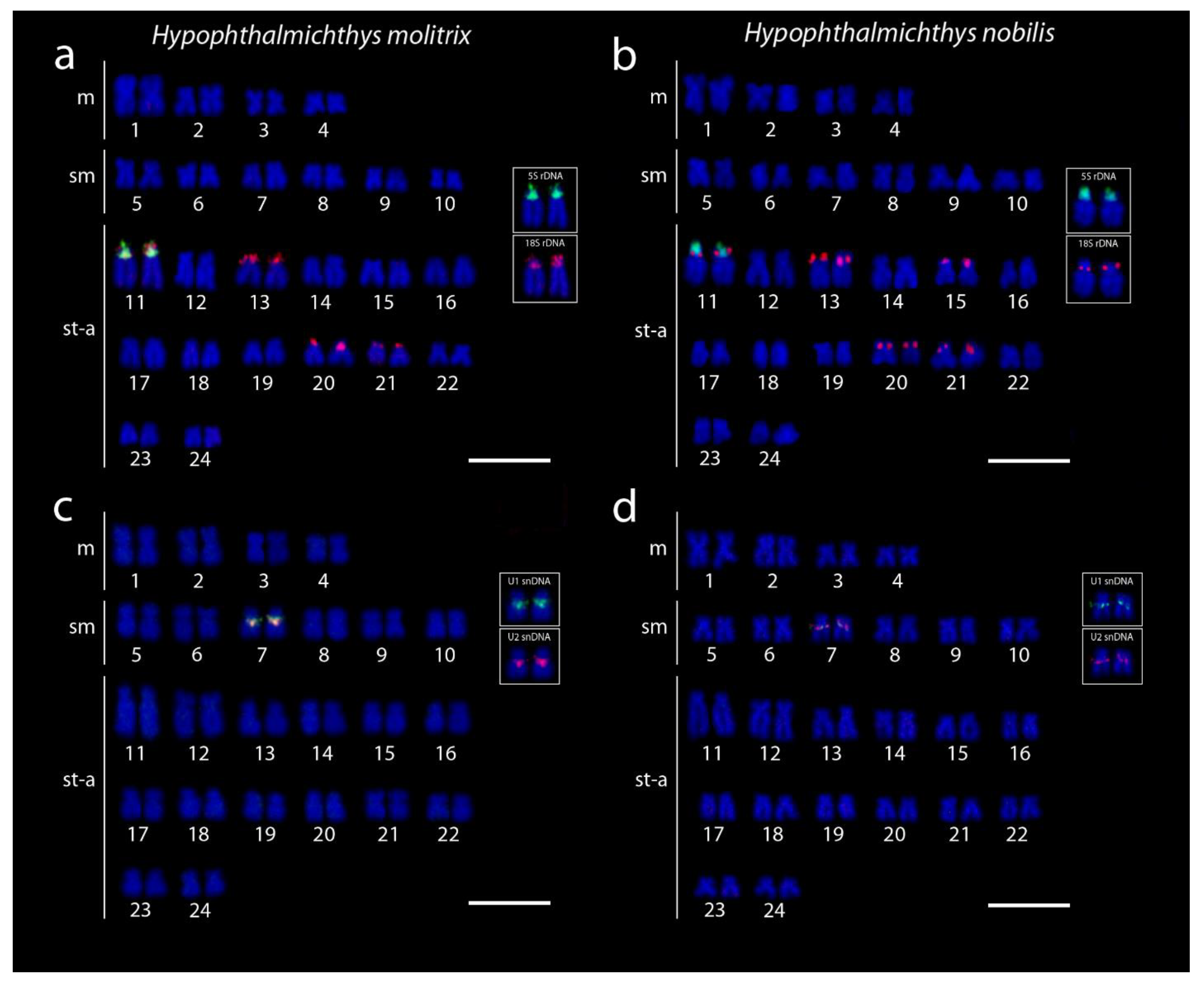
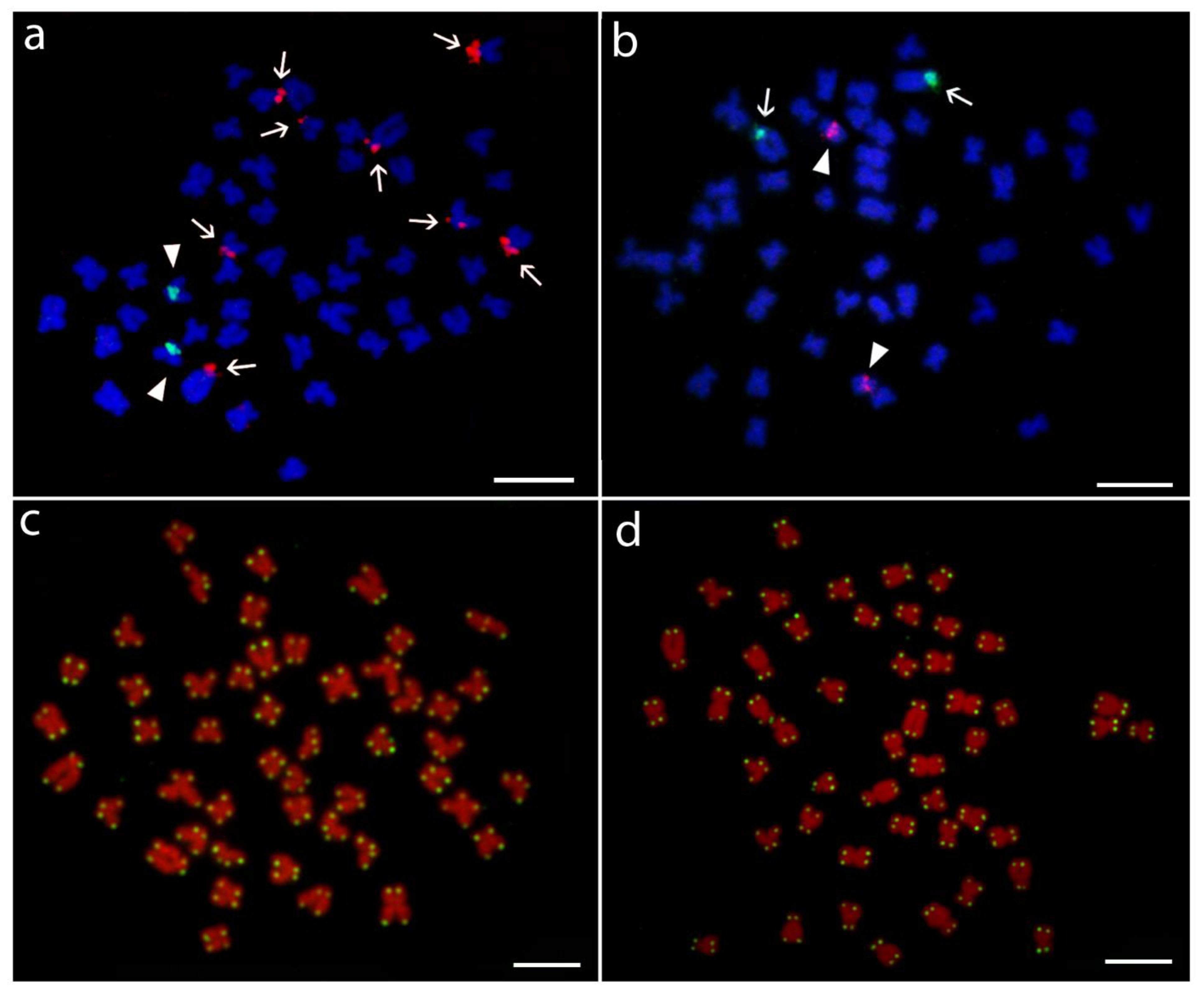
3.1. Karyotypes and Chromosome Banding Characteristics
3.2. Sequence Analysis of Repetitive DNA Fragments
3.3. Hybridization Patterns of Repetitive DNA Probes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Howes, G.J. Anatomy and phylogeny of the Chinese major carps Ctenopharyngodon Steind., 1866 and Hypophthalmichthys Blkr., 1860. Bull. Br. Mus. (Nat. Hist.) Zool 1981, 41, 1–52. [Google Scholar]
- Tan, M.; Armbruster, J.W. Phylogenetic classification of extant genera of fishes of the order Cypriniformes (Teleostei. Ostariophysi). Zootaxa 2018, 4476, 6–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, W.; Zou, M.; Wang, X.; Gan, X.; Mayden, R.L.; He, S. Phylogenomic analysis resolves the formerly intractable adaptive diversification of the endemic clade of East Asian Cyprinidae (Cypriniformes). PLoS ONE 2010, 5, e13508. [Google Scholar] [CrossRef] [PubMed]
- Kottelat, M. The fishes of the inland waters of Southeast Asia. In A Catalogue and Core Bibliography of the Fishes Known to Occur in Freshwaters, Mangroves and Estuaries; Raffles Bull. Zool: Kent Ridge, Singapore, 2013; pp. 1–663. [Google Scholar]
- Yen, M.D. Species composition and distribution of the freshwater fish fauna of the North of Vietnam. Hydrobiologia 1985, 121, 281–286. [Google Scholar] [CrossRef]
- FAO Statistics Food and Agriculture. Available online: http://www.fao.org/fishery/culturedspecies/Hypophthalmichthys_molitrix/en (accessed on 29 January 2020).
- Stepien, C.A.; Snyder, M.R.; Elz, A.E. Invasion genetics of the silver carp Hypophthalmichthys molitrix across North America: Differentiation of fronts, introgression, and eDNA metabarcode detection. PLoS ONE 2019, 14, e0203012. [Google Scholar] [CrossRef] [Green Version]
- Sing, A.K.; Kumar, D.; Srivastava, S.C.; Ansan, A.; Jena, T.K.; Sarkar, U.K. Invasion and impacts of alien fish species in the Ganga River, India. Aquat. Ecosyst. Health Manag. 2013, 16, 408–414. [Google Scholar] [CrossRef]
- Luebcker, N.; Zengeva, T.A.; Dabrowski, J.; Robertson, J.P. Predicting the potential distribution of invasive silver carp Hypophthalmichthys molitrix in South Africa. Afr. J. Aquat. Sci. 2014, 39, 157–165. [Google Scholar] [CrossRef]
- Garvey, J.E. Bigheads of the genus Hypophthalmichthys. In A Handbook of Global Freshwater Invasive Species; Francis, R.A., Ed.; Earthscan: New York, NY, USA, 2012; pp. 235–245. [Google Scholar]
- Hayer, C.A.; Breeggemann, J.J.; Jason, J.; Klumb, R.A.; Graeb, B.D.S.; Bertrand, K.N. Population characteristics of bighead and silver carp on the northwestern front of their North American invasion. Aquat. Invasions 2014, 9, 289–303. [Google Scholar] [CrossRef]
- Farrington, H.L.; Edwards, C.E.; Bartron, M.; Lance, R.F. Phylogeography and population genetics of introduced silver carp (Hypophthalmichthys molitrix) and bighead carp (H. nobilis) in North America. Biol. Invasions 2017, 19, 2789–2811. [Google Scholar] [CrossRef]
- DeBoer, J.A.; Anderson, A.M.; Casper, A.F. Multi-trophic response to invasive silver carp (Hypophthalmichthys molitrix) in large floodplain river. Freshw. Biol. 2018, 63, 597–611. [Google Scholar] [CrossRef]
- Tristano, E.P.; Coulter, A.A.; Newton, T.J.; Garvey, J.E. Invasive silver carp may compete with unionid mussels for algae: First experimental evidence. Aquat. Conserv. 2019, 29, 1749–1757. [Google Scholar] [CrossRef]
- Marián, T.; Krasznai, Z. Karyological investigation on Ctenopharyngodon idella and Hypophthalmichthys nobilis and their cross-breeding. Aquac. Hung. (Szarvas) 1978, 1, 44–50. [Google Scholar]
- Marián, T.; Krasznai, Z. Comparative karyological studies on Chinese carps. Aquaculture 1979, 13, 325–336. [Google Scholar] [CrossRef]
- Marián, T.; Krasznai, Z. Zylogische Untersuchungen bei der Familie Cyprinidae (Pisces). Biol. Zent. 1987, 97, 205–214. [Google Scholar]
- Marián, T.; Krasznai, Z.; Oláh, J. Characteristic karyological, biochemical and morphological markers of silver carp (Hypophthalmichthys molitrix), bighead carp (Aristicthys nobilis) and their hybrids. Aquac. Hung. 1986, 5, 15–30. [Google Scholar]
- Vasiliev, V.P.; Makeeva, A.P.; Ryabov, I.N. The study of chromosome complexes in cyprinid fish and their hybrids. Genetika 1978, 14, 1450–1460. (In Russian) [Google Scholar]
- Beck, M.L.; Biggers, C.J.; Dupree, H.K. Karyological analysis of Ctenopharyngodon idella, Aristichthys nobilis and their F1 hybrid. Trans. Am. Fish. Soc. 1980, 109, 433–438. [Google Scholar] [CrossRef]
- Beck, M.L.; Biggers, C.J. Chromosomal investigation of Ctenopharyngodon idella × Aristichthys nobilis hybrids. Experientia 1982, 38, 319. [Google Scholar] [CrossRef]
- Zan, R.G.; Song, Z. Analysis and comparison between karyotypes of Cyprinus carpio and Carassius auratus and Aristichthys nobilis and Hypophthalmichthys molitrix. Acta Genet. Sin. 1980, 7, 72–77. [Google Scholar]
- Zan, R.G.; Song, Z.; Liu, W.G. Studies on karyotypes and nuclear DNA contents of some cyprinoid fishes, with notes on fish polyploids in China. In Indo-Pacific Fish Biology: Proceedings of the Second International Conference on Indo-Pacific Fish; Uyeno, T., Arai, R., Taniuchi, T., Matsuura, K., Eds.; Ichthyological Society of Japan: Tokyo, Japan, 1986; pp. 877–885. [Google Scholar]
- Zhou, T. Analysis of karyotype in bighead carp. Freshw. Fish. (Shashi) 1980, 4, 3–7. (In Chinese) [Google Scholar]
- Zhou, T.; Lin, J.; Yang, Y. The karyotype of black carp [Mylopharyngodon piceus (Rich.)]. J. Wuhan Univ. 1980, 4, 112–116, (In Chinese with English Abstract). [Google Scholar]
- Liu, L.Y. Karyotype analysis of Aristichthys nobilis by the leucocyte culture method. J. Beijing Norm. Univ. (Nat. Sci.) 1981, 3, 79–83, (In Chinese with English Abstract). [Google Scholar]
- Almeida-Toledo, L.F.; Bigoni, A.P.V.; Bernardino, G.; De Almeida Filho, S. Chromosomal location of NORs and C bands in F1 hybrids of bighead carp and silver carp reared in Brazil. Aquaculture 1995, 135, 277–284. [Google Scholar] [CrossRef]
- Anonymous. A preliminary report on the chromosomes number of 52 species of fishes of the Pearl River system in Guandong Province. J. Wuhan Univ. 1983, 3, 123–125. [Google Scholar]
- Manna, G.K.; Khuda-Bukhsh, A.R. Karyomorphology of cyprinid fishes and cytological evaluation of the family. Nucleus 1978, 43, 119–127. [Google Scholar]
- Liu, L.Y. Karyotype analysis of Hypophthalmichthys molitrix. Acta Genet. Sin. 1981, 8, 251–255, (In Chinese with English Abstract). [Google Scholar]
- Su, Z.; Xu, K.; Chen, S.; Bai, G. Studies on triploid silver carp and its karyotype. Chin. Zool. Res. (Suppl.) Kunming 1984, 5, 15–19, (In Chinese with English Abstract). [Google Scholar]
- Yu, X.L.; Zhou, T.; Li, Z.C.; Li, K.; Zhou, M. On the karyosystematics of cyprinid fishes and a summary of fish chromosome studies in China. Genetica 1987, 72, 225–236. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Y.C.; Zhou, T.; Yu, X.J. A BrdU-BSG method for G-banding in fish chromosomes and an idiogram of G-banded karyotype of silver carp. Acta Genet. Sin. 1989, 16, 184–187. [Google Scholar]
- Ren, X.H.; Cui, J.; Yu, Q. Chromosomal heterochromatin differentiation of cyprinid fishes. Genetica 1992, 87, 47–51. [Google Scholar] [CrossRef]
- Khuda-Bukhsh, A.R.; Chakrabarti, C. Induction of serial bands akin to G-type on metaphase chromosomes of two species of fish, Rita rita (Bagridae) and Hypophthalmichthys molitrix (Cyprinidae). Indian J. Exp. Biol. 1996, 34, 1271–1273. [Google Scholar]
- Šlechtová, V.; Šlechta, V.; Hiep, D.D.; Valenta, M. Biochemical genetic comparison of bighead (Aristichthys nobilis) and silver carp (Hypophthalmichthys molitrix) and their hybrids in Czechoslovakia. J. Fish Biol. 1991, 39, 349–357. [Google Scholar] [CrossRef]
- Fujiwara, A.; Nishida-Umehara, C.; Sakamoto, T.; Okamoto, N.; Nakayama, I.; Abe, S. Improved fish lymphocyte culture for chromosome preparation. Genetica 2001, 111, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Flajšhans, M.; Sember, A.; Havelka, M.; Gela, D.; Kořínková, T.; Rodina, M.; Rábová, M.; Ráb, P. Molecular cytogenetics in artificial hybrid and highly polyploid sturgeons: An evolutionary story narrated by repetitive sequences. Cytogenet. Genome Res. 2013, 141, 153–162. [Google Scholar] [CrossRef]
- Ráb, P.; Roth, P. Cold-blooded vertebrates. In Methods of Chromosome Analysis; Balicek, P., Forejt, J., Rubeš, J., Eds.; Cytogenetická Sekce Československé Biologické Společnosti pri. CSAV: Brno, Czech Republic, 1988; pp. 115–124. [Google Scholar]
- Bertollo, L.A.C.; Cioffi, M.B.; Moreira-Filho, O. Direct chromosome preparation from freshwater teleost fishes. In Fish Cytogenetic Techniques (Ray-Fin Fishes and Chondrichthyans), 2nd ed.; Ozouf-Costaz, C., Pisano, E., Foresti, F., Toledo, L.F.A., Eds.; CRC Press: Enfield, CT, USA, 2015; Volume 1, pp. 21–26. [Google Scholar]
- Haaf, T.; Schmid, M. An early stage of ZZ/ZW sex chromosomes differentiation in Poecilia sphenops var. melanistica (Poeciliidae, Cyprinodontiformes). Chromosoma 1984, 89, 37–41. [Google Scholar] [CrossRef]
- Mayr, B.; Ráb, P.; Kalat, M. Localisation of NORs and counterstain-enhanced fluorescence studies in Perca fluviatilis (Pisces, Percidae). Genetica 1985, 67, 51–56. [Google Scholar] [CrossRef]
- Sola, L.; Rossi, A.R.; Iaselli, V.; Rasch, E.M.; Monaco, P.J. Cytogenetics of bisexual/unisexual species of Poecilia. II. Analysis of heterochromatin and nucleolar organizer regions in Poecilia mexicana mexicana by C-banding and DAPI, quinacrine, chromomycin A3, and silver staining. Cytogenet. Cell Genet 1992, 60, 229–235. [Google Scholar] [CrossRef]
- Howell, W.M.; Black, D.A. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: A 1-step method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef]
- Sember, A.; Bohlen, J.; Šlechtová, V.; Altmanová, M.; Symonová, R.; Ráb, P. Karyotype differentiation in 19 species of river loach fishes (Nemacheilidae, Teleostei): Extensive variability associated with rDNA and heterochromatin distribution and its phylogenetic and ecological interpretation. BMC Evol. Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [Green Version]
- Cioffi, M.B.; Martins, C.; Centofante, L.; Jacobina, U.; Bertollo, L.A.C. Chromosomal variability among allopatric populations of Erythrinidae fish Hoplias malabaricus: Mapping of three classes of repetitive DNAs. Cytogenet. Genome Res. 2009, 125, 132–141. [Google Scholar] [CrossRef]
- Silva, D.M.Z.A.; Utsunomia, R.; Pansonato-Alves, J.C.; Oliveira, C.; Foresti, F. Chromosomal mapping of repetitive DNA sequences in five species of Astyanax (Characiformes, Characidae) reveals independent location of U1 and U2 snRNA sites and association of U1 snRNA and 5S rDNA. Cytogenet. Genome Res. 2015, 146, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, C.F.; Bertollo, L.A.C.; Cioffi, M.B. Fish-FISH: Molecular cytogenetics in fish species. In Fluorescence In Situ Hybridization (FISH)—Application Guide, 2nd ed.; Liehr, T., Ed.; Springer: Berlin, Germany, 2017; pp. 429–443. [Google Scholar]
- Sember, A.; Bohlen, J.; Šlechtová, V.; Altmanová, M.; Pelikánová, Š.; Ráb, P. Dynamics of tandemly repeated DNA sequences during evolution of diploid and tetraploid botiid loaches (Teleostei: Cobitoidea: Botiidae). PLoS ONE 2018, 13, e0195054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. Available online: http://blast.ncbi.nlm.nih.gov/blast (accessed on 20 March 2018). [CrossRef]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Matthey, R. L’ evolution de la formule chromosomiale chez les vertebrees. Experientia 1945, 1, 78–86. [Google Scholar] [CrossRef]
- Liu, H.; Pang, M.; Yu, X.; Zhou, Y.; Tong, J.; Fu, B. Sex-specific markers developed by next-generation sequencing confirmed an XX/XY sex determination system in bighead carp (Hypophthalmichehys nobilis) and silver carp (Hypophthalmichthys molitrix). DNA Res. 2018, 25, 257–264. [Google Scholar] [CrossRef]
- Ráb, P.; Collares-Pereira, M.J. Chromosomes of European cyprinid fishes (Cyprinidae, Cypriniformes). A review. Folia Zool. 1995, 44, 193–214. [Google Scholar]
- Knytl, M.; Kalous, L.; Rylková, K.; Choleva, L.; Merilä, J.; Ráb, P. Morphologically indistinguishable hybrid Carassius female with 156 chromosomes: A threat for the threatened crucian carp, C. carassius, L. PLoS ONE 2018, 13, e0190924. [Google Scholar] [CrossRef] [Green Version]
- Saenjundaeng, P.; Oliveira, E.A.; Tanomtong, A.; Supiwong, W.; Phimphan, S.; Collares-Pereira, M.J.; Sember, A.; Yano, C.F.; Hatanaka, T.; Bertollo, L.A.C.; et al. Chromosomes of Asian cyprinid fishes: Cytogenetic analysis of two representatives of small paleotetraploid tribe Probarbini. Mol. Cytogenet. 2018, 11, 51. [Google Scholar] [CrossRef]
- Rocchi, M.; Archidiacono, N.; Schempp, W.; Capozzi, O.; Stanyon, R. Centromere repositioning in mammals. Heredity 2012, 108, 59–67. [Google Scholar] [CrossRef]
- Schubert, I. What is behind “centromere repositioning”? Chromosoma 2018, 127, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Souza, I.L.; Moreira-Filho, O. Constitutive heterochromatin and Ag-NOR polymorphisms in the small characid fish Astyanax scabripinnis (Jenyns, 1842). Comp. Gen. Pharm. 2007, 72, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, D.T.; Porto-Foresti, F. Chromosome polymorphism of heterochromatin and nucleolar regions in two populations of the fish Astyanax bockmanni (Teleostei: Characiformes). Neotrop. Ichthyol. 2010, 8, 861–866. [Google Scholar] [CrossRef]
- Baumgärtner, L.; Paiz, L.M.; Zawadzki, C.H.; Margarido, V.P.; Castro, A.L.B.P. Heterochromatin polymorphism and physical mapping of 5S and 18S ribosomal DNA in four populations of Hypostomus strigaticeps (Regan, 1907) from the Paraná River basin, Brazil: Evolutionary and environmental correlation. Zebrafish 2014, 11, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Utsunomia, R.; Pansonato-Alves, J.C.; Costa-Silva, G.J.; Mendonça, F.F.; Scacchetti, P.C.; Oliveira, C.; Foresti, F. Molecular and cytogenetic analyses of cryptic species within the Synbranchus marmoratus Bloch, 1795 (Synbranchiformes: Synbranchidae) grouping: Species delimitations, karyotypic evolution and intraspecific diversification. Neotrop. Ichthyol. 2014, 12, 903–911. [Google Scholar] [CrossRef] [Green Version]
- Cioffi, M.B.; Bertollo, L.A.C. Chromosomal distribution and evolution of repetitive DNAs in fish. Genome Dyn. 2012, 7, 197–221. [Google Scholar] [CrossRef]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.; Rodriguez, S.; Pardo, B.G.; Sanchez, L.; Martinez, P. Population analysis of an unusual NOR-site polymorphism in brown trout (Salmo trutta L.). Heredity 2001, 86, 291–302. [Google Scholar] [CrossRef]
- Piscor, D.; Alves, A.L.; Parise-Maltempi, P.P. Chromosomal microstructure diversity in three Astyanax (Characiformes, Characidae) species: Comparative analysis of the chromosomal locations of the 18S and 5S rDNAs. Zebrafish 2015, 12, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Traldi, J.B.; Vicari, M.R.; Martinez, J.D.F.; Blanco, D.R.; Lui, R.L.; Moreira-Filho, O. Chromosome analyses of Apareiodon argenteus and Apareiodon davisi (Characiformes, Parodontidae): An extensive chromosomal polymorphism of 45S and 5S ribosomal DNAs. Zebrafish 2016, 13, 19–25. [Google Scholar] [CrossRef]
- Da Silva, A.F.; Feldberg, E.; Moura Carvalho, N.D.; Hernández Rangel, S.M.; Schneider, C.H.; Carvalho-Zilse, G.A.; Fonsêca da Silva, V.; Gross, M.C. Effects of environmental pollution on the rDNAomics of Amazonian fish. Environ. Pollut. 2019, 252, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.A.; Aboim, M.A.; Ráb, P.; Collares-Pereira, M.J. Introgressive hybridization as a promoter of genome reshuffling in natural homoploid fish hybrids (Cyprinidae, Leuciscinae). Heredity 2014, 112, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cioffi, M.B.; Molina, W.F.; Artoni, R.F.; Bertollo, L.A.C. Chromosomes as tools for discovering biodiversity—The case of Erythrinidae fish family. Recent Trends Cytogenet. Stud. Methodol. Appl. 2012, 125–146. [Google Scholar] [CrossRef] [Green Version]
- Prizon, A.C.; Bruschi, D.P.; Borin-Carvalho, L.A.; Cius, A.; Barbosa, L.M.; Ruiz, H.B.; Zawadzki, C.H.; Fenocchio, A.S.; Portela-Castro, A.L.B. Hidden diversity in the populations of the armored catfish Ancistrus Kner, 1854 (Loricariidae, Hypostominae) from the Paraná River Basin revealed by molecular and cytogenetic data. Front. Genet. 2017, 8, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirchio, M.; Paim, F.G.; Milana, V.; Rossi, A.R.; Oliveira, C. Identification of a new mullet species complex based on an integrative molecular and cytogenetic investigation of Mugil hospes (Mugilidae: Mugiliformes). Front. Genet. 2018, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.P.; Gui, J.F. Identification of genome organization in the unusual allotetraploid form of Carassius auratus gibelio. Aquaculture 2007, 265, 109–117. [Google Scholar] [CrossRef]
- Zhang, C.; Ye, L.; Chen, Y.; Xiao, J.; Wu, Y.; Tao, M.; Xiao, Y.; Liu, S. The chromosomal constitution of fish hybrid lineage revealed by 5S rDNA FISH. BMC Genet. 2015, 16, 140. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.P.; Ma, D.M.; Gui, J.F. Triploid origin of the gibel carp as revealed by 5S rDNA localization and chromosome painting. Chromosome Res. 2006, 14, 767–776. [Google Scholar] [CrossRef]
- Li, Y.J.; Tian, Y.; Zhang, M.Z.; Tian, P.P.; Yu, Z.; Abe, S.; Arai, K. Chromosome banding and FISH with rDNA probe in the diploid and tetraploid loach Misgurnus anguillicaudatus. Ichthyol. Res. 2010, 57, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, M.; Matoso, D.A.; Ludwig, L.A.M.; Gomes, E.; Almeida, M.C.; Vicari, M.R.; Artoni, R.F. Natural triploidy in Rhamdia quelen identified by cytogenetic monitoring in Iguaçu basin, southern Brazil. Environ. Biol. Fishes 2011, 91, 361–366. [Google Scholar] [CrossRef]
- Spóz, A.; Boroń, A.; Ocalewicz, K.; Kirtiklis, L. Polymorphism of the rDNA chromosomal regions in the weatherfish Misgurnus fossilis (Teleostei: Cobitidae). Folia Biol. 2017, 65, 63–70. [Google Scholar] [CrossRef]
- Symonová, R.; Majtánová, Z.; Sember, A.; Staaks, G.B.; Bohlen, J.; Freyhof, J.; Rábová, M.; Ráb, P. Genome differentiation in a species pair of coregonine fishes: An extremely rapid speciation driven by stress-activated retrotransposons mediating extensive ribosomal DNA multiplications. BMC Evol. Biol. 2013, 13, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima-Filho, P.A.; Bertollo, L.A.C.; Cioffi, M.B.; Costa, G.W.W.F.; Molina, W.F. Karyotype divergence and spreading of 5S rDNA sequences between genomes of two species: Darter and emerald gobies (Ctenogobius, Gobiidae). Cytogenet. Genome Res. 2014, 142, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Mazzei, F.; Ghigliotti, L.; Bonillo, C.; Coutanceau, J.-P.; Ozouf-Costaz, C.; Pisano, E. Chromosomal patterns of major and 5S ribosomal DNA in six icefish species (Perciformes, Notothenioidei, Channichthyidae). Polar Biol. 2004, 28, 47–55. [Google Scholar] [CrossRef]
- Ziemniczak, K.; Barros, A.V.; Rosa, K.O.; Nogaroto, V.; Almeida, M.C.; Cestari, M.M.; Moreira-Filho, O.; Artoni, R.F.; Vicari, M.R. Comparative cytogenetics of Loricariidae (Actinopterygii: Siluriformes): Emphasis in Neoplecostominae and Hypoptopomatinae. Ital. J. Zool. 2012, 79, 492–501. [Google Scholar] [CrossRef]
- Piscor, D.; Fernandes, C.A.; Parise-Maltempi, P.P. Conserved number of U2 snDNA sites in Piabina argentea, Piabarchus stramineus and two Bryconamericus species (Characidae, Stevardiinae). Neotrop. Ichthyol. 2018, 16, e170066. [Google Scholar] [CrossRef] [Green Version]
- Yano, C.F.; Merlo, M.A.; Portela-Bens, S.; Cioffi, M.B.; Bertollo, L.A.C.; Santos-Júnior, C.D.; Rebordinos, L.; Esteban, M.A.; Albert, J.S. Evolutionary dynamics of multigene families in Triportheus (Characiformes, Triportheidae): A transposon mediated mechanism? Front. Mar. Sci. 2020, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Inafuku, J.; Nabeyama, M.; Kikuma, Y.; Saitoh, J.; Kubota, S.; Kohno, S.I. Chromosomal location and nucleotide sequences of 5S ribosomal DNA of two cyprinid species (Osteichthyes, Pisces). Chromosome Res. 2000, 8, 193–199. [Google Scholar] [CrossRef]
- He, W.; Qin, Q.; Liu, S.; Li, T.; Wang, J.; Xiao, J.; Xie, L.; Zhang, C.; Liu, Y. Organization and variation analysis of 5S rDNA in different ploidy-level hybrids of red crucian carp × topmouth culter. PLoS ONE 2012, 7, e38976. [Google Scholar] [CrossRef] [Green Version]
- Rábová, M.; Ráb, P.; Ozouf-Costaz, C. Extensive polymorphism and chromosomal characteristics of ribosomal DNA in a loach fish, Cobitis vardarensis (Ostariophysi, Cobitidae) detected by different banding techniques and fluorescence in situ hybridization (FISH). Genetica 2001, 111, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Boroń, A.; Ozouf-Costaz, C.; Coutanceau, J.-P.; Woroniecka, K. Gene mapping of 28S and 5S rDNA sites in the spined loach Cobitis taenia (Pisces, Cobitidae) from a diploid population and a diploid-tetraploid population. Genetica 2006, 128, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Gromicho, M.; Coutanceau, J.-P.; Ozouf-Costaz, C.; Collares-Pereira, M.J. Contrast between extensive variation of 28S rDNA and stability of 5S rDNA and telomeric repeats in the diploid-polyploid Squalius alburnoides complex and in its maternal ancestor Squalius pyrenaicus (Teleostei, Cyprinidae). Chromosome Res. 2006, 14, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.A.; Ráb, P.; Collares-Pereira, M.J. Chromosomes of European cyprinid fishes: Comparative cytogenetics and chromosomal characteristics of ribosomal DNAs in nine Iberian chondrostomine species (Leuciscinae). Genetica 2012, 140, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.R.; Milana, V.; Hett, A.K.; Tancioni, L. Molecular cytogenetic analysis of the Appenine endemic cyprinid fish Squalius lucumonis and three other Italian leuciscines using chromosome banding and FISH with rDNA probes. Genetica 2012, 140, 469–476. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, R.; Nagpure, N.S.; Kushwaha, B.; Mani, I.; Lakra, W.S. Extensive NOR site polymorphism in geographically isolated populations of Golden mahseer, Tor putitora. Genome 2009, 52, 783–789. [Google Scholar] [CrossRef]
- Libertini, A.; Sola, L.; Rampin, M.; Rossi, A.R.; Iijima, K.; Ueda, T. Classical and molecular cytogenetic characterization of allochthonous European bitterling Rhodeus amarus (Cyprinidae, Acheilognathinae) from Northern Italy. Genes Genet. Syst. 2008, 83, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Han, C.C.; Yen, T.B.; Chen, N.C.; Tseng, M.C. Comparative studies of 5S rDNA profiles and Cyt b sequences in two Onychostoma species (Cyprinidae). Int. J. Mol. Sci. 2015, 16, 29663–29672. [Google Scholar] [CrossRef]
- Kirtiklis, L.; Porycka, K.; Boroń, A.; Coutanceau, J.-P.; Dettai, A. Use of the chromosomal co-location of the minor 5S and the major 28S rDNA as a cytogenetic marker within the genus Leuciscus (Pisces, Cyprinidae). Folia Biol. 2010, 58, 245–249. [Google Scholar] [CrossRef]
- Merlo, M.A.; Cross, I.; Rodríguez-Rúa, A.; Manchado, M.; Rebordinos, L. First approach to studying the genetics of the meagre (Argyrosomus regius; Asso, 1801) using three multigene families. Aquac. Res. 2013, 44, 974–984. [Google Scholar] [CrossRef]
- Scacchetti, P.C.; Utsunomia, R.; Pansonato-Alves, J.C. Repetitive DNA sequences and evolution of ZZ/ZW sex chromosomes in Characidium (Teleostei: Characiformes). PLoS ONE 2015, 10, e0137231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, C.F.; Bertollo, L.A.C.; Rebordinos, L.; Merlo, M.A.; Liehr, T.; Portela-Bens, S.; Cioffi, M.B. Evolutionary dynamics of rDNAs and U2 small nuclear DNAs in Triportheus (Characiformes, Triportheidae): High variability and particular syntenic organization. Zebrafish 2017, 14, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Araya-Jaime, C.; Mateussi, N.T.B.; Utsunomia, R.; Costa-Silva, G.J.; Oliveira, C.; Foresti, F. ZZ/Z0: The new system of sex chromosomes in Eigenmannia aff. trilineata (Teleostei: Gymnotiformes: Sternopygidae) characterized by molecular cytogenetics and DNA barcoding. Zebrafish 2017, 14, 464–470. [Google Scholar] [CrossRef]
- Ponzio, J.C.; Piscor, D.; Parise-Maltempi, P.P. Chromosomal locations of U2 snDNA clusters in Megaleporinus, Leporinus and Schizodon (Characiformes: Anostomidae). Biologia 2018, 73, 295–298. [Google Scholar] [CrossRef]
- Cabral-de-Mello, D.C.; Valente, G.T.; Nakajima, R.T.; Martins, C. Genomic organization and comparative chromosome mapping of the U1 snRNA gene in cichlid fish, with an emphasis in Oreochromis niloticus. Chromosome Res. 2012, 20, 279–292. [Google Scholar] [CrossRef]
- Carvalho, P.C.; Oliveira, E.A.; Bertollo, L.A.C.; Yano, C.F.; Al-Rikabi, A.B.H.; Cioffi, M.B. First chromosomal analysis in Hepsetidae (Actinopterygii, Characiformes): Insights into relationship between African and Neotropical fish groups. Front. Genet. 2017, 8, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Souto, D.; Troncoso, T.; Pérez, M.; Pasantes, J.J. Molecular cytogenetic analysis of the european hake Merluccius merluccius (Merlucciidae, Gadiformes): U1 and U2 snRNA gene clusters map to the same location. PLoS ONE 2015, 10, e0146150. [Google Scholar] [CrossRef]
- Piscor, D.; Paiz, L.M.; Baumgärtner, L.; Cerqueira, F.J.; Fernandes, C.A.; Lui, R.L.; Parise-Maltempi, P.P.; Margarido, V.P. Chromosomal mapping of repetitive sequences in Hyphessobrycon eques (Characiformes, Characidae): A special case of the spreading of 5S rDNA clusters in a genome. Genetica 2020, 148, 25–32. [Google Scholar] [CrossRef]
- Usso, M.C.; dos Santos, A.R.; Gouveia, J.G.; Frantine-Silva, W.; Araya-Jaime, C.; Oliveira, M.L.M.; Foresti, F.; Giuliano-Caetano, L.; Dias, A.L. Genetic and chromosomal differentiation of Rhamdia quelen (Siluriformes, Heptapteridae) revealed by repetitive molecular markers and DNA barcoding. Zebrafish 2019, 16, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Cremer, M.; Von Hase, J.; Volm, T.; Brero, A.; Kreth, G.; Walter, J.; Fischer, C.; Solovei, I.; Cremer, C.; Cremer, T. Non-random radial higher-order chromatin arrangements in nuclei of diploid human cells. Chromosome Res. 2001, 9, 541–567. [Google Scholar] [CrossRef]
- Úbeda-Manzanaro, M.; Merlo, M.A.; Palazón, J.L.; Cross, I.; Sarasquete, C.; Rebordinos, L. Chromosomal mapping of the major and minor ribosomal genes, (GATA)n and U2 snRNA gene by double-colour FISH in species of the Batrachoididae family. Genetica 2010, 138, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Malimpensa, G.C.; Traldi, J.B.; Toyama, D.; Henrique-Silva, F.; Vicari, M.R.; Moreira-Filho, O. Chromosomal mapping of repeat DNA in Bergiaria westermanni (Pimelodidae, Siluriformes): Localization of 45S rDNA in B chromosomes. Cytogenet. Genome Res. 2018, 154, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Manchado, M.; Zuasti, E.; Cross, I.; Merlo, A.; Infante, C.; Rebordinos, L. Molecular characterization and chromosomal mapping of the 5S rRNA gene in Solea senegalensis: A new linkage to the U1, U2, and U5 small nuclear RNA genes. Genome 2006, 49, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, A.; Nergadze, S.G.; Santagostino, M.; Giulotto, E. Telomeric repeats far from the ends: Mechanisms of origin and role in evolution. Cytogenet. Genome Res. 2009, 122, 219–228. [Google Scholar] [CrossRef]
- Zhou, M.; Kang, Y.; Li, Y.C.; Zhou, T. Studies on silver-stained karyotypes of 7 species in Cyprinidae (Pisces). Zool. Res 1988, 9, 225–229. (In Chinese) [Google Scholar]
- Ren, X.H.; Yu, X.J. Characterization of nucleolar organizer regions of twelve species of Chinese cyprinid fishes. Caryologia 1993, 46, 201–207. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, K.; Hong, Y.H.; Gui, J.F.; Zhou, T. Studies on karyotypes of Chinese cyprinid fishes. VII. Karyotypic analyses of seven species in the subfamily Leuciscinae with a consideration for the phylogenetic relationships of some cyprinid fishes concerned. Acta Genet. Sin. 1985, 12, 367–372. (In Chinese) [Google Scholar]
- Li, K.; Li, Y.C.; Zhou, M.; Zhou, T. Studies on the karyotypes of Chinese cyprinid fishes. II. Karyotypes of four species of Xenocyprininae. Acta Zool. Sin. 1983, 29, 207–213. (In Chinese) [Google Scholar]
- Zan, R.G.; Song, Z. Analysis and comparison between the karyotypes of Ctenopharyngodon idella and Megalobrama amblycephala. Acta Genet. Sin. 1979, 6, 205–210. (In Chinese) [Google Scholar]
- Lu, R.H.; Li, Y.J.; Xu, K.S. A chromosome study of Megalobrama amblycephala. Oceanol. Limnol. Sin. 1984, 15, 487–492. (In Chinese) [Google Scholar]
- Liu, Y.U. On the karyotype of the grass-carp Ctenopharyngodon Idella. Acta Zool. Sin 1980, 26, 14–16. (In Chinese) [Google Scholar]
- Lou, Y.; Zhang, K.; Wu, Y.; Wang, Y. Studies on karyotype of black carp (Mylopharyngodon piceus). J. Fish. China 1983, 7, 77–81. (In Chinese) [Google Scholar]
- Arai, R. Fish Karyotypes: A Check List, 1st ed.; Springer: Tokyo, Japan, 2011. [Google Scholar]
- Ráb, P.; Crossman, E.J. Chromosomal NOR phenotypes in North American pikes and pickerels, genus Esox with notes on Umbridae (Euteleostei: Esocae). Can. J. Zool. 1994, 72, 1951–1956. [Google Scholar] [CrossRef]
- Symonová, R.; Ocalewicz, K.; Kirtiklis, L.; Delmastro, G.B.; Pelikánová, Š.; Garcia, S.; Kovařík, A. Higher-order organisation of extremely amplified, potentially functional and massively methylated 5S rDNA in European pikes (Esox sp.). BMC Genom. 2017, 18, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, R.B.; Ráb, P. Chromosome evolution in the Salmonidae (Pisces): An update. Biol. Rev. Camb. Philos. Soc. 2001, 76, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Dion-Côté, A.-M.; Symonová, R.; Lamaze, F.; Pelikánová, Š.; Ráb, P.; Bernatchez, L. Standing chromosomal variation in Lake Whitefish species pairs: The role of historical contingency and relevance for speciation. Mol. Ecol. 2017, 26, 178–192. [Google Scholar] [CrossRef]
- Barby, F.F.; Bertollo, L.A.C.; Oliveira, E.A.; Yano, C.F.; Hatanaka, T.; Ráb, P.; Sember, A.; Ezaz, T.; Artoni, R.F.; Liehr, T.; et al. Emerging patterns of genome organization in Notopteridae species (Teleostei, Osteoglossiformes) as revealed by Zoo-FISH and Comparative Genomic Hybridization (CGH). Sci. Rep. 2019, 9, 1112. [Google Scholar] [CrossRef]
- Molina, W.F. Chromosomal changes and stasis in marine fish groups. In Fish Cytogenetics, 1st ed.; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; Science Publishers: Enfield, CT, USA, 2007; pp. 69–110. [Google Scholar]
- Motta-Neto, C.C.; Cioffi, M.B.; Bertollo, L.A.C.; Molina, W.F. Extensive chromosomal homologies and evidence of karyotypic stasis in Atlantic grunts of the genus Haemulon (Perciformes). J. Exp. Mar. Biol. Ecol. 2011, 401, 75–79. [Google Scholar] [CrossRef]
- Motta-Neto, C.C.; Cioffi, M.B.; Bertollo, L.A.C.; Molina, W.F. Molecular cytogenetic analysis of Haemulidae fish (Perciformes): Evidence of evolutionary conservation. J. Exp. Mar. Biol. Ecol. 2011, 407, 97–100. [Google Scholar] [CrossRef]
- Motta-Neto, C.C.; Lima-Filho, P.A.; Araújo, W.C.; Bertollo, L.A.C.; Molina, W.F. Differentiated evolutionary pathways in Haemulidae (Perciformes): Karyotype stasis versus morphological differentiation. Rev. Fish Biol. Fish. 2012, 22, 457–465. [Google Scholar] [CrossRef]
- Motta-Neto, C.C.; Cioffi, M.B.; Costa, G.W.W.F.; Amorim, K.D.J.; Bertollo, L.A.C.; Artoni, R.F.; Molina, W.F. Overview on karyotype stasis in Atlantic grunts (Eupercaria, Haemulidae) and the evolutionary extensions for other marine fish groups. Front. Mar. Sci. 2019, 6, 628. [Google Scholar] [CrossRef] [Green Version]
- Ellegren, H. Evolutionary stasis: The stable chromosomes of birds. Trends Ecol. Evol. 2010, 25, 283–291. [Google Scholar] [CrossRef]
- Tian, Y.; Nie, W.; Wang, J.; Ferguson-Smith, M.A.; Yang, F. Chromosome evolution in bears: Reconstructing phylogenetic relationships by cross-species chromosome painting. Chromosome Res. 2004, 12, 55–63. [Google Scholar] [CrossRef]
- Razin, S.V.; Gavrilov, A.A.; Vassetzky, Y.S.; Ulianov, S.V. Topologically-associating domains: Gene warehouses adapted to serve transcriptional regulation. Transcription 2016, 7, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Rosin, L.F.; Crocker, O.; Isenhart, R.L.; Nguyen, S.C.; Xu, Z.; Joyce, E.F. Chromosome territory formation attenuates the translocation potential of cells. eLife 2019, 8, e49553. [Google Scholar] [CrossRef]
- Alves, M.J.; Coelho, M.M.; Collares-Pereira, M.J. Evolution in action through hybridization and polyploid in an Iberain freshwater fish: A genetic review. Genetica 2001, 111, 375–385. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | 2n | Karyotype Composition | NF | References | |||
|---|---|---|---|---|---|---|---|
| m | sm | st | a | ||||
| H. nobilis | 48 | 20 | 16 | 12 | 84 | [15,16,17,18] | |
| 48 | - 26 - | 20 | 2 | 74 | [19] | ||
| 48 | 18 | 30 | 96 | [20,21] | |||
| 48 | 14 | 24 | 10 | 86 | [22,23] | ||
| 48 | 6 | 36 | 6 | 96 | [24,25] | ||
| 48 | 26 | 20 | 2 | 96 | [26] | ||
| 48 | - 24 - | 24 | 72 | [27] | |||
| H. molitrix | 48 | 10 | - 26 - | 12 | 84 | [28] | |
| 48 | 20 | 12 | 6 | 10 | 82 | [29] | |
| 48 | - 20 - | - 28 - | 68 | [19] | |||
| 48 | 22 | 14 | 12 | 84 | [15,16] | ||
| 48 | 14 | 24 | 10 | 86 | [22,23] | ||
| 48 | 24 | 16 | 8 | 96 | [30] | ||
| 48 | 20 | 24 | 4 | 96 | [31] | ||
| 48 | 18 | 22 | 8 | 88 | [32] | ||
| 48 | - 24 - | 24 | 72 | [27] | |||
| 48 | 18 | 22 | 8 | 92 | [33] | ||
| Species | 2n | Karyotype Composition | NF | References | |||
|---|---|---|---|---|---|---|---|
| m | sm | st | a | ||||
| Elopichthys bambusa | 48 | 10 | 24 | 12 | 2 | 82 | [112,114] |
| Luciobrama microcephalus | 48 | 12 | 22 | 12 | 2 | 82 | [114] |
| Ochetobius elongates | 48 | 10 | 16 | 22 | 74 | [114] | |
| Squaliobarbus curriculus | 48 | 14 | 30 | 4 | 92 | [114] | |
| Culter oxycephaloides | 48 | 20 | 24 | 4 | 92 | [115] | |
| Xenocypris macrolepis | 48 | 20 | 26 | 2 | 94 | [112,115] | |
| Xenocypris davidi | 48 | 18 | 26 | 4 | 92 | [112,115] | |
| Xenocypris fangi | 48 | 16 | 28 | 4 | 92 | [112] | |
| Xenocypris sechuanensis | 48 | 18 | 26 | 4 | 92 | [112] | |
| Megalobrama amblycephala | 48 | 18 | 26 | 4 | 92 | [115,116,117] | |
| Megalobrama terminalis | 48 | 18 | 22 | 8 | 88 | [115] | |
| Ctenopharyngodon idella | 48 | 18 | 24 | 6 | 90 | [115] | |
| 48 | 18 | 30 | 96 | [118] | |||
| 48 | 18 | 22 | 8 | 88 | [116] | ||
| Mylopharyngodon piceus | 48 | 14 | - 34 - | 96 | [25] | ||
| 48 | 16 | 28 | 4 | 92 | [119] | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sember, A.; Pelikánová, Š.; de Bello Cioffi, M.; Šlechtová, V.; Hatanaka, T.; Do Doan, H.; Knytl, M.; Ráb, P. Taxonomic Diversity Not Associated with Gross Karyotype Differentiation: The Case of Bighead Carps, Genus Hypophthalmichthys (Teleostei, Cypriniformes, Xenocyprididae). Genes 2020, 11, 479. https://doi.org/10.3390/genes11050479
Sember A, Pelikánová Š, de Bello Cioffi M, Šlechtová V, Hatanaka T, Do Doan H, Knytl M, Ráb P. Taxonomic Diversity Not Associated with Gross Karyotype Differentiation: The Case of Bighead Carps, Genus Hypophthalmichthys (Teleostei, Cypriniformes, Xenocyprididae). Genes. 2020; 11(5):479. https://doi.org/10.3390/genes11050479
Chicago/Turabian StyleSember, Alexandr, Šárka Pelikánová, Marcelo de Bello Cioffi, Vendula Šlechtová, Terumi Hatanaka, Hiep Do Doan, Martin Knytl, and Petr Ráb. 2020. "Taxonomic Diversity Not Associated with Gross Karyotype Differentiation: The Case of Bighead Carps, Genus Hypophthalmichthys (Teleostei, Cypriniformes, Xenocyprididae)" Genes 11, no. 5: 479. https://doi.org/10.3390/genes11050479