Fibrillarin Ribonuclease Activity is Dependent on the GAR Domain and Modulated by Phospholipids

 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines, Cell Culture, and Transfection Assays
2.2. Microscopy
2.3. Structured Illumination Microscopy
2.4. Cloning
2.5. Recombinant Protein Expression and Purification
2.6. Exponential Megaprimer PCR (EMP) Strategy to Introduce the GAR Domain Coding Region into RNP Complex
2.7. In Vitro Ligation and Transformation of the EMP Product
2.8. Recombinant RNP Complex Expression and Purification
2.9. SNAP-Tag-Fibrillarin Purification from HeLa Cells
2.10. Western Blot Assay
2.11. Fibrillarin Mutagenesis
2.12. RNA In Vitro Transcription
2.13. RNA Extraction from HeLa Human Cells
2.14. In-Gel RNAse Activity Assay
2.15. In Vitro RNA Activity Assay
2.16. Fat Blot Assay
2.17. Fluorescence Recovery after Photobleaching
2.18. Bioinformatic Analysis
3. Results
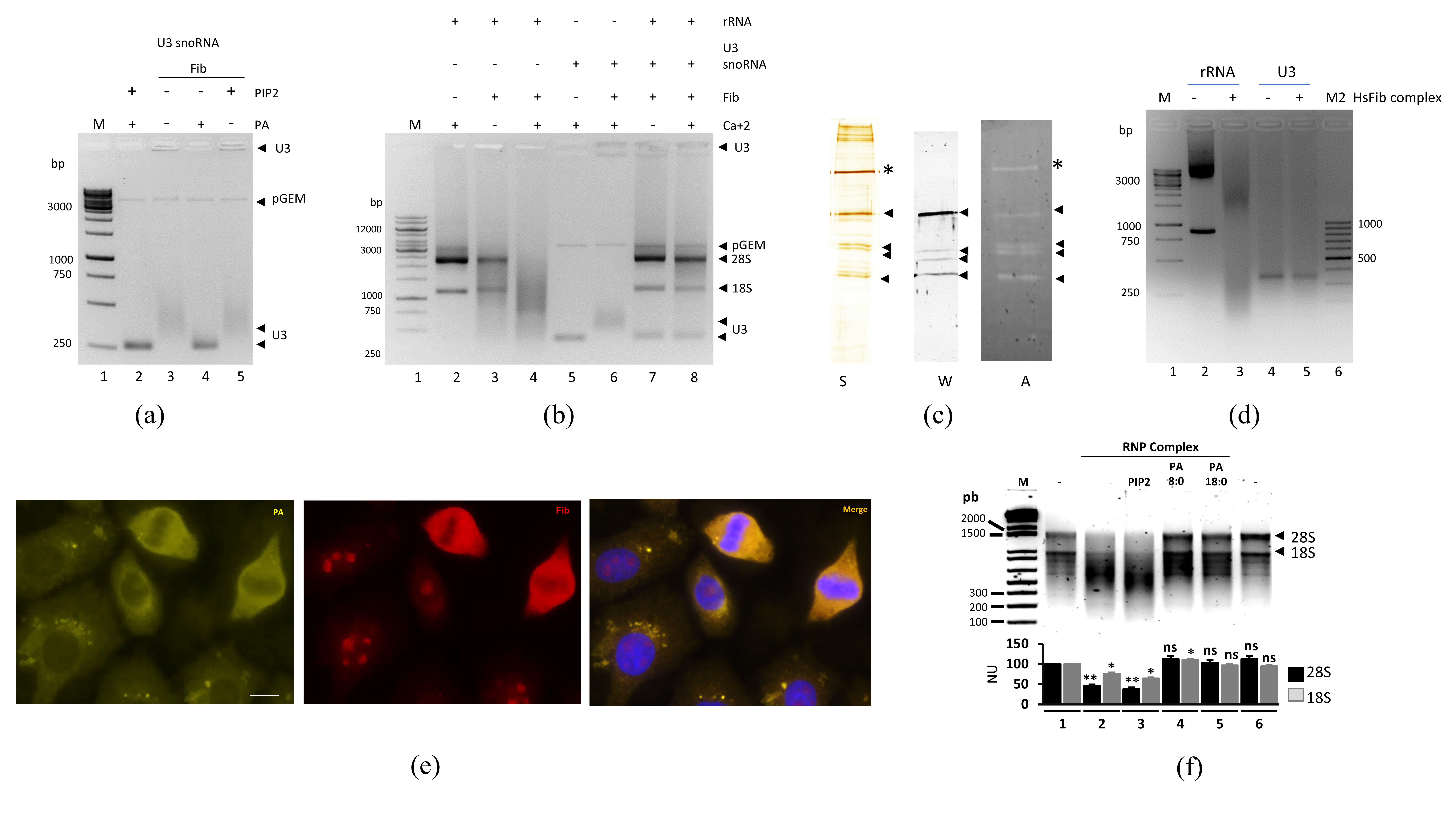
3.1. Fibrillarin as a Ribonuclease
3.2. Ribonuclease Activity of the Recombinant Ribonucleoparticle Complex Involving Fibrillarin
3.3. Fibrillarin Specificity Ribonuclease Activity in Complex with RNA Guide
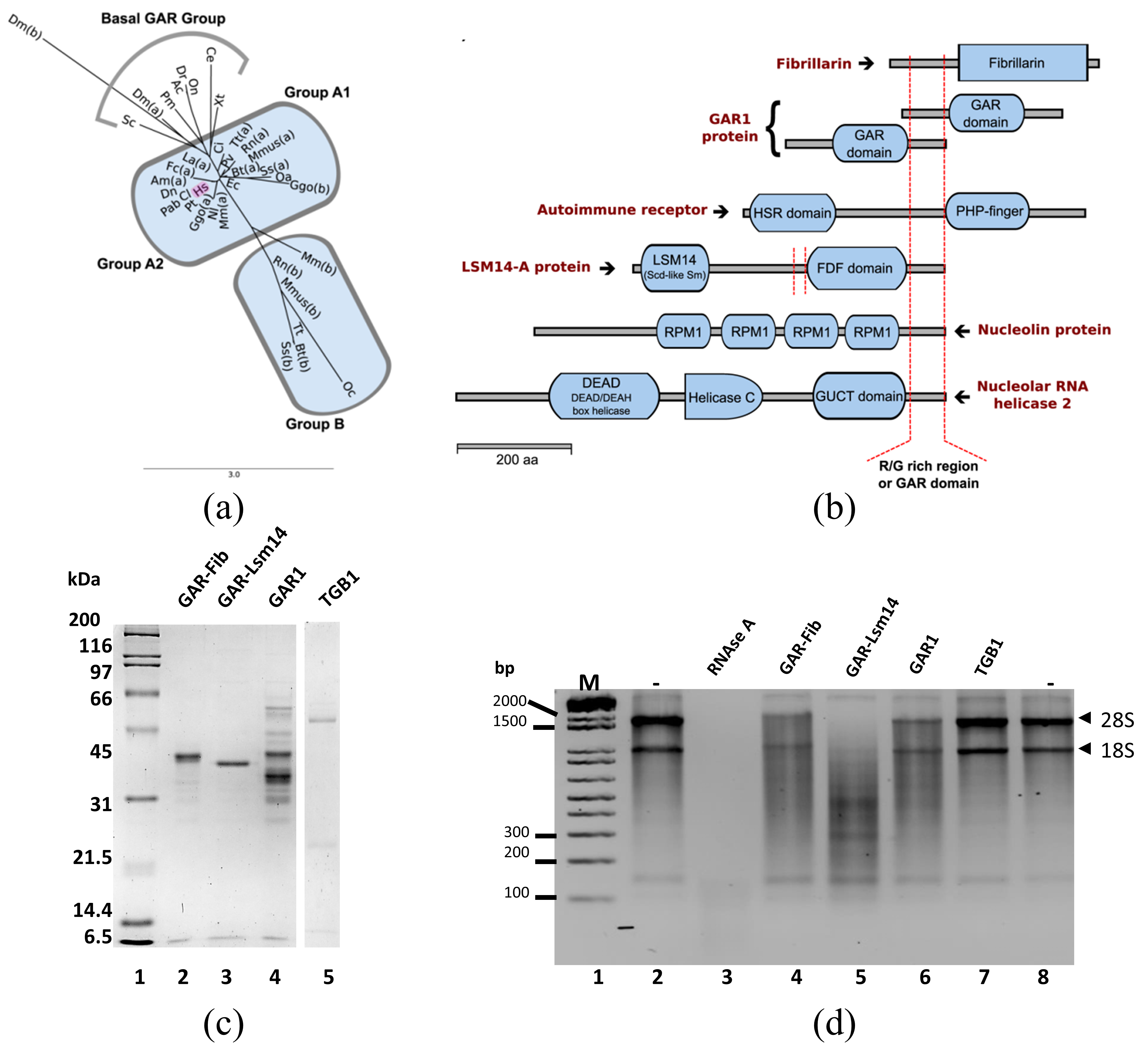
3.4. GAR Domain, Modular in Fibrillarin
3.5. Mutation of the GAR Domain
3.6. Ribonuclease Activity of GAR-Like Domains Containing Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schimmang, T.; Tollervey, D.; Kern, H.; Frank, R.; Hurt, E.C. A yeast nucleolar protein related to mammalian fibrillarin is associated with small nucleolar RNA and is essential for viability. EMBO J. 1989, 8, 4015–4024. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Verdun, D.; Roussel, P.; Thiry, M.; Sirri, V.; Lafontaine, D.L.J. The nucleolus: Structure/function relationship in RNA metabolism. Wiley Interdiscip. Rev. RNA 2010, 1, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, A.; Speckmann, W.; Terns, R.; Terns, M.P. Role of the Box C/D Motif in Localization of Small Nucleolar RNAs to Coiled Bodies and Nucleoli. Mol. Biol. Cell 1999, 10, 2131–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, S.C.; Brangwynne, C.P. Inverse size scaling of the nucleolus by a concentration-dependent phase transition. Curr. Biol. 2015, 25, 641–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brangwynne, C.P.; Mitchison, T.J.; Hyman, A.A. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 4334–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falahati, H.; Wieschaus, E. Independent active and thermodynamic processes govern the nucleolus assembly in vivo. Proc. Natl. Acad. Sci. USA 2017, 114, 1335–1340. [Google Scholar] [CrossRef] [Green Version]
- Berry, J.; Weber, S.C.; Vaidya, N.; Haataja, M.; Brangwynne, C.P. RNA transcription modulates phase transition-driven nuclear body assembly. Proc. Natl. Acad. Sci. USA 2015, 112, E5237–E5245. [Google Scholar] [CrossRef] [Green Version]
- Mitrea, D.M.; Kriwacki, R.W. Phase separation in biology; functional organization of a higher order. Cell Commun. Signal. 2016, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Mitrea, D.M.; Cika, J.A.; Stanley, C.B.; Nourse, A.; Onuchic, P.L.; Banerjee, P.R.; Phillips, A.H.; Park, C.G.; Deniz, A.A.; Kriwacki, R.W. Self-interaction of NPM1 modulates multiple mechanisms of liquid-liquid phase separation. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Rodriguez-Corona, U.; Sobol, M.; Rodriguez-Zapata, L.C.; Hozak, P.; Castano, E. Fibrillarin from Archaea to human. Biol. Cell 2015, 107, 159–174. [Google Scholar] [CrossRef]
- Aris, J.P.; Blobel, G. cDNA cloning and sequencing of human fibrillarin, a conserved nucleolar protein recognized by autoimmune antisera. Proc. Natl. Acad. Sci. USA 1991, 88, 931–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tessarz, P.; Santos-Rosa, H.; Robson, S.C.; Sylvestersen, K.B.; Nelson, C.J.; Nielsen, M.L.; Kouzarides, T. Glutamine methylation in histone H2A is an RNA-polymerase-I-dedicated modification. Nature 2014, 505, 564–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loza-Muller, L.; Rodríguez-Corona, U.; Sobol, M.; Rodríguez-Zapata, L.C.; Hozak, P.; Castano, E. Fibrillarin methylates H2A in RNA polymerase I trans-active promoters in Brassica oleracea. Front. Plant Sci. 2015, 6, 976. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Wurm, T.; Britton, P.; Brooks, G.; Hiscox, J.A. Interaction of the coronavirus nucleoprotein with nucleolar antigens and the host cell. J. Virol. 2002, 76, 5233–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponti, D.; Troiano, M.; Bellenchi, G.C.; Battaglia, P.A.; Gigliani, F. The HIV Tat protein affects processing of ribosomal RNA precursor. BMC Cell Biol. 2008, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melén, K.; Tynell, J.; Fagerlund, R.; Roussel, P.; Hernandez-Verdun, D.; Julkunen, I. Influenza A H3N2 subtype virus NS1 protein targets into the nucleus and binds primarily via its C-terminal NLS2/NoLS to nucleolin and fibrillarin. Virol. J. 2012, 9, 167. [Google Scholar] [CrossRef]
- Fournier, M.J.; Maxwell, E.S. The nucleolar snRNAs: Catching up with the spliceosomal snRNAs. Trends Biochem. Sci. 1993, 18, 131–135. [Google Scholar] [CrossRef]
- Kass, S.; Tyc, K.; Steitz, J.A.; Sollner-Webb, B. The U3 small nucleolar ribonucleoprotein functions in the first step of preribosomal RNA processing. Cell 1990, 60, 897–908. [Google Scholar] [CrossRef]
- Hughes, J.M.; Ares, M. Depletion of U3 small nucleolar RNA inhibits cleavage in the 5’ external transcribed spacer of yeast pre-ribosomal RNA and impairs formation of 18S ribosomal RNA. EMBO J. 1991, 10, 4231–4239. [Google Scholar] [CrossRef]
- Dragon, F.; Lemay, V.; Trahan, C. snoRNAs: Biogenesis, structure and function. Encycl. Life Sci. 2006, 1–7. [Google Scholar] [CrossRef]
- Granneman, S.; Vogelzangs, J.; Lührmann, R.; van Venrooij, W.J.; Pruijn, G.J.M.; Watkins, N.J. Role of pre-rRNA base pairing and 80S complex formation in subnucleolar localization of the U3 snoRNP. Mol. Cell. Biol. 2004, 24, 8600–8610. [Google Scholar] [CrossRef] [Green Version]
- Sáez-Vasquez, J.; Caparros-Ruiz, D.; Barneche, F.; Echeverría, M. Characterization of a crucifer plant pre-rRNA processing complex. Biochem. Soc. Trans. 2004, 32, 578–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Protter, D.S.W.; Rosen, M.K.; Parker, R. Formation and maturation of phase-separated liquid droplets by RNA-binding proteins. Mol. Cell 2015, 60, 208–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.-L.; Erce, M.A.; Lai, Y.-W.; Tomasetig, F.; Hart-Smith, G.; Hamey, J.J.; Wilkins, M.R. Crosstalk of phosphorylation and arginine methylation in disordered SRGG repeats of S. cerevisiae fibrillarin and its association with nucleolar localisation. J. Mol. Biol. 2019, 432, 448–466. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, S.; Castano, E.; Sobol, M.; Philimonenko, V.V.; Dzijak, R.; Venit, T.; Hozak, P.; Hozák, P. Involvement of phosphatidylinositol 4,5-bisphosphate in RNA polymerase I transcription. J. Cell Sci. 2013, 126, 2730–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobol, M.; Yildirim, S.; Philimonenko, V.V.; Marášek, P.; Castaño, E.; Hozák, P. UBF complexes with phosphatidylinositol 4,5-bisphosphate in nucleolar organizer regions regardless of ongoing RNA polymerase I activity. Nucleus 2013, 4, 478–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamann, B.L.; Blind, R.D. Nuclear phosphoinositide regulation of chromatin. J. Cell. Physiol. 2017, 233, 107–123. [Google Scholar] [CrossRef]
- Castano, E.; Yildirim, S.; Fáberová, V.; Krausová, A.; Uličná, L.; Paprčková, D.; Sztacho, M.; Hozák, P. Nuclear phosphoinositides—Versatile regulators of genome functions. Cells 2019, 8, 649. [Google Scholar] [CrossRef] [Green Version]
- Kalasova, I.; Fáberová, V.; Kalendová, A.; Yildirim, S.; Uličná, L.; Venit, T.; Hozák, P. Tools for visualization of phosphoinositides in the cell nucleus. Histochem. Cell Biol. 2016, 145, 485–496. [Google Scholar] [CrossRef]
- Sobol, M.; Krausová, A.; Yildirim, S.; Kalasová, I.; Fáberová, V.; Vrkoslav, V.; Philimonenko, V.; Marášek, P.; Pastorek, L.; Čapek, M.; et al. Nuclear phosphatidylinositol 4,5-bisphosphate islets contribute to efficient RNA polymerase II-dependent transcription. J. Cell Sci. 2018, 131, jcs211094. [Google Scholar] [CrossRef] [Green Version]
- Arraiano, C.M.; Mauxion, F.; Viegas, S.C.; Matos, R.G.; Séraphin, B. Intracellular ribonucleases involved in transcript processing and decay: Precision tools for RNA. Biochim. Biophys. Acta 2013, 1829, 491–513. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, M.P. Twenty years of bacterial RNases and RNA processing: How we’ve matured. RNA 2015, 21, 597–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moelling, K.; Broecker, F. The reverse transcriptase-RNase H: From viruses to antiviral defense. Ann. N. Y. Acad. Sci. 2015, 1341, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Bubeck, D.; Reijns, M.A.M.; Graham, S.C.; Astell, K.R.; Jones, E.Y.; Jackson, A.P. PCNA directs type 2 RNase H activity on DNA replication and repair substrates. Nucleic Acids Res. 2011, 39, 3652–3666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houseley, J.; Tollervey, D. The many pathways of RNA degradation. Cell 2009, 136, 763–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henras, A.K.; Plisson-Chastang, C.; O’Donohue, M.F.; Chakraborty, A.; Gleizes, P.E. An overview of pre-ribosomal RNA processing in eukaryotes. Wiley Interdiscip. Rev. RNA 2015, 6, 225–242. [Google Scholar] [CrossRef]
- Fatica, A.; Galardi, S.; Altieri, F.; Bozzoni, I. Fibrillarin binds directly and specifically to U16 box C/D snoRNA. RNA 2000, 6, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Tschochner, H.; Hurt, E. Pre-ribosomes on the road from the nucleolus to the cytoplasm. Trends Cell Biol. 2003, 13, 255–263. [Google Scholar] [CrossRef]
- Kassas, N.; Tanguy, E.; Thahouly, T.; Fouillen, L.; Heintz, D.; Chasserot-Golaz, S.; Bader, M.-F.; Grant, N.J.; Vitale, N. Comparative characterization of phosphatidic acid sensors and their localization during frustrated phagocytosis. J. Biol. Chem. 2017, 292, 4266–4279. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, A.; Andersen, K.R.; Schwartz, T.U. Exponential Megapriming PCR (EMP) Cloning-Seamless DNA Insertion into Any Target Plasmid without Sequence Constraints. PLoS ONE 2012, 7, e0053360. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Yu, G.; Tian, S.; Li, H. Co-expression and co-purification of archaeal and eukaryal box C/D RNPs. PLoS ONE 2014, 9, e0103096. [Google Scholar] [CrossRef] [PubMed]
- Dudkina, E.; Ulyanova, V.; Shah Mahmud, R.; Khodzhaeva, V.; Dao, L.; Vershinina, V.; Kolpakov, A.; Ilinskaya, O. Three-step procedure for preparation of pure Bacillus altitudinis ribonuclease. FEBS Open Bio 2016, 6, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Huang, S. Nucleolar components involved in ribosome biogenesis cycle between the nucleolus and nucleoplasm in interphase cells. J. Cell Biol. 2001, 153, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Koonin, E.V. Iterated profile searches with PSI-BLAST--a tool for discovery in protein databases. Trends Biochem. Sci. 1998, 23, 444–447. [Google Scholar] [CrossRef]
- Rosenhouse-Dantsker, A.; Logothetis, D.E.; Levitan, I. Cholesterol sensitivity of KIR2.1 is controlled by a belt of residues around the cytosolic pore. Biophys. J. 2011, 100, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Morales, J.; Sobol, M.; Rodriguez-Zapata, L.C.; Hozak, P.; Castano, E. Aromatic amino acids and their relevance in the specificity of the PH domain. J. Mol. Recognit. 2017, 30. [Google Scholar] [CrossRef]
- Min, J.; Wu, H.; Zeng, H.; Loppnau, P.; Weigelt, J.; Sundstrom, M.; Arrowsmith, C.H.; Edwards, A.M.; Bochkarev, A.; Plotnikov, A.N.; et al. Resour. Stud. Biol. Macromol. Protein Data Bank. Available online: https://www.rcsb.org/structure/2IPX. (accessed on 20 April 2019).
- Schwarz, D.S.; Blower, M.D. The calcium-dependent ribonuclease XendoU promotes ER network formation through local RNA degradation. J. Cell Biol. 2014, 207, 41–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidel, C.W.; Peck, L.J. Purification of a calcium dependent ribonuclease from Xenopus laevis. Nucleic Acids Res. 1994, 22, 1456–1462. [Google Scholar] [CrossRef] [Green Version]
- Rosta, E.; Yang, W.; Hummer, G. Calcium inhibition of ribonuclease H1 two-metal ion catalysis. J. Am. Chem. Soc. 2014, 136, 3137–3144. [Google Scholar] [CrossRef]
- Rodriguez-Corona, U.; Pereira-Santana, A.; Sobol, M.; Rodriguez-Zapata, L.C.; Hozak, P.; Castano, E. Novel Ribonuclease Activity Differs between Fibrillarins from Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1878. [Google Scholar] [CrossRef] [Green Version]
- Tran, E.J.; Zhang, X.; Maxwell, E.S. Efficient RNA 2’-O-methylation requires juxtaposed and symmetrically assembled archaeal box C/D and C’/D’ RNPs. EMBO J. 2003, 22, 3930–3940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, N.J.; Bohnsack, M.T. The box C/D and H/ACA snoRNPs: Key players in the modification, processing and the dynamic folding of ribosomal RNA. Wiley Interdiscip. Rev. RNA 2012, 3, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.R.; Kuhn, J.F.; Shanab, G.M.; Maxwell, E.S. Box C/D snoRNA-associated proteins: Two pairs of evolutionarily ancient proteins and possible links to replication and transcription. RNA 2000, 6, 861–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomkuviene, M.; Ličyte, J.; Olendraite, I.; Liutkevičiute, Z.; Clouet-D’Orval, B.; Klimašauskas, S. Archaeal fibrillarin-Nop5 heterodimer 2′-O-methylates RNA independently of the C/D guide RNP particle. RNA 2017, 23, 1329–1337. [Google Scholar] [CrossRef]
- Thandapani, P.; O’Connor, T.R.; Bailey, T.L.; Richard, S. Defining the RGG/RG Motif. Mol. Cell 2013, 50, 613–623. [Google Scholar] [CrossRef] [Green Version]
- McBride, A.E.; Conboy, A.K.; Brown, S.P.; Ariyachet, C.; Rutledge, K.L. Specific sequences within arginine-glycine-rich domains affect mRNA-binding protein function. Nucleic Acids Res. 2009, 37, 4322–4330. [Google Scholar] [CrossRef] [Green Version]
- Gendra, E.; Moreno, A.; Albà, M.M.; Pages, M. Interaction of the plant glycine-rich RNA-binding protein MA16 with a novel nucleolar DEAD box RNA helicase protein from Zea mays. Plant J. 2004, 38, 875–886. [Google Scholar] [CrossRef]
- Donald, R.G.; Lawrence, D.M.; Jackson, A.O. The barley stripe mosaic virus 58-kilodalton beta (b) protein is a multifunctional RNA binding protein. J. Virol. 1997, 71, 1538–1546. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.M.; Jackson, A.O. Requirements for cell-to-cell movement of Barley stripe mosaic virus in monocot and dicot hosts. Mol. Plant Pathol. 2001, 2, 65–75. [Google Scholar] [CrossRef]
- Lawrence, D.M.; Jackson, A.O. Interactions of the TGB1 protein during cell-to-cell movement of Barley stripe mosaic virus. J. Virol. 2001, 75, 8712–8723. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Y.; Jiang, Z.; Jin, X.; Zhang, K.; Wang, X.; Han, C.; Yu, J.; Li, D. Hijacking of the nucleolar protein fibrillarin by TGB1 is required for cell-to-cell movement of Barley stripe mosaic virus. Mol. Plant Pathol. 2018, 19, 1222–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Macfarlane, S.; Kalinina, N.O.; Rakitina, D.V.; Ryabov, E.V.; Gillespie, T.; Haupt, S.; Brown, J.W.S.; Taliansky, M. Interaction of a plant virus-encoded protein with the major nucleolar protein fibrillarin is required for systemic virus infection. Proc. Natl. Acad. Sci. USA 2007, 104, 11115–11120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmott, E.; Hiscox, J.A. Nucleolar targeting: The hub of the matter. EMBO Rep. 2009, 10, 231–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, S.E.; Bhatti, M.M.; Uversky, V.N.; Dunker, A.K.; Sullivan, W.J., Jr. Regions of intrinsic disorder help identify a novel nuclear localization signal in Toxoplasma gondii histone acetyltransferase TgGCN5-B. Mol. Biochem. Parasitol. 2011, 175, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.P.; Hurt, E.C.; Kern, H.; Lehtonen, H.; Carmo-Fonseca, M.; Lapeyre, B.; Tollervey, D. Evolutionary conservation of the human nucleolar protein fibrillarin and its functional expression in yeast. J. Cell Biol. 1991, 113, 715–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tollervey, D.; Lehtonen, H.; Jansen, R.; Kern, H.; Hurt, E.C. Temperature-sensitive mutations demonstrate roles for yeast fibrillarin in pre-rRNA processing, pre-rRNA methylation, and ribosome assembly. Cell 1993, 72, 443–457. [Google Scholar] [CrossRef]
- Peña, C.; Hurt, E.; Panse, V.G. Eukaryotic ribosome assembly, transport and quality control. Nat. Struct. Mol. Biol. 2017, 24, 689–699. [Google Scholar] [CrossRef]
- Correll, C.C.; Bartek, J.; Dundr, M. The Nucleolus: A multiphase condensate balancing ribosome synthesis and translational capacity in health, aging and ribosomopathies. Cells 2019, 8, 869. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.-W.; Xu, G.; Wang, Y.; Shan, L.; Luan, P.-F.; Wang, Y.; Wu, M.; Yang, L.-Z.; Xing, Y.-H.; Yang, L.; et al. Nascent Pre-rRNA Sorting via Phase Separation Drives the Assembly of Dense Fibrillar Components in the Human Nucleolus. Mol. Cell 2019, 76, 767–783. [Google Scholar] [CrossRef]
- Xing, Y.-H.; Yao, R.-W.; Zhang, Y.; Guo, C.-J.; Jiang, S.; Xu, G.; Dong, R.; Yang, L.; Chen, L.-L. SLERT Regulates DDX21 Rings Associated with Pol I Transcription. Cell 2017, 169, 664–678.e16. [Google Scholar] [CrossRef]
- Tollervey, D.; Lehtonen, H.; Carmo-Fonseca, M.; Hurt, E.C. The small nucleolar RNP protein NOP1 (fibrillarin) is required for pre-rRNA processing in yeast. EMBO J. 1991, 10, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Tiku, V.; Kew, C.; Mehrotra, P.; Ganesan, R.; Robinson, N.; Antebi, A. Nucleolar fibrillarin is an evolutionarily conserved regulator of bacterial pathogen resistance. Nat. Commun. 2018, 9, 3607. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.S.; Diloknawarit, P.; Park, B.S.; Chua, N.H. ELF18-INDUCED LONG NONCODING RNA 1 evicts fibrillarin from mediator subunit to enhance PATHOGENESIS-RELATED GENE 1 (PR1) expression. New Phytol. 2019, 221, 2067–2079. [Google Scholar] [CrossRef] [PubMed]
- Lapeyre, B.; Bourbon, H.; Amalric, F. Nucleolin, the major nucleolar protein of growing eukaryotic cells: An unusual protein structure revealed by the nucleotide sequence. Proc. Natl. Acad. Sci. USA 1987, 84, 1472–1476. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.C.; Xue, Z.X.; Mélèse, T. The NSR1 gene encodes a protein that specifically binds nuclear localization sequences and has two RNA recognition motifs. J. Cell Biol. 1991, 113, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jong, A.Y.; Clark, M.W.; Gilbert, M.; Oehm, A.; Campbell, J.L. Saccharomyces cerevisiae SSB1 protein and its relationship to nucleolar RNA-binding proteins. Mol. Cell. Biol. 1987, 7, 2947–2955. [Google Scholar] [CrossRef] [Green Version]
- Girard, J.P.; Lehtonen, H.; Caizergues-Ferrer, M.; Amalric, F.; Tollervey, D.; Lapeyre, B. GAR1 is an essential small nucleolar RNP protein required for pre-rRNA processing in yeast. EMBO J. 1992, 11, 673–682. [Google Scholar] [CrossRef]
- Bugler, B.; Bourbon, H.; Lapeyre, B.; Wallace, M.O.; Chang, J.H.; Amalric, F.; Olson, M.O. RNA binding fragments from nucleolin contain the ribonucleoprotein consensus sequence. J. Biol. Chem. 1987, 262, 10922–10925. [Google Scholar]
- Snaar, S.; Wiesmeijer, K.; Jochemsen, A.G.; Tanke, H.J.; Dirks, R.W. Mutational analysis of fibrillarin and its mobility in living human cells. J. Cell Biol. 2000, 151, 653–662. [Google Scholar] [CrossRef]
- Fiume, R.; Keune, W.J.; Faenza, I.; Bultsma, Y.; Ramazzotti, G.; Jones, D.R.; Martelli, A.M.; Somner, L.; Follo, M.Y.; Divecha, N.; et al. Nuclear phosphoinositides: Location, regulation and function. Subcell. Biochem. 2012, 59, 335–361. [Google Scholar]
- Sagaram, U.S.; El-Mounadi, K.; Buchko, G.W.; Berg, H.R.; Kaur, J.; Pandurangi, R.S.; Smith, T.J.; Shah, D.M. Structural and functional studies of a phosphatidic acid-binding antifungal plant defensin MtDef4: Identification of an RGFRRR motif governing fungal cell entry. PLoS ONE 2013, 8, e82485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wawrzyniak, A.M.; Kashyap, R.; Zimmermann, P. Phosphoinositides and PDZ domain scaffolds. Adv. Exp. Med. Biol. 2013, 991, 41–57. [Google Scholar] [PubMed]
- Warner, J.R. The economics of ribosome biosynthesis in yeast. Trends Biochem. Sci. 1999, 24, 437–440. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guillen-Chable, F.; Rodríguez Corona, U.; Pereira-Santana, A.; Bayona, A.; Rodríguez-Zapata, L.C.; Aquino, C.; Šebestová, L.; Vitale, N.; Hozak, P.; Castano, E. Fibrillarin Ribonuclease Activity is Dependent on the GAR Domain and Modulated by Phospholipids. Cells 2020, 9, 1143. https://doi.org/10.3390/cells9051143
Guillen-Chable F, Rodríguez Corona U, Pereira-Santana A, Bayona A, Rodríguez-Zapata LC, Aquino C, Šebestová L, Vitale N, Hozak P, Castano E. Fibrillarin Ribonuclease Activity is Dependent on the GAR Domain and Modulated by Phospholipids. Cells. 2020; 9(5):1143. https://doi.org/10.3390/cells9051143
Chicago/Turabian StyleGuillen-Chable, Francisco, Ulises Rodríguez Corona, Alejandro Pereira-Santana, Andrea Bayona, Luis Carlos Rodríguez-Zapata, Cecilia Aquino, Lenka Šebestová, Nicolas Vitale, Pavel Hozak, and Enrique Castano. 2020. "Fibrillarin Ribonuclease Activity is Dependent on the GAR Domain and Modulated by Phospholipids" Cells 9, no. 5: 1143. https://doi.org/10.3390/cells9051143